The Comparative Immunotropic Activity of Carrageenan, Chitosan and Their Complexes


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characterisation of the Initial Polysaccharides
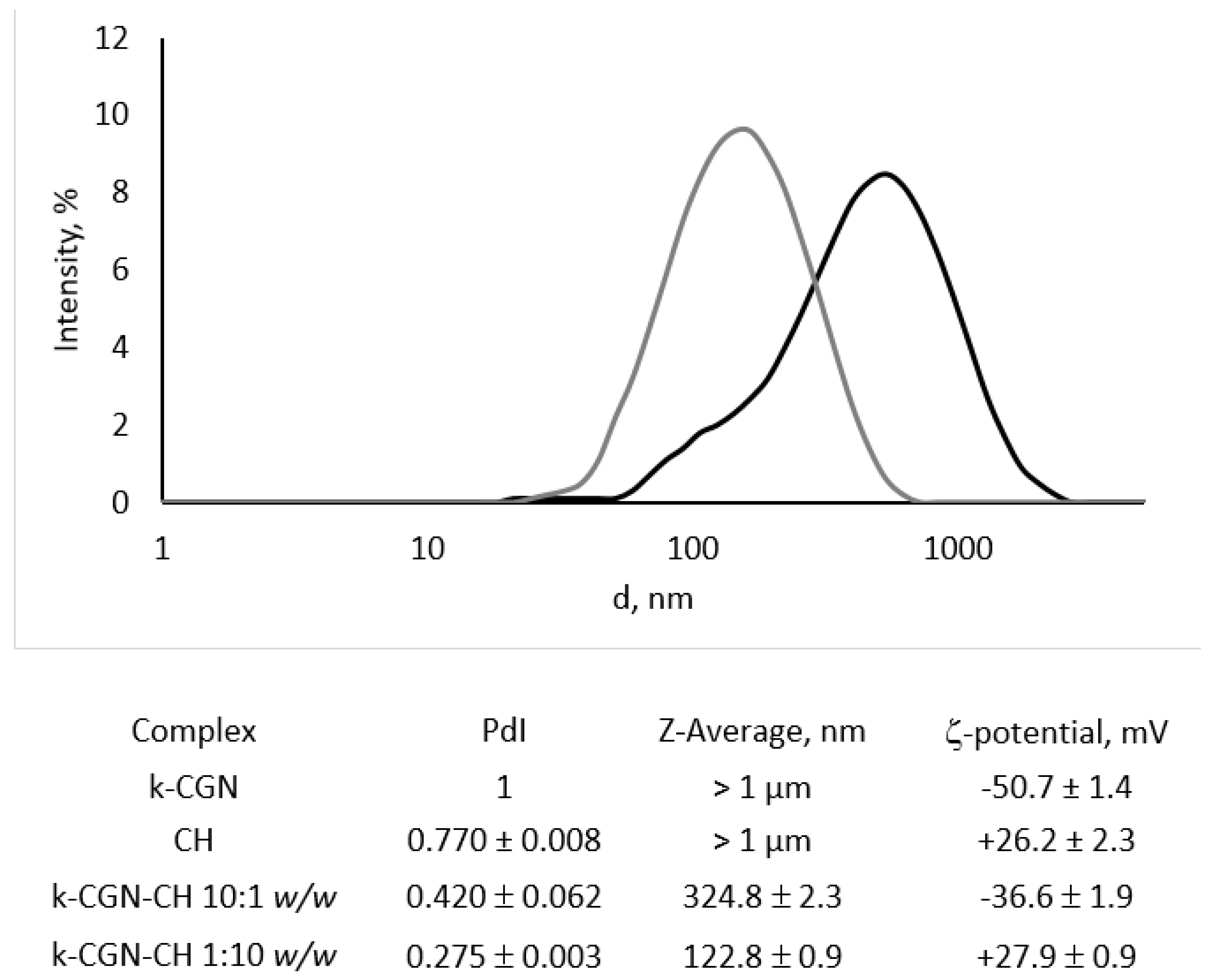
2.2. Characterization of PEC κ-CGN-CH
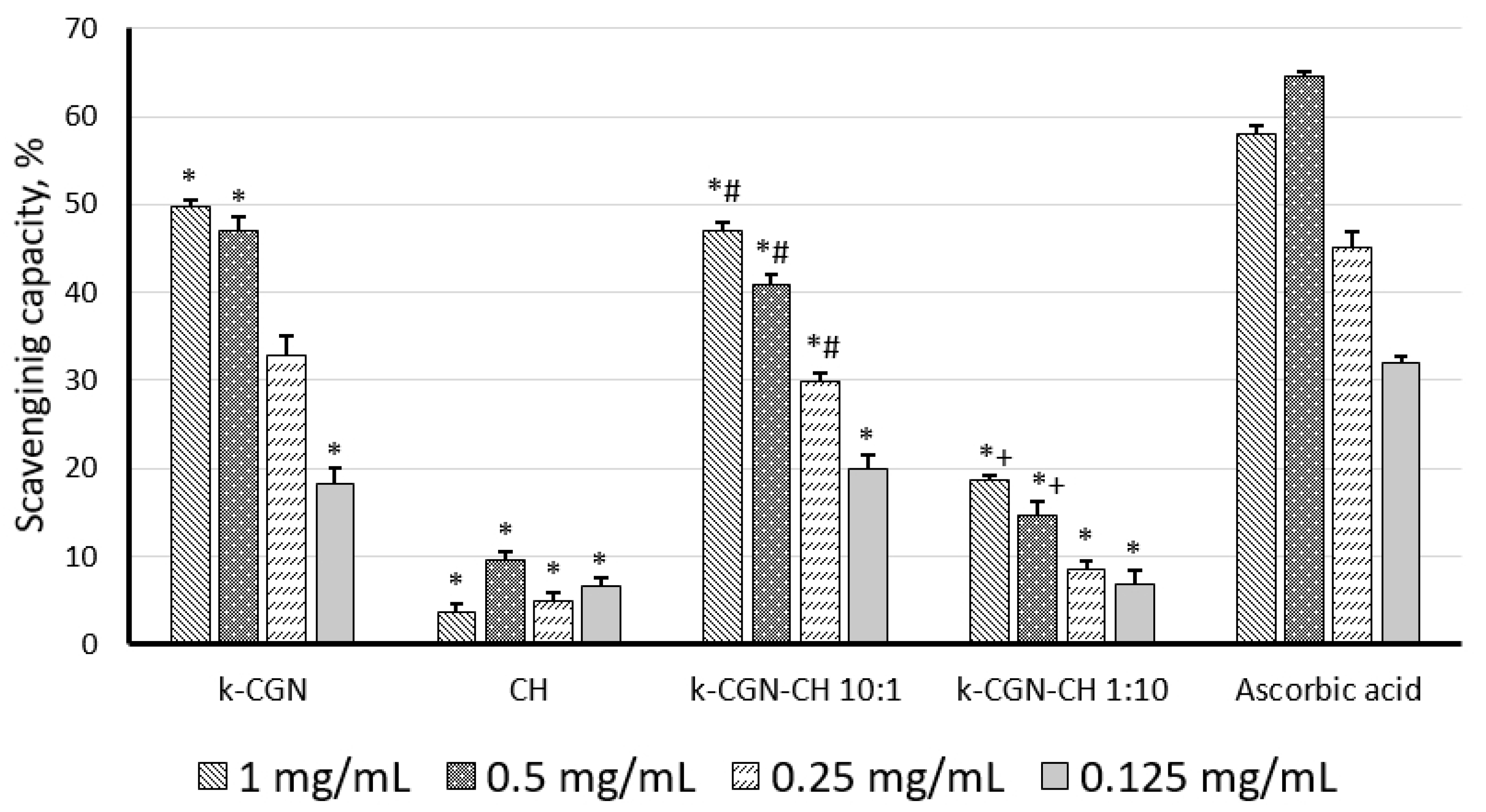
2.3. In Vitro Activity of the Initial Polysaccharides and Their Complexes
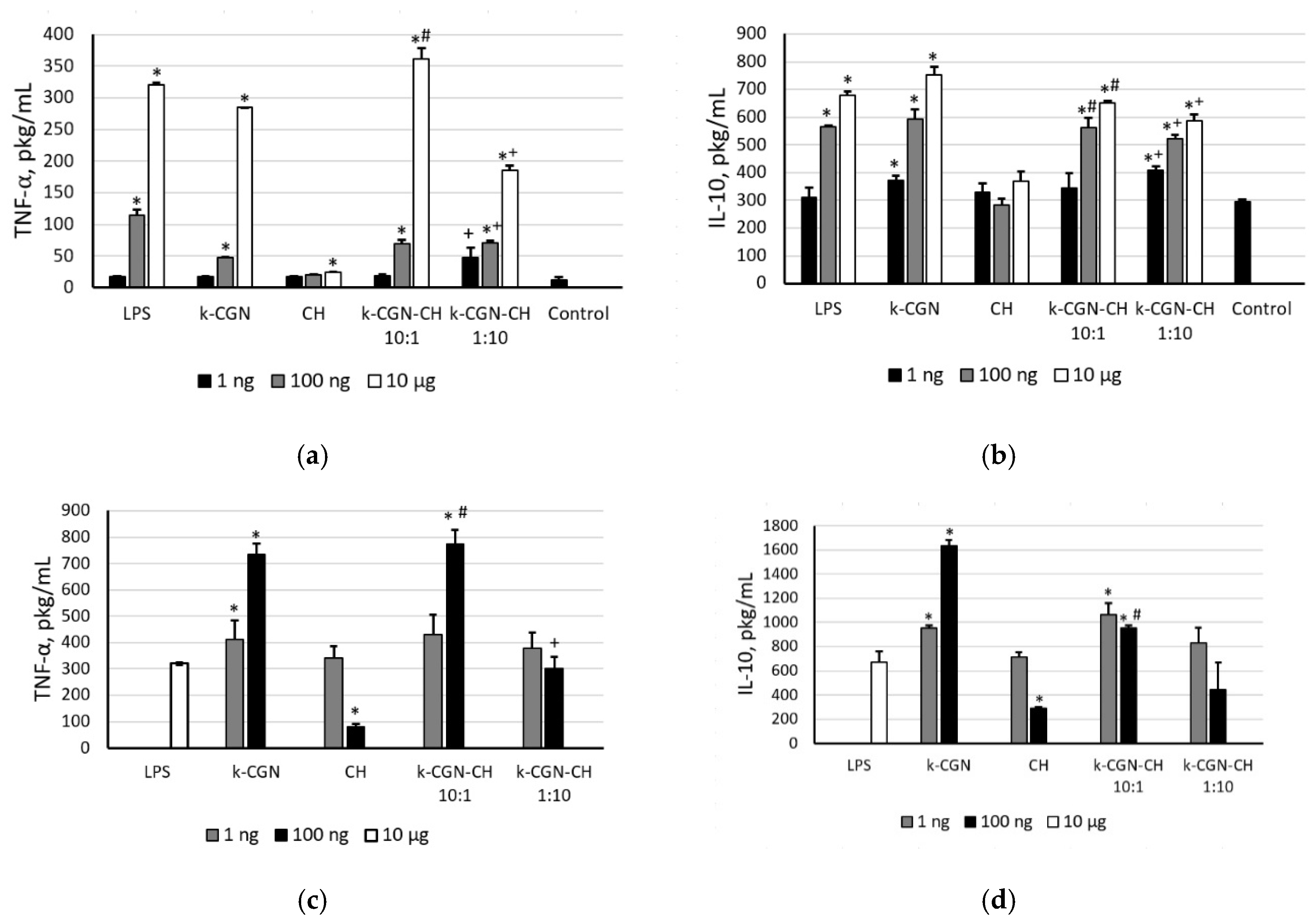
2.4. Ex Vivo Activity of the Initial Polysaccharides and Their Complexes
2.5. In Vivo Anti-Inflammatory Activity of the Initial Polysaccharides and Their Complexes
3. Discussion
4. Materials and Methods
4.1. Polysacharides
4.2. Complexes CGN:CH
4.3. Dynamic Light Scattering (DLS) and Electrophoretic Properties of the CGN:CH Complexes
4.4. Determination of NO Scavenging Capacity (Microplate)
4.5. IL-10 and TNF-α Inducing Activity on the Human Blood Cells
4.6. Biological Activity In Vivo
4.6.1. Animals
4.6.2. Anti-Inflammatory Activity
4.6.3. Histamine-Induced Mouse Paw Edema Test
4.6.4. Concavaline A-Induced Mouse Paw Edema Test
4.6.5. Delayed Type Hypersensitivity Reaction (DTH) Test
4.6.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Yermak, I.M.; Khotimchenko, Y.S. Chemical properties, biological activities and applications of carrageenans from red algae. Recent Adv. Mar. Biotechnol. 2003, 9, 207–255. [Google Scholar]
- Van de Velde, F.; Knutsen, S. 1H and 13C high resolution NMR spectroscopy of carrageenans: Application in research and industry. Trends Food Sci. Technol. 2002, 13, 73–92. [Google Scholar] [CrossRef]
- Yermak, I.M.; Barabanova, A.O.; Aminin, D.L.; Davydova, V.N.; Sokolova, E.V.; Solov’Eva, T.F.; Kim, Y.H.; Shin, K.S. Effects of structural peculiarities of carrageenans on their immunomodulatory and anticoagulant activities. Carbohydr. Polym. 2012, 87, 713–720. [Google Scholar] [CrossRef]
- Sokolova, E.V.; Byankina, A.O.; Kalitnik, A.A.; Kim, Y.H.; Bogdanovich, L.N.; Solov’Eva, T.F.; Yermak, I.M. Influence of red algal sulfated polysaccharides on blood coagulation and platelets activation in vitro. J. Biomed. Mater. Res. Part A 2014, 102, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. Focus on antivirally active sulfated polysaccharides: From structure-activity analysis to clinical evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, H.H.A.; Elshoubaky, G.A. Antiviral activity of sulfated polysaccharides carrageenan from some marine seaweeds. Int. J. Curr. Pharm. Rev. Res. 2016, 7, 34–42. [Google Scholar]
- Yuan, H.; Song, J.; Li, X.; Li, N.; Liu, S. Enhanced immunostimulatory and antitumor activity of different derivatives of κ-carrageenan oligosaccharides from Kappaphycus striatum. J. Appl. Phycol. 2011, 23, 59–65. [Google Scholar] [CrossRef]
- Lahaye, M.; Kaeffer, B. Seaweed dietary fibres: Structure, physico-chemical and biological properties relevant to intestinal physiology. Sci. Aliment. 1997, 17, 563–584. [Google Scholar]
- Weiner, M.L. Food additive carrageenan: Part II: A critical review of carrageenan in vivo safety studies. Crit. Rev. Toxicol. 2014, 44, 244–269. [Google Scholar] [CrossRef]
- Prajapati, V.D.; Maheriya, P.M.; Jani, G.K.; Solanki, H.K. Carrageenan: A natural seaweed polysaccharide and its applications. Carbohydr. Polym. 2014, 105, 97–112. [Google Scholar] [CrossRef]
- Singla, A.K.; Chawla, M. Chitosan: Some pharmaceutical and biological aspects-an update. J. Pharm. Pharmacol. 2001, 53, 1047–1067. [Google Scholar] [CrossRef] [PubMed]
- Kunio, N.R.; Riha, G.M.; Watson, K.M.; Differding, J.A.; Schreiber, M.A.; Watters, J.M. Chitosan based advanced hemostatic dressing is associated with decreased blood loss in a swine uncontrolled hemorrhage model. Am. J. Surg. 2013, 205, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Skjak-Braek, G.; Anthonsen, T.; Sandford, P.A. Chitin and Chitosan: Sources, Chemistry, Biochemistry, Physical Properties and Applications; Springer: Berlin/Heidelberg, Germany, 1989; ISBN 1851663959. [Google Scholar]
- Pérez-Recalde, M.; Matulewicz, M.C.; Pujol, C.A.; Carlucci, M.J. In vitro and in vivo immunomodulatory activity of sulfated polysaccharides from red seaweed Nemalion helminthoides. Int. J. Biol. Macromol. 2014, 63, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Xu, S.; Li, L.; Pan, T.; Shi, C.L.; Liu, H.; Cao, M.; Su, W.; Liu, G. In vitro and in vivo immunomodulatory activity of sulfated polysaccharide from Porphyra haitanensis. Carbohydr. Polym. 2017, 165, 189–196. [Google Scholar] [CrossRef]
- Suganya, A.M.; Sanjivkumar, M.; Chandran, M.N.; Palavesam, A.; Immanuel, G. Pharmacological importance of sulphated polysaccharide carrageenan from red seaweed Kappaphycus alvarezii in comparison with commercial carrageenan. Biomed. Pharmacother. 2016, 84, 1300–1312. [Google Scholar] [CrossRef]
- Sokolova, E.V.; Barabanova, A.O.; Homenko, V.A.; Solov’eva, T.F.; Bogdanovich, R.N.; Yermak, I.M. In Vitro and Ex Vivo Studies of Antioxidant Activity of Carrageenans, Sulfated Polysaccharides from Red Algae. Bull. Exp. Biol. Med. 2011, 150, 426–428. [Google Scholar] [CrossRef]
- Debnath, T.; Kim, D.; Lim, B. Natural Products as a Source of Anti-Inflammatory Agents Associated with Inflammatory Bowel Disease. Molecules 2013, 18, 7253–7270. [Google Scholar] [CrossRef]
- Bruno, A.; Tacconelli, S.; Patrignani, P. Variability in the Response to Non-Steroidal Anti-Inflammatory Drugs: Mechanisms and Perspectives. Basic Clin. Pharmacol. Toxicol. 2014, 114, 56–63. [Google Scholar] [CrossRef]
- Davydova, V.N.; Kalitnik, A.A.; Markov, P.A.; Volod’ko, A.V.; Popov, S.V.; Ermak, I.M. Cytokine-inducing and anti-inflammatory activity of chitosan and its low-molecular derivative. Appl. Biochem. Microbiol. 2016, 52, 476–482. [Google Scholar] [CrossRef]
- Kalitnik, A.A.; Anastyuk, S.D.; Barabanova, A.O.B.; Glazunov, V.P.; Yermak, I.M.; Marcov, P.A.; Popov, S.V.; Ovodov, Y.S. Gelling polysaccharide from Chondrus armatus and its oligosaccharides: The structural peculiarities and anti-inflammatory activity. Carbohydr. Polym. 2015, 115, 768–775. [Google Scholar] [CrossRef]
- Yermak, I.M.; Volod’ko, A.V.; Khasina, E.I.; Davydova, V.N.; Chusovitin, E.A.; Goroshko, D.L.; Kravchenko, A.O.; Solov’eva, T.F.; Maleev, V.V. Inhibitory effects of carrageenans on endotoxin-induced inflammation. Mar. Drugs 2020, 18, 248. [Google Scholar] [CrossRef] [PubMed]
- McKim, J.M.; Wilga, P.C.; Pregenzer, J.F.; Blakemore, W.R. The common food additive carrageenan is not a ligand for Toll-Like- Receptor 4 (TLR4) in an HEK293-TLR4 reporter cell-line model. Food Chem. Toxicol. 2015, 78, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.L. Toxicological properties of carrageenan. Agents Actions 1991, 32, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Matricardi, P.; Di Meo, C.; Coviello, T.; Hennink, W.E.; Alhaique, F. Interpenetrating polymer networks polysaccharide hydrogels for drug delivery and tissue engineering. Adv. Drug Deliv. Rev. 2013, 65, 1172–1187. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.; Sanchez, C.; Desobry-Banon, S.; Hardy, J. Structure and technofunctional properties of protein-polysaccharide complexes: A review. Crit. Rev. Food Sci. Nutr. 1998, 38, 689–753. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.J.; Snowden, M.J. The preparation, characterisation and applications of colloidal microgels. Adv. Colloid Interface Sci. 1995, 54, 73–91. [Google Scholar] [CrossRef]
- Volod’ko, A.V.; Davydova, V.N.; Chusovitin, E.; Sorokina, I.V.; Dolgikh, M.P.; Tolstikova, T.G.; Balagan, S.A.; Galkin, N.G.; Yermak, I.M. Soluble chitosan–carrageenan polyelectrolyte complexes and their gastroprotective activity. Carbohydr. Polym. 2014, 101, 1087–1093. [Google Scholar] [CrossRef]
- Volod’ko, A.V.; Davydova, V.N.; Nedashkovskaya, O.I.; Terentieva, N.A.; Chusovitin, E.A.; Galkin, N.G.; Yermak, I.M. Morphology, electrokinetic characteristics and the effect on biofilm formation of carrageenan:chitosan polyelectrolyte complexes. Int. J. Biol. Macromol. 2018, 117, 1118–1124. [Google Scholar] [CrossRef]
- Yermak, I.M.; Kim, Y.H.; Titlynov, E.A.; Isakov, V.V.; Solov’eva, T.F. Chemical structure and gel properties of carrageenans from algae belonging to the Gigartinaceae and Tichocarpaceae, collected from the Russian Pacific Coast. J. Appl. Phycol. 1999, 11, 41–48. [Google Scholar] [CrossRef]
- Wolfrom, M.; Han, T. The Sulfonation of Chitosan. J. Am. Chem. Soc. 1959, 870, 1764–1766. [Google Scholar] [CrossRef]
- Domszy, J.; Roberts, G. Evaluation of infrared spectroscopic techniques for analysing chitosan. Die Makromol. Chem. 1985, 186, 1671–1677. [Google Scholar] [CrossRef]
- Volod’Ko, A.V.; Davydova, V.N.; Barabanova, A.O.; Soloveva, T.F.; Ermak, I.M. Formation of soluble chitosan-carrageenan polyelectrolyte complexes. Chem. Nat. Compd. 2012, 48. [Google Scholar] [CrossRef]
- Davydova, V.N.; Volod’ko, A.V.; Sokolova, E.V.; Chusovitin, E.A.; Balagan, S.A.; Gorbach, V.I.; Galkin, N.G.; Yermak, I.M.; Solov’Eva, T.F. The supramolecular structure of LPS-chitosan complexes of varied composition in relation to their biological activity. Carbohydr. Polym. 2015, 123. [Google Scholar] [CrossRef] [PubMed]
- Ratanathanawongs Williams, S.K.; Lee, D. Field-flow fractionation of proteins polysaccharides, synthetic polymers, and supramolecular assemblies. J. Sep. Sci. 2006, 29, 1720–1732. [Google Scholar] [CrossRef]
- Kalim, M.D.; Bhattacharyya, D.; Banerjee, A.; Chattopadhyay, S. Oxidative DNA damage preventive activity and antioxidant potential of plants used in Unani system of medicine. BMC Complement. Altern. Med. 2010, 10. [Google Scholar] [CrossRef] [Green Version]
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef]
- Chen, S.K.; Tsai, M.L.; Huang, J.R.; Chen, R.H. In Vitro Antioxidant Activities of Low-Molecular-Weight Polysaccharides with Various Functional Groups. J. Agric. Food Chem. 2009, 57, 2699–2704. [Google Scholar] [CrossRef]
- Kim, K.W.; Thomas, R.L. Antioxidative activity of chitosans with varying molecular weights. Food Chem. 2007, 101, 308–313. [Google Scholar] [CrossRef]
- Duque, G.A.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.; Reist, M.; Mayer, J.M.; Felt, O.; Gurny, R. Structure and interactions in chitosan hydrogels formed by complexation or aggregation for biomedical applications. Eur. J. Pharm. Biopharm. 2004, 57, 35–52. [Google Scholar] [CrossRef]
- Rabson, A.; Roitt, I.M.; Delves, P.J. Really Essential Medical Immunology, 2nd ed.; Blackwell Publishing Ltd.: Oxford, UK, 2005. [Google Scholar]
- Moncada, S.; Palmer, R.M.J.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar] [PubMed]
- Stuehr, D.J.; Nathan, C.F. Nitric oxide: A macrophage product responsible for cytostasis and respiratory inhibition in tumor target cells. J. Exp. Med. 1989, 169, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Ajisaka, K.; Agawa, S.; Nagumo, S.; Kurato, K.; Yokoyama, T.; Arai, K.; Miyazaki, T. Evaluation and comparison of the antioxidative potency of various carbohydrates using different methods. J. Agric. Food Chem. 2009, 57, 3102–3107. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of In Vitro Antioxidant Activity of Polysaccharides. Oxid. Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gouwy, M.; Struyf, S.; Proost, P.; Van Damme, J. Synergy in cytokine and chemokine networks amplifies the inflammatory response. Cytokine Growth Factor Rev. 2005, 16, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Lin, C.; Bian, Z.; Xu, B. An insight into anti-inflammatory effects of fungal beta-glucans. Trends Food Sci. Technol. 2015, 41, 49–59. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Li, Y.; Hou, X.; Zeng, F. Correlation of structure to antitumor activities of five derivatives of a β-glucan from Poria cocos sclerotium. Carbohydr. Res. 2004, 339, 2567–2574. [Google Scholar] [CrossRef]
- Amann, R.; Schuligoi, R.; Lanz, I.; Donnerer, J. Histamine-induced edema in the rat paw-effect of capsaicin denervation and a CGRP receptor antagonist. Eur. J. Pharmacol. 1995, 279, 227–231. [Google Scholar] [CrossRef]
- Sokolova, E.V.; Kravchenko, A.O.; Sergeeva, N.V.; Davydova, V.N.; Bogdanovich, L.N.; Yermak, I.M. Effect of carrageenans on some lipid metabolism components in vitro. Carbohydr. Polym. 2020, 230. [Google Scholar] [CrossRef]
- Dumonde, D.C.; Maini, R.N. The clinical significance of mediators of cellular immunity. Clin. Exp. Allergy 1971, 1, 123–139. [Google Scholar] [CrossRef]
- Andreis, M.; Stastny, P.; Ziff, M. Experimental arthritis produced by injection of mediators of delayed hypersensitivity. Rheum.Annu. Rev. 1975, 6, 303–307. [Google Scholar] [CrossRef]
- Bento, C.A.M.; Cavada, B.S.; Oliveira, J.T.A.; Moreira, R.A.; Barja-Fidalgo, C. Rat paw edema and leukocyte immigration induced by plant lectins. Agents Actions 1993, 38, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Bernfield, M.; Park, P.W.; Reizes, O.; Fitzgerald, M.L.; Lincecum, J.; Zako, M.; Gotte, M.; Park, P.W.; Reizes, O.; Fitzgerald, M.L.; et al. Functions of cell surface heparan sulfate proteoglycans. Annu. Rev. Biochem. 1999, 68, 729–777. [Google Scholar] [CrossRef]
- Hartig, S.M.; Greene, R.R.; Dikov, M.M.; Prokop, A.; Davidson, J.M. Multifunctional nanoparticulate polyelectrolyte complexes. Pharm. Res. 2007, 24, 2353–2369. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Kamm, W.; Breitenbach, A.; Kaiserling, E.; Xiao, J.X.; Kissel, T. Biodegradable nanoparticles for oral delivery of peptides: Is there a role for polymers to affect mucosal uptake? Eur. J. Pharm. Biopharm. 2000, 50, 147–160. [Google Scholar] [CrossRef]
- Klein, J. Probing the interactions of proteins and nanoparticles. Proc. Natl. Acad. Sci. USA 2007, 104, 2029–2030. [Google Scholar] [CrossRef] [Green Version]
- Tenzer, S.; Docter, D.; Kuharev, J.; Musyanovych, A.; Fetz, V.; Hecht, R.; Schlenk, F.; Fischer, D.; Kiouptsi, K.; Reinhardt, C.; et al. Rapid formation of plasma protein corona critically affects nanoparticle pathophysiology. Nat. Nanotechnol. 2013, 8, 772–781. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, X.; Li, Y.; Chen, Q.; Liu, F.; Zhu, X.; Mei, L.; Song, X.; Liu, X.; Song, Z.; et al. Anti-inflammatory effects of a Mytilus coruscus α-D-glucan (MP-A) in activated macrophage cells via TLR4/NF-κB/MAPK pathway inhibition. Mar. Drugs 2017, 15, 294. [Google Scholar] [CrossRef] [Green Version]
- Naberezhnykh, G.A.; Gorbach, V.I.; Likhatskaya, G.N.; Davidova, V.N.; Solov’eva, T.F. Interaction of chitosans and their N-acylated derivatives with lipopolysaccharide of gram-negative bacteria. Biochemistry 2008, 73, 432–441. [Google Scholar] [CrossRef]
- Barabanova, A.O.; Shashkov, A.S.; Glazunov, V.P.; Isakov, V.V.; Nebylovskaya, T.B.; Helbert, W.; Solov’eva, T.F.; Yermak, I.M. Structure and properties of carrageenan-like polysaccharide from the red alga Tichocarpus crinitus (Gmel.) Rupr. (Rhodophyta, Tichocarpaceae). J. Appl. Phycol. 2008, 20, 1013–1020. [Google Scholar] [CrossRef]
- Sokolova, E.V.; Barabanova, A.O.; Bogdanovich, R.N.; Khomenko, V.A.; Solov’eva, T.F.; Yermak, I.M. In vitro antioxidant properties of red algal polysaccharides. Biomed. Prev. Nutr. 2011, 1, 161–167. [Google Scholar] [CrossRef]
- De Groote, D.; Zangerle, P.F.; Gevaert, Y.; Fassotte, M.F.; Beguin, Y.; Noizat-Pirenne, F.; Pirenne, J.; Gathy, R.; Lopez, M.; Dehart, I.; et al. Direct stimulation of cytokines (IL-1β, TNF-α, IL-6, IL-2, IFN-γ and GM-CSF) in whole blood. I. Comparison with isolated PBMC stimulation. Cytokine 1992, 4, 239–248. [Google Scholar] [CrossRef]
- Khlebnicova, T.S.; Piven, Y.A.; Baranovsky, A.V.; Lakhvich, F.A.; Sorokina, I.V.; Tolstikova, T.G. Fluorine-containing lupane triterpenoid acid derivatives: Design, synthesis and biological evaluation as potential anti-inflammatory agents. Steroids 2019, 147, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk-Cwynar, B.; Zaprutko, L.; Marciniak, J.; Lewandowski, G.; Szulc, M.; Kaminska, E.; Wachowiak, N.; Mikolajczak, P.L. The analgesic and anti-inflammatory effect of new oleanolic acid acyloxyimino derivative. Eur. J. Pharm. Sci. 2012, 47, 549–555. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Agent | Histamine-Induced Inflammation Test | Concanavalin A-Induced Inflammation Test | Delayed Type Hypersensitivity Reaction Test | |||
|---|---|---|---|---|---|---|
| IEI, % | AIA, % | IEI, % | AIA, % | IEI, % | AIA, % | |
| Control | 29.6 ± 2.0 | 0 | 18.6 ± 2.0 | 0 | 14.6 ± 0.6 | 0 |
| κ-CGN:CH 10:1 | 22.44 ± 1.53 ** ## | 37 | 20.0 ± 2.3 ## | 0 | 14.6 ± 1.3 | 0 |
| κ-CGN:CH 1:10 | 17.45 ± 3.04 ** | 51 | 20.9 ± 2.3 ## | 0 | 14.0 ± 1.6 | 4 |
| κ-CGN | 20.77 ± 2.56 **# | 37 | 9.8 ± 1.1 ** # | 47 | 6.4 ± 1.0 *** | 56 |
| CH | 16.70 ± 3.66 ** | 53 | 10.9 ± 1.6 * # | 41 | 15.7 ± 1.7 | 0 |
| Indomethacin | 12.61 ± 1.11 *** | 65 | − | − | − | − |
| Diclofenac | − | − | 7.2 ± 1.4 *** | 61 | − | − |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davydova, V.N.; Sorokina, I.V.; Volod’ko, A.V.; Sokolova, E.V.; Borisova, M.S.; Yermak, I.M. The Comparative Immunotropic Activity of Carrageenan, Chitosan and Their Complexes. Mar. Drugs 2020, 18, 458. https://doi.org/10.3390/md18090458
Davydova VN, Sorokina IV, Volod’ko AV, Sokolova EV, Borisova MS, Yermak IM. The Comparative Immunotropic Activity of Carrageenan, Chitosan and Their Complexes. Marine Drugs. 2020; 18(9):458. https://doi.org/10.3390/md18090458
Chicago/Turabian StyleDavydova, Viktoriya N., Irina V. Sorokina, Aleksandra V. Volod’ko, Ekaterina V. Sokolova, Marina S. Borisova, and Irina M. Yermak. 2020. "The Comparative Immunotropic Activity of Carrageenan, Chitosan and Their Complexes" Marine Drugs 18, no. 9: 458. https://doi.org/10.3390/md18090458
APA StyleDavydova, V. N., Sorokina, I. V., Volod’ko, A. V., Sokolova, E. V., Borisova, M. S., & Yermak, I. M. (2020). The Comparative Immunotropic Activity of Carrageenan, Chitosan and Their Complexes. Marine Drugs, 18(9), 458. https://doi.org/10.3390/md18090458