
Selection of Culture Conditions and Cell Morphology for Biocompatible Extraction of β-Carotene from Dunaliella salina




Abstract
:1. Introduction
2. Results
2.1. β-Carotene Extraction from Cells
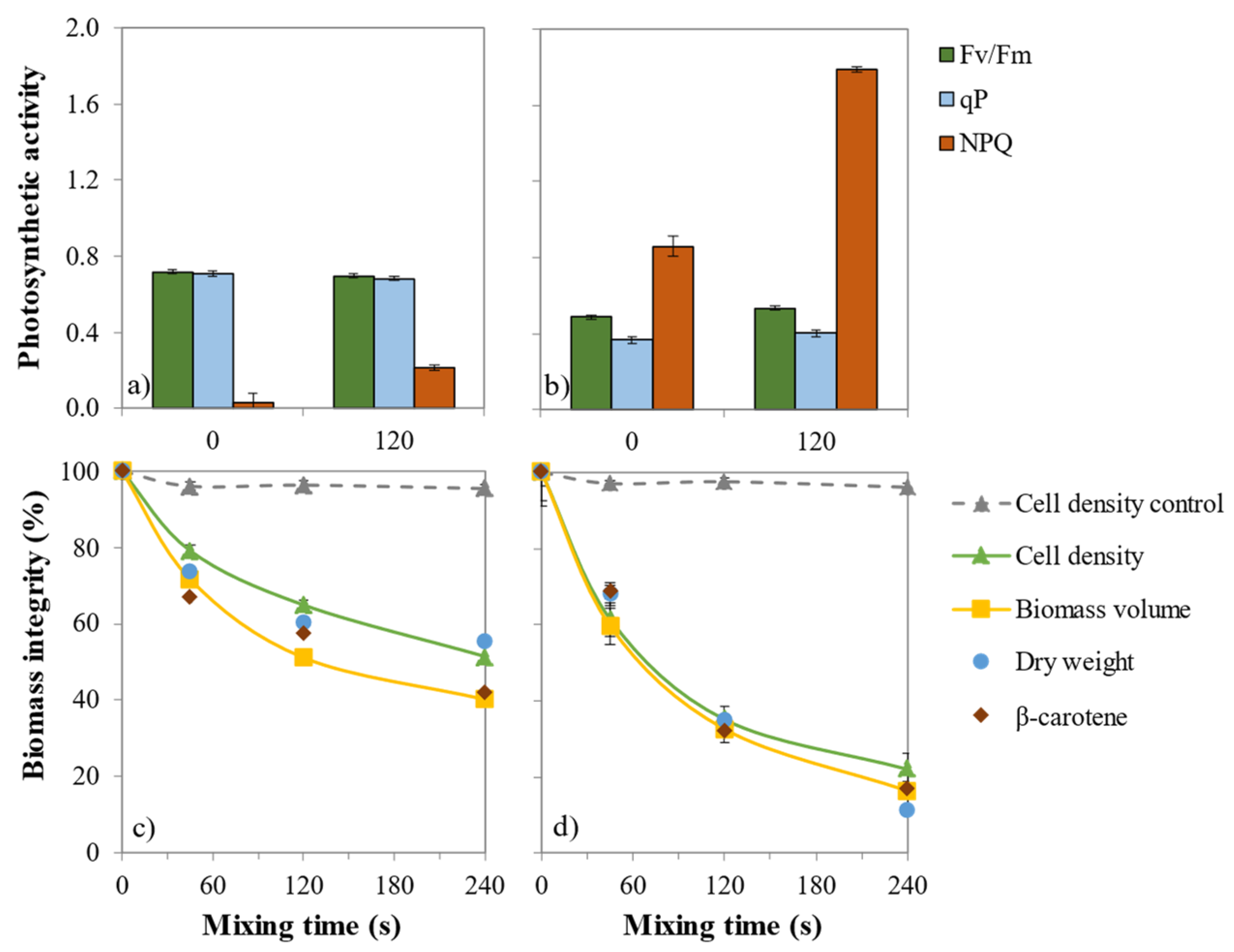
2.2. Impact of β-Carotene Extraction on Photosynthetic Activity
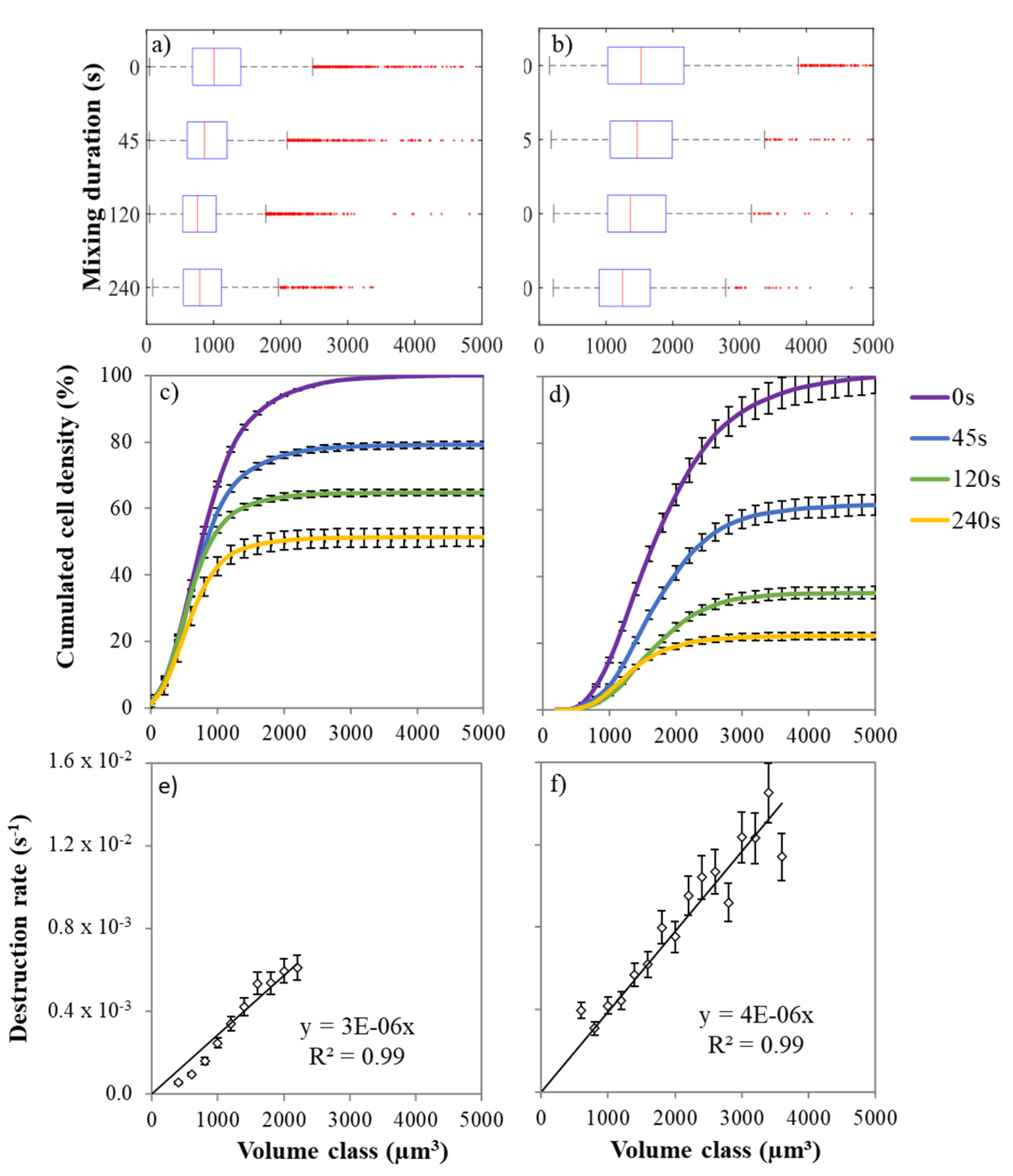
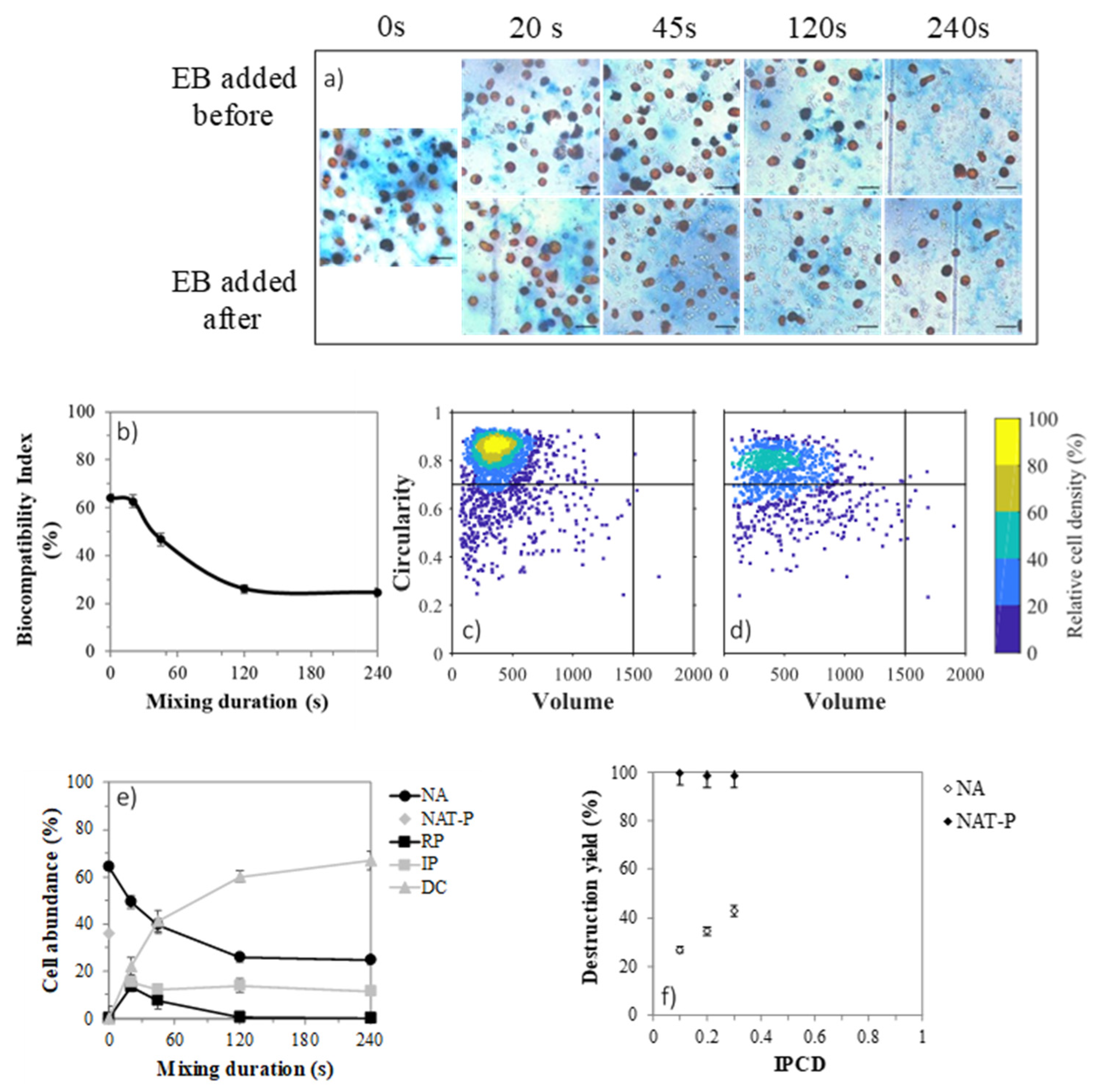
2.3. Impact of β-Carotene Extraction on Cell Integrity
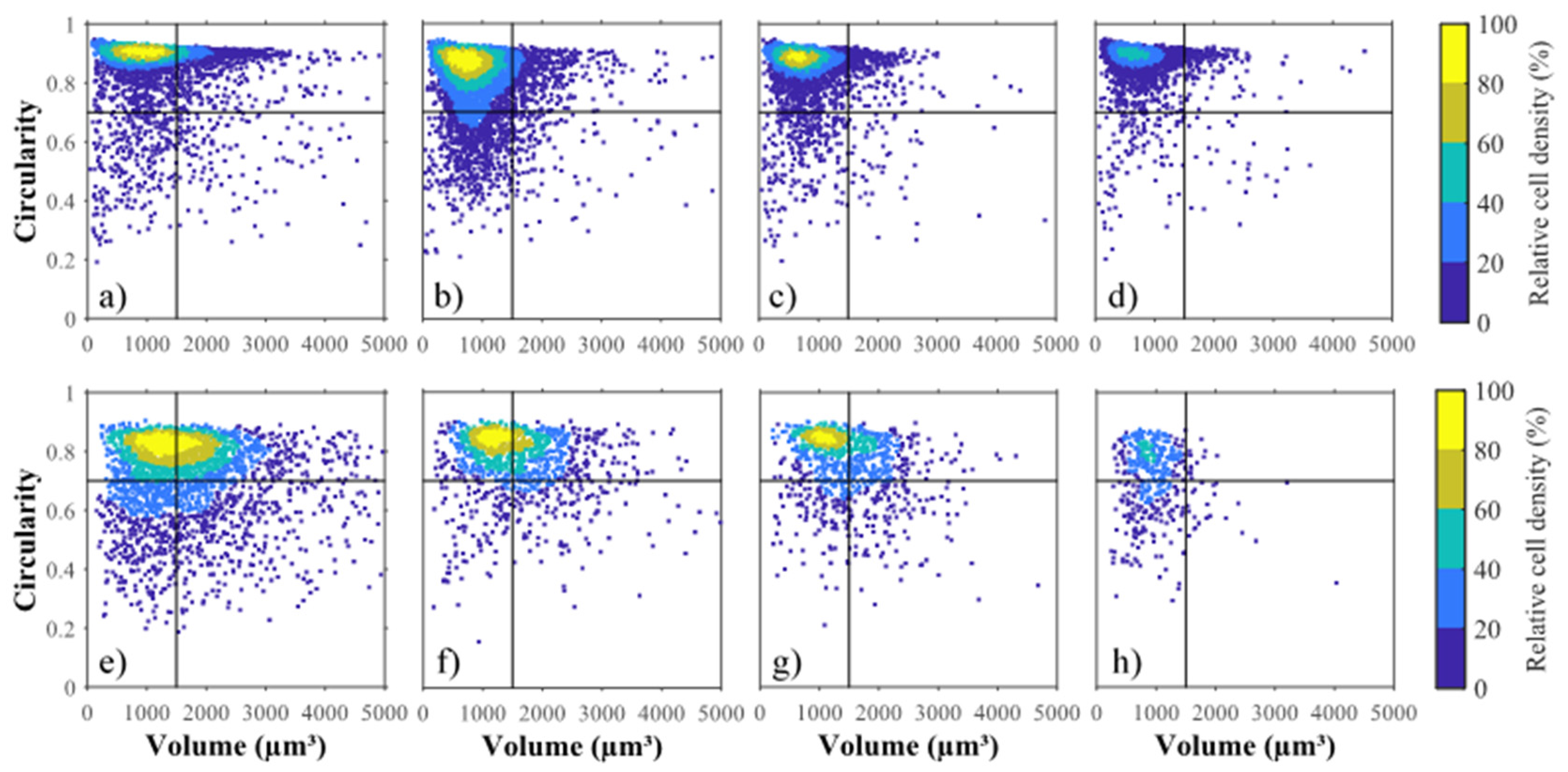
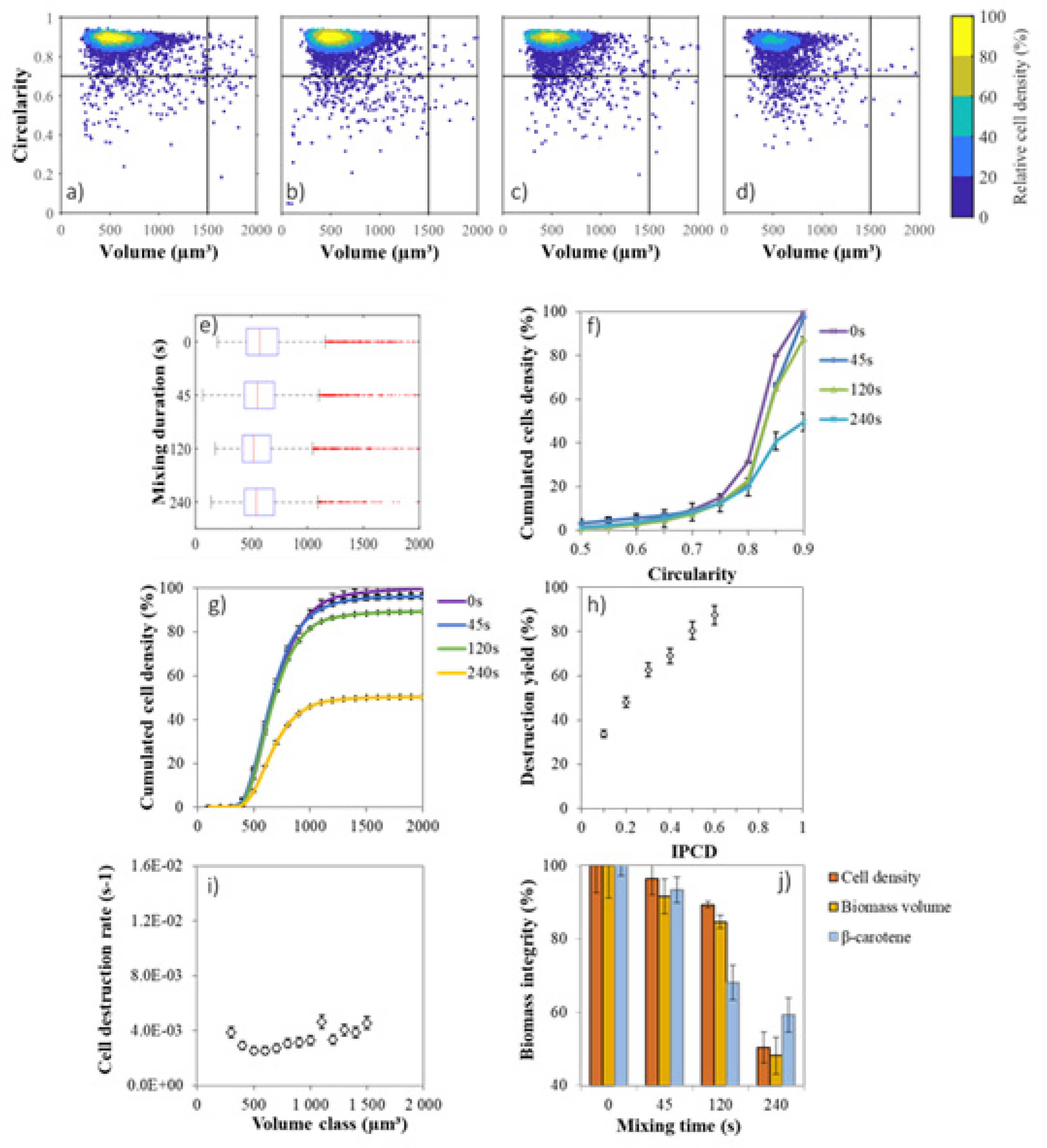
2.4. Cell Size Distribution
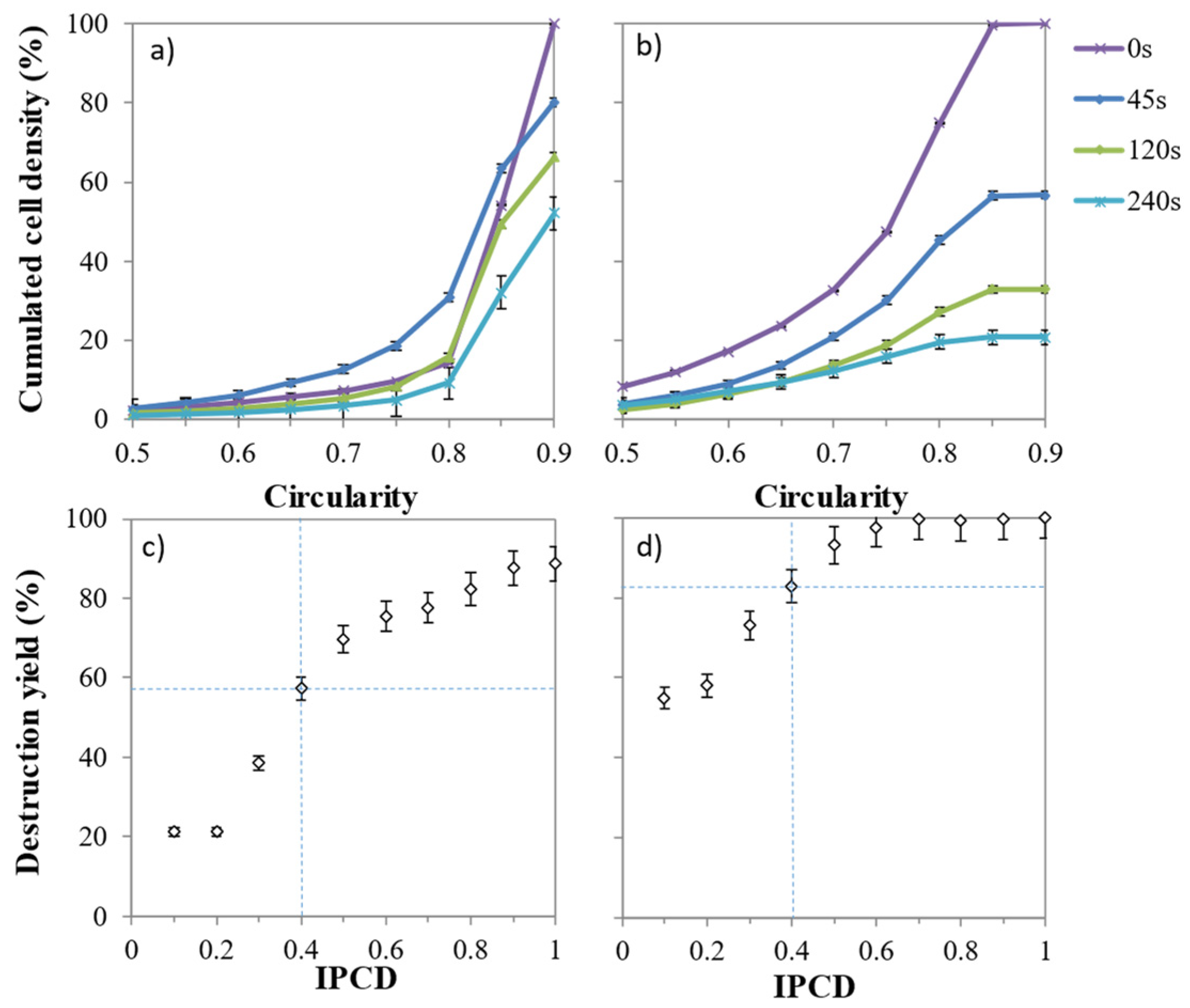
2.5. Influence of Volume and Circularity on Cell Weakness
2.6. Biocompatible Extraction from Photobioreactor Production
2.7. Cell Permeabilization with Solvent
3. Materials and Methods
3.1. Strain and Cultures in Flask
3.2. Culture in Photobioreactor
3.3. In Situ Extraction Experiment
3.4. Characterization of the Aqueous and Solvent Phases
3.4.1. Photosynthetic Activity
3.4.2. β-Carotene Quantification in Biomass
3.4.3. Dry Mass
3.4.4. Cell Density, Volume and Circularity
Cell Density
Circularity
Cell Volume
Index of Potential Disruption
Biomass Integrity, Cell Disruption Yield and Rate
3.4.5. Cell Membrane Permeabilization
3.4.6. Statistics and Plots
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.-P.; Morant-Manceau, A.; Schoefs, B. The Potential of Microalgae for the Production of Bioactive Molecules of Pharmaceutical Interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef] [PubMed]
- Scarsini, M.; Marchand, J.; Schoefs, B. Carotenoid Overproduction in Microalgae: Biochemical and Genetic Engineering. In Pigments from Microalgae Handbook; Springer International Publishing: Cham, Swizterland, 2020; pp. 81–126. [Google Scholar]
- Gateau, H.; Solymosi, K.; Marchand, J.; Schoefs, B. Carotenoids of Microalgae Used in Food Industry and Medicine. Mini-Rev. Med. Chem. 2016, 17, 1140–1172. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Lers, A.; Avron, M. Stereoisomers of β-Carotene and Phytoene in the Alga Dunaliella bardawil. Plant Physiol. 1988, 86, 1286–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinayak, V.; Manoylov, K.M.; Gateau, H.; Blanckaert, V.; Hérault, J.; Pencréac’H, G.; Marchand, J.; Gordon, R.; Schoefs, B. Diatom milking? A review and new approaches. Mar. Drugs 2015, 13, 2629–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini Tafreshi, A.; Shariati, M. Dunaliella biotechnology: Methods and applications. J. Appl. Microbiol. 2009, 107, 14–35. [Google Scholar] [CrossRef]
- Zhang, R.; Marchal, L.; Lebovka, N.; Vorobiev, E.; Grimi, N. Two-step procedure for selective recovery of bio-molecules from microalga Nannochloropsis oculata assisted by high voltage electrical discharges. Bioresour. Technol. 2020, 302, 122893. [Google Scholar] [CrossRef] [PubMed]
- Gateau, H.; Blanckaert, V.; Veidl, B.; Burlet-Schiltz, O.; Pichereaux, C.; Gargaros, A.; Marchand, J.; Schoefs, B. Application of pulsed electric fields for the biocompatible extraction of proteins from the microalga Haematococcus pluvialis. Bioelectrochemistry 2021, 137, 107588. [Google Scholar] [CrossRef] [PubMed]
- Vinayak, V.; Kumar, V.; Kashyap, M.; Joshi, K.B.; Gordon, R.; Schoefs, B. Fabrication of resonating microfluidic chamber for biofuel production in diatoms (Resonating device for biofuel production). In Proceedings of the 2016 3rd International Conference on Emerging Electronics, ICEE 2016, Mumbai, India, 27–30 December 2016; Institute of Electrical and Electronics Engineers Inc.: Piscataway, NJ, USA, 2017. [Google Scholar]
- Frenz, J.; Largeau, C.; Casadevall, E. Hydrocarbon recovery by extraction with a biocompatible solvent from free and immobilized cultures of Botryococcus braunii. Enzyme Microb. Technol. 1989, 11, 717–724. [Google Scholar] [CrossRef]
- Atta, M.; Bukhari, A.; Idris, A. Enhanced lipid selective extraction from Chlorella vulgaris without cell sacrifice. Algal Res. 2016, 20, 7–15. [Google Scholar] [CrossRef]
- León, R.; Martín, M.; Vigara, J.; Vilchez, C.; Vega, J.M. Microalgae mediated photoproduction of β-carotene in aqueous-organic two phase systems. Biomol. Eng. 2003, 20, 177–182. [Google Scholar] [CrossRef]
- Mojaat, M.; Foucault, A.; Pruvost, J.; Legrand, J. Optimal selection of organic solvents for biocompatible extraction of β-carotene from Dunaliella salina. J. Biotechnol. 2008, 133, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, M.A.; De Lamarliere, C.; Rocha, J.M.S.; Vermuë, M.; Tramper, J.; Wijffels, R.H. Selective extraction of carotenoids from the microalga Dunaliella salina with retention of viability. Biotechnol. Bioeng. 2002, 79, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinegris, D.M.M.; van Es, M.A.; Janssen, M.; Brandenburg, W.A.; Wijffels, R.H. Phase toxicity of dodecane on the microalga Dunaliella salina. J. Appl. Phycol. 2011, 23, 949–958. [Google Scholar] [CrossRef] [Green Version]
- Hejazi, M.A.; Andrysiewicz, E.; Tramper, J.; Wijffels, R.H. Effect of Mixing Rate on β-Carotene Production and Extraction by Dunaliella Salina in Two-Phase Bioreactors. Biotechnol. Bioeng. 2003, 84, 591–596. [Google Scholar] [CrossRef]
- Miazek, K.; Kratky, L.; Sulc, R.; Jirout, T.; Aguedo, M.; Richel, A.; Goffin, D. Effect of organic solvents on microalgae growth, metabolism and industrial bioproduct extraction: A review. Int. J. Mol. Sci. 2017, 18, 1429. [Google Scholar] [CrossRef] [Green Version]
- Dodge, J.D. The Fine Structure of Algal Cells; Elsevier: Amsterdam, The Netherlands, 1973. [Google Scholar]
- León, R.; Garbayo, I.; Hernández, R.; Vigara, J.; Vilchez, C. Organic solvent toxicity in photoautotrophic unicellular microorganisms. Enzyme Microb. Technol. 2001, 29, 173–180. [Google Scholar] [CrossRef]
- Bar, R. Effect of interphase mixing on a water–organic solvent two-liquid phase microbial system: Ethanol fermentation. J. Chem. Technol. Biotechnol. 1988, 43, 49–62. [Google Scholar] [CrossRef]
- Sikkema, J.; De Bont, J.A.M.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Lorente De Nó, R. Studies on the structure of the cerebral cortex. II. Continuation of the study of the ammonic system. J. Psychol. Neurol. 1934, 46, 113–117. [Google Scholar]
- Ben-Amotz, A.; Avron, M. On the Factors Which Determine Massive β-Carotene Accumulation in the Halotolerant Alga Dunaliella bardawi. Plant Physiol. 1983, 72, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Melis, A. Biosynthesis and Distribution of Chlorophyll among the Photosystems during Recovery of the Green Alga. Society 2002, 128, 603–614. [Google Scholar] [CrossRef]
- Gray, G.R.; Huner, N.P.A. Excitation Pressure Determines Tolerance to Photoinhibition of Photosynthesis. In Photosynthesis: From Light to Biosphere; Springer: Dordrecht, The Netherlands, 1995; pp. 3335–3338. [Google Scholar]
- Kleinegris, D.M.M.; Janssen, M.; Brandenburg, W.A.; Wijffels, R.H. Continuous production of carotenoids from Dunaliella salina. Enzyme Microb. Technol. 2011, 48, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojaat Guemir, M. Production par Dunaliella salina et extraction en continu de β-carotène par couplage d’un photobioréacteur et d’une chromatographie de partage centrifuge. Ph.D. Thesis, University of Wageningen, Wageningen, The Netherlands, 2008. [Google Scholar]
- Halter, M.; Elliott, J.T.; Hubbard, J.B.; Tona, A.; Plant, A.L. Cell volume distributions reveal cell growth rates and division times. J. Theor. Biol. 2009, 257, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, M.A.; Kleinegris, D.; Wijffels, R.H. Mechanism of extraction of β-carotene from microalga Dunaliellea salina in two-phase bioreactors. Biotechnol. Bioeng. 2004, 88, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, Y.; Zhang, X.; Tan, T. Lipid extraction from non-broken and high water content microalgae Chlorella spp. by three-phase partitioning. Algal Res. 2015, 10, 218–223. [Google Scholar] [CrossRef]
- Hadj-Romdhane, F.; Jaouen, P.; Pruvost, J.; Grizeau, D.; Van Vooren, G.; Bourseau, P. Development and validation of a minimal growth medium for recycling Chlorella vulgaris culture. Bioresour. Technol. 2012, 123, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Çelekli, A.; Dönmez, G. Effect of pH, light intensity, salt and nitrogen concentrations on growth and β-carotene accumulation by a new isolate of Dunaliella sp. World J. Microbiol. Biotechnol. 2006, 22, 183–189. [Google Scholar] [CrossRef]
- Pruvost, J.; Cornet, J.-F.; Pilon, L. Large-Scale Production of Algal Biomass: Photobioreactors. Algae Biotechnol. 2016, 10, 41–66. [Google Scholar] [CrossRef]
- Xu, F.; Yuan, Q.P.; Zhu, Y. Improved production of lycopene and β-carotene by Blakeslea trispora with oxygen-vectors. Process Biochem. 2007, 42, 289–293. [Google Scholar] [CrossRef]
- Roháček, K.; Soukupová, J.; Barták, M. Chlorophyll Fluorescence: A Wonderful Tool to Study Plant Physiology and Plant Stress; Research Signpost: Kerala, India, 2008; Volume 661, ISBN 9788130801049. [Google Scholar]
- Roháček, K.; Bertrand, M.; Moreau, B.; Jacquette, B.; Caplat, C.; Morant-Manceau, A.; Schoefs, B. Relaxation of the non-photochemical chlorophyll fluorescence quenching in diatoms: Kinetics, components and mechanisms. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichthentaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Mojaat, M.; Pruvost, J.; Foucault, A.; Legrand, J. Effect of organic carbon sources and Fe2+ ions on growth and β-carotene accumulation by Dunaliella salina. Biochem. Eng. J. 2008, 39, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.J.; Lee, Y.K. Determination of biomass dry weight of marine microalgae. J. Appl. Phycol. 1997, 9, 189–194. [Google Scholar] [CrossRef]
- Hyka, P.; Lickova, S.; Přibyl, P.; Melzoch, K.; Kovar, K. Flow cytometry for the development of biotechnological processes with microalgae. Biotechnol. Adv. 2013, 31, 2–16. [Google Scholar] [CrossRef] [Green Version]
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The inverted microscope method of estimating algal numbers and the statistical basis of estimations by counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- De Carvalho, C.C.C.R.; Da Fonseca, M.M.R. Solvent toxicity in organic-aqueous systems analysed by multivariate analysis. Bioprocess Biosyst. Eng. 2004, 26, 361–375. [Google Scholar] [CrossRef]
- Syed, M.S.; Rafeie, M.; Vandamme, D.; Asadnia, M.; Henderson, R.; Taylor, R.A.; Warkiani, M.E. Selective separation of microalgae cells using inertial microfluidics. Bioresour. Technol. 2018, 252, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Liu, D. Geometric models for calculating cell biovolume and surface area for phytoplankton. J. Plankton Res. 2003, 25, 1331–1346. [Google Scholar] [CrossRef] [Green Version]
- Hamer, P.W.; McGeachie, J.M.; Davies, M.J.; Grounds, M.D. Evans Blue Dye as an in vivo marker of myofibre damage: Optimising parameters for detecting initial myofibre membrane permeability. J. Anat. 2002, 200, 69–79. [Google Scholar] [CrossRef]
- Luengo, E.; Condón-Abanto, S.; Álvarez, I.; Raso, J. Effect of Pulsed Electric Field Treatments on Permeabilization and Extraction of Pigments from Chlorella vulgaris. J. Membr. Biol. 2014, 247, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Eilers, P.H.C.; Goeman, J.J. Enhancing scatterplots with smoothed densities. Bioinformatics 2004, 20, 623–628. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Category | Abbreviation | EB Alone (A) | EB before Solvent (B) | EB after Solvent (C) |
|---|---|---|---|---|
| Non-affected | NA | − | − | − |
| Reversibly permeabilized | RP | − | + | − |
| Irreversibly permeabilized | IP | − | + | + |
| Naturally permeabilized | NAT-P | + | + | + |
| Disrupted cells | DC | / | / | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanguy, G.; Legat, A.; Gonçalves, O.; Marchal, L.; Schoefs, B. Selection of Culture Conditions and Cell Morphology for Biocompatible Extraction of β-Carotene from Dunaliella salina. Mar. Drugs 2021, 19, 648. https://doi.org/10.3390/md19110648
Tanguy G, Legat A, Gonçalves O, Marchal L, Schoefs B. Selection of Culture Conditions and Cell Morphology for Biocompatible Extraction of β-Carotene from Dunaliella salina. Marine Drugs. 2021; 19(11):648. https://doi.org/10.3390/md19110648
Chicago/Turabian StyleTanguy, Guillaume, Aline Legat, Olivier Gonçalves, Luc Marchal, and Benoît Schoefs. 2021. "Selection of Culture Conditions and Cell Morphology for Biocompatible Extraction of β-Carotene from Dunaliella salina" Marine Drugs 19, no. 11: 648. https://doi.org/10.3390/md19110648
APA StyleTanguy, G., Legat, A., Gonçalves, O., Marchal, L., & Schoefs, B. (2021). Selection of Culture Conditions and Cell Morphology for Biocompatible Extraction of β-Carotene from Dunaliella salina. Marine Drugs, 19(11), 648. https://doi.org/10.3390/md19110648