Fucoidan Induces Apoptosis of HT-29 Cells via the Activation of DR4 and Mitochondrial Pathway


Abstract
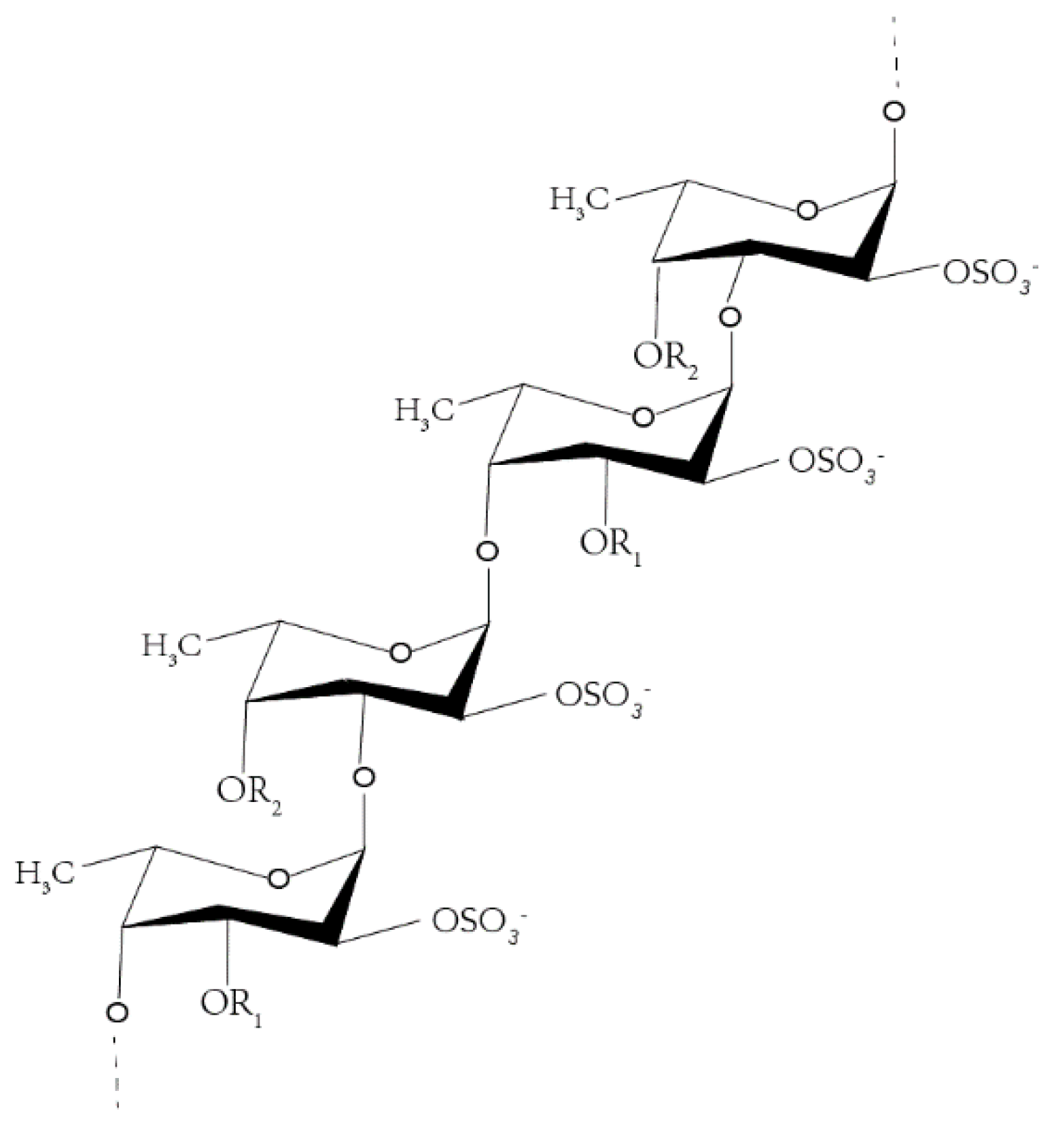
:1. Introduction
2. Results
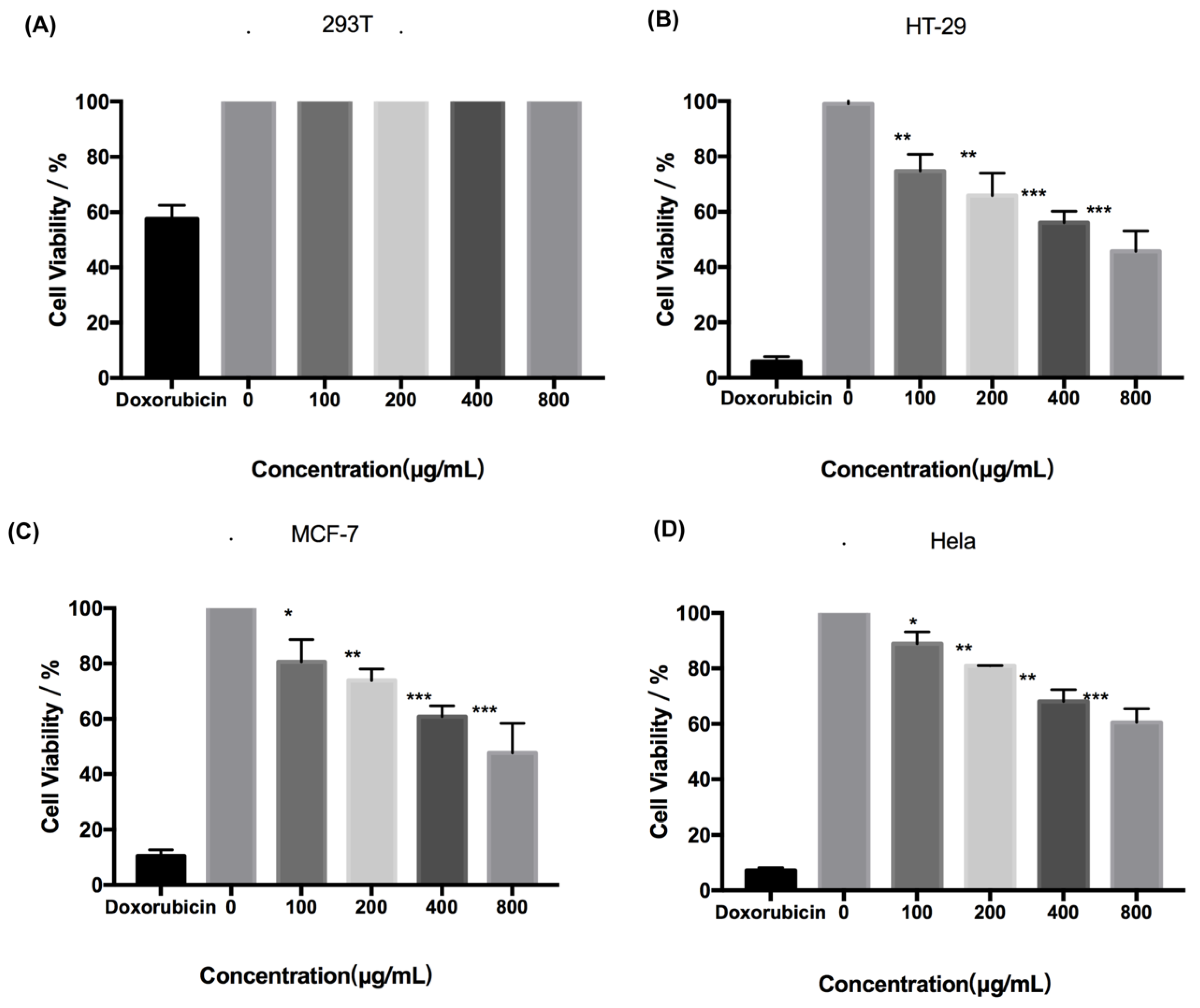
2.1. Cytotoxicity of Fucoidan
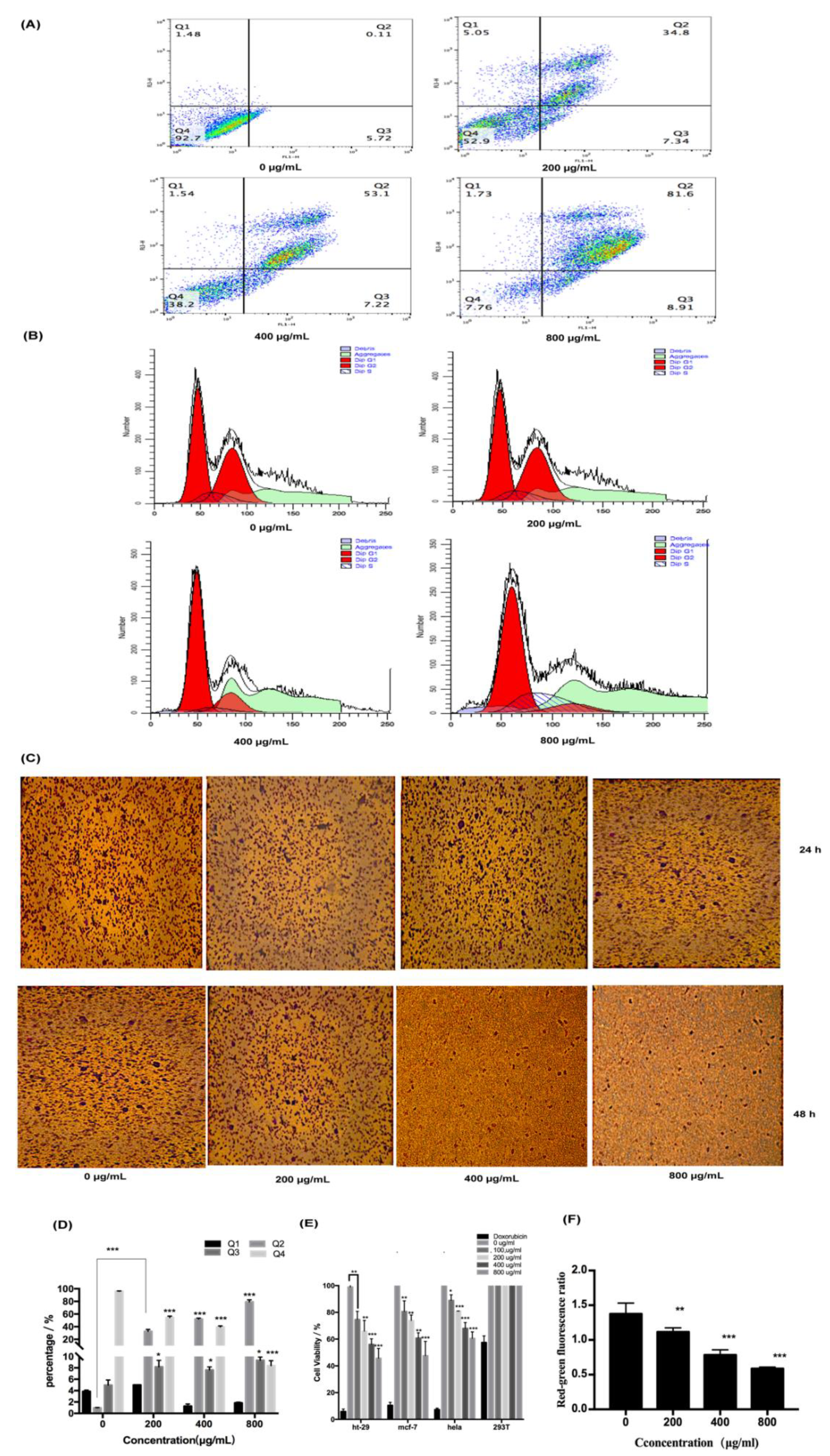
2.2. Pharmacological Activity of Fucoidan on HT-29 Cells
2.3. Analysis of Fucoidan-Induced Apoptosis of HT-29 Cells
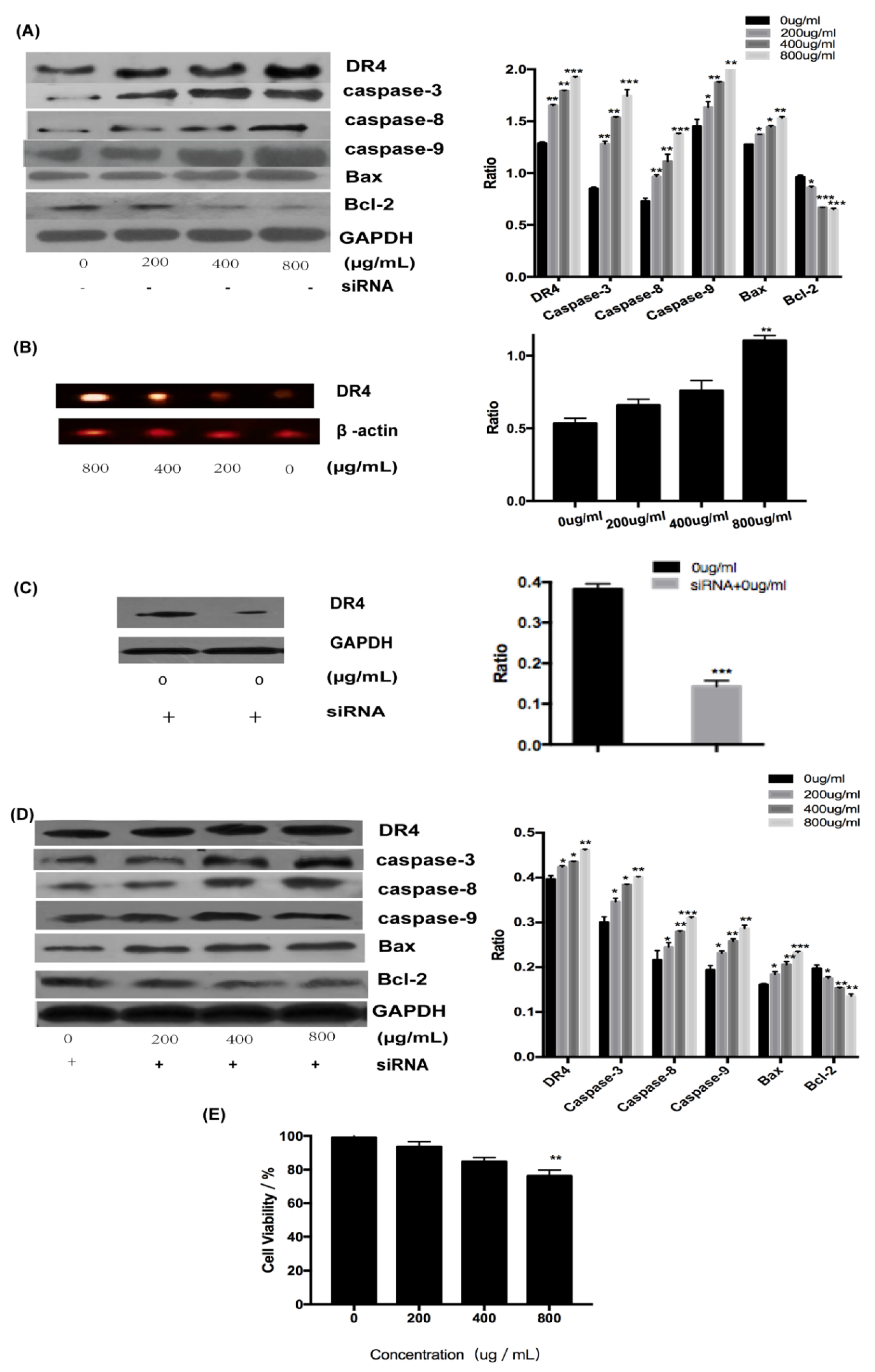
2.3.1. Fucoidan Can Induce Apoptosis Through the Extrinsic Pathway
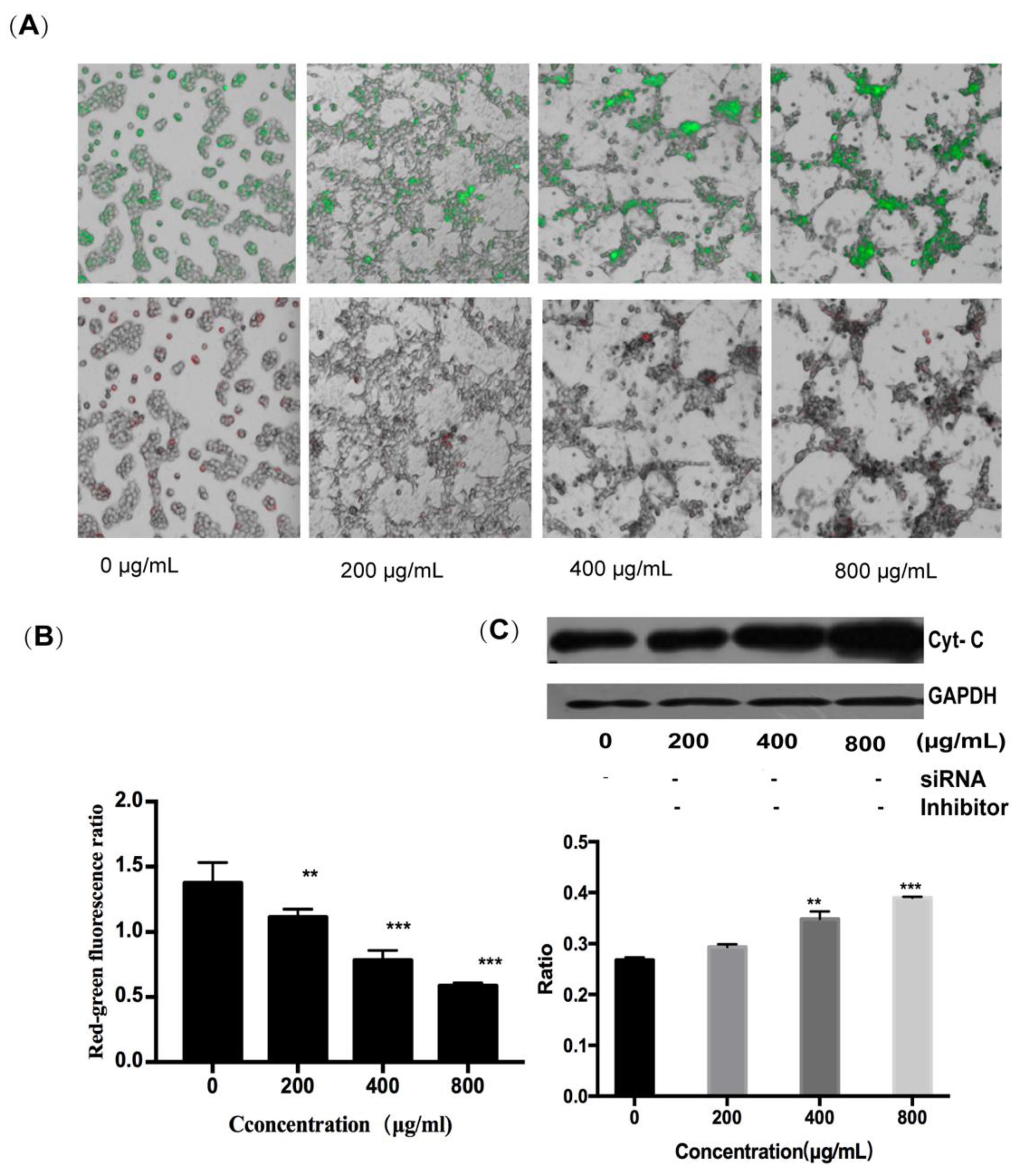
2.3.2. Fucoidan Can Induce Apoptosis Through the Intrinsic Pathway
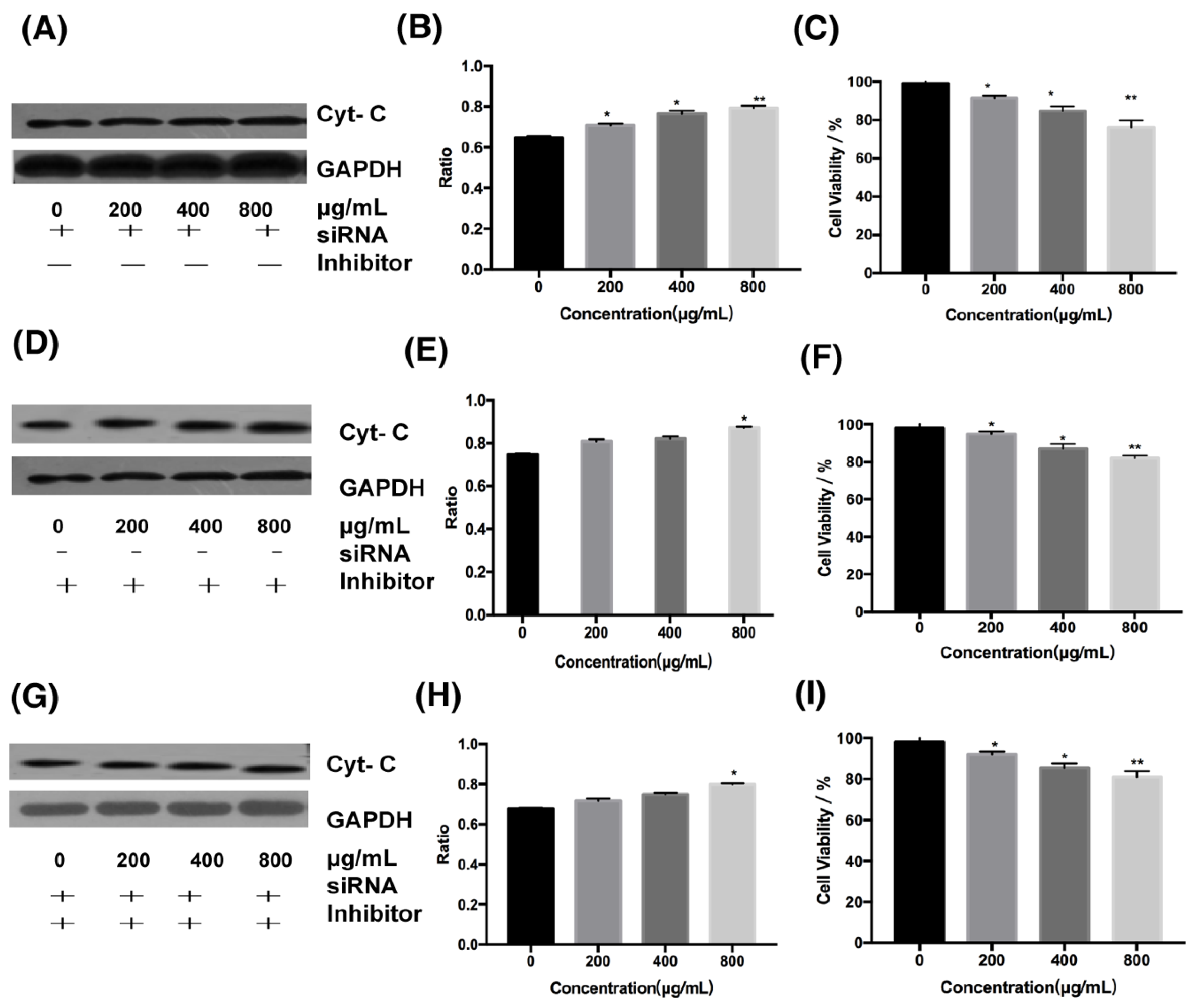
2.3.3. Relationship Between the Extrinsic and Intrinsic Pathways
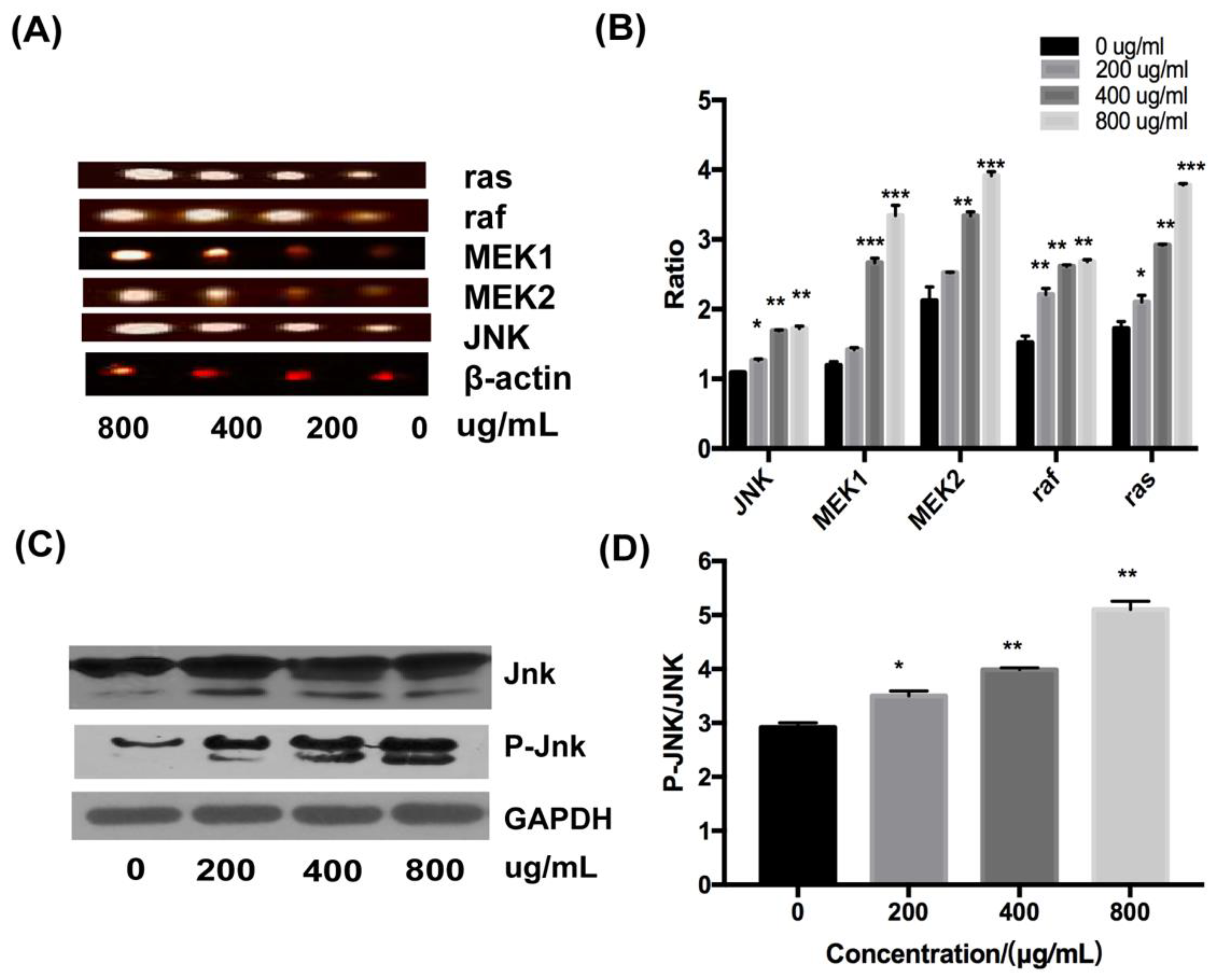
2.4. Effect of Fucoidan on the JNK Signaling Pathway in HT-29 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. MTT Assay for Cell Viability
4.4. Cell Cycle Analysis
4.5. Cell Migration Assay
4.6. Quantification of Apoptosis with the Annexin V/Propidium Iodide Assay
4.7. Total RNA Extraction and Reverse Transcription-Polymerase Chain Reaction
4.8. Silencing the Expression of DR4 by siRNA
4.9. Detection of Mitochondrial Membrane Potential by JC-1 Staining
4.10. Inhibition of Cytochrome C Expression
4.11. Western Blot Analysis
4.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bai, X.; Zhang, E.; Hu, B.; Liang, H.; Song, S.; Ji, A. Study on Absorption Mechanism and Tissue Distribution of Fucoidan. Molecules 2020, 25, 1087. [Google Scholar] [CrossRef] [Green Version]
- Gabbia, D.; Saponaro, M.; Sarcognato, S.; Guido, M.; Ferri, N.; Carrara, M.; de Martin, S. Fucus vesiculosus and Ascophyllum nodosum Ameliorate Liver Function by Reducing Diet-Induced Steatosis in Rats. Mar. Drugs 2020, 18, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.; Lee, J.Y.; Yang, C.; Song, G.; Lim, W. Fucoidan Derived from Fucus vesiculosus Inhibits the Development of Human Ovarian Cancer via the Disturbance of Calcium Homeostasis, Endoplasmic Reticulum Stress, and Angiogenesis. Mar. Drugs 2020, 18, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and Tissue Distribution of Fucoidan from Fucus vesiculosus after Oral Administration to Rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic Effects of Fucoidan: A Review on Recent Studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [Green Version]
- Usoltseva, R.V.; Shevchenko, N.M.; Malyarenko, O.S.; Ishina, I.A.; Ivannikova, S.I.; Ermakova, S.P. Structure and anticancer activity of native and modified polysaccharides from brown alga Dictyota dichotoma. Carbohydr. Polym. 2018, 180, 21–28. [Google Scholar] [CrossRef]
- Dobrincic, A.; Balbino, S.; Zoric, Z.; Pedisic, S.; Kovacevic, D.B.; Garofulic, I.E.; Dragovic-Uzelac, V. Advanced Technologies for the Extraction of Marine Brown Algal Polysaccharides. Mar. Drugs 2020, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vaquero, M.; Rajauria, G.; Tiwari, B.; Sweeney, T.; O’Doherty, J. Extraction and Yield Optimisation of Fucose, Glucans and Associated Antioxidant Activities from Laminaria digitata by Applying Response Surface Methodology to High Intensity Ultrasound-Assisted Extraction. Mar. Drugs 2018, 16, 257. [Google Scholar] [CrossRef] [Green Version]
- Obluchinskaya, E.D.; Makarova, M.N.; Pozharitskaya, O.N.; Shikov, A.N. Effects of Ultrasound Treatment on the Chemical Composition and Anticoagulant Properties of Dry Fucus Extract. Pharm. Chem. J. 2015, 49, 183–186. [Google Scholar] [CrossRef]
- Blaszczak, W.; Lach, M.S.; Barczak, W.; Suchorska, W.M. Fucoidan Exerts Anticancer Effects against Head and Neck Squamous Cell Carcinoma In Vitro. Molecules 2018, 23, 3302. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Abeta25-35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Mar. Drugs 2017, 15, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Q.; Li, G.; Li, C.; Li, Q.; Li, J.; Liu, C.; Pan, L.; Li, S.; Cai, C.; Hao, J.; et al. Two different fucosylated chondroitin sulfates: Structural elucidation, stimulating hematopoiesis and immune-enhancing effects. Carbohydr. Polym. 2020, 230, 115698. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.Y.; Chen, L.; Li, Q.J.; Shen, Y.Z.; Jin, L.; Zhang, X.; Chen, P.C.; Wu, M.J.; Choi, J.I.; et al. Different extraction methods bring about distinct physicochemical properties and antioxidant activities of Sargassum fusiforme fucoidans. Int. J. Biol. Macromol. 2019. [Google Scholar] [CrossRef]
- Amin, M.L.; Mawad, D.; Dokos, S.; Koshy, P.; Martens, P.J.; Sorrell, C.C. Immunomodulatory properties of photopolymerizable fucoidan and carrageenans. Carbohydr. Polym. 2020, 230, 115691. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, Y.; Cao, M.J.; Liu, G.M.; Chen, Q.; Sun, L.; Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohydr. Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef]
- Ji, X.; Peng, Q.; Wang, M. Anti-colon-cancer effects of polysaccharides: A mini-review of the mechanisms. Int. J. Biol. Macromol. 2018, 114, 1127–1133. [Google Scholar] [CrossRef]
- Bovet, L.; Samer, C.; Daali, Y. Preclinical Evaluation of Safety of Fucoidan Extracts from Undaria pinnatifida and Fucus vesiculosus for Use in Cancer Treatment. Integr. Cancer 2019, 18, 1534735419876325. [Google Scholar] [CrossRef] [Green Version]
- Paternot, S.; Bockstaele, L.; Bisteau, X.; Kooken, H.; Coulonval, K.; Roger, P.P. Rb inactivation in cell cycle and cancer: The puzzle of highly regulated activating phosphorylation of CDK4 versus constitutively active CDK-activating kinase. Cell Cycle 2010, 9, 689–699. [Google Scholar] [CrossRef]
- Ishikawa, C.; Senba, M.; Mori, N. Mitotic kinase PBK/TOPK as a therapeutic target for adult Tcell leukemia/lymphoma. Int. J. Oncol. 2018, 53, 801–814. [Google Scholar]
- Zayed, A.; Ulber, R. Fucoidan production: Approval key challenges and opportunities. Carbohydr. Polym. 2019, 211, 289–297. [Google Scholar] [CrossRef]
- Chen, M.C.; Hsu, W.L.; Hwang, P.A.; Chou, T.C. Low Molecular Weight Fucoidan Inhibits Tumor Angiogenesis through Downregulation of HIF-1/VEGF Signaling under Hypoxia. Mar. Drugs 2015, 13, 4436–4451. [Google Scholar] [CrossRef]
- Wang, F.; Ye, X.; Zhai, D.; Dai, W.; Wu, Y.; Chen, J.; Chen, W. Curcumin-loaded nanostructured lipid carrier induced apoptosis in human HepG2 cells through activation of the DR5/caspase-mediated extrinsic apoptosis pathway. Acta Pharm. 2020, 70, 227–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Ji, X.; Xue, C.; Liang, H.; Ge, Y.; He, X.; Zhang, L.; Bian, K.; Zhang, L. Caspase-dependent and caspase-independent induction of apoptosis in breast cancer by fucoidan via the PI3K/AKT/GSK3beta pathway in vivo and in vitro. Biomed. Pharm. 2017, 94, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, M.; Wang, W.; Pang, L.; Wang, Z.; Yuan, C.; Liu, K. Overexpression of apoptosis-inducing factor mitochondrion-associated 1 (AIFM1) induces apoptosis by promoting the transcription of caspase3 and DRAM in hepatoma cells. Biochem. Biophys. Res. Commun. 2018, 498, 453–457. [Google Scholar] [CrossRef]
- Wu, Q.; Deng, J.; Fan, D.; Duan, Z.; Zhu, C.; Fu, R.; Wang, S. Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem. Pharm. 2018, 148, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Amarante-Mendes, G.P.; Griffith, T.S. Therapeutic applications of TRAIL receptor agonists in cancer and beyond. Pharmacol. Ther. 2015, 155, 117–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittner, S.; Knoll, G.; Ehrenschwender, M. Death receptor 3 mediates necroptotic cell death. Cell Mol. Life Sci. 2017, 74, 543–554. [Google Scholar] [CrossRef]
- Flusberg, D.A.; Sorger, P.K. Surviving apoptosis: Life-death signaling in single cells. Trends Cell Biol. 2015, 25, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.O.; Song, M.G.; Kim, Y.N.; Park, J.I.; Kwak, J.Y. The mechanism of fucoidan-induced apoptosis in leukemic cells: Involvement of ERK1/2, JNK, glutathione, and nitric oxide. Mol. Carcinog. 2010, 49, 771–782. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. Sulfated polysaccharides from brown seaweeds Saccharina japonica and Undaria pinnatifida: Isolation, structural characteristics, and antitumor activity. Carbohydr. Res. 2011, 346, 2769–2776. [Google Scholar] [CrossRef]
- Lim, S.J.; Aida, W.M.W.; Maskat, M.Y.; Latip, J.; Badri, K.H.; Hassan, O.; Yamin, B.M. Characterisation of fucoidan extracted from Malaysian Sargassum binderi. Food Chem. 2016, 209, 267–273. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences | Product Size | |
|---|---|---|---|
| Ras | Forward | 5’-CGACACAGCAGGTCAAGAGG-3’ | 20 |
| Reverse | 5’-GGCATCATCAACACCCTGTCT-3’ | 21 | |
| Raf | Forward | 5’-CAGCGAATCAGCCTCACCTTCAG-3’ | 23 |
| Reverse | 5’-CGCAGAACAGCCACCTCATTCC-3’ | 22 | |
| β-actin | Forward | 5’-CGTGGACATCCGCAAAGAC-3’ | 19 |
| Reverse | 5’-GCATTTGCGGTGGACGAT-3’ | 18 | |
| DR4 | Forward | 5’-CTGATCACCCAACAAGACCTAG-3’ | 22 |
| Reverse | 5’-GATGCAATCTCTACCGCTTCT-3’ | 22 | |
| JNK | Forward | 5’-GGAATGGCCTGCCTTACGATGAC-3’ | 23 |
| Reverse | 5’-GGCTCTGTTGCTGCCACTGC-3’ | 20 | |
| MEK1 | Forward | 5’-CAGCTCTGCGGAGACCAACTTG-3’ | 22 |
| Reverse | 5’-CTGATCTCGCCATCGCTGTAGAAC-3’ | 24 | |
| MEK2 | Forward | 5’-ACTTGACGAGCAGCAGAAGAAGC-3’ | 23 |
| Reverse | 5’-GAGCCGCCGTCCATGTGTTC-3’ | 20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Wang, Y.; Hu, B.; Cao, Q.; Xing, M.; Song, S.; Ji, A. Fucoidan Induces Apoptosis of HT-29 Cells via the Activation of DR4 and Mitochondrial Pathway. Mar. Drugs 2020, 18, 220. https://doi.org/10.3390/md18040220
Bai X, Wang Y, Hu B, Cao Q, Xing M, Song S, Ji A. Fucoidan Induces Apoptosis of HT-29 Cells via the Activation of DR4 and Mitochondrial Pathway. Marine Drugs. 2020; 18(4):220. https://doi.org/10.3390/md18040220
Chicago/Turabian StyleBai, Xu, Yu Wang, Bo Hu, Qi Cao, Maochen Xing, Shuliang Song, and Aiguo Ji. 2020. "Fucoidan Induces Apoptosis of HT-29 Cells via the Activation of DR4 and Mitochondrial Pathway" Marine Drugs 18, no. 4: 220. https://doi.org/10.3390/md18040220
APA StyleBai, X., Wang, Y., Hu, B., Cao, Q., Xing, M., Song, S., & Ji, A. (2020). Fucoidan Induces Apoptosis of HT-29 Cells via the Activation of DR4 and Mitochondrial Pathway. Marine Drugs, 18(4), 220. https://doi.org/10.3390/md18040220