Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications



Abstract
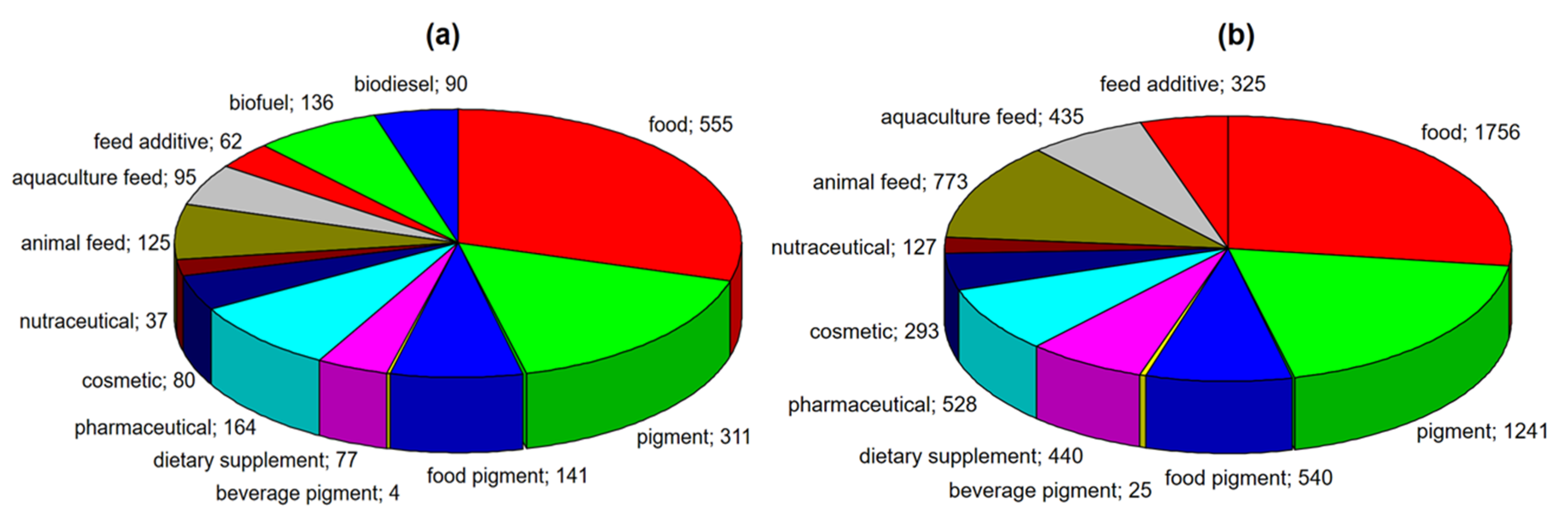
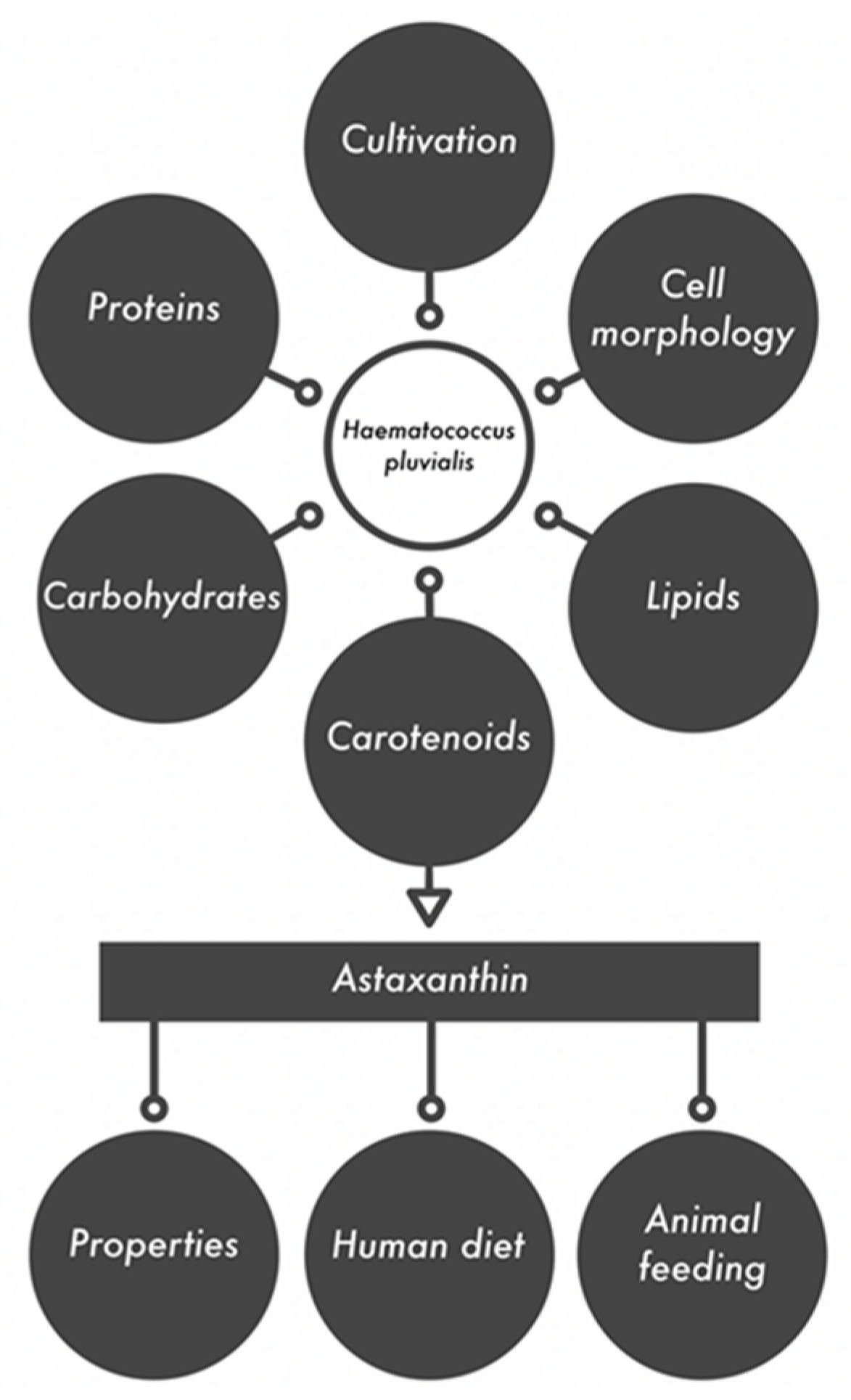
:1. Introduction
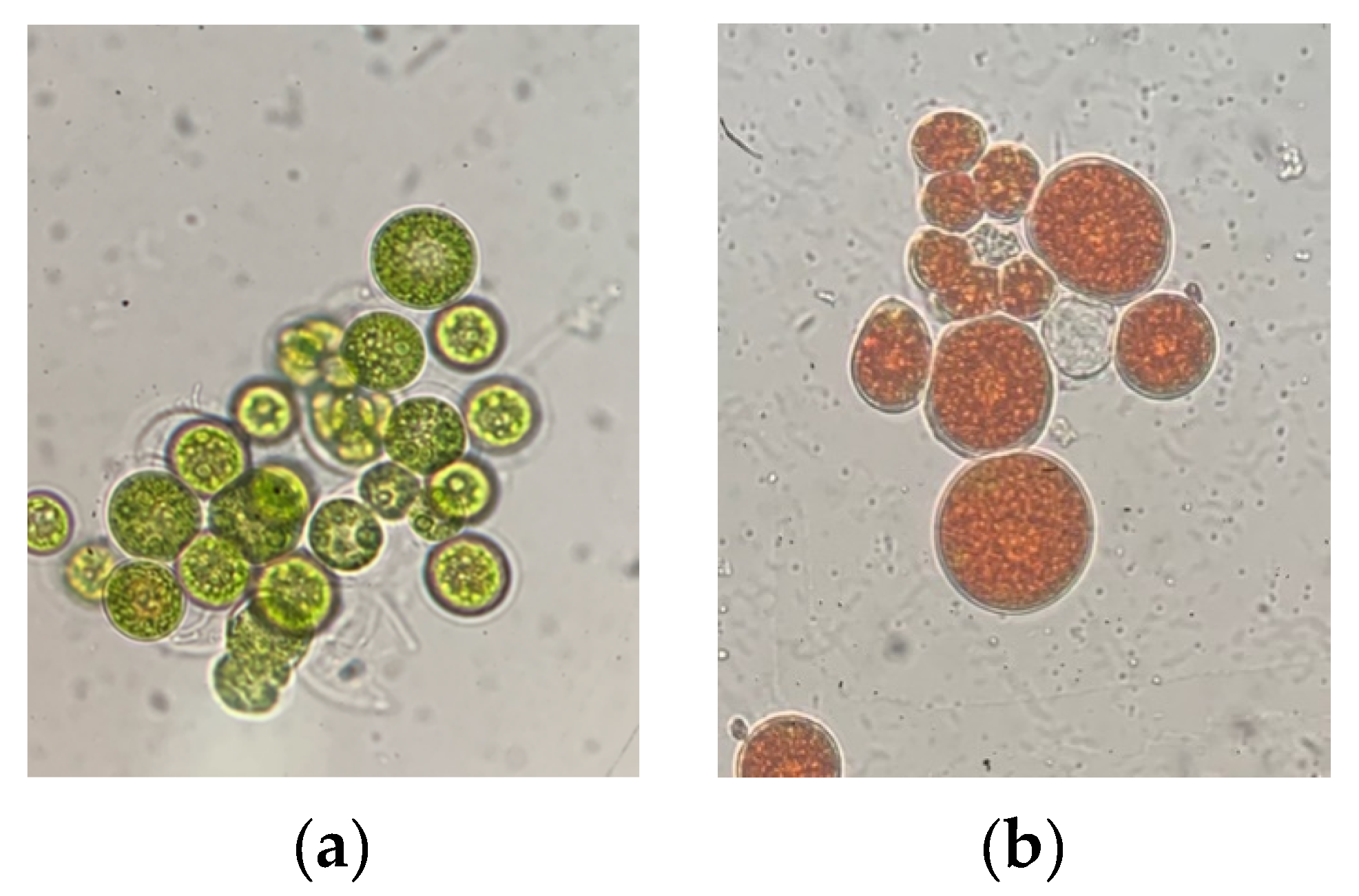
2. Cell Morphology of Haematococcus Pluvialis
3. Cultivation of Haematococcus Pluvialis
4. Biochemical Composition of Haematococcus Pluvialis
4.1. Proteins and Carbohydrates
4.2. Lipids
4.3. Carotenoids
5. Astaxanthin as a Valuable Biologically Active Compound
5.1. Astaxanthin Accumulation
5.2. Astaxanthin Recovery
6. Biological Properties of Astaxanthin
6.1. Anti-Lipid Peroxidation Properties
6.2. Anti-Inflammatory Effects
6.3. Anti–Diabetic Activity
6.4. Anticancer Activity
6.5. Other Potential Applications of Astaxanthin
7. Astaxanthin in the Human and Animal Diet
7.1. Human Diet
7.2. Animal Feed
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruiz-Dominguez, M.C.; Espinosa, C.; Paredes, A.; Palma, J.; Jaime, C.; Vilchez, C.; Cerezal, P. Determining the potential of Haematococcus pluvialis oleoresin as a rich source of antioxidants. Molecules 2019, 24, 4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariede, M.B.; Candido, T.M.; Morocho-Jacome, A.L.; Baby, A.R. Cosmetic attributes of algae—A review. Algal Res. 2017, 25, 483–489. [Google Scholar] [CrossRef]
- Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae characterization for consolidated and new application in human food, animal feed and nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahira, Y.; Ehsan, A.; Afifi, Z.; Masita, M.; Mohd, T. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. Thessalon. 2014, 21, 4. [Google Scholar]
- Skjånes, K.; Rebours, C.; Lindblad, P. Potential for green microalgae to produce hydrogen, pharmaceuticals and other high value products in a combined process. Crit. Rev. Biotechnol. 2013, 33, 172–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.; Miller, C.D. Microalgae as a source of bioplastics. Algal Green Chem. 2017, 6, 121–138. [Google Scholar]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Matos, Â.P. The Impact of microalgae in food science and technology. J. Am. Oil Chem. Soc. 2017, 94, 1333–1350. [Google Scholar] [CrossRef]
- Krichen, E.; Rapaport, A.; Le Floc’h, E.; Fouilland, E. Demonstration of facilitation between microalgae to face environmental stress. Sci. Rep. 2019, 9, 16076. [Google Scholar] [CrossRef] [Green Version]
- Nawrocka, D.; Kornicka, K.; Śmieszek, A.; Marycz, K. Spirulina platensis improves mitochondrial function impaired by elevated oxidative stress in adipose-derived mesenchymal stromal cells (ASCs) and intestinal epithelial cells (IECs), and enhances insulin sensitivity in equine metabolic syndrome (EMS) horses. Mar. Drugs 2017, 15, 237. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Ducan, J.; Barber, J. Astaxanthin accumulation in the green alga Haematococcus pluvialis: Effects of cultivation parameters. J. Integr. Plant Biol. 2007, 49, 447–451. [Google Scholar] [CrossRef]
- Specht, E.A.; Mayfield, S.P. Algae-based oral recombinant vaccines. Front. Microbiol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harasym, J.; Bogacz-Radomska, L. Colorants in foods—From past to present. Eng. Sci. Technol. 2016, 3, 21–35. [Google Scholar]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis. Single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [Green Version]
- Aflalo, C.; Meshulam, Y.; Zarka, A.; Boussiba, S. On the relative efficiency of two- vs. one-stage production of astaxanthin by the green alga Haematococcus pluvialis. Biotechnol. Bioeng. 2007, 98, 300–305. [Google Scholar] [CrossRef]
- Ota, S.; Kawano, S. Three-dimensional ultrastructure and hyperspectral imaging of metabolite accumulation and dynamics in Haematococcus and Chlorella. Microscopy 2019, 68, 57–68. [Google Scholar] [CrossRef]
- Fábregas, J.; Otero, A.; Maseda, A.; Domínguez, A. Two-stage cultures for the production of astaxanthin from Haematococcus pluvialis. J. Biotechnol. 2001, 89, 65–71. [Google Scholar] [CrossRef]
- Hagen, C.; Siegmund, S.; Braune, W. Ultrastructural and chemical changes in the cell wall of Haematococcus pluvialis (Volvocales, Chlorophyta) during aplanospore formation. Eur. J. Phycol. 2002, 37, 217–226. [Google Scholar] [CrossRef]
- Kim, J.H.; Affan, M.A.; Jang, J.; Kang, M.H.; Ko, A.R.; Jeon, S.M. Morphological, molecular, and biochemical characterization of astaxanthin-producing green microalga Haematococcus sp. KORDI03 Haematococcaceae, Chlorophyta) isolated from Korea. J. Microbiol. Biotechnol. 2015, 25, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Butler, T.; McDougall, G.; Campbell, R.; Stanley, M.; Day, J. Media screening for obtaining Haematococcus pluvialis red motile macrozooids rich in astaxanthin and fatty acids. Biology 2018, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Li, Y.; Hu, O. Astaxanthin in microalgae: Pathways, functions and biotechnological implications. Algae 2013, 28, 131–147. [Google Scholar] [CrossRef]
- Hanan, N.A.; Al-Shorgani, N.K.; Shukor, H.; Rahman, N.A.; Kali, M.S. Pre-optimization conditions for Haematococcus pluvialis growth. Int. J. Adv. Sci. Eng. Inf. Technol. 2013, 3, 168–171. [Google Scholar] [CrossRef]
- Hata, N.; Ogbonna, J.C.; Hasegawa, Y.; Taroda, H.; Tanaka, H. Production of astaxanthin by Haematococcus pluvialis in a sequential heterotrophic-photoautotrophic culture. J. Appl. Phycol. 2001, 13, 395–402. [Google Scholar] [CrossRef]
- Saha, S.K.; McHugh, E.; Hayes, J.; Moane, S.; Walsh, D.; Murray, P. Effect of various stress-regulatory factors on biomass and lipid production in microalga Haematococcus pluvialis. Bioresour. Technol. 2013, 128, 118–124. [Google Scholar] [CrossRef]
- Do, T.T.; Ong, B.N.; Nguyen Tran, M.L.; Nguyen, D.; Melkonian, M.; Tran, H.D. Biomass and astaxanthin productivities of Haematococcus pluvialis in an angled twin-layer porous substrate photobioreactor: Effect of inoculum density and storage time. Biology 2019, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, U.; Sarada, R.; Ramachandra Rao, S.; Ravishankar, G.A. Production of astaxanthin in Haematococcus pluvialis cultured in various media. Bioresour. Technol. 1999, 68, 197–199. [Google Scholar] [CrossRef]
- Bashan, Y.; Perez-Garcia, O. Microalgal heterotrophic and mixotrophic culturing for bio-refining: From metabolic routes to techno-economics. Algal Bioref. 2015, 2, 61–65. [Google Scholar]
- Orosa, M.; Franqueira, D.; Cid, A.; Abalde, J. Analysis and enhancement of astaxanthin accumulation in Haematococcus pluvialis. Bioresour. Technol. 2004, 96, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Aitken, D.; Antizar-Ladislao, B. Achieving a green solution: Limitations and focus points for sustainable algal fuels. Energies 2012, 5, 1613–1647. [Google Scholar] [CrossRef] [Green Version]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal carotenoids: A review of production, current markets, regulations, and future direction Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [Green Version]
- Dore, J.E.; Cysewski, G.R.; Cyanotech Corporation. Haematococcus Algae Meal as a Source of Natural Astaxanthin for Aquaculture Feeds. Available online: http://www.ruscom.com/cyan/web02/pdfs/naturose/nrtl09.pdf (accessed on 14 December 2019).
- Lee, K.Y.; Lee, S.H.; Lee, J.E.; Lee, S.Y. Biosorption of radioactive cesium from contaminated water by microalgae Haematococcus pluvialis and Chlorella vulgaris. J. Environ. Manag. 2019, 233, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Grewe, C.B.; Griehl, C. The Carotenoid Astaxanthin from Haematococcus Pluvialis. In Microalgal Biotechnology: Integration and Economy; Walter, C., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2012; Volume 1, pp. 129–144. [Google Scholar]
- Recht, L.; Zarka, A.; Boussiba, S. Patterns of carbohydrate and fatty acid changes under nitrogen starvation in the microalgae Haematococcus pluvialis and Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2012, 94, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Boussiba, S.; Vonshak, A. Astaxanthin accumulation in the green alga Haematococcus pluvialis. Plant Cell Physiol. 1991, 32, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Damiani, C.; Popovich, C.; Constenla, D.; Leonardi, P. Lipid analysis in Haematococcus pluvialis to assess its potential use as a biodiesel feedstock. Bioresour. Technol. 2010, 101, 3801–3807. [Google Scholar] [CrossRef]
- Cerón, M.C.; García-Malea, M.C.; Rivas, J.; Acien, F.G.; Fernandez, J.M.; Del Río, E.; Guerrero, M.G.; Molina, E. Antioxidant activity of Haematococcus pluvialis cells grown in continuous culture as a function of their carotenoid and fatty acid content. Appl. Microbiol. Biotechnol. 2007, 74, 1112–1119. [Google Scholar] [CrossRef]
- Liang, C.; Zhai, Y.; Xu, D.; Ye, N.; Zhang, X.; Wang, Y.; Zhang, W.; Yu, J. Correlation between lipid and carotenoid synthesis and photosynthetic capacity in Haematococcus pluvialis grown under high light and nitrogen deprivation stress. Grasas Aceites 2015, 66, 77. [Google Scholar]
- Lorenz, R.T. A Technical review of Haematococcus algae. In NatuRose™ Technical Bulletin #060; Cyanotech Corporation: Kailua-Kona, HI, USA, 1999. [Google Scholar]
- Scodelaro Bilbao, P.G.; Damiani, C.; Salvador, G.A.; Leonardi, P. Haematococcus pluvialis as a source of fatty acids and phytosterols: Potential nutritional and biological implications. J. Appl. Phycol. 2016, 28, 3283–3294. [Google Scholar] [CrossRef]
- Lei, A.; Chen, H.; Shen, G.; Hu, Z.; Chen, L.; Wang, J. Expression of fatty acid synthesis genes and fatty acid accumulation in Haematococcus pluvialis under different stressors. Biotech. Biofuels. 2012, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Dragos, N.; Bercea, V.; Bica, A.; Druga, B.; Nicoara, A.; Coman, C. Astaxanthin production from a new strain of Haematococcus pluvialis grown in batch culture. Ann. Roman. Soc. Cell Biol. 2010, 15, 353–361. [Google Scholar]
- Harker, M.; Tsavalos, A.J.; Young, A.J. Factors responsible for astaxanthin formation in the chlorophyte Haematococcus pluvialis. Bioresour. Technol. 1996, 55, 207–214. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Artaria, C. Astaxanthin in cardiovascular health and disease: Mechanisms of action, therapeutic merits, and knowledge gaps. Food Funct. 2017, 8, 39–63. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Yoshida, H.; Kondo, K. Potential anti-atherosclerotic properties of astaxanthin. Mar. Drugs 2016, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, P. Life cycle assessment of the production of the red antioxidant carotenoid astaxanthin by microalgae: From lab to pilot scale. J. Clean. Prod. 2014, 64, 332–344. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.E.; Choi, H.; Kwak, S.; Hwang, S.W.; Sung, J.Y.; Chang, S.; Sim, S.J. Rapid selection of astaxanthin-hyperproducing Haematococcus mutant via azide-based colorimetric assay combined with oil-based astaxanthin extraction. Bioresour. Technol. 2018, 267, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Miki, W. Biological functions and activities of animal carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Droop, M.R. Conditions governing haematochrome formation and loss in the alga Haematococcus pluvialis flotow. Arch. Mikrobiol. 1954, 20, 391–397. [Google Scholar] [CrossRef]
- Cerón García, M.; González López, C.; Fernández Sevilla, J.; Molina Grima, E. Preparative Recovery of Carotenoids from Microalgal Biomass. In Microbial Carotenoids. Methods in Molecular Biology; Barreiro, C., Barredo, J.L., Eds.; Humana Press: New York, NY, USA, 2018; Volume 1852. [Google Scholar]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Molino, A.; Mehariya, S.; Iovine, A.; Larocca, V.; Di Sanzo, G.; Martino, M.; Casella, P.; Chianese, S.; Musmarra, D. Extraction of astaxanthin and lutein from microalga Haematococcus pluvialis in the red phase using CO2 supercritical fluid extraction technology with ethanol as co-solvent. Mar. Drugs 2018, 16, 432. [Google Scholar] [CrossRef] [Green Version]
- Ruen-ngam, D.; Shotipruk, A.; Pavasant, P. Comparison of extraction methods for recovery of astaxanthin from Haematococcus pluvialis. Sep. Sci. Technol. 2011, 46, 64–70. [Google Scholar] [CrossRef]
- Khoo, K.S.; Lee, S.Y.; Ooi, C.W.; Fu, X.T.; Miao, X.L.; Ling, T.C.; Show, P.L. Recent advances in biorefinery of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2019, 288, 121606. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Yan, X.; Sun, L.; Yang, T.; Hu, X.; He, Z.; Liu, F.; Liu, X. Research progress on extraction, biological activities and delivery systems of natural astaxanthin. Trends Food Sci. Technol. 2019, 91, 354–361. [Google Scholar] [CrossRef]
- Dong, S.; Huang, Y.; Zhang, R.; Wang, S.; Liu, Y. Four different methods comparison for extraction astaxanthin from green alga Haematococcus pluvialis. J. Sci. World. 2014, 2014, 694305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yang, B.; Yan, B.; Yao, X. Supercritical fluid extraction of astaxanthin from Haematococcus pluvialis and its antioxidant potential in sunflower oil. Inno. Food Sci. Emerg. 2012, 13, 120–127. [Google Scholar] [CrossRef]
- Di Sanzo, G.; Mehariya, S.; Martino, M.; Larocca, V.; Casella, P.; Chianese, S.; Musmarra, D.; Balducchi, R.; Molino, A. Supercritical carbon dioxide extraction of astaxanthin, lutein, and fatty acids from Haematococcus pluvialis microalgae. Mar. Drugs 2018, 16, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beoletto, V.G.; de las Mercedes Oliva, M.; Marioli, J.M.; Demo, M.S. Antimicrobial natural products against bacterial biofilms. Antibiot. Resist. 2016, 14, 291–307. [Google Scholar]
- Bendicho, C.; Lavilla, I. Application of ultrasound assisted metal extractions. In Encyclopedia of Separation Science; Wilson, I.D., Adlard, E.R., Cooke, M., Poole, C., Eds.; Academic Press: London, UK, 2000; Volume 9, pp. 4421–4426. [Google Scholar]
- Zhao, X.; Zhang, X.; Liu, H.; Zhu, H.; Zhu, Y. Enzyme-assisted extraction of astaxanthin from Haematococcus pluvialis and its stability and antioxidant activity. Food Sci. Biotechnol. 2019, 28, 1637–1647. [Google Scholar] [CrossRef]
- Karpińska, A.; Gromadzka, G. Oxidative stress and natural antioxidant mechanisms: The role in neurodegeneration. From molecular mechanisms to therapeutic strategies. Postepy Hig Med. Dosw. 2013, 67, 43–53. [Google Scholar] [CrossRef]
- Das, K.C.; White, C.W. Redox system of the cell: Possible links and implications. Proc. Nat. Acad. Sci. USA 2002, 99, 9617–9618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutowicz, M. The influence of reactive oxygen species on the central nervous system. Postepy Hig Med. Dosw. 2011, 65, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Pashkow, F.J.; Watumull, D.G.; Campbell, C.L. Astaxanthin: A novel potential treatment for oxidative stress and inflammation in cardiovascular disease. Am. J. Cardiol. 2008, 101, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Kogure, K.; Abe, K.; Kimata, Y.; Kitahama, K.; Yamashita, E.; Terada, H. Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochim. Biophys. Acta 2001, 1512, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Dose, J.; Matsugo, S.; Yokokawa, H.; Koshida, Y.; Okazaki, S.; Seidel, U.; Eggersdorfer, M.; Rimbach, G.; Esatbeyoglu, T. Free radical scavenging and cellular antioxidant properties of astaxanthin. Int. J. Mol. Sci. 2016, 17, 103. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M.A. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Uszkadzanie składników komórek przez reaktywne formy tlenu. Druga twarz tlenu. Warszawa: Wyd. Nauk. PWN 2008, 2, 103–104. [Google Scholar]
- Bartosz, G. Jakie jest biologiczne znaczenie peroksydacji lipidów? Druga twarz tlenu. Warszawa: Wyd. Nauk. PWN 2008, 2, 101–103. [Google Scholar]
- Mano, C.M.; Guaratin, T.; Cardozo, K.; Colepicolo, P.; Bechara, E.; Barros, M. Astaxanthin restrains nitrative-oxidative peroxidation in mitochondrial-mimetic liposomes: A pre-apoptosis model. Mar. Drugs 2018, 16, 126. [Google Scholar] [CrossRef] [Green Version]
- Kamath, B.S.; Srikanta, B.M.; Dharmesh, S.M.; Sarada, R.; Ravishankar, G.A. Ulcer preventive and antioxidative properties of astaxanthin from Haematococcus pluvialis. Eur. J. Pharmacol. 2008, 590, 387–395. [Google Scholar] [CrossRef]
- Olson, J.A. Carotenoids: Absorption, transport, and metabolism of carotenoids in humans. Pure App. Chem. 2004, 66, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, T.; Hosoda, K.; Hirano, R. Inhibition of low density lipoprotein oxidation by astaxanthin. J. Atheroscler. Thromb. 2000, 7, 216–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, B.P.; Mathison, B.D.; Hayek, M.G.; Massimino, S.; Reinhart, G.A.; Park, J.S. Dietary astaxanthin enhances immune response in dogs. Vet. Immunol. Immunopathol. 2011, 140, 199–206. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, Y.S.; Song, G.G.; Park, J.J.; Chang, H.I. Protective effect of astaxanthin on naproxen-induced gastric antral ulceration in rats. Eur. J. Pharmacol. 2005, 514, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Chew, B.P.; Park, J.S. Carotenoids: Nutrition and health. In Carotenoids against Disease: Part C: The Immune System and Disease; Britton, G., Liaanen-Jensen, S., Pfander, H., Eds.; Birkhauser Press: Basel, Switzerland, 2009; Volume 5, pp. 363–382. [Google Scholar]
- Kettenmann, H.; Hanisch, U.K.; Noda, M.; Verkhratsky, A. Physiology of microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef]
- Grimmig, B.; Kim, S.H.; Nash, K.; Bickford, P.C.; Douglas Shytle, D.R. Neuroprotective mechanisms of astaxanthin: A potential therapeutic role in preserving cognitive function in age and neurodegeneration. GeroScience 2017, 39, 19–32. [Google Scholar] [CrossRef]
- Choi, S.K.; Park, Y.S.; Choi, D.K.; Chang, H.I. Effects of astaxanthin on the production of NO and the expression of COX-2 and iNOS in LPS-stimulated BV2 microglial cells. J. Microbiol. Biotechnol. 2008, 18, 1990–1996. [Google Scholar]
- Kim, Y.H.; Koh, H.K.; Kim, D.S. Down-regulation of IL-6 production by astaxanthin via ERK-, MSK-, and NFkappaB- mediated signals in activated microglia. Int. Immunopharmacol. 2010, 10, 1560–1572. [Google Scholar] [CrossRef]
- Park, S.K.; Kim, K.; Page, G.P.; Allison, D.B.; Weindruch, R.; Prolla, T.A. Gene expression profiling of aging in multiple mouse strains: Identification of aging biomarkers and impact of dietary antioxidants. Aging Cell 2009, 8, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Kozek, E. Insulin resistance and hyperinsulinemia-clinical aspects. Przegląd Lek. 1996, 53, 647–652. [Google Scholar]
- Uchiyama, K.; Naito, Y.; Hasegawa, G.; Nakamura, N.; Takahashi, J.; Yoshikawa, T. Astaxanthin protects β-cells against glucose toxicity in diabetic db/db mice. Redox Rep. 2002, 7, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Otton, R.; Marin, D.P.; Bolin, A.P.; Santos, R.C.; Polotow, T.G.; Sampaio, S.C.; De Barros, M.P. Astaxanthin ameliorates the redox imbalance in lymphocytes of experimental diabetic rats. Chem. Biol. Interact. 2010, 186, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneswari, S.; Anuradha, C.V. Astaxanthin prevents loss of insulin signaling and improves glucose metabolism in liver of insulin resistant mice. Can. J. Physiol. Pharmacol. 2012, 90, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneswari, S.; Arunkumar, E.; Viswanathan, P.; Anuradha, C.V. Astaxanthin restricts weight gain, promotes insulin sensitivity and curtails fatty liver disease in mice fed a obesity-promoting diet. Process. Biochem. 2010, 45, 1406–1414. [Google Scholar] [CrossRef]
- Landon, R.; Gueguen, V.; Petite, H.; Letourneur, D.; Pavon-Djavid, G.; Anagnostou, F. Impact of astaxanthin on diabetes pathogenesis and chronic complications. Mar. Drugs 2020, 18, 357. [Google Scholar] [CrossRef]
- Arunkumar, E.; Bhuvaneswari, S.; Anuradha, C.V. An intervention study in obese mice with astaxanthin, a marine carotenoid–effects on insulin signaling and pro-inflammatory cytokines. Food Funct. 2012, 3, 120–126. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Koyama, T.; Takahashi, J.; Yazawa, K. Effects of astaxanthin in obese mice fed a high-fat diet. Biosci. Biotechnol. Biochem. 2007, 71, 893–899. [Google Scholar] [CrossRef] [Green Version]
- Bandeira, S.d.; da Fonseca, L.J.S.; Guedes, G.d.; Rabelo, L.A.; Goulart, M.O.F.; Vasconcelos, S.M.L. Oxidative stress as an underlying contributor in the development of chronic complications in diabetes mellitus. Int. J. Mol. Sci. 2013, 14, 3265–3284. [Google Scholar] [CrossRef]
- Palloza, P.; Torelli, C.; Boninsegna, A.; Simone, R.; Catalano, A.; Mele, M.C.; Picci, N. Growth inhibitory effects of the astaxanthin-rich alga Haematococcus pluvialis in human colon cancer cells. Cancer Lett. 2009, 283, 108–117. [Google Scholar] [CrossRef]
- Chew, B.P.; Park, J.S. Carotenoid action on the immune response. J. Nutr. 2004, 134, 257–261. [Google Scholar] [CrossRef]
- Chew, B.P.; Park, J.S.; Wong, M.W.; Wong, T.S. A comparison of the anticancer activities of dietary β-carotene, canthaxanthin and astaxanthin in mice in vivo. Anticancer Res. 1999, 19, 1849–1853. [Google Scholar]
- Nakano, M.; Onodera, A.; Saito, E.; Tanabe, M.; Yajima, K.; Takahashi, J.; Ngyuen, V.C. Effect of astaxanthin in combination with alpha-tocopherol or ascorbic acid against oxidative damage in diabetic ODS rats. J. Nutr. Sci. Vitaminol. 2008, 54, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jyonouchi, H.; Sun, S.; Iijima, K.; Gross, M.D. Antitumor activity of astaxanthin and its mode of action. Nutr. Cancer 2008, 36, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Song, X.D.; Zhang, J.J.; Wang, M.R.; Liu, W.B.; Gu, X.B.; Lv, C.J. Astaxanthin induces mitochondria-mediated apoptosis in rat hepatocellular carcinoma CBRH-7919 cells. Biol. Pharm. Bull. 2011, 34, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, G.; Nakamura, M.; Zhao, Q.; Iguchi, T.; Goto, H.; Sankawa, U.; Watanabe, H. Antihypertensive and neuroprotective effects of astaxanthin in experimental animals. Biol. Pharm. Bull. 2005, 28, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, G.; Goto, H.; Oda, S.; Sankawa, U.; Matsumoto, K.; Watanabe, H. Antihypertensive potential and mechanism of action of astaxanthin: III. Antioxidant and histopathological effects in spontaneously hypertensive rats. Biol. Pharm. Bull. 2006, 29, 684–688. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gao, H.; Zhang, L.; Chen, C.; Yang, W.; Deng, Q.; Huang, Q.; Huang, F. A combination of flaxseed oil and astaxanthin alleviates atherosclerosis risk factors in high fat diet fed rats. Lipids Health Dis. 2014, 13, 63. [Google Scholar] [CrossRef] [Green Version]
- Saito, M.; Yoshida, K.; Saito, W.; Fujiya, A.; Ohgami, K.; Kitaichi, N.; Tsukahara, H.; Ishida, S.; Ohno, S. Astaxanthin increases choroidal blood flow velocity. Graef. Archive Clin. Exp. 2012, 250, 239–245. [Google Scholar] [CrossRef]
- Mosaad, Y.O.; Abd El Khalik Gobba, N.; Hussein, M.A. Astaxanthin; a promising protector against gentamicin-induced nephrotoxicity in rats. Curr. Pharm. Biotechnol. 2016, 17, 1189–1197. [Google Scholar] [CrossRef]
- Otsuka, T.; Shimazawa, M.; Inoue, Y.; Nakano, Y.; Ojino, K.; Izawa, H.; Tsuruma, K.; Ishibashi, T.; Hara, H. Astaxanthin protects against retinal damage: Evidence from in vivo and in vitro retinal ischemia and reperfusion models. Curr. Eye Res. 2016, 41, 1465–1472. [Google Scholar] [CrossRef]
- Curek, G.D.; Cort, A.; Yucel, G.; Demir, N.; Ozturk, S.; Elpek, G.O.; Savas, B.; Aslan, M. Effect of astaxanthin on hepatocellular injury following ischemia/reperfusion. Toxicology 2010, 267, 147–153. [Google Scholar] [CrossRef]
- Giannaccare, G.; Pellegrini, M.; Senni, C.; Bernabei, F.; Scorcia, V.; Cicero, A.F.G. Clinical applications of astaxanthin in the treatment of ocular diseases: Emerging insights. Mar. Drugs 2020, 18, 239. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, A.; Dargahi, L.; Abbaszadeh, F.; Pourgholami, M.H.; Asgari, A.; Manoochehri, M.; Jorjani, M. Neuroprotective effects of astaxanthin in a rat model of spinal cord injury. Behav. Brain Res. 2017, 329, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Zhang, X.; Wu, Q.; Li, W.; Wang, C.X.; Xie, G.B.; Zhou, X.M.; Shi, J.X.; Zhou, M.L. Astaxanthin offers neuroprotection and reduces neuroinflammation in experimental subarachnoid hemorrhage. J. Surg. Res. 2014, 192, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Aneva, I.Y.; Farzaei, M.H.; Sobarzo-Sánchez, E. The Neuroprotective effects of astaxanthin: Therapeutic targets and clinical perspective. Molecules 2019, 24, 2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanoni, F.; Vakarelova, M.; Zocctelli, G. Development and characterization of astaxanthin-containing whey protein based nanoparticles. Mar. Drugs. 2019, 17, 627. [Google Scholar] [CrossRef] [Green Version]
- Hossain, A.K.M.M.; Brennan, M.A.; Mason, S.L.; Guo, X.; Zeng, X.A.; Brennan, C.S. The effect of astaxanthin-rich microalgae “Haematococcus pluvialis” and wholemeal flours incorporation in improving the physical and functional properties of cookies. Foods 2017, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Mercke Odeberg, J.; Lignell, A.; Pettersson, A.; Höglund, P. Oral bioavailability of the antioxidant astaxanthin in humans is enhanced by incorporation of lipid based formulations. Eur. J. Pharm. Sci. 2003, 19, 299–304. [Google Scholar] [CrossRef]
- Ranga Rao, A.; Harshvardhan Reddy, A.; Aradhya, S.M. Antibacterial properties of Spirulina platensis, Haematococcus pluvialis, Botryococcus braunii micro algal extracts. Curr. Trends Biotechnol. Pharm. 2010, 4, 809–819. [Google Scholar]
- Furr, H.C.; Clark, R.M. Intestinal absorption and tissue distribution of carotenoids. J. Nutr. Biochem. 1997, 8, 364–377. [Google Scholar] [CrossRef]
- Barros, M.P.; Marin, D.P.; Bolin, A.P.; de Cássia Santos Macedo, R.; Campoio, T.R.; Fineto, C., Jr.; Guerra, B.A.; Polotow, T.G.; Vardaris, C.; Mattei, R.; et al. Combined astaxanthin and fish oil supplementation improves glutathione-based redox balance in rat plasma and neutrophils. Chem. Biol. Interact. 2012, 197, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Ishikura, M.; Maoka, T. Bioavailability of astaxanthin in Haematococcus algal extract: The effects of timing of diet and smoking habits. Biosci. Biotechnol. Biochem. 2009, 73, 1928–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van het Hof, K.H.; West, C.E.; Weststrate, J.A.; Hautvast, J.G. Dietary factors that affect the bioavailability of carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Thyssandier, V.; Lyan, B.; Borel, P. Main factors governing the transfer of carotenoids from emulsion lipid droplets to micelles. Biochim. Biophys. Acta. 2001, 1533, 285–292. [Google Scholar] [CrossRef]
- Harrison, E.H. Mechanisms involved in the intestinal absorption of dietary vitamin A and provitamin A carotenoids. BBA 2012, 1821, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.C.; Yusoff, F.; Shariff, M.; Kamarudin, M.S. Astaxanthin as feed supplement in aquatic animals. Rev. Aquacult. 2018, 10, 738–773. [Google Scholar] [CrossRef]
- Ranga Rao, A.; Baskaran, V.; Sarada, R.; Ravishankar, G.A. In vivo bioavailability and antioxidant activity of carotenoids from micro algal biomass—A repeated dose study. Food Res. Int. 2013, 54, 711–717. [Google Scholar] [CrossRef]
- Stewart, J.S.; Lignell, A.; Pettersson, A.; Elfving, E.; Soni, M.G. Safety assessment of astaxanthin rich microalgae biomass: Acute and subchronic toxicity studies in rats. Food Chem. Toxicol. 2008, 46, 3030–3036. [Google Scholar] [CrossRef]
- EFSA, NDA Panel. Scientific opinion on the safety of astaxanthin-rich ingredients (AstaREAL A1010 and AstaREAL L10) as novel food ingredients EFSA panel on dietetic products, nutrition and allergies (NDA). EFSA J. 2014, 12, 1–35. [Google Scholar]
- Spiller, G.A.; Dewell, A. Safety of an astaxanthin rich Haemaotoccu pluvialis algal extract: A randomized clinical trial. J. Med. Food. 2003, 6, 51–56. [Google Scholar] [CrossRef]
- Kupcinskas, L.; Lafolie, P.; Lignell, A.; Kiudelis, G.; Jonaitis, L.; Adamonis, K.; Andersen, L.P.; Wadström, T. Efficacy of the natural antioxidant astaxanthin in the treatment of functional dyspepsia in patients with or without Helicobacter pylori infection: A prospective, randomized, double blind, and placebo-controlled study. Phytomedicine 2008, 15, 391–399. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, H.; Zhang, X.; Zhang, G.; Zhu, H. Astaxanthin from Haematococcus pluvialis microencapsulated by spray drying: Characterization and antioxidant activity. J. Am. Oil Chem. Soc. 2019, 96, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Ranga Rao, A.; Sarada, R.; Ravishankar, G.A. Stabilization of astaxanthin in edible oils and its use as an antioxidant. J. Sci. Food Agric. 2007, 87, 957–965. [Google Scholar]
- Higuera-Ciapara, I.; Felix-Valenzuela, L.; Goycoolea, F.M.; Argüelles-Monal, W. Microencapsulation of astaxanthin in a chitosan matrix. Carbohydr. Polym. 2004, 56, 41–45. [Google Scholar] [CrossRef]
- Tachaprutinun, A.; Udomsup, T.; Luadthong, C.; Wanichwecharungruang, S. Preventing the thermal degradation of astaxanthin through nanoencapsulation. Int. J. Pharm. 2009, 374, 119–124. [Google Scholar] [CrossRef]
- Pogorzelska, E.; Hamułka, J.; Wawrzyniak, A. Astaksantyna—Budowa, właściwości i możliwości zastosowania w żywności funkcjonalnej. Żywność. Nauka. Technol. Jakość. 2016, 1, 5–16. [Google Scholar]
- Hasan, M.R.; Chakrabarti, R. Use of algae and aquatic macrophytes as feed in small-scale aquaculture: A review. FAO Fish. Aquacult. Tech. Paper. 2009, 531, 100–123. [Google Scholar]
- Hansen, O.J.; Puvanendran, V.; Bangera, R. Broodstock diet with water and astaxanthin improve condition and egg output of brood fish and larval survival in Atlantic cod, Gadus morhua. L. Aquac. Res. 2016, 47, 819–829. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Deng, D.F.; Dominy, W. A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture 2012, 354, 50–55. [Google Scholar] [CrossRef]
- Torrissen, O.J. Dietary delivery of carotenoids. In Antioxidants in Muscle Foods: Nutritional Strategies to Improve Quality; Decker, E., Faustman, C., Lopez-Bote, C.J., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2000; pp. 289–313. [Google Scholar]
- Sheikhzadeh, N.; Tayefi-Nasrabadi, H.; Oushani, A.K.; Enferadi, M.H.N. Effects of Haematococcus pluvialis supplementation on antioxidant system and metabolism in rainbow trout (Oncorhynchus mykiss). Fish. Physiol. Biochem. 2012, 38, 413–419. [Google Scholar] [CrossRef]
- Yang, Y.X.; Kim, Y.J.; Jin, Z.; Lohakare, J.D.; Kim, C.H.; Ohh, S.H.; Lee, S.H.; Choi, J.Y.; Chae, B.J. Effects of dietary supplementation of astaxanthin on production performance, egg quality in layers and meat quality in finishing pigs. AJAS 2006, 19, 1019–1025. [Google Scholar] [CrossRef]
- Inborr, J. Haematococcus, the Poultry Pigmentor. Feed Mix. 1998, 6, 31–34. [Google Scholar]
- Bergstrom, J.R.; Nelssen, J.L.; Houser, T.; Gunderson, J.A.; Gipe, A.N.; Jacela, J.; Benz, J.M.; Sulabo, R.C.; Tokac, M.D. Effects of dietary astaxanthin on the growth performance and carcass characteristics of finishing pigs. Fish. Pig Nutr. Manag. 2009, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Waldenstedt, L.; Inborr, J.; Hansson, I.; Elwinger, K. Effects of astaxanthin-rich algal meal (Haematococcus pluvalis) on growth performance, caecal campylobacter and clostridial counts and tissue astaxanthin concentration of broiler chickens. Anim. Feed Sci. Technol. 2003, 108, 119–132. [Google Scholar] [CrossRef]
- Smith, G.C.; Belk, K.E.; Sofos, J.N.; Tatum, J.D.; Williams, S.N. Economic implications of improved color stability in beef. Antioxid. Muscle Foods Nutr. Strateg. Improv. Qual. 2000, 118, 397–426. [Google Scholar]
- Higgins, F.M.; Kerry, J.P.; Buckley, D.J.; Morrissey, P.A. Dietary supplementation versus direct postmortem addition of α-tocopherol on lipid and colour stability in cooked turkey breast patties. Food Res. 1998, 30, 373–383. [Google Scholar]
- Kim, K.; Lim, J.; Shin, M.; Choi, Y.; Lee, S.; Cho, S. Effect of dietary combined probiotics (AnyLac) supplementation contained with Phaffia rhodozyma on the growth performance and meat quality of pigs. Kor. J. Anim. Sci. Technol. 2008, 50, 657–666. [Google Scholar]
- Carr, C.C.; Johnson, D.D.; Brendemuhl, J.H.; Gonzalez, J.M. Fresh pork quality and shelf-life characteristics of meat from pigs supplemented with natural astaxanthin in the diet. Prof. Anim. Sci. 2010, 26, 18–25. [Google Scholar] [CrossRef]
- Serwotka-Suszczak, A.M.; Marcinkowska, K.A.; Śmieszek, A.; Michalak, I.; Grzebyk, M.; Wiśniewski, M.; Marycz, K.M. The Haematococcus pluvialis extract enriched by bioaccumulation process with Mg (II) ions improves insulin resistance in equine adipose-derived stromal cells (EqASCs). Biomed. Pharmacother. 2019, 116, 108972. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Kim et al., 2015 [19] | Lorenz, 1999 [39] | Scodelaro Bilbao et al., 2016 [40] | Lei et al. 2012 [41] |
|---|---|---|---|---|
| C12:0 lauric | N/A | 0.1 | N/A | 0.28 |
| C14:0 myristic | 0.1 | 0.5 | 1.99 | 0.65 |
| C15:0 pentadecanoic acid | 0.1 | N/A | N/A | 0.25 |
| C16:0 palmitic | 13.7 | 29 | 22.9 | 12.7 |
| C16:1 palmitoleic | 0.5 | 0.6 | 0.35 | 0.7 |
| C16:2 | 0.4 | N/A | N/A | N/A |
| C16:3 | 3.5 | N/A | N/A | N/A |
| C16:4 | 3.3 | N/A | N/A | N/A |
| C17:0 margaric | N/A | 0.2 | N/A | 0.23 |
| C17:1 margaroleic | N/A | 1.3 | N/A | 0.0 |
| C18:0 stearic | 0.7 | 2.1 | 1.15 | 4.79 |
| C18:1 oleic | 4.9 | 25.9 | 16.3 | 11.2 |
| C18:2 linoleic | 24.9 | 20.8 | 23.9 | 13.0 |
| C18:3 linolenic | 39.7 | 12.8 | 12.5 | 2.84 |
| C18:4 octadecatetraenoic | 5.8 | 1.4 | N/A | N/A |
| C20:0 arachidic | N/A | 0.6 | N/A | 0.35 |
| C20:1 gadoleic | 0.5 | 0.3 | N/A | 1.3 |
| C20:2 eicosadenoic | N/A | 1.2 | 2.21 | 0.87 |
| C20:3 eicosatrienoic gamma | N/A | 0.5 | N/A | 0.18 |
| C20:4 arachidonic | 0.9 | 1.4 | 1.92 | 1.77 |
| C20:5 eicosapentaenoic | 0.6 | 0.6 | 0.66 | 0.99 |
| C22:0 behenic | N/A | 0.4 | N/A | 0.16 |
| C24:0 lignoceric | 0.3 | 0.2 | 0.33 | 0.4 |
| C24:1 nervonic acid | 0.1 | 0.1 | N/A | 0.14 |
| Σ SFAs | 15 | 33.2 | 25.3 | 19.8 |
| Σ ΜUFAs | 6 | 28.1 | 16.6 | 12.97 |
| Σ PUFAs | 79.1 | 38.7 | 41.2 | 19.65 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications. Mar. Drugs 2020, 18, 459. https://doi.org/10.3390/md18090459
Mularczyk M, Michalak I, Marycz K. Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications. Marine Drugs. 2020; 18(9):459. https://doi.org/10.3390/md18090459
Chicago/Turabian StyleMularczyk, Malwina, Izabela Michalak, and Krzysztof Marycz. 2020. "Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications" Marine Drugs 18, no. 9: 459. https://doi.org/10.3390/md18090459
APA StyleMularczyk, M., Michalak, I., & Marycz, K. (2020). Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications. Marine Drugs, 18(9), 459. https://doi.org/10.3390/md18090459