Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp

 , and
, and
Abstract
:1. Introduction
2. Results
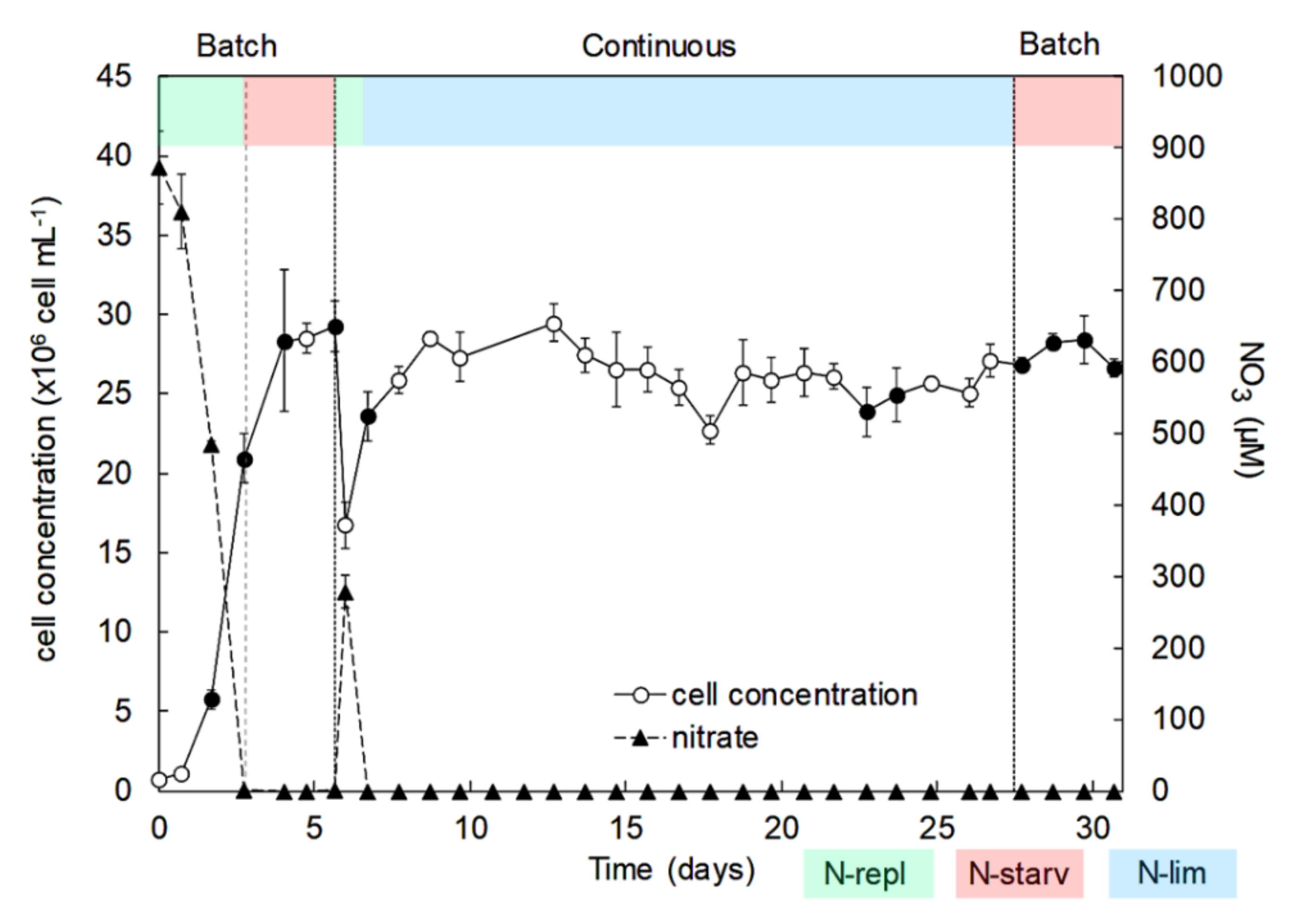
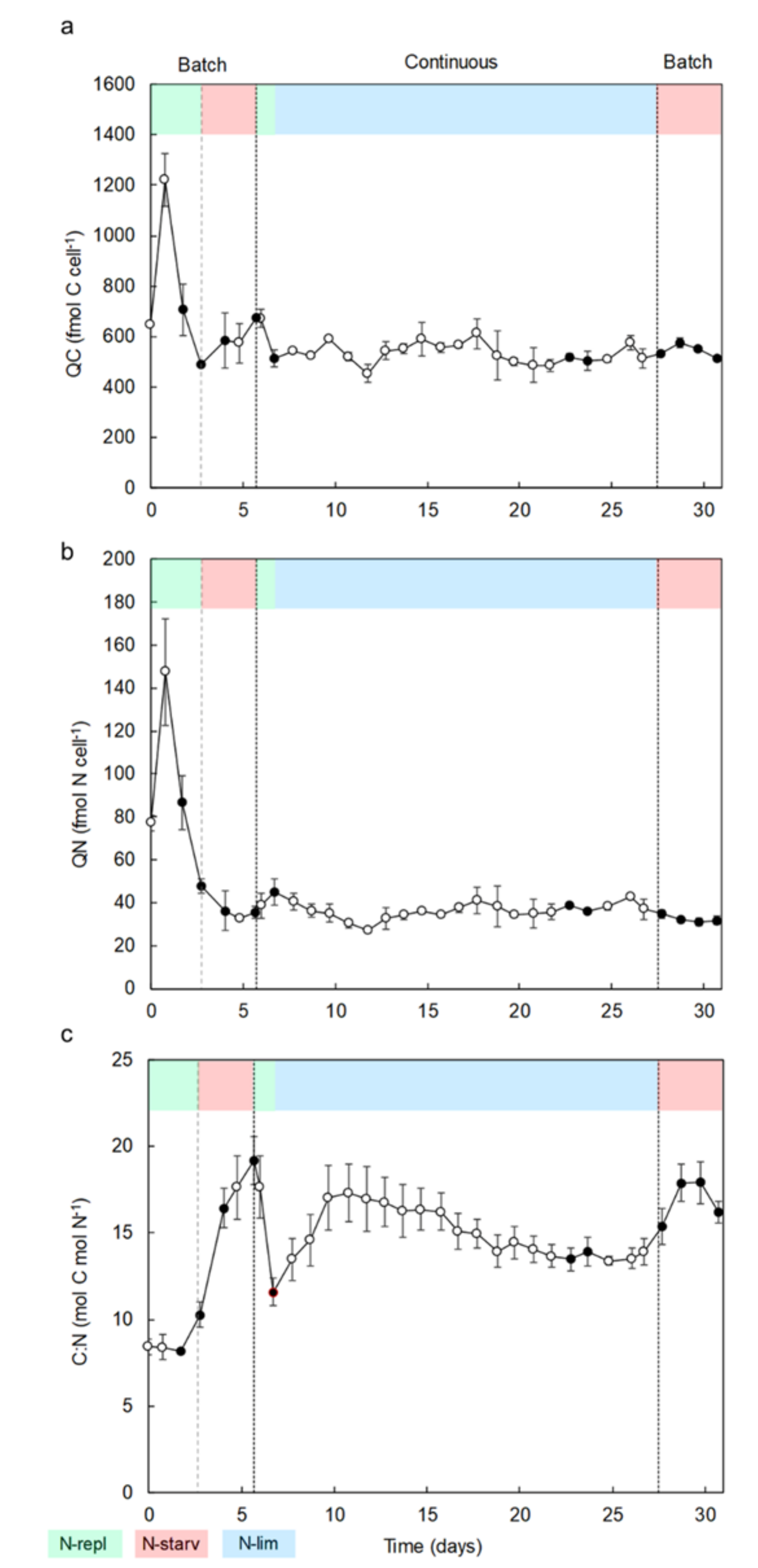
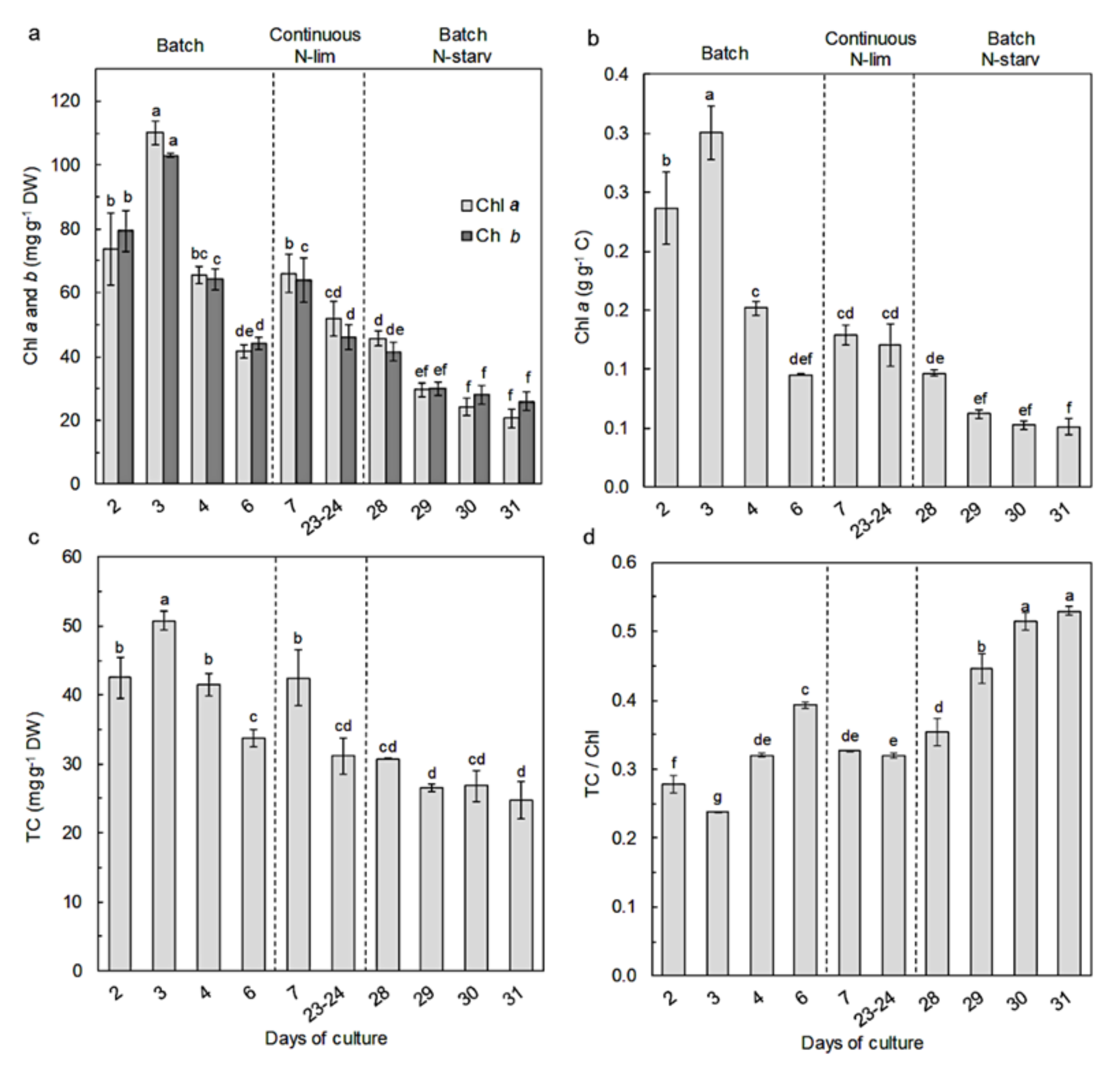
2.1. Identification, Growth, and Cellular Elemental Composition
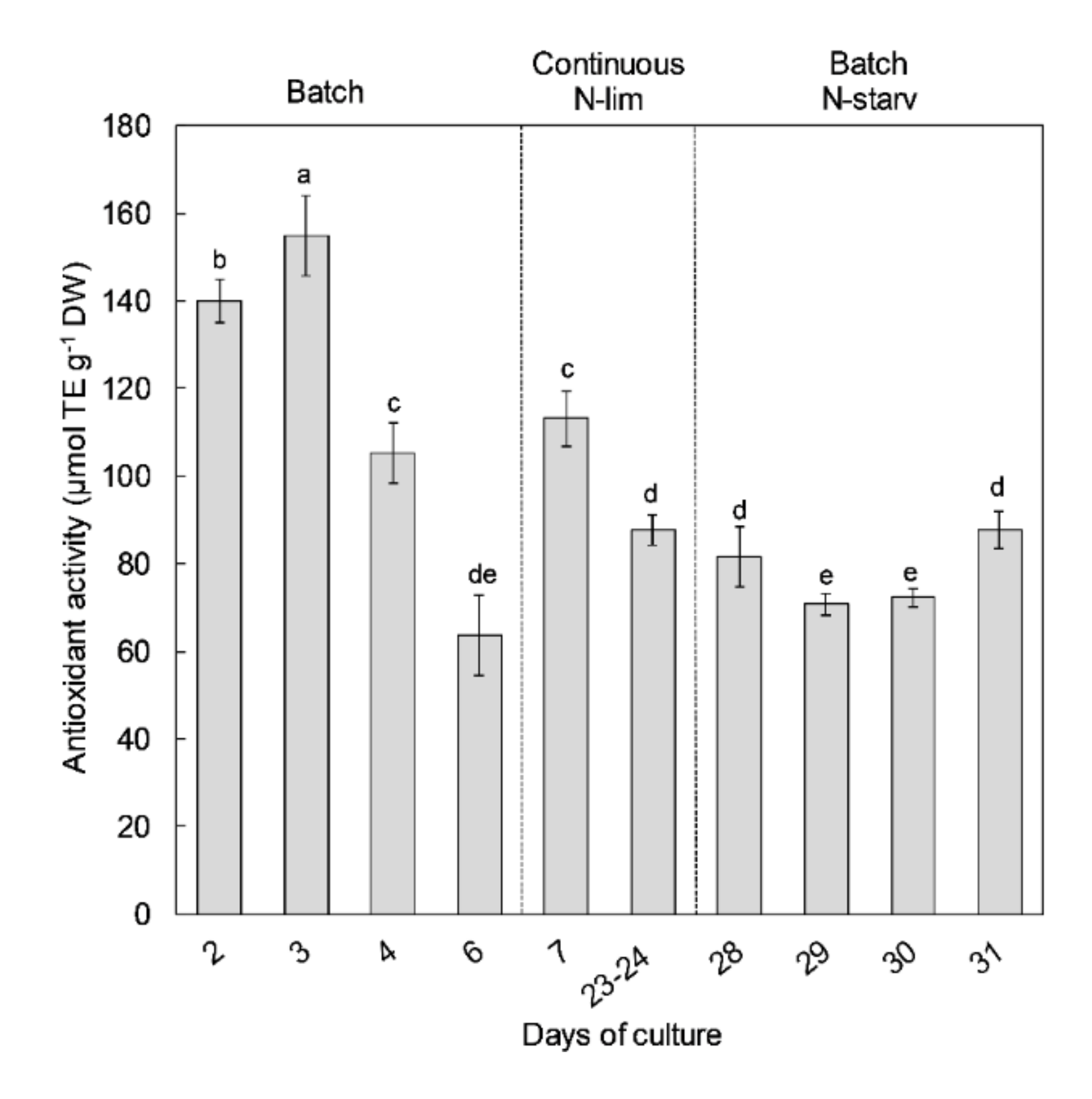
2.2. Antioxidant Activity
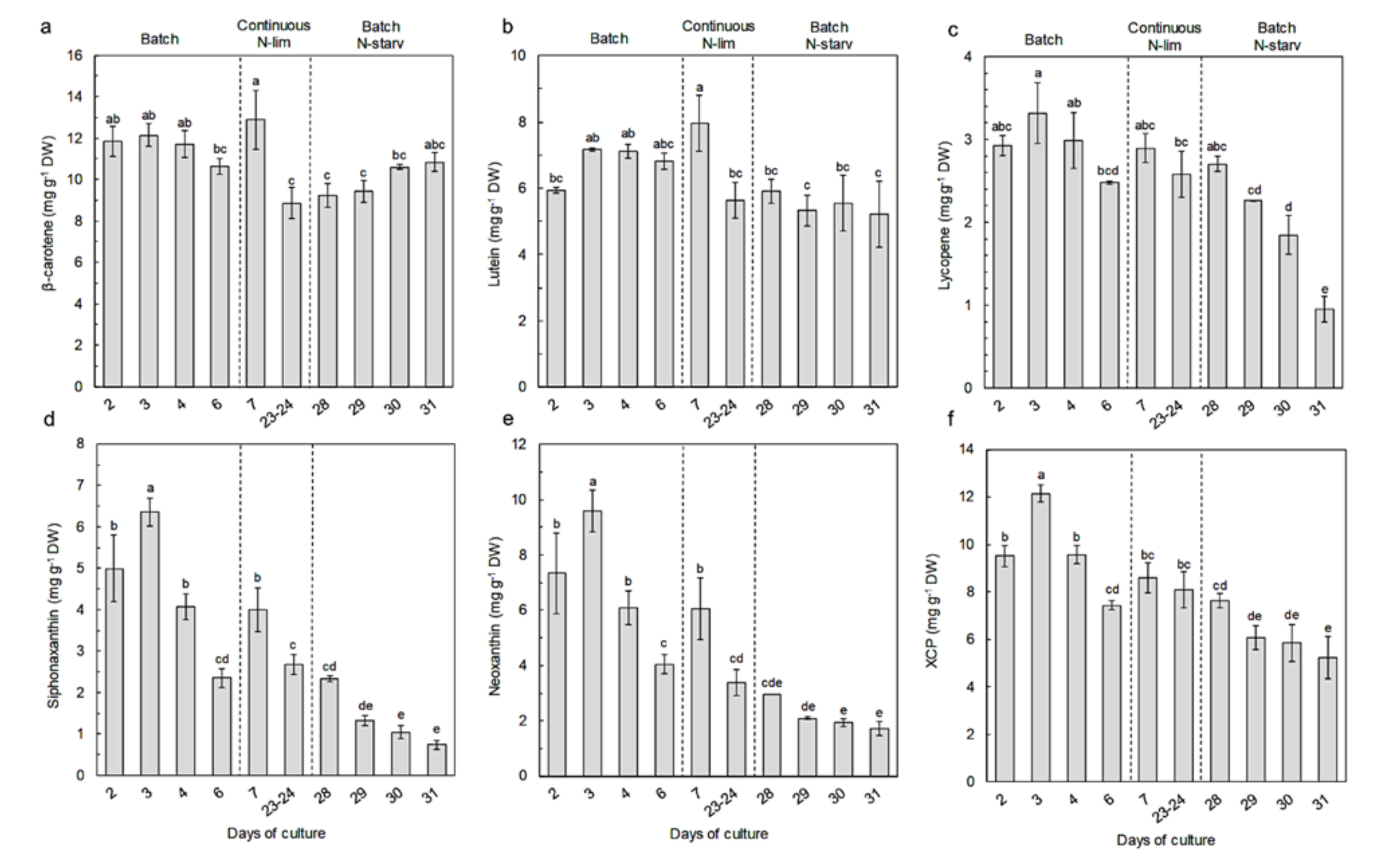
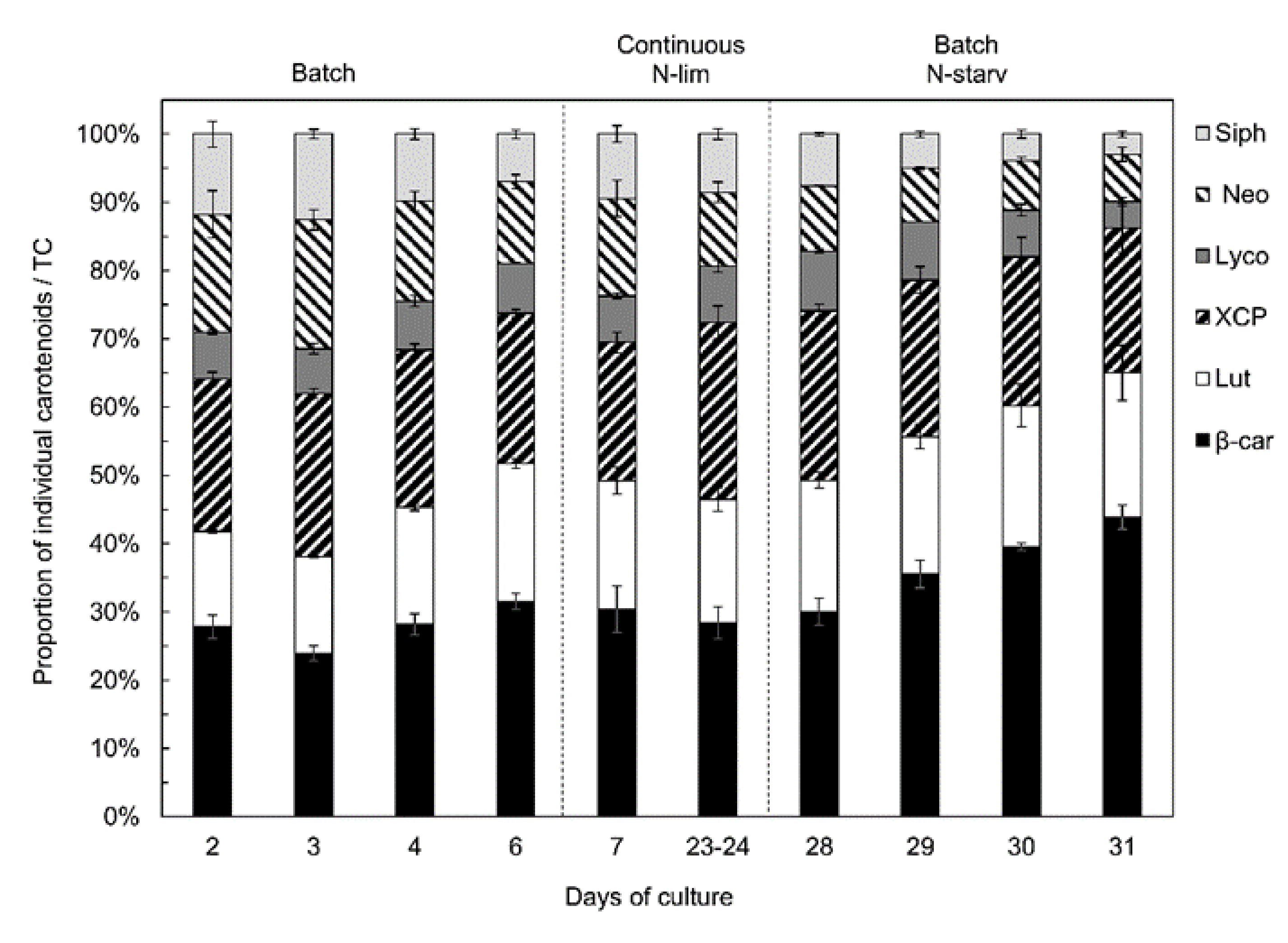
2.3. Pigment Content and Composition
2.4. Correlation between Antioxidant Activity and Carotenoid Content
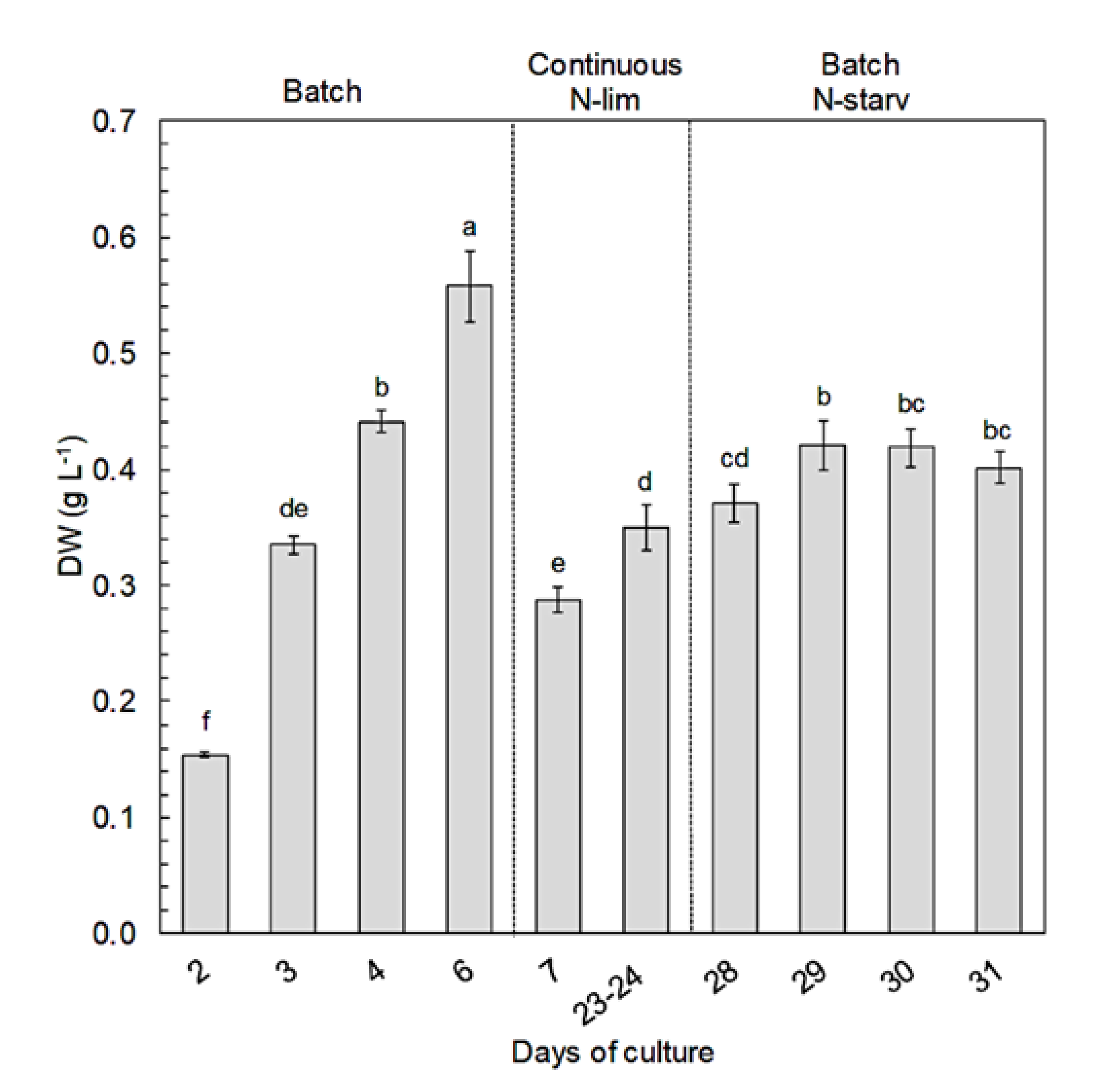
2.5. Biomass and Carotenoid Productivities
3. Discussion
4. Materials and Methods
4.1. Strain
4.2. Nephroselmis sp. Molecular Identification
4.3. Culture Conditions and Experimental Protocol
- (1)
- A first batch culture period (day 0 to day 6) was applied to study the effects of N-replete to N-starvation conditions. At the end of this period, and owing to sample collections during this first batch culture, PBRs volumes were restored to 9.5 L (day 6) by adding filtered sea water (0.2 µm) enriched with 1 mL L−1 of Walne’s medium at 1.18 mM-N to ensure that the temperature and pH probes remained submerged;
- (2)
- The PBRs were then switched to a continuous mode of culture in chemostat to study the effects of N-limitation (day 7 to day 27). During this period, a dilution rate of 0.3 day−1 was applied. The culture medium was composed of filtered seawater (0.2 µm) enriched in 1 mL L−1 of Walne’s medium at 1.18 mM-N [99]. The light intensity was increased to 1100 µmol m−2 s−1 on day 8 to ensure that there was no light limitation owing to the very high cellular concentration (>25 × 106 cell mL−1). Continuous culture was maintained until analyses at steady state (i.e., at least 3 days with less than 10% variation of cellular concentration and absorbance);
- (3)
- Finally, the PBRs were switched back to a second batch culture period (day 28 to day 31) in N-starvation conditions until beginning of senescence.
4.4. Cell Growth Measurements
4.5. Particulate Organic Carbon and Nitrogen and Residual Nitrate and Phosphate
4.6. Measurement of Antioxidant Activity and Pigments Analysis
4.6.1. Sampling
4.6.2. Antioxidant Activity
4.6.3. Pigments Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morais, D.; Greque, M.; Vaz, B.D.S.; Morais, D.; Greque, E.; Costa, J.A.V. Biologically Active Metabolites Synthesized by Microalgae. Available online: https://www.hindawi.com/journals/bmri/2015/835761/abs/ (accessed on 5 February 2018).
- Sansone, C.; Brunet, C. Promises and Challenges of Microalgal Antioxidant Production. Antioxidants 2019, 8, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aklakur, M. Natural antioxidants from sea: A potential industrial perspective in aquafeed formulation. Rev. Aquacult. 2016, 10, 385–399. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of high added-value compounds—A brief review of recent work. Biotechnol. Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.-P.; Morant-Manceau, A.; Schoefs, B. The potential of microalgae for the production of bioactive molecules of pharmaceutical interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef] [PubMed]
- Safafar, H.; van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, Phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [Green Version]
- Abalde, J.; Fabregas, J.; Herrero, C. β-Carotene, vitamin C and vitamin E content of the marine microalga Dunaliella tertiolecta cultured with different nitrogen sources. Bioresour. Technol. 1991, 38, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; Brabanter, J.D.; Cooman, L.D. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Durmaz, Y. Vitamin E (α-tocopherol) production by the marine microalgae Nannochloropsis oculata (Eustigmatophyceae) in nitrogen limitation. Aquaculture 2007, 272, 717–722. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef]
- Sandmann, G. Antioxidant Protection from UV- and light-stress related to carotenoid structures. Antioxidants 2019, 8, 219. [Google Scholar] [CrossRef] [Green Version]
- Young, A.J.; Lowe, G.M. Antioxidant and Prooxidant Properties of Carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Adams, W.W. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends Plant. Sci. 1996, 1, 21–26. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The carotenoids as anti-oxidants—A review. J. Photochem. Photobiol. B Biol. 1997, 41, 189–200. [Google Scholar] [CrossRef]
- Patias, L.D.; Fernandes, A.S.; Petry, F.C.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Carotenoid profile of three microalgae/cyanobacteria species with peroxyl radical scavenger capacity. Food Res. Int. 2017. [Google Scholar] [CrossRef]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic pigment production with microalgae: Biological constraints and opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef]
- Borowitzka, M.A. The ‘stress’ concept in microalgal biology—Homeostasis, acclimation and adaptation. J. Appl. Phycol. 2018, 30, 2815–2825. [Google Scholar] [CrossRef]
- Salguero, A.; de la Morena, B.; Vigara, J.; Vega, J.M.; Vilchez, C.; León, R. Carotenoids as protective response against oxidative damage in Dunaliella bardawil. Biomol. Eng. 2003, 20, 249–253. [Google Scholar] [CrossRef]
- Shaish, A.; Avron, M.; Pick, U.; Ben-Amotz, A. Are active oxygen species involved in induction of β-carotene in Dunaliella bardawil? Planta 1993, 190, 363–368. [Google Scholar] [CrossRef]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, V.S.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic stresses as tools for metabolites in microalgae. Bioresour. Technol. 2017. [Google Scholar] [CrossRef]
- Solovchenko, A.E. Physiology and adaptive significance of secondary carotenogenesis in green microalgae. Russ. J. Plant. Physiol. 2013, 60, 1–13. [Google Scholar] [CrossRef]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Nitrogen starvation-induced cellular crosstalk of ROS-scavenging antioxidants and phytohormone enhanced the biofuel potential of green microalga Acutodesmus dimorphus. Biotechnol. Biofuels 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-M.; Chen, H.; He, C.-L.; Wang, Q. Nitrogen starvation induced oxidative stress in an oil-producing green alga Chlorella sorokiniana C3. PLoS ONE 2013, 8, e69225. [Google Scholar] [CrossRef] [PubMed]
- Çakmak, Z.E.; Ölmez, T.T.; Çakmak, T.; Menemen, Y.; Tekinay, T. Antioxidant response of Chlamydomonas reinhardtii grown under different element regimes. Phycol. Res. 2015, 63, 202–211. [Google Scholar] [CrossRef]
- Hamid, S.; Sibi, G. Antioxidant System Response in Green Microalga Chlorococcopsis minuta Against Nutrient Stress in Growth Media. Asian J. Biol. Sci. 2018, 11, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Cui, Y.; Wan, M.; Wang, W.; Li, Y. Lipid accumulation and biosynthesis genes response of the oleaginous Chlorella pyrenoidosa under three nutrition stressors. Biotechnol. Biofuels 2014, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative Stress Is a Mediator for Increased Lipid Accumulation in a Newly Isolated Dunaliella salina Strain. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Goiris, K.; Van Colen, W.; Wilches, I.; León-Tamariz, F.; De Cooman, L.; Muylaert, K. Impact of nutrient stress on antioxidant production in three species of microalgae. Algal Res. 2015, 7, 51–57. [Google Scholar] [CrossRef]
- Lamers, P.P.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in nitrogen-starved Dunaliella salina, a unicellular green microalga. J. Biotechnol. 2012, 162, 21–27. [Google Scholar] [CrossRef]
- Ruiz-Domínguez, M.C.; Vaquero, I.; Obregón, V.; de la Morena, B.; Vílchez, C.; Vega, J.M. Lipid accumulation and antioxidant activity in the eukaryotic acidophilic microalga Coccomyxa sp. (strain onubensis) under nutrient starvation. J. Appl. Phycol. 2015, 27, 1099–1108. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M.N. Effects of light and nitrogen starvation on the content and composition of carotenoids of the green microalga Parietochloris incisa. Russ. J. Plant. Physiol. 2008, 55, 455–462. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, H.; Yuan, C.; Li, T.; Li, A. Growth, biochemical composition, and photosynthetic performance of Scenedesmus acuminatus during nitrogen starvation and resupply. J. Appl. Phycol. 2019, 31, 2797–2809. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. On the Factors Which Determine Massive β-Carotene Accumulation in the Halotolerant Alga Dunaliella bardawil. Plant. Physiol. 1983, 72, 593–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aremu, A.O.; Neményi, M.; Stirk, W.A.; Ördög, V.; van Staden, J. Manipulation of nitrogen levels and mode of cultivation are viable methods to improve the lipid, fatty acids, phytochemical content, and bioactivities in Chlorella minutissima. J. Phycol. 2015, 51, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Aremu, A.O.; Masondo, N.A.; Molnár, Z.; Stirk, W.A.; Ördög, V.; van Staden, J. Changes in phytochemical content and pharmacological activities of three Chlorella strains grown in different nitrogen conditions. J. Appl. Phycol. 2016, 28, 149–159. [Google Scholar] [CrossRef]
- Coulombier, N.; Nicolau, E.; Le Déan, L.; Antheaume, C.; Jauffrais, T.; Lebouvier, N. Impact of Light Intensity on Antioxidant Activity of Tropical Microalgae. Mar. Drugs 2020, 18, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshii, Y.; Takaichi, S.; Maoka, T.; Suda, S.; Sekiguchi, H.; Nakayama, T.; Inouye, I. Variation of siphonaxanthin series among the genus Nephroselmis (prasinophyceae, Chlorophyta), including a novel primary methoxy carotenoid. J. Phycol. 2005, 41, 827–834. [Google Scholar] [CrossRef]
- Dambeck, M.; Sandmann, G. Antioxidative activities of algal keto carotenoids acting as antioxidative protectants in the chloroplast. Photochem. Photobiol. 2014, 90, 814–819. [Google Scholar] [CrossRef]
- Ganesan, P.; Matsubara, K.; Ohkubo, T.; Tanaka, Y.; Noda, K.; Sugawara, T.; Hirata, T. Anti-angiogenic effect of siphonaxanthin from green alga, Codium fragile. Phytomedicine 2010, 17, 1140–1144. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-S.; Noda, K.; Fujita, E.; Manabe, Y.; Hirata, T.; Sugawara, T. The green algal carotenoid siphonaxanthin inhibits adipogenesis in 3T3-L1 preadipocytes and the accumulation of lipids in white adipose tissue of KK-Ay mice. J. Nutr. 2015. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Li, Z.; Manabe, Y.; Kim, M.; Goto, T.; Kawada, T.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae, inhibits lipogenesis in hepatocytes via the suppression of liver x receptor α activity. Lipids 2018, 41–52. [Google Scholar] [CrossRef]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae Codium cylindricum, protects Ob/Ob mice fed on a high-fat diet against lipotoxicity by ameliorating somatic stresses and restoring anti-oxidative capacity. Nutr. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Manabe, Y.; Takii, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae, suppresses advanced glycation end product-induced inflammatory responses. J. Nat. Med. 2020, 74, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a marine carotenoid from green algae, effectively induces apoptosis in human leukemia (HL-60) cells. Biochim. Biophys. Acta 2011, 1810, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Marin, B.; Melkonian, M. Molecular phylogeny and classification of the Mamiellophyceae class. nov. (Chlorophyta) based on sequence comparisons of the nuclear- and plastid-encoded rRNA operons. Protist 2010, 161, 304–336. [Google Scholar] [CrossRef] [PubMed]
- Young, E.B.; Beardall, J. Photosynthetic Function in Dunaliella Tertiolecta (chlorophyta) during a nitrogen starvation and recovery cycle. J. Phycol. 2003, 39, 897–905. [Google Scholar] [CrossRef]
- De La Rocha, C.; Terbrüggen, A.; Völker, C.; Hohn, S. Response to and recovery from nitrogen and silicon starvation in Thalassiosira weissflogii: Growth rates, nutrient uptake and C, Si and N content per cell. Mar. Ecol. Prog. Ser. 2010, 412, 57–68. [Google Scholar] [CrossRef]
- Dortch, Q.; Clayton, J.R.; Thoresen, S.S.; Ahmed, S.I. Species differences in accumulation of nitrogen pools in phytoplankton. Mar. Biol. 1984, 81, 237–250. [Google Scholar] [CrossRef]
- Hockin, N.L.; Mock, T.; Mulholland, F.; Kopriva, S.; Malin, G. The response of diatom central carbon metabolism to nitrogen starvation is different from that of green algae and higher plants. Plant. Physiol. 2012, 158, 299–312. [Google Scholar] [CrossRef] [Green Version]
- Redfield, A. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 230A, 205–221. [Google Scholar]
- Geider, R.; La Roche, J. Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis. Eur. J. Phycol. 2002, 37, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cullen, J.J.; Yang, X.; MacIntyre, H.L. Nutrient limitation of marine photosynthesis. In Primary Productivity and Biogeochemical Cycles in the Sea; Falkowski, P.G., Woodhead, A.D., Vivirito, K., Eds.; Environmental Science Research; Springer US: Boston, MA, USA, 1992; pp. 69–88. ISBN 978-1-4899-0762-2. [Google Scholar]
- MacIntyre, H.; Cullen, J. Using cultures to investigate the physiological ecology of microalgae. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2005; pp. 287–326. [Google Scholar]
- Jiang, Y.; Yoshida, T.; Quigg, A. Photosynthetic performance, lipid production and biomass composition in response to nitrogen limitation in marine microalgae. Plant. Physiol. Biochem. 2012, 54, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Breuer, G.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour. Technol. 2012, 124, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Jerez, C.G.; Malapascua, J.R.; Sergejevová, M.; Figueroa, F.L.; Masojídek, J. Effect of Nutrient Starvation under High Irradiance on Lipid and Starch Accumulation in Chlorella fusca (Chlorophyta). Mar. Biotechnol 2016, 18, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zheng, Y.; Zhan, J.; He, C.; Wang, Q. Comparative metabolic profiling of the lipid-producing green microalga Chlorella reveals that nitrogen and carbon metabolic pathways contribute to lipid metabolism. Biotechnol. Biofuels 2017, 10, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant. J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Berges, J.A.; Charlebois, D.O.; Mauzerall, D.C.; Falkowski, P.G. Differential effects of nitrogen limitation on photosynthetic efficiency of photosystems I and II in microalgae. Plant. Physiol. 1996, 110, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Liefer, J.D.; Garg, A.; Campbell, D.A.; Irwin, A.J.; Finkel, Z.V. Nitrogen starvation induces distinct photosynthetic responses and recovery dynamics in diatoms and prasinophytes. PLoS ONE 2018, 13, e0195705. [Google Scholar] [CrossRef] [Green Version]
- Coleman, L.W.; Rosen, B.H.; Schwartzbach, S.D. Preferential loss of chloroplast proteins in nitrogen deficient Euglena. Plant. Cell Physiol. 1988, 29, 1007–1014. [Google Scholar] [CrossRef]
- Geider, R.; Macintyre; Graziano, L.; McKay, R.M. Responses of the photosynthetic apparatus of Dunaliella tertiolecta (Chlorophyceae) to nitrogen and phosphorus limitation. Eur. J. Phycol. 1998, 33, 315–332. [Google Scholar] [CrossRef]
- Kim, S.-H.; Liu, K.-H.; Lee, S.-Y.; Hong, S.-J.; Cho, B.-K.; Lee, H.; Lee, C.-G.; Choi, H.-K. Effects of light intensity and nitrogen starvation on glycerolipid, glycerophospholipid, and carotenoid composition in Dunaliella tertiolecta culture. PLoS ONE 2013, 8, e72415. [Google Scholar] [CrossRef] [Green Version]
- Ben-Amotz, A. Effect of irradiance and nutrient deficiency on the chemical composition of Dunaliella bardawil Ben-Amotz and Avron (Volvocales, Chlorophyta). J. Plant. Physiol. 1987, 131, 479–487. [Google Scholar] [CrossRef]
- Yokohama, Y.; Kageyama, A.; Ikawa, T.; Shimura, S. A carotenoid characteristic of chlorophycean seaweeds living in deep coastal waters. Bot. Mar. 1977, 20, 433–436. [Google Scholar] [CrossRef]
- Anderson, J.M. Chlorophyll-protein complexes of a Codium species, including a light-harvesting siphonaxanthin-Chlorophylla ab-protein complex, an evolutionary relic of some Chlorophyta. Biochim. Et Biophys. Acta (Bba) Bioenerg. 1983, 724, 370–380. [Google Scholar] [CrossRef]
- Chen, G.; Niu, X.; Chen, X.; Li, L.; Kuang, T.; Li, S. Characterization of chlorophyll–protein complexes isolated from a Siphonous green alga, Bryopsis corticulans. Photosynth. Res. 2008, 96, 75. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Qin, X.; Sang, M.; Chen, D.; Wang, K.; Lin, R.; Lu, C.; Shen, J.-R.; Kuang, T. Spectral and functional studies on siphonaxanthin-type light-harvesting complex of photosystem II from Bryopsis corticulans. Photosynth. Res. 2013, 117, 267–279. [Google Scholar] [CrossRef]
- Yoshii, Y. Diversity and evolution of photosynthetic antenna systems in green plants. Phycol. Res. 2006, 54, 220–229. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Takeoka, G.R.; Dao, L.; Flessa, S.; Gillespie, D.M.; Jewell, W.T.; Huebner, B.; Bertow, D.; Ebeler, S.E. Processing effects on lycopene content and antioxidant activity of tomatoes. J. Agric. Food Chem. 2001, 49, 3713–3717. [Google Scholar] [CrossRef]
- Miller, N.J.; Sampson, J.; Candeias, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Muzandu, K.; Ishizuka, M.; Sakamoto, K.Q.; Shaban, Z.; El Bohi, K.; Kazusaka, A.; Fujita, S. Effect of lycopene and β-carotene on peroxynitrite-mediated cellular modifications. Toxicol. Appl. Pharmacol. 2006, 215, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Stahl, W. Vitamins E and C, beta-carotene, and other carotenoids as antioxidants. Am. J. Clin. Nutr. 1995, 62, 1315S–1321S. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Paliwal, C.; Mishra, S. Growth medium and nitrogen stress sparked biochemical and carotenogenic alterations in Scenedesmus sp. CCNM 1028. Bioresour. Technol. Rep. 2019, 7, 100194. [Google Scholar] [CrossRef]
- Telfer, A. What is β–carotene doing in the photosystem II reaction centre? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2002, 357, 1431–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Et Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Ho, S.-H.; Chen, C.-N.N.; Chen, C.-Y.; Ng, I.-S.; Jing, K.-J.; Chang, J.-S.; Lu, Y. Phototrophic cultivation of a thermo-tolerant Desmodesmus sp. for lutein production: Effects of nitrate concentration, light intensity and fed-batch operation. Bioresour. Technol. 2013, 144, 435–444. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Moreno, J.; Rodríguez, H.; Angeles Vargas, M.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: Factors determining lutein accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef]
- Ho, S.-H.; Xie, Y.; Chan, M.-C.; Liu, C.-C.; Chen, C.-Y.; Lee, D.-J.; Huang, C.-C.; Chang, J.-S. Effects of nitrogen source availability and bioreactor operating strategies on lutein production with Scenedesmus obliquus FSP-3. Bioresour. Technol. 2015, 184, 131–138. [Google Scholar] [CrossRef]
- Solovchenko, A.; Lukyanov, A.; Solovchenko, O.; Didi-Cohen, S.; Boussiba, S.; Khozin-Goldberg, I. Interactive effects of salinity, high light, and nitrogen starvation on fatty acid and carotenoid profiles in Nannochloropsis oceanica CCALA 804: Interactive effects of stresses on N. oceanica. Eur. J. Lipid Sci. Technol. 2014, 116, 635–644. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Polle, J.E.W.; Subba Rao, D.V. (Eds.) The Alga Dunaliella: Biodiversity, Physiology, Genomics and Biotechnology, 1st ed.; Science Publishers: Enfield, NH, USA, 2009; ISBN 978-1-57808-545-3. [Google Scholar]
- Rodrigues, E.; Mariutti, L.R.B.; Chisté, R.C.; Mercadante, A.Z. Development of a novel micro-assay for evaluation of peroxyl radical scavenger capacity: Application to carotenoids and structure–activity relationship. Food Chem. 2012, 135, 2103–2111. [Google Scholar] [CrossRef] [Green Version]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Naguib, Y.M.A. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Telfer, A.; Pascal, A.; Gall, A. Carotenoids in photosynthesis. In Carotenoids, Volume 4: Natural Functions; Carotenoids; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 2008; pp. 265–308. ISBN 978-3-7643-7499-0. [Google Scholar]
- Banskota, A.H.; Sperker, S.; Stefanova, R.; McGinn, P.J.; O’Leary, S.J.B. Antioxidant properties and lipid composition of selected microalgae. J. Appl. Phycol. 2018, 1–10. [Google Scholar] [CrossRef]
- Do Nascimento, T.C.; Nass, P.P.; Fernandes, A.S.; Vieira, K.R.; Wagner, R.; Jacob-Lopes, E.; Zepka, L.Q. Exploratory data of the microalgae compounds for food purposes. Data Brief. 2020, 29, 105182. [Google Scholar] [CrossRef] [PubMed]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Barba, F.J.; Lorenzo, J.M. Antioxidant potential of extracts obtained from macro- (Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata) and micro-algae (Chlorella vulgaris and Spirulina platensis) assisted by ultrasound. Medicines 2018, 5, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loganathan, B.G.; Orsat, V.; Lefsrud, M. Evaluation and interpretation of growth, biomass productivity and lutein content of Chlorella variabilis on various media. J. Environ. Chem. Eng. 2020, 8, 103750. [Google Scholar] [CrossRef]
- Schüler, L.M.; Santos, T.; Pereira, H.; Duarte, P.; Katkam, N.G.; Florindo, C.; Schulze, P.S.C.; Barreira, L.; Varela, J.C.S. Improved production of lutein and β-carotene by thermal and light intensity upshifts in the marine microalga Tetraselmis sp. CTP4. Algal Res. 2020, 45, 101732. [Google Scholar] [CrossRef]
- Lin, J.-H.; Lee, D.-J.; Chang, J.-S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef]
- Ho, S.-H.; Chan, M.-C.; Liu, C.-C.; Chen, C.-Y.; Lee, W.-L.; Lee, D.-J.; Chang, J.-S. Enhancing lutein productivity of an indigenous microalga Scenedesmus obliquus FSP-3 using light-related strategies. Bioresour. Technol. 2014, 152, 275–282. [Google Scholar] [CrossRef]
- Sánchez, J.F.; Fernández-Sevilla, J.M.; Acién, F.G.; Cerón, M.C.; Pérez-Parra, J.; Molina-Grima, E. Biomass and lutein productivity of Scenedesmus almeriensis: Influence of irradiance, dilution rate and temperature. Appl. Microbiol. Biotechnol. 2008, 79, 719–729. [Google Scholar] [CrossRef]
- Casal, C.; Cuaresma, M.; Vega, J.M.; Vilchez, C. Enhanced productivity of a lutein-enriched novel acidophile microalga grown on urea. Mar. Drugs 2011, 9, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, T.; Ganesan, P.; Li, Z.; Manabe, Y.; Hirata, T. Siphonaxanthin, a green algal carotenoid, as a novel functional compound. Mar. Drugs 2014, 12, 3660–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walne, P.R. Experiments in the Large Scale Culture of the Larvae of Ostrea Edulis L.; Fishery investigations (Great Britain. Ministry of Agriculture, Fisheries and Food); H.M.S.O.: London, UK, 1966; Volume 25. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jauffrais, T.; LeKieffre, C.; Schweizer, M.; Jesus, B.; Metzger, E.; Geslin, E. Response of a kleptoplastidic foraminifer to heterotrophic starvation: Photosynthesis and lipid droplet biogenesis. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauffrais, T.; LeKieffre, C.; Schweizer, M.; Geslin, E.; Metzger, E.; Bernhard, J.M.; Jesus, B.; Filipsson, H.L.; Maire, O.; Meibom, A. Kleptoplastidic benthic foraminifera from aphotic habitats: Insights into assimilation of inorganic C, N and S studied with sub-cellular resolution. Environ. Microbiol. 2019, 21, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Watanabe, J.; Oki, T.; Takebayashi, J.; Yada, H.; Wagaki, M.; Takano-Ishikawa, Y.; Yasui, A. Improvement and interlaboratory validation of the lipophilic oxygen radical absorbance capacity: Determination of antioxidant capacities of lipophilic antioxidant solutions and food extracts. Anal. Sci. 2016, 32, 171–175. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Van Heukelem, L.; Thomas, C.S. Computer-assisted high-performance liquid chromatography method development with applications to the isolation and analysis of phytoplankton pigments. J. Chromatogr. A 2001, 910, 31–49. [Google Scholar] [CrossRef]
- Serive, B.; Nicolau, E.; Bérard, J.-B.; Kaas, R.; Pasquet, V.; Picot, L.; Cadoret, J.-P. Community analysis of pigment patterns from 37 microalgae strains reveals new carotenoids and porphyrins characteristic of distinct strains and taxonomic groups. PLoS ONE 2017, 12, e0171872. [Google Scholar] [CrossRef]
- Welschmeyer, N.; Roy, S.; Llewellyn, C.; Egeland, E.S.; Johnsen, G. Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Cambridge University Press: Cambridge, UK, 2013; Volume 49, pp. 213–214. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antioxidant Activity | ||
|---|---|---|
| Correlation Coefficient | Adjusted R² | |
| Siph | 0.82 ** | 0.66 |
| Neo | 0.82 ** | 0.66 |
| XCP | 0.75 ** | 0.55 |
| Lut | 0.51 * | 0.22 |
| Lyco | 0.65 ** | 0.40 |
| β-car | 0.60 * | 0.33 |
| Total carotenoids | 0.80 ** | 0.62 |
| Biomass | Siph | Neo | XCP | Lut | Lyco | β-car | TC | ||
|---|---|---|---|---|---|---|---|---|---|
| Batch | Day 2 | 89.7 ± 0.7 c | 0.45 ± 0.08 b | 0.66 ± 0.10 b | 0.85 ± 0.05 bc | 0.53 ± 0.01 c | 0.26 ± 0.01 b | 1.06 ± 0.07 bc | 3.82 ± 0.30 bc |
| Day 3 | 120.8 ± 1.5 a | 0.77 ± 0.05 a | 1.16 ± 0.08 a | 1.47 ± 0.06 a | 0.87 ± 0.00 a | 0.40 ± 0.04 a | 1.47 ± 0.08 a | 6.13 ± 0.24 a | |
| Day 4 | 108.4 ± 1.1 b | 0.44 ± 0.04 b | 0.66 ± 0.05 b | 1.04 ± 0.05 b | 0.77 ± 0.03 ab | 0.32 ± 0.03 ab | 1.27 ± 0.08 ab | 4.50 ± 0.22 b | |
| Day 6 | 95.5 ± 2.6 c | 0.22 ± 0.03 c | 0.39 ± 0.03 c | 0.71 ± 0.04 b | 0.65 ± 0.04 bc | 0.24 ± 0.01 b | 1.02 ± 0.07 bc | 3.22 ± 0.21 c | |
| Continuous at steady state (Days 23–24) | 110.6 ± 5.7 ab | 0.29 ± 0.02 bc | 0.37 ± 0.03 c | 0.88 ± 0.09 bc | 0.61 ± 0.08 c | 0.27 ± 0.05 b | 0.98 ± 0.06 c | 3.40 ± 0.34 c | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coulombier, N.; Nicolau, E.; Le Déan, L.; Barthelemy, V.; Schreiber, N.; Brun, P.; Lebouvier, N.; Jauffrais, T. Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp. Mar. Drugs 2020, 18, 453. https://doi.org/10.3390/md18090453
Coulombier N, Nicolau E, Le Déan L, Barthelemy V, Schreiber N, Brun P, Lebouvier N, Jauffrais T. Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp. Marine Drugs. 2020; 18(9):453. https://doi.org/10.3390/md18090453
Chicago/Turabian StyleCoulombier, Noémie, Elodie Nicolau, Loïc Le Déan, Vanille Barthelemy, Nathalie Schreiber, Pierre Brun, Nicolas Lebouvier, and Thierry Jauffrais. 2020. "Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp" Marine Drugs 18, no. 9: 453. https://doi.org/10.3390/md18090453
APA StyleCoulombier, N., Nicolau, E., Le Déan, L., Barthelemy, V., Schreiber, N., Brun, P., Lebouvier, N., & Jauffrais, T. (2020). Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp. Marine Drugs, 18(9), 453. https://doi.org/10.3390/md18090453