
Recent Advances in Small Peptides of Marine Origin in Cancer Therapy

 ,
,  and
and
Abstract
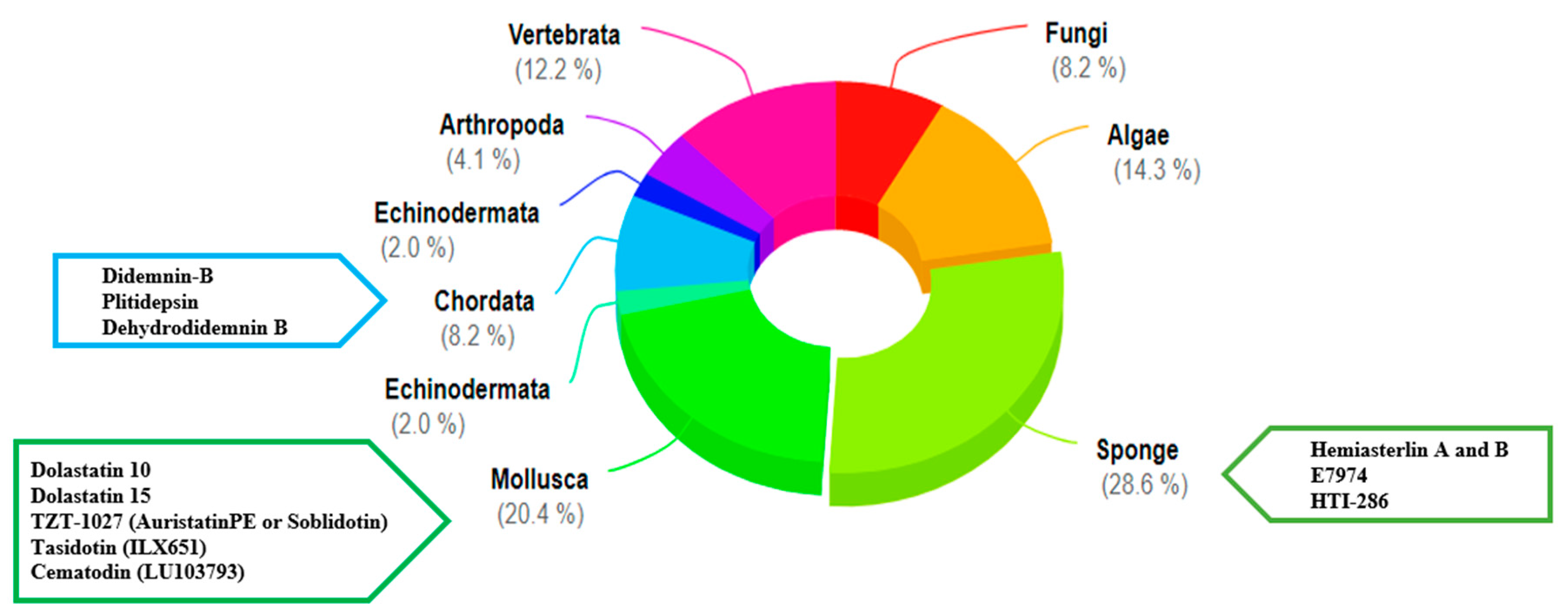
:1. Introduction
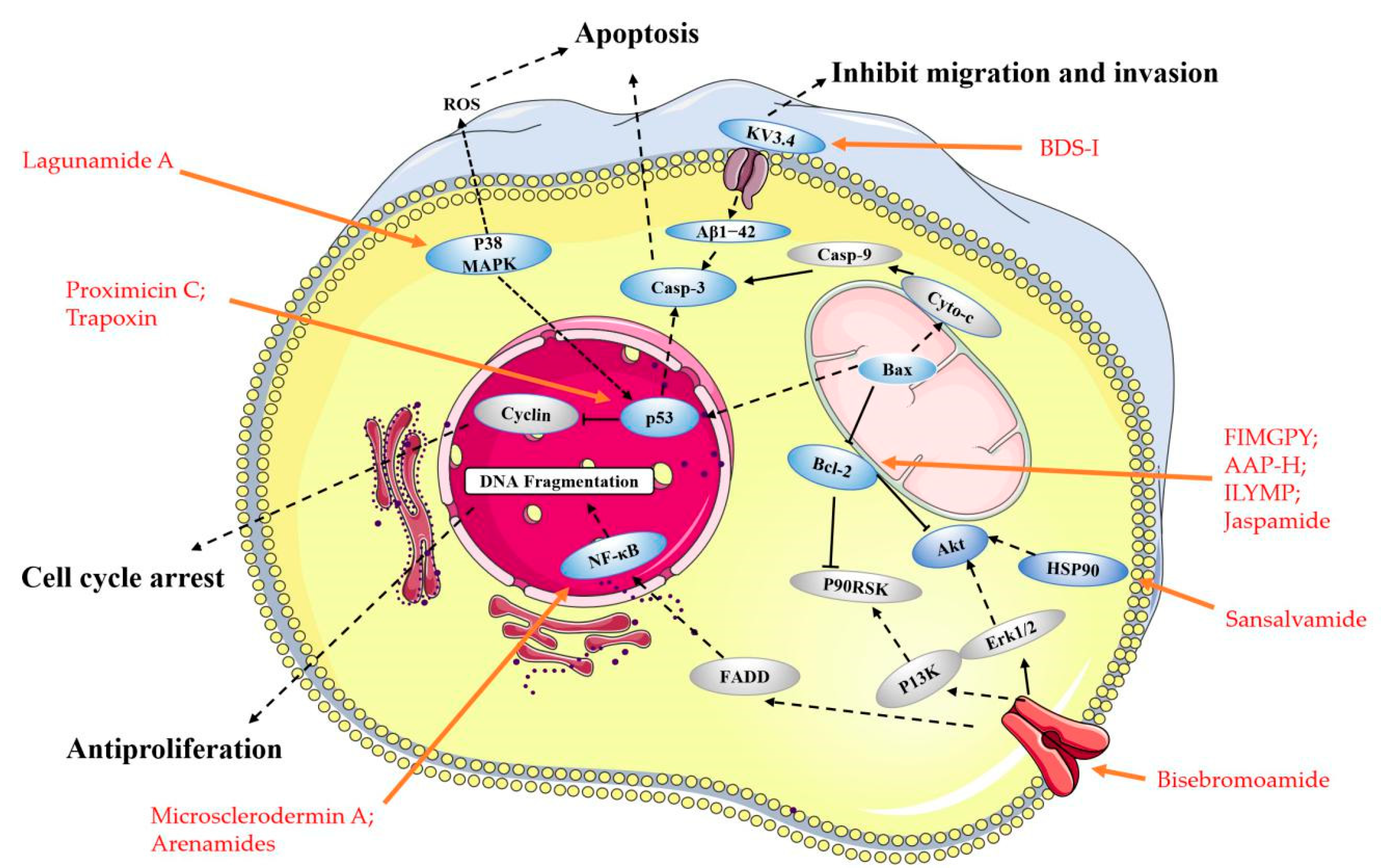
2. Linear Peptides and Derivatives
2.1. Animals
2.2. Fungi and Bacteria
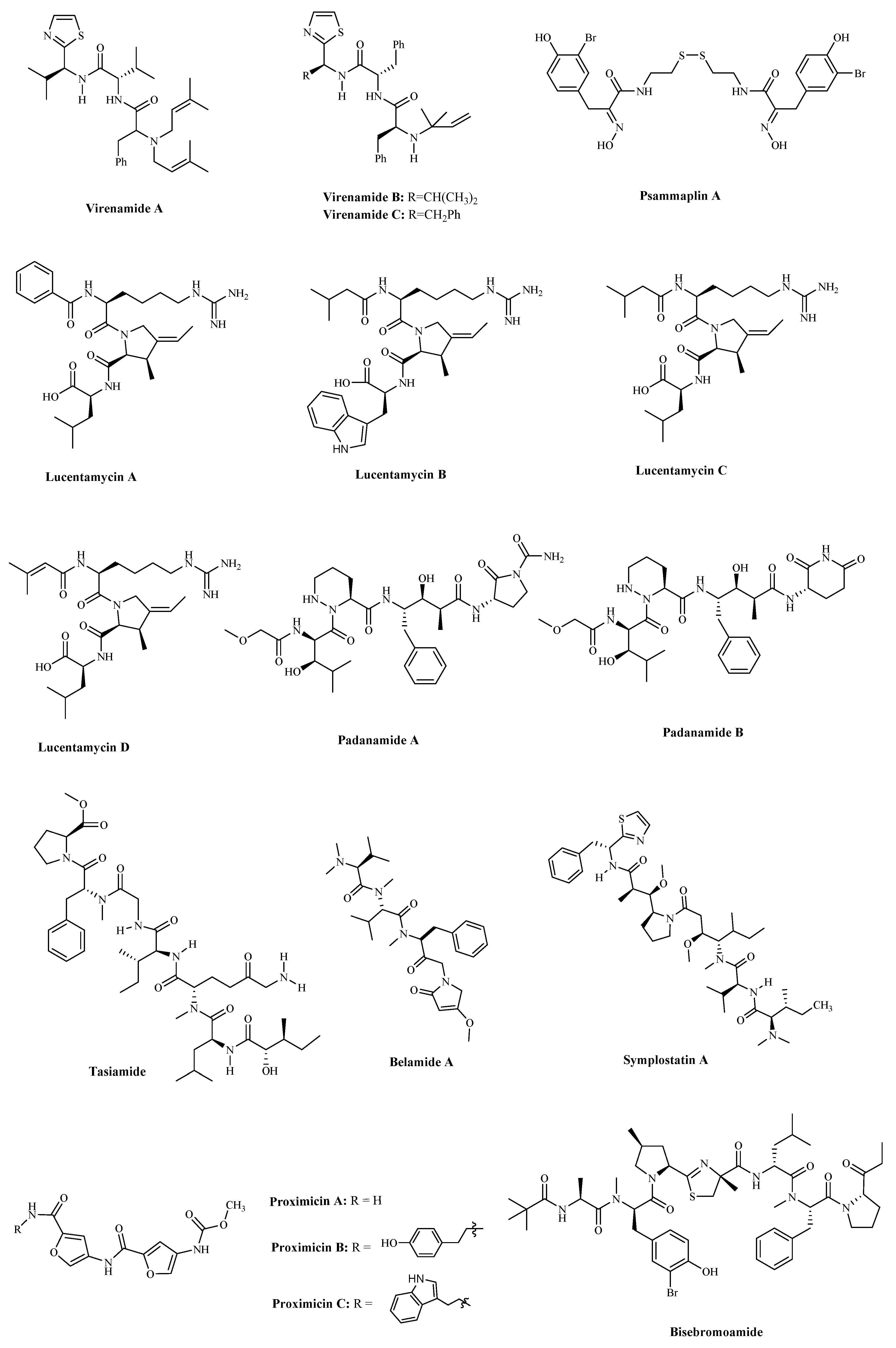
2.3. Other Small Peptides
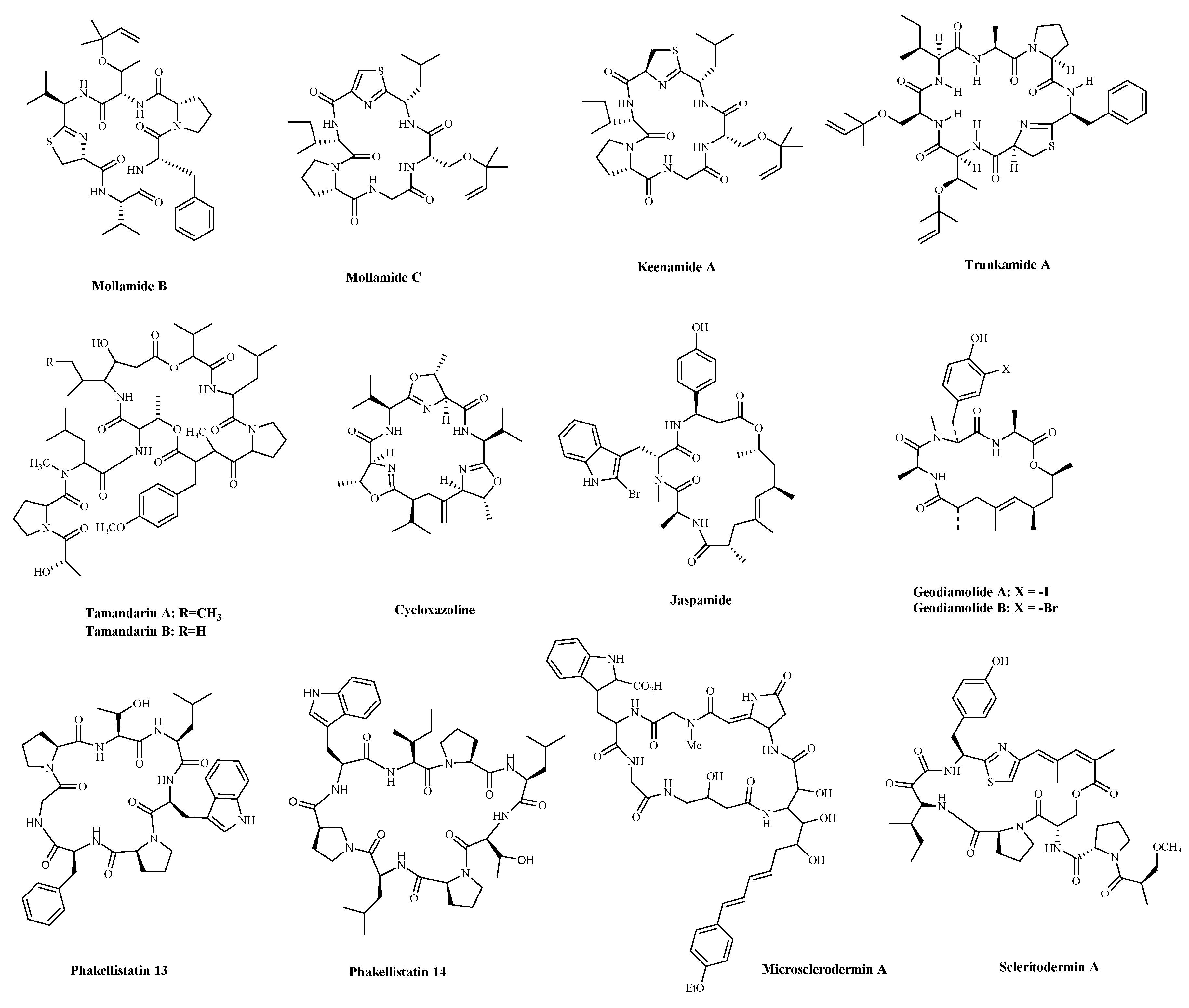
3. Cyclic Peptides and Derivatives
3.1. Animals
3.1.1. Metabolites of Ascidians
3.1.2. Metabolites of Sponges
3.2. Fungi
3.3. Bacteria
Cyanobacterial Metabolites
4. Marine-Derived Small Peptides in Clinical Trials
4.1. Linear Peptides
4.2. Cyclic Peptides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| ADCs | Antibody–drug conjugates |
| A549 | Human non-small cell lung cancer cells |
| AGS | Human gastric cancer adenocytes cells |
| APD3 | Database of antimicrobial peptides |
| A-10 | Rat aortic smooth muscle cells |
| AsPC-1 | Human metastatic pancreatic cancer cells |
| AVPdb | Database of antiviral peptides |
| BACE1 | Beta-secretase 1 |
| BEL-7404 | Human liver cancer cells |
| BxPC-3 | Human in situ adenocarcinoma cells |
| CAR-T | Chimeric Antigen Receptor T-Cell Immunotherapy |
| CancerPPD | Database of anticancer peptides and proteins |
| Cath D | Cathepsin D |
| Cath E | Cathepsin E |
| CDX-011 | Glembatumumab vedotin |
| CV1 | African green monkey kidney cells |
| DLD-1 | Human colonic adenocarcinoma cells |
| DU-145 | Human prostate cancer cells |
| FDA | United States Food and Drug Administration |
| 5-FU | 5-Fluorouracil |
| GI50 | Half-maximal growth inhibitory concentration |
| GPNMB | Glycoprotein NMB |
| HCT 8 | Human cecal adenocarcinoma cell |
| HCT-116 | Human colon cancer cells |
| HDAC | Histone Deacetylase |
| HeLa | Human cervical cancer cells |
| Hep G2 | Human liver cancer cells |
| H460 | Human non-small cell lung cancer cells |
| H-1299 | Human non-small cell lung cancer cells |
| HSP90A | Heat Shock Protein 90 Alpha Family Class A |
| HT-29 | Human colon cancer cells |
| IC50 | Half-maximal inhibitory concentration |
| Jurkat 16 | T-cell leukemia cells |
| JFCR39 | A panel of 39 human cancer cell lines |
| KB | Human nasopharyngeal cancer cells |
| KDC | Knottin peptide–drug conjugate |
| LNCaP | Human prostate cancer cells |
| LO2 | Normal liver cells |
| L1236 | Hodgkin’s Lymphoma cells |
| LoVo | Human colon cancer cells |
| MCF-7 | Human breast cancer cells |
| MDA-MB-231 | Human breast cancer cells |
| MDR | Multidrug resistance |
| MDs | Molecular dynamics simulation |
| MMAE | Monomethyl auristatin E |
| MOLT4 | Human acute T lymphoblastic leukaemia cells |
| NF-κB | Nuclear factor kappa B |
| PANC-1 | Human pancreatic cancer cells |
| PC-3 | Human prostate cancer cells |
| P388 | Mouse leukemia cells |
| PsA | Psammaplin A |
| RBC | Red blood cell |
| ROS | Overproduction of reactive oxygen species |
| RXF 393 | Human kidney cancer cells |
| SGC-7901 | Human gastric cancer cells |
| SK-OV3 | Human ovarian cancer cell |
| SF-268 | Human neurocancer cells |
| TNF | Tumor necrosis factor |
| 293/NFκB- | Stably transfected NFκB human embryonic kidney cells |
| U-87 MG | Human glioblastoma cells |
| U937 | Human histiocytic lymphoma cells |
| WBC | White blood cell |
References
- Molinski, T. Marine natural products. Clin. Adv. Hematol. Oncol. 2009, 7, 383–385. [Google Scholar]
- de Vries, D.J.; Beart, P.M. Fishing for drugs from the sea: Status and strategies. Trends Pharmacol. Sci. 1995, 16, 275–279. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [Green Version]
- Dyshlovoy, S.A.; Honecker, F. Marine compounds and cancer: 2017 updates. Mar. Drugs 2018, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Nazish, S.; Mohamed, A. Personalized medicine in cardio-oncology: The role of induced pluripotent stem cell. Cardiovasc. Res. 2019, 115, 949–959. [Google Scholar] [CrossRef]
- Vora, C.; Gupta, S. Targeted therapy in cervical cancer. ESMO Open 2019, 3, e000462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, K.A.; Kalafatis, M. The case back on the TRAIL: Death receptors as markers for rhTRAIL sensitivity. J. Appl. Lab. Med. 2017, 2, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Salazar, R.; Cortés-Funes, H.; Casado, E.; Pardo, B.; López-Martín, A.; Cuadra, C.; Tabernero, J.; Coronado, C.; García, M.; Soto Matos-Pita, A.; et al. Phase I study of weekly kahalalide F as prolonged infusion in patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2013, 72, 75–83. [Google Scholar] [CrossRef]
- Jiao, S.; Wang, H.; Shi, Z.; Dong, A.; Zhang, W.; Song, X.; He, F.; Wang, Y.; Zhang, Z.; Wang, W.; et al. A peptide mimicking VGLL4 function acts as a YAP antagonist therapy against gastric cancer. Cancer Cell 2014, 25, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Cox, N.; Kintzing, J.R.; Smith, M.; Grant, G.A.; Cochran, J.R. Integrin-targeting knottin peptide-drug conjugates are potent inhibitors of tumor cell proliferation. Angew. Chem. Int. Ed. Engl. 2016, 55, 9894–9897. [Google Scholar] [CrossRef] [PubMed]
- Bjork, R.L. Bi-Specific Monoclonal Antibody (Specific for Both CD3 and CD11b) Therapeutic Drug. U.S. Patent WO2007US14524, 22 June 2007. [Google Scholar]
- Isazadeh, A.; Hajazimian, S.; Garshasbi, H.; Shadman, B.; Baradaran, B. Resistance mechanisms to immune checkpoints blockade by monoclonal antibody drugs in cancer immunotherapy: Focus on myeloma. J. Cell. Physiol. 2020, 236, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Shahidian, A.; Ghassemi, M.; Mohammadi, J.; Hashemi, M. Immunotherapy. In Bio-Engineering Approaches to Cancer Diagnosis and Treatment; Academic Press: Cambridge, MA, USA, 2020; pp. 69–114. [Google Scholar]
- Lian, Q.; Cheng, Y.; Zhong, C.; Wang, F. Inhibition of the IgE-mediated activation of RBL-2H3 cells by TIPP, a novel thymic immunosuppressive pentapeptide. Int. J. Mol. Sci. 2015, 16, 2252–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, S.; Vhora, I.; Amrutiya, J.; Lalani, R.; Misra, A. Role of nanotechnology in delivery of protein and peptide drugs. Curr. Pharm. Des. 2015, 21, 4155–4173. [Google Scholar] [CrossRef]
- Tatiana, R.; Andreas, B.; Yves, S.; Christopher, K.; Alison, B.; Fabien, F.; Luca, M.; Ismael, Z.; Andrea, M. Software-aided approach to investigate peptide structure and metabolic susceptibility of amide bonds in peptide drugs based on high resolution mass spectrometry. PLoS ONE 2017, 12, e0186461. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Seebah, S.; Suresh, A.; Zhuo, S.; Choong, Y.H.; Chua, H.; Chuon, D.; Beuerman, R.; Verma, C. Defensins knowledgebase: A manually curated database and information source focused on the defensins family of antimicrobial peptides. Nucleic Acids Res. 2007, 35, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.; Thakur, N.; Tandon, H.; Kumar, M. AVPdb: A database of experimentally validated antiviral peptides targeting medically important viruses. Nucleic Acids Res. 2014, 42, D1147–D1153. [Google Scholar] [CrossRef] [Green Version]
- Divya, M.; Priya, A.; Vineet, K.; Anshika, J.; Deepika, M.; Sandeep, S.; Abhishek, T.; Kumardeep, C.; Gautam, S.K.; Ankur, G. ParaPep: A web resource for experimentally validated antiparasitic peptide sequences and their structures. Database (Oxf.) 2014. [Google Scholar] [CrossRef]
- Atul, T.; Abhishek, T.; Priya, A.; Sudheer, G.; Minakshi, S.; Deepika, M.; Anshika, J.; Sandeep, S.; Ankur, G.; Raghava, G.P.S. CancerPPD: A database of anticancer peptides and proteins. Nucleic Acids Res. 2015, 43, 837–843. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.J.; Wang, S.; Kang, C.Z.; Lv, C.G.; Zhou, L.; Huang, L.Q.; Guo, L.P. Pharmacodynamic material basis of traditional chinese medicine based on biomacromolecules: A review. Plant Methods 2020, 16, 26. [Google Scholar] [CrossRef] [Green Version]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Míguez, A.; Gutiérrez-Jácome, A.; Pérez-Pérez, M.; Pérez-Rodríguez, G.; Catalán-García, S.; Fdez-Riverola, F.; Lourenço, A.; Sánchez, B. From amino acid sequence to bioactivity: The biomedical potential of antitumor peptides. Protein Sci. 2016, 25, 1084–1095. [Google Scholar] [CrossRef]
- Nasrolahi Shirazi, A.; Tiwari, R.; Chhikara, B.S.; Mandal, D.; Parang, K. Design and biological evaluation of cell-penetrating peptide-doxorubicin conjugates as prodrugs. Mol. Pharm. 2013, 10, 488–499. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Mani-Varnosfaderani, A.; Khayamian, T.; Gharaghani, S. An in silico approach to design peptide mimetics based on docking and molecular dynamics simulation of EGFR-matuzumab complex. J. Iran Chem. Soc. 2016, 13, 1805–1817. [Google Scholar] [CrossRef]
- Gellerman, G.; Baskin, S.; Galia, L.; Gilad, Y.; Firer, M.A. Drug resistance to chlorambucil in murine B-cell leukemic cells is overcome by its conjugation to a targeting peptide. Anticancer Drugs 2013, 24, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine natural products: A source of novel anticancer drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A.R.; Mohanram, M.S.G.; Balasubramanian, B.; Kim, I.H.; Seedevi, P.; Mohan, K.; Kanagasabai, S.; Arasu, M.V.; Al-Dhabi, N.A.; Ignacimuthu, S. Marine invertebrates’ proteins: A recent update on functional property. J. King Saud Univ. Sci. 2020, 32, 1496–1502. [Google Scholar] [CrossRef]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, A.; Balde, A.; Raghavender, P.; Shashanth, D.; Abraham, J.; Joshi, I.; Nazeer, R.A. Isolation of marine crab (Charybdis natator) leg muscle peptide and its anti-inflammatory effects on macrophage cells. Biocatal. Agric. Biotechnol. 2020, 25, 101577. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, G.J.; Liu, W.D.; Yang, X.Y.; Zhu, N.; Shen, J.M.; Wang, Z.C.; Liu, Y.; Cheng, S.; Yu, G.L.; et al. Recent progress in research and development of marine drugs. Chin. J. Mar. Drugs 2019, 38, 35–69. [Google Scholar]
- Kim, S.M. Antioxidant and anticancer activities of enzymatic hydrolysates of solitary tunicate (Styela clava). Food Sci. Biotechnol. 2011, 20, 1075–1085. [Google Scholar] [CrossRef]
- Shakeel, E.; Arora, D.; Jamal, Q.M.S.; Akhtar, S.; Khan, M.K.A.; Kamal, M.A.; Siddiqui, M.H.; Lohani, M.; Arif, J.M. Marine drugs: A hidden wealth and a new epoch for cancer management. Curr. Drug Metab. 2018, 19, 523–543. [Google Scholar] [CrossRef]
- Dyshlovoy, S.; Honecker, F. Marine compounds and cancer: Updates 2020. Mar. Drugs 2020, 18, 643. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.; Honecker, F. Marine compounds and cancer: The first two decades of XXI century. Mar. Drugs 2019, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, N.; Dhanalakshmi, S.; Pandian, P. Encouraging leads from marine sources for cancer therapy—A review approach. Pharmacogn. J. 2020, 12, 1475–1481. [Google Scholar] [CrossRef]
- Ming-Jia, Z.; Dong-Mei, Z.; Jun-Hui, C. Research advances of antitumor peptides. Chin. J. Biochem. Pharm. 2007, 28, 139–141. [Google Scholar] [CrossRef]
- Ding, G.-F.; Huang, F.-F.; Yang, Z.-S.; Yu, D.; Yang, Y.-F. Anticancer activity of an oligopeptide isolated from hydrolysates of Sepia Ink. Chin. J. Nat. Med. 2011, 9, 151–155. [Google Scholar] [CrossRef]
- Quah, Y. Purification and identification of novel cytotoxic oligopeptides from soft coral Sarcophyton glaucum. J. Zhejiang Univ. Sci. B Biomed. Biotechnol. 2019, 20, 59–70. [Google Scholar] [CrossRef]
- Gan, H. Concise and efficient total syntheses of virenamides A and D. J. Adv. Chem. 2008, 4, 488–493. [Google Scholar] [CrossRef]
- Umayaparvathi, S.; Arumugam, M.; Meenakshi, S.; Dräger, G.; Kirschning, A.; Balasubramanian, T. Purification and characterization of antioxidant peptides from oyster (Saccostrea cucullata) hydrolysate and the anticancer activity of hydrolysate on human colon cancer cell lines. Int. J. Pept. Res. Ther. 2014, 20, 231–243. [Google Scholar] [CrossRef]
- Umayaparvathi, S.; Meenakshi, S.; Vimalraj, V.; Arumugam, M.; Sivagami, G.; Balasubramanian, T. Antioxidant activity and anticancer effect of bioactive peptide from enzymatic hydrolysate of oyster (Saccostrea cucullata). Biomed. Prev. Nutr. 2014, 4, 343–353. [Google Scholar] [CrossRef]
- Song, R.; Wei, R.B.; Luo, H.Y.; Yang, Z.S. Isolation and identification of an antiproliferative peptide derived from heated products of peptic hydrolysates of half-fin anchovy (Setipinna taty). J. Funct. Foods 2014, 10, 104–111. [Google Scholar] [CrossRef]
- Ratih, P.; Se-Kwon, K. Bioactive peptide of marine origin for the prevention and treatment of non-communicable diseases. Mar. Drugs 2017, 15, 67. [Google Scholar] [CrossRef]
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Li, T.; Ding, G.-F. Antioxidant and anticancer peptides from the protein hydrolysate of blood clam (Tegillarca granosa) muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Aissaoui, D.; Mlayah Bellalouna, S.; Jebali, J.; Abdelkafi Koubaa, Z.; Souid, S.; Moslah, W.; Othman, H.; Luis, J.; Elayeb, M.; Marrakchi, N. Functional role of Kv1.1 and Kv1.3 channels in the neoplastic progression steps of three cancer cell lines, elucidated by scorpion peptides. Int. J. Biol. Macromol. 2018, 111, 1146–1155. [Google Scholar] [CrossRef]
- Ciccone, R.; Piccialli, I.; Grieco, P.; Merlino, F.; Annunziato, L.; Pannaccione, A. Synthesis and pharmacological evaluation of a novel peptide based on anemonia sulcata BDS-I toxin as a new KV3.4 inhibitor exerting a neuroprotective effect against amyloid-β peptide. Front. Chem. 2019, 9, 497. [Google Scholar] [CrossRef]
- Min, S.; Su, P.; Jeong, P.; Jin, B.; Hee, J.; Seung, S.; Pan, R.; So, L. Kv3.1 and Kv3.4, are involved in cancer cell migration and invasion. Int. J. Mol. Sci. 2018, 19, 1061. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Zhao, Y.Q.; Hu, F.Y.; Chi, C.F.; Wang, B. Anticancer activity of a hexapeptide from skate (Raja porosa) cartilage protein hydrolysate in HeLa cells. Mar. Drugs 2016, 14, 153. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Z.; Ding, G.F.; Huang, F.F.; Yang, Z.S.; Yu, F.M.; Tang, Y.P.; Jia, Y.L.; Zheng, Y.Y.; Chen, R. Anticancer activity of anthopleura anjunae oligopeptides in prostate cancer DU-145 cells. Mar. Drugs 2018, 16, 125. [Google Scholar] [CrossRef] [Green Version]
- Fangmiao, Y.; Yaru, Z.; Lei, Y.; Yunping, T.; Guofang, D.; Xiaojun, Z.; Zuisu, Y. A novel antiproliferative pentapeptide (ILYMP) isolated from cyclinasinensis protein hydrolysate induces apoptosis of DU145 prostate cancer cells. Mol. Med. Rep. 2018, 18, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Ding, G.; Yang, Z.; Yu, F. Two novel peptides derived from Sinonovacula constricta inhibit the proliferation and induce apoptosis of human prostate cancer cells. Mol. Med. Rep. 2017, 16, 6697–6707. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Sun, L.; Zhou, G.; Xie, P.; Ye, J. Sepia ink oligopeptide induces apoptosis and growth inhibition in human lung cancer cells. Oncotarget 2017, 8, 23202–23212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia ink oligopeptide induces apoptosis in prostate cancer cell lines via caspase-3 activation and elevation of Bax/Bcl-2 ratio. Mar Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Jing, Y.; Ding, G.; Yang, Z. Isolation and purification of novel peptides derived from Sepia ink: Effects on apoptosis of prostate cancer cell PC-3. Mol. Med. Rep. 2017, 16, 4222–4228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, M.G.J.; Leiser, T.; Haus, P.; Samlal, S.; Wong, A.C.; Wood, R.J.; Petrucci, V.; Gunaratnam, M.; Hughes, S.M.; Buluwela, L. Defining the mechanism of action and enzymatic selectivity of Psammaplin A against its epigenetic targets. J. Med. Chem. 2012, 55, 1731–1750. [Google Scholar] [CrossRef] [PubMed]
- Kumar M, S.L.; Ali, K.; Chaturvedi, P.; Meena, S.; Datta, D.; Panda, G. Design, synthesis and biological evaluation of oxime lacking Psammaplin inspired chemical libraries as anti-cancer agents. J. Mol. Struct. 2021, 1225. [Google Scholar] [CrossRef]
- Simmons, T.L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine natural products as anticancer drugs. Mol. Cancer Ther. 2005, 4, 333–342. [Google Scholar] [PubMed]
- Youssef, F.S.; Ashour, M.L.; Singab, A.N.B.; Wink, M. A Comprehensive review of bioactive peptides from marine fungi and their biological significance. Mar. Drugs 2019, 17, 559. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.Y.; Williams, P.G.; Kwon, H.C.; Jensen, P.R.; Fenical, W. Lucentamycins A–D, cytotoxic peptides from the marine-derived actinomycete nocardiopsis lucentensis. J. Nat. Prod. 2007, 70, 1321–1328. [Google Scholar] [CrossRef]
- Williams, D.E.; Dalisay, D.S.; Patrick, B.O.; Matainaho, T.; Andrusiak, K.; Deshpande, R.; Myers, C.L.; Piotrowski, J.S.; Boone, C.; Yoshida, M.; et al. Padanamides A and B, highly modified linear tetrapeptides produced in culture by a Streptomyces sp. isolated from a marine sediment. Org. Lett. 2011, 13, 3936–3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Tasiamide, a cytotoxic peptide from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2002, 65, 1336. [Google Scholar] [CrossRef]
- Pranjol, M.Z.I.; Gutowski, N.J.; Hannemann, M.; Whatmore, J.L. Cathepsin D non-proteolytically induces proliferation and migration in human omental microvascular endothelial cells via activation of the ERK1/2 and PI3K/AKT pathways. Biochim. Biophys. Acta. Mol. Cell. Res. 2018, 1865, 25–33. [Google Scholar] [CrossRef]
- Li, Z.; Bao, K.; Xu, H.; Wu, P.; Li, W.; Liu, J.; Zhang, W. Design, synthesis, and bioactivities of tasiamide B derivatives as cathepsin D inhibitors. J. Pept. Sci. 2019, 25, e3154. [Google Scholar] [CrossRef]
- Simmons, T.L.; Mcphail, K.L.; Ortega-Barría, E.; Mooberry, S.L.; Gerwick, W.H. Belamide A, a new antimitotic tetrapeptide from a Panamanian marine cyanobacterium. Tetrahedron Lett. 2006, 37, 3387–3390. [Google Scholar] [CrossRef]
- Corbett, T.H. The molecular pharmacology of symplostatin 1: A new antimitotic dolastatin 10 analog. Int. J. Oncol. 2010, 104, 512–521. [Google Scholar] [CrossRef]
- Brucoli, F.; Natoli, A.; Marimuthu, P.; Borrello, M.T.; Stapleton, P.; Gibbons, S.; Schätzlein, A. Efficient synthesis and biological evaluation of proximicins A, B and C. Bioorg. Med. Chem. 2012, 20, 2019–2024. [Google Scholar] [CrossRef]
- Teruya, T.; Sasaki, H.; Fukazawa, H.; Suenaga, K. Bisebromoamide, a potent cytotoxic peptide from the marine cyanobacterium Lyngbya sp.: Isolation, stereostructure, and biological activity. Org. Lett. 2010, 41, 5062–5065. [Google Scholar] [CrossRef]
- Suzuki, K.; Mizuno, R.; Suenaga, K.; Teruya, T.; Tanaka, N.; Kosaka, T.; Oya, M. Bisebromoamide, an extract from Lyngbya species, induces apoptosis through ERK and mTOR inhibitions in renal cancer cells. Cancer Med. 2013, 2, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X. Isolation and identification of anti-proliferative peptides from Spirulina platensis using three-step hydrolysis. J. Sci. Food Agric. 2017, 97, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Joo, S.H. Recent trends in cyclic peptides as therapeutic agents and biochemical tools. Biomol. Ther. 2020, 28, 18–24. [Google Scholar] [CrossRef]
- Donia, M.S.; Wang, B.; Dunbar, D.C.; Desai, P.V.; Hamann, M.T. Mollamides B and C, cyclic hexapeptides from the indonesian tunicate Didemnum molle. Planta Med. 2008, 71, 941–945. [Google Scholar] [CrossRef]
- Wesson, K.J.; Hamann, M.T. Keenamide A, a bioactive cyclic peptide from the marine mollusk Pleurobranchus forskalii. J. Nat. Prod. 1996, 59, 629–631. [Google Scholar] [CrossRef] [PubMed]
- Mckeever, B.; Pattenden, G. Total synthesis of trunkamide A, a novel thiazoline-based prenylated cyclopeptide metabolite from Lissoclinum sp. Tetrahedron 2003, 59, 2713–2727. [Google Scholar] [CrossRef]
- Liang, B.; Richard, D.J.; Portonovo, P.S.; Joullié, M.M. Total syntheses and biological investigations of tamandarins A and B and tamandarin A analogs. J. Am. Chem. Soc. 2001, 123, 4469–4474. [Google Scholar] [CrossRef]
- Watters, D.J.; Beamish, H.J. Accumulation of HL-60 leukemia cells in G2/M and inhibition of cytokinesis caused by two marine compounds, bistratene A and cycloxazoline. Cancer Chemoth. Pharm. 1994, 33, 399–409. [Google Scholar] [CrossRef]
- Robinson, S.J.; Morinaka, B.I.; Amagata, T.; Tenney, K.; Crews, P. New structures and bioactivity properties of jasplakinolide (jaspamide) analogues from marine sponges. J. Med. Chem. 2010, 53, 1651–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, M.; Ionta, M.; Pfister, C.S.; Ferreira, A.S.A.R.; Machado-Santelli, M.G. Marine sponge depsipeptide increases gap junction length in HTC cells transfected with Cx43–GFP. Cell Biol. Int. Rep. 2013, 17, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettit, G.R.; Cichacz, Z.; Barkoczy, J.; Dorsaz, A.C.; Herald, D.L.; Williams, M.D.; Doubek, D.L.; Schmidt, J.M.; Tackett, L.P.; Brune, D.C. Isolation and structure of the marine sponge cell growth inhibitory cyclic peptide phakellistatin 1. J. Nat. Prod. 1993, 56, 260–267. [Google Scholar] [CrossRef]
- Guzmán, E.A.; Maers, K.; Roberts, J.; Kemami-Wangun, H.V.; Harmody, D.; Wright, A.E. The marine natural product microsclerodermin A is a novel inhibitor of the nuclear factor kappa B and induces apoptosis in pancreatic cancer cells. Investig. New Drugs. 2015, 33, 86. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.W.; Raventos Suarez, C.; Bifano, M.; Menendez, A.T.; Fairchild, C.R.; Faulkner, D.J. Scleritodermin A, a cytotoxic cyclic peptide from the lithistid sponge scleritoderma nodosum. J. Nat. Prod. 2004, 67, 475–478. [Google Scholar] [CrossRef]
- Wang, S.; Crevenna, A.H.; Ugur, I.; Marion, A.; Antes, I.; Kazmaier, U.; Hoyer, M.; Lamb, D.C.; Gegenfurtner, F.; Kliesmete, Z.; et al. Actin stabilizing compounds show specific biological effects due to their binding mode. Sci. Rep. 2019, 9, 9731. [Google Scholar] [CrossRef] [PubMed]
- Schweikart, K.; Guo, L.; Shuler, Z.; Abrams, R.; Chiao, E.T.; Kolaja, K.L.; Davis, M. The effects of jaspamide on human cardiomyocyte function and cardiac ion channel activity. Toxicol. In Vitro 2013, 27, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Cioca, D.P.; Kitano, K. Induction of apoptosis and CD10/neutral endopeptidase expression by jaspamide in HL-60 line cells. Cell Mol. Life Sci. 2002, 59, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.R.; Tinto, W.F.; Manchand, P.S.; Todaro, L.J. Stereostructures of Geodiamolides A and B, novel cyclodepsipeptides from the marine sponge Geodia sp. J. Org. Chem. 1987, 52, 3091–3093. [Google Scholar] [CrossRef]
- Tinto, W.F.; Lough, A.J.; McLean, S.; Reynolds, W.F.; Yu, M.; Chan, W.R. Geodiamolides H and I, further cyclodepsipeptides from the marine sponge Geodia sp. Tetrahedron 1998, 54, 4451–4458. [Google Scholar] [CrossRef]
- Rangel, M.; Prado, M.P.; Konno, K.; Naoki, H.; Freitas, J.C.; Machado-Santelli, G.M. Cytoskeleton alterations induced by Geodia corticostylifera depsipeptides in breast cancer cells. Peptides 2006, 27, 2047–2057. [Google Scholar] [CrossRef]
- Freitas, V.M.; Rangel, M.; Bisson, L.F.; Jaeger, R.G.; Machado-Santelli, G.M. The geodiamolide H, derived from Brazilian sponge Geodia corticostylifera, regulates actin cytoskeleton, migration and invasion of breast cancer cells cultured in three-dimensional environment. J. Cell. Physiol. 2008, 216, 583–594. [Google Scholar] [CrossRef]
- Meli, A.; Tedesco, C.; Della Sala, G.D.; Schettini, R.; Albericio, F.; De Riccardis, F.; Izzo, I. Phakellistatins: An underwater unsolved puzzle. Mar. Drugs 2017, 15, 78. [Google Scholar] [CrossRef] [Green Version]
- Pettit, G.R.; Tan, R. Isolation and structure of phakellistatin 14 from the western pacific marine sponge Phakellia sp. J. Nat. Prod. 2005, 68, 60–63. [Google Scholar] [CrossRef]
- Bao, Y.; Jiang, S.; Zhao, L.; Jin, Y.; Yan, R.; Wang, Z. Photoinduced synthesis and antitumor activity of a phakellistatin 18 analog with an isoindolinone fragment. New J. Chem. 2020, 44, 19174–19178. [Google Scholar] [CrossRef]
- Tian, T.; Takada, K.; Ise, Y.; Ohtsuka, S.; Okada, S.; Matsunaga, S. Microsclerodermins N and O, cytotoxic cyclic peptides containing a p-ethoxyphenyl moiety from a deep-sea marine sponge Pachastrella sp. Tetrahedron 2020, 76, 130997. [Google Scholar] [CrossRef]
- Vinothkumar, S.; Parameswaran, P.S. Recent advances in marine drug research. Biotechnol. Adv. 2013, 31, 1826–1845. [Google Scholar] [CrossRef]
- Oh, D.C.; Jensen, P.R.; Fenical, W. Zygosporamide, a cytotoxic cyclic depsipeptide from the marine-derived fungus Zygosporium masonii. Tetrahedron Lett. 2006, 47, 8625–8628. [Google Scholar] [CrossRef]
- Chen, Z.; Song, Y.; Chen, Y.; Huang, H.; Zhang, W.; Ju, J. Cyclic heptapeptides, cordyheptapeptides C–E, from the marine-derived fungus Acremonium persicinum SCSIO 115 and their cytotoxic activities. J. Nat. Prod. 2012, 75, 1215–1219. [Google Scholar] [CrossRef]
- He, F.; Bao, J.; Zhang, X.Y.; Tu, Z.C.; Shi, Y.M.; Qi, S.H. Asperterrestide A, a cytotoxic cyclic tetrapeptide from the marine-derived fungus Aspergillus terreus SCSGAF0162. J. Nat. Prod. 2013, 76, 1182–1186. [Google Scholar] [CrossRef]
- Ujiki, M.B.; Milam, B.; Ding, X.Z.; Roginsky, A.B.; Salabat, M.R.; Talamonti, M.S.; Bell, R.H.; Gu, W.; Silverman, R.B.; Adrian, T.E. A novel peptide sansalvamide analogue inhibits pancreatic cancer cell growth through G0/G1 cell-cycle arrest. Biochem. Biophys. Res. Commun. 2006, 340, 1224–1228. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, J.; Wu, H.; Li, Y.; Conti, P.S.; Chen, K. PET imaging of Hsp90 expression in pancreatic cancer using a new 64 Cu-labeled dimeric Sansalvamide A decapeptide. Amino Acids 2018, 50, 897–907. [Google Scholar] [CrossRef]
- Song, K.H.; Oh, S.J.; Kim, S.; Cho, H.; Lee, H.J.; Song, J.S.; Chung, J.Y.; Cho, E.; Lee, J.; Jeon, S.; et al. HSP90A inhibition promotes anti-tumor immunity by reversing multi-modal resistance and stem-like property of immune-refractory tumors. Nat. Commun. 2020, 11, 562. [Google Scholar] [CrossRef]
- Porter, N.J.; Christianson, D.W. Binding of the microbial cyclic tetrapeptide Trapoxin A to the class i histone deacetylase HDAC8. ACS Chem. Biol. 2017, 12, 2281–2286. [Google Scholar] [CrossRef] [PubMed]
- Itazaki, H.; Nagashima, K.; Sugita, K.; Yoshida, H.; Kawamura, Y.; Yasuda, Y.; Matsumoto, K.; Ishii, K.; Uotani, N.; Nakai, H.; et al. Isolation and structural elucidation of new cyclotetrapeptides, trapoxins A and B, having detransformation activities as antitumor agents. J. Antibiot. (Tokyo) 1990, 43, 1524–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijima, M.; Yoshida, M.; Sugita, K.; Horinouchi, S.; Beppu, T. Trapoxin, an antitumor cyclic tetrapeptide, is an irreversible inhibitor of mammalian histone deacetylase. J. Biol. Chem. 1993, 268, 22429–22435. [Google Scholar] [CrossRef]
- Jung, M.; Hoffmann, K.; Brosch, G.; Loidl, P. Analogues of trichostatin A and trapoxin B as histone deacetylase inhibitors. Bioorg. Med. Chem. Lett. 1997, 7, 1655–1658. [Google Scholar] [CrossRef]
- Gu, W.; Cueto, M.; Jensen, P.R.; Fenical, W.; Silverman, R.B. Microsporins A and B: New histone deacetylase inhibitors from the marine-derived fungus Microsporum cf. gypseum and the solid-phase synthesis of microsporin A. Tetrahedron 2007, 63, 6535–6541. [Google Scholar] [CrossRef]
- Karpiński, T.; Adamczak, A. Anticancer activity of bacterial proteins and peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.L.; Hua, H.M.; Pei, Y.H.; Yao, X.S. Three new cytotoxic cyclic acylpeptides from marine Bacillus sp. Chem. Pharm. Bull. (Tokyo) 2004, 52, 1029–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanoh, K.; Matsuo, Y.; Adachi, K.; Imagawa, H.; Nishizawa, M.; Shizuri, Y. Mechercharmycins A and B, cytotoxic substances from marine-derived Thermoactinomyces sp. YM3-251. J. Antibiot. (Tokyo) 2005, 58, 289–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberheide, A.; Schwenk, S.; Ronco, C.; Semmrau, L.M.; Gorls, H.; Arndt, H.D. Synthesis, structure, and cytotoxicity of urukthapelstatin A polyazole cyclopeptide analogs. Eur. J. Org. Chem. 2019, 2019, 4320–4326. [Google Scholar] [CrossRef]
- Asolkar, R.N.; Freel, K.C.; Jensen, P.R.; Fenical, W.; Pezzuto, J.M. Arenamides A–C, cytotoxic NFkappaB inhibitors from the marine actinomycete Salinispora arenicola. J. Nat. Prod. 2009, 72, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Medina, R.A.; Goeger, D.E.; Hills, P.; Mooberry, S.L.; Huang, N.; Romero, L.I.; Ortega-BarriA, E.; Gerwick, W.H.; Mcphail, K.L. Coibamide A, a potent antiproliferative cyclic depsipeptide from the Panamanian marine cyanobacterium Leptolyngbya sp. J. Am. Chem. Soc. 2008, 130, 6324–6325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, F.; Zhang, C.; Zhu, L.; Li, W.; Song, P.; Tao, Y.; Du, G. Synthesis and antitumor activity of cyclic octapeptide, samoamide A, and its derivatives. Med. Chem. Res. 2019, 28, 768–777. [Google Scholar] [CrossRef]
- Naman, C.B.; Rattan, R.; Nikoulina, S.E.; Lee, J.; Miller, B.W.; Moss, N.A.; Armstrong, L.; Boudreau, P.D.; Debonsi, H.M.; Valeriote, F.A.; et al. Integrating molecular networking and biological assays to target the isolation of a cytotoxic cyclic octapeptide, Samoamide A, from an American samoan marine cyanobacterium. J. Nat. Prod. 2017, 80, 625–633. [Google Scholar] [CrossRef]
- Huang, W.; Ren, R.-G.; Dong, H.-Q.; Wei, B.-G.; Lin, G.-Q. Diverse synthesis of marine cyclic depsipeptide Lagunamide A and its analogues. J. Org. Chem. 2014, 45, 10747–10762. [Google Scholar] [CrossRef]
- Luo, D.; Putra, M.Y.; Ye, T.; Paul, V.J.; Luesch, H. Isolation, structure elucidation and biological evaluation of lagunamide D: A new cytotoxic macrocyclic depsipeptide from marine cyanobacteria. Mar. Drugs 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, cancer cell toxins from a papua new guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, M.; Sueyoshi, K.; Teruya, T.; Ohno, H.; Fujii, N.; Oishi, S. Total synthesis of odoamide, a novel cyclic depsipeptide, from an okinawan marine cyanobacterium. Org. Biomol. Chem. 2016, 14, 9093–9104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.G.; Yoshida, W.Y.; Quon, M.K.; Moore, R.E.; Paul, V.J. The structure of Palau’amide, a potent cytotoxin from a species of the marine cyanobacterium Lyngbya. J. Nat. Prod. 2003, 66, 1545–1549. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Liu, Y.; Luesch, H. Systematic chemical mutagenesis identifies a potent novel Apratoxin A/E hybrid with improved in vivo antitumor activity. ACS Med. Chem. Lett. 2011, 2, 861–865. [Google Scholar] [CrossRef]
- Cai, W.; Ratnayake, R.; Gerber, M.H.; Chen, Q.Y.; Yu, Y.; Derendorf, H.; Trevino, J.G.; Luesch, H. Development of apratoxin S10 (Apra S10) as an anti-pancreatic cancer agent and its preliminary evaluation in an orthotopic patient-derived xenograft (PDX) model. Investig. New Drugs 2019, 37, 364–374. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L. Isolation, structure determination, and biological activity of Lyngbyabellin A from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 611–615. [Google Scholar] [CrossRef]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; Mcphail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine Cyanobacterium symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Ying, Y.; Taori, K.; Kim, H.; Hong, J.; Luesch, H. Total synthesis and molecular target of largazole, a histone deacetylase inhibitor. J. Am. Chem. Soc. 2008, 130, 8455–8459. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Park, H.; Salvador, L.A.; Serrano, P.E.; Kwan, J.C.; Zeller, S.L.; Chen, Q.Y.; Ryu, S.; Liu, Y.; Byeon, S.; et al. Evaluation of class I HDAC isoform selectivity of largazole analogues. Bioorg. Med. Chem. Lett. 2014, 24, 3728–3731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
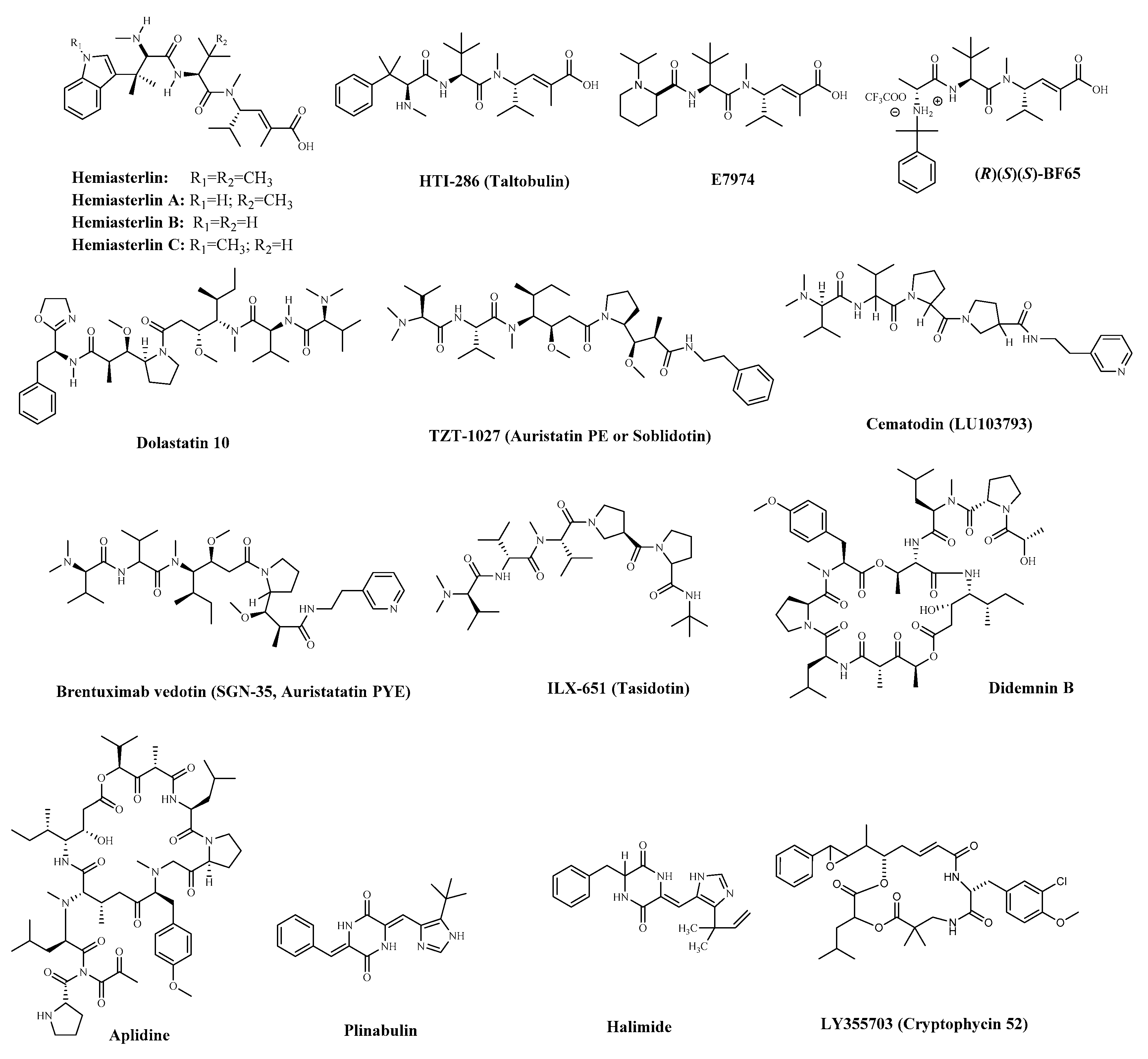
- Anderson, H.J.; Coleman, J.E.; Andersen, R.J.; Roberge, M. Cytotoxic peptides hemiasterlin, hemiasterlin A and hemiasterlin B induce mitotic arrest and abnormal spindle formation. Cancer Chemother. Pharmacol. 1996, 39, 223–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank LoVecchio, D.O.; Jane Klemens, R.N.; Ba, B.A.R.; Annie Klemens, B.A. HTI-286, a synthetic analogue of the tripeptide hemiasterlin, is a potent antimicrotubule agent that circumvents P-glycoprotein-mediated resistance in vitro and in vivo. Cancer Res. 2003, 63, 1838–1845. [Google Scholar] [CrossRef]
- Bai, R.; Durso, N.A.; Sackett, D.L.; Hamel, E. Interactions of the sponge-derived antimitotic tripeptide hemiasterlin with tubulin: Comparison with dolastatin 10 and cryptophycin 1. Biochemistry 1999, 38, 14302. [Google Scholar] [CrossRef]
- Kuznetsov, G.; TenDyke, K.; Towle, M.J.; Cheng, H.; Liu, J.; Marsh, J.P.; Schiller, S.E.R.; Spyvee, M.R.; Yang, H.; Seletsky, B.M.; et al. Tubulin-based antimitotic mechanism of E7974, a novel analogue of the marine sponge natural product hemiasterlin. Mol. Cancer Ther. 2009, 8, 2852–2860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poruchynsky, M.S.; Kim, J.H.; Nogales, E.; Annable, T.; Loganzo, F.; Greenberger, L.M.; Sackett, D.L.; Fojo, T. Tumor cells resistant to a microtubule-depolymerizing hemiasterlin analogue, HTI-286, have mutations in alpha- or beta-tubulin and increased microtubule stability. Biochemistry 2004, 43, 13944–13954. [Google Scholar] [CrossRef]
- Lai, W.T.; Cheng, K.L.; Baruchello, R.; Rondanin, R.; Marchetti, P.; Simoni, D.; Lee, R.M.; Guh, J.H.; Hsu, L.C. Hemiasterlin derivative (R)(S)(S)-BF65 and Akt inhibitor MK-2206 synergistically inhibit SKOV3 ovarian cancer cell growth. Biochem. Pharmacol. 2016, 113, 12–23. [Google Scholar] [CrossRef]
- Hsu, L.C.; Durrant, D.E.; Huang, C.C.; Chi, N.W.; Baruchello, R.; Rondanin, R.; Rullo, C.; Marchetti, P.; Grisolia, G.; Simoni, D.; et al. Development of hemiasterlin derivatives as potential anticancer agents that inhibit tubulin polymerization and synergize with a stilbene tubulin inhibitor. Investig. New Drugs 2012, 30, 1379–1388. [Google Scholar] [CrossRef]
- Pettit, G.; Kamano, Y.; Herald, C.; Tuinman, A.; Boettner, F.; Kizu, H.; Schmidt, J.; Baczynskyj, L.; Tomer, K.; Bontems, R. The isolation and structure of a remarkable marine animal antineoplastic constituent: Dolastatin 10. J. Am. Chem. Soc. 1986, 109, 6883–6885. [Google Scholar] [CrossRef]
- Hamel, E. Interactions of antimitotic peptides and depsipeptides with tubulin. Biopolymers 2002, 66, 142–160. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.L.; Pettit, G.R.; Hamel, E. Binding of dolastatin 10 to tubulin at a distinct site for peptide antimitotic agents near the exchangeable nucleotide and vinca alkaloid sites. J. Biol. Chem. 1990, 265, 17141–17149. [Google Scholar] [CrossRef]
- Ebbinghaus, S.; Hersh, E.; Cunningham, C.C.; O’Day, S.; Mcdermott, D.; Stephenson, J.; Richards, D.A.; Eckardt, J.; Haider, O.L.; Hammond, L.A. Phase II study of synthadotin (SYN-D; ILX651) administered daily for 5 consecutive days once every 3 weeks (qdx5q3w) in patients (Pts) with inoperable locally advanced or metastatic melanoma. J. Clin. Oncol. 2004, 22, 7530. [Google Scholar] [CrossRef]
- Mross, K.; Herbst, K.; Berdel, W.E.; Korfel, A.; von Broen, I.M.; Bankmann, Y.; Hossfeld, D.K. Phase I clinical and pharmacokinetic study of LU103793 (Cemadotin Hydrochloride) as an intravenous bolus injection in patients with metastatic solid tumors. Onkologie 1996, 19, 490–495. [Google Scholar] [CrossRef]
- Natsume, T.; Watanabe, J.I.; Koh, Y.; Fujio, N.; Kobayashi, M. Antitumor activity of TZT-1027 (Soblidotin) against vascular endothelial growth factor-secreting human lung cancer in vivo. Cancer Sci. 2010, 94, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.; Cragg, G. Natural products from marine invertebrates and microbes as modulators of antitumor targets. Curr. Drug Targets 2006, 7, 279–304. [Google Scholar] [CrossRef]
- Beckwith, M.; Urba, W.J.; Longo, D.L. Growth inhibition of human lymphoma cell lines by the marine products, Dolastatins 10 and 15. J. Natl. Cancer Inst. 1993, 85, 483–488. [Google Scholar] [CrossRef]
- Keir, C.H.; Vahdat, L.T. The use of an antibody drug conjugate, glembatumumab vedotin (CDX-011), for the treatment of breast cancer. Expert. Opin. Biol. Ther. 2012, 12, 259–263. [Google Scholar] [CrossRef]
- Yamamoto, N.; Andoh, M.; Kawahara, M.; Fukuoka, M.; Niitani, H. Phase I study of TZT-1027, a novel synthetic dolastatin 10 derivative and inhibitor of tubulin polymerization, given weekly to advanced solid tumor patients for 3 weeks. Cancer Sci. 2009, 100, 316–321. [Google Scholar] [CrossRef]
- Shnyder, S.D.; Cooper, P.A.; Millington, N.J.; Pettit, G.R.; Bibby, M.C. Auristatin PYE, a novel synthetic derivative of dolastatin 10, is highly effective in human colon tumour models. Int. J. Oncol. 2007, 31, 353. [Google Scholar] [CrossRef] [Green Version]
- Prokopiou, E.M.; Cooper, P.A.; Pettit, G.R.; Bibby, M.C.; Shnyder, S.D. Potentiation of the activity of cisplatin in a human colon tumour xenograft model by auristatin PYE, a structural modification of dolastatin 10. Mol. Med. Rep. 2010, 3, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Okouneva, T.; Manna, T.; Miller, H.P.; Schmid, S.; Arthaud, L.; Luduena, R.; Jordan, M.A.; Wilson, L. Mechanism of action of the microtubule-targeted antimitotic depsipeptide tasidotin (formerly ILX651) and its major metabolite tasidotin C-carboxylate. Cancer Res. 2007, 67, 3767–3776. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.; Appleman, L.J.; Kirvan-Visovatti, M.; Ryan, D.P.; Regan, E.; Vukelja, S.; Bonate, P.L.; Ruvuna, F.; Fram, R.J.; Jekunen, A.; et al. Phase I and pharmacokinetic study of the dolastatin-15 analogue tasidotin (ILX651) administered intravenously on days 1, 3, and 5 every 3 weeks in patients with advanced solid tumors. Clin. Cancer Res. 2005, 11, 7825–7833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebbinghaus, S.; Rubin, E.; Hersh, E.; Cranmer, L.D.; Bonate, P.L.; Fram, R.J.; Jekunen, A.; Weitman, S.; Hammond, L.A. A phase I study of the dolastatin-15 analogue tasidotin (ILX651) administered intravenously daily for 5 consecutive days every 3 weeks in patients with advanced solid tumors. Clin. Cancer Res. 2005, 11, 7807–7816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mita, A.C.; Hammond, L.A.; Bonate, P.L.; Weiss, G.; McCreery, H.; Syed, S.; Garrison, M.; Chu, Q.S.C.; DeBono, J.S.; Jones, C.B.; et al. Phase I and pharmacokinetic study of tasidotin hydrochloride (ILX651), a third-generation dolastatin-15 analogue, administered weekly for 3 weeks every 28 days in patients with advanced solid tumors. Clin. Cancer Res. 2006, 12, 5207–5215. [Google Scholar] [CrossRef] [Green Version]
- Naumovski, L.; Junutula, J.R. Glembatumumab vedotin, a conjugate of an anti-glycoprotein non-metastatic melanoma protein B mAb and monomethyl auristatin E for the treatment of melanoma and breast cancer. Curr. Opin. Mol. Ther. 2010, 12, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Hasanov, M.; Rioth, M.J.; Kendra, K.; Hernandez-Aya, L.; Joseph, R.W.; Williamson, S.; Chandra, S.; Shirai, K.; Turner, C.D.; Lewis, K.; et al. A phase ii study of glembatumumab vedotin for metastatic uveal melanoma. Cancers 2020, 12, 2270. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, S.M.; Advani, R.H.; Bartlett, N.L.; Jacobsen, E.D.; Sharman, J.P.; O’Connor, O.A.; Siddiqi, T.; Kennedy, D.A. Objective responses in relapsed T-cell lymphomas with single-agent brentuximab vedotin. Blood 2014, 123, 3095–3100. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, K.L.J.; Gloer, J.B.; Cook, J.C.J. Structures of the didemnins, antiviral and cytotoxic depsipeptides from a Caribbean tunicate. J. Am. Chem. Soc. 1981, 12, 1857–1859. [Google Scholar] [CrossRef]
- Urdiales, J.; Morata, P.; Castro, I.N.D.; Sánchez-Jiménez, F. Antiproliferative effect of dehydrodidemnin B (DDB), a depsipeptide isolated from Mediterranean tunicates. Cancer Lett. 1996, 102, 31–37. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, M.; Joshi, P.; Rawat, D.S. Clinical status of anti-cancer agents derived from marine sources. Anticancer Agents Med. Chem. 2008, 8, 603–617. [Google Scholar] [CrossRef]
- Maroun, J.A.; Belanger, K.; Seymour, L.; Matthews, S.; Roach, J.; Dionne, J.; Soulieres, D.; Stewart, D.; Goel, R.; Charpentier, D. Phase I study of Aplidine in a daily×5 one-hour infusion every 3 weeks in patients with solid tumors refractory to standard therapy. Ann. Oncol. 2006, 17, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Toulmonde, M.; Le Cesne, A.; Piperno-Neumann, S.; Penel, N.; Chevreau, C.; Duffaud, F.; Bellera, C.; Italiano, A. Aplidin in patients with advanced dedifferentiated liposarcomas: A french sarcoma group single-arm phase II study. Ann. Oncol. 2015, 26, 1465–1470. [Google Scholar] [CrossRef]
- White, K.M.; Rosales, R.; Yildiz, S.; Kehrer, T.; Miorin, L.; Moreno, E.; Jangra, S.; Uccellini, M.B.; Rathnasinghe, R.; Coughlan, L.; et al. Plitidepsin has potent preclinical efficacy against SARS-CoV-2 by targeting the host protein eEF1A. Science 2021, 10, 1126. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Sumikura, M.; Hidaka, K.; Yasui, H.; Kiso, Y.; Yakushiji, F.; Hayashi, Y. Anti-microtubule ‘plinabulin’ chemical probe KPU-244-B3 labeled both α- and β-tubulin. Bioorg. Med. Chem. 2010, 18, 3169–3174. [Google Scholar] [CrossRef] [PubMed]
- Millward, M.; Mainwaring, P.; Mita, A.; Federico, K.; Lloyd, G.K.; Reddinger, N.; Nawrocki, S.; Mita, M.; Spear, M.A. Phase 1 study of the novel vascular disrupting agent plinabulin (NPI-2358) and docetaxel. Investig. New Drugs 2012, 30, 1065–1073. [Google Scholar] [CrossRef]
- Fenical, W.; Jensen, P.R.; Cheng, X.C. Halimide, a Cytotoxic Marine Natural Product, and Derivatives Thereof. U.S. Patent No. 6,069,146, 30 May 2000. [Google Scholar]
- Mooberry, S.L.; Busquets, L.; Tien, G. Induction of apoptosis by cryptophycin 1, a new antimicrotubule agent. Int. J. Cancer 1997, 73, 440–448. [Google Scholar] [CrossRef]
- Ding, Y.; Rath, C.M.; Bolduc, K.L.; Hakansson, K.; Sherman, D.H. Chemoenzymatic synthesis of cryptophycin anticancer agents by an ester bond-forming non-ribosomal peptide synthetase module. J. Am. Chem. Soc. 2011, 133, 14492–14495. [Google Scholar] [CrossRef] [Green Version]
- Sessa, C.; Weigang-Köhler, K.; Pagani, O.; Greim, G.; Mora, O.; De Pas, T.; Burgess, M.; Weimer, I.; Johnson, R. Phase I and pharmacological studies of the cryptophycin analogue LY355703 administered on a single intermittent or weekly schedule. Eur. J. Cancer 2002, 38, 2388–2396. [Google Scholar] [CrossRef]
- Liu, L.; Bao, Y.; Zhang, Y.; Xiao, C.; Chen, L. Acid-responsive dextran-based therapeutic nanoplatforms for photodynamic-chemotherapy against multidrug resistance. Int. J. Biol. Macromol. 2020, 155, 233–240. [Google Scholar] [CrossRef]
- Pei, X.; Zhu, Z.; Gan, Z.; Chen, J.; Zhang, X.; Cheng, X.; Wan, Q.; Wang, J. PEGylated nano-graphene oxide as a nanocarrier for delivering mixed anticancer drugs to improve anticancer activity. Sci. Rep. 2020, 10, 2717. [Google Scholar] [CrossRef] [PubMed]
- Conibear, A.C.; Schmid, A.; Kamalov, M.; Becker, C.F.W.; Bello, C. Recent advances in peptide-based approaches for cancer treatment. Curr. Med. Chem. 2020, 27, 1174–1205. [Google Scholar] [CrossRef]
- Zhao, Y.; Cai, C.; Liu, M.; Zhao, Y.; Wu, Y.; Fan, Z.; Ding, Z.; Zhang, H.; Wang, Z.; Han, J. Drug-binding albumins forming stabilized nanoparticles for co-delivery of paclitaxel and resveratrol: In vitro/in vivo evaluation and binding properties investigation. Int. J. Biol. Macromol. 2020, 153, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, A.; Deyle, K.; Heinis, C. Cyclic peptide therapeutics: Past, present and future. Curr. Opin. Chem. Biol. 2017, 38, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogervorst, T.P.; Li, R.J.E.; Marino, L.; Bruijns, S.C.M.; Meeuwenoord, N.J.; Filippov, D.V.; Overkleeft, H.S.; van der Marel, G.A.; van Vliet, S.J.; van Kooyk, Y.; et al. C-Mannosyl lysine for solid phase assembly of mannosylated peptide conjugate cancer vaccines. ACS Chem. Biol. 2020, 15, 728–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, G.M.; Sedlik, C.; Baharom, F.; Zhu, Y.; Ramirez-Valdez, R.A.; Coble, V.L.; Tobin, K.; Nichols, S.R.; Itzkowitz, Y.; Zaidi, N.; et al. Peptide-TLR-7/8a conjugate vaccines chemically programmed for nanoparticle self-assembly enhance CD8 T-cell immunity to tumor antigens. Nat. Biotechnol. 2020, 38, 320–332. [Google Scholar] [CrossRef]
- Habault, J.; Kaci, A.; Pasquereau-Kotula, E.; Fraser, C.; Chomienne, C.; Dombret, H.; Braun, T.; Pla, M.; Poyet, J.L. Prophylactic and therapeutic antileukemic effects induced by the AAC-11-derived Peptide RT53. Oncoimmunology 2020, 9, 1728871. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Source | Mechanism | Cell Lines | IC50/(GI50) b | Reference |
|---|---|---|---|---|---|
| QPK | Sepia ink | Cytotoxicity a | DU-145 | 9.50 mg/mL (24 h); | [40] |
| 1.00 mg/mL (48 h) | |||||
| AGAPGG; | Sarcophyton glaucum | Cytotoxicity a | HeLa | 8.6 mmol/L; | [41] |
| AERQ; | 4.9 mmol/L; | ||||
| RDTQ | 5.6 mmol/L | ||||
| Virenamides A; | The Didemnid ascidian Diplosoma virens | Inhibiting the Topoisomerase II | P388; | 2.5 µg/mL | [42] |
| A549; | 10 µg/mL | ||||
| HT-29; | 10 µg/mL | ||||
| CV1 | 10 µg/mL | ||||
| Virenamides B | The Didemnid ascidian Diplosoma virens | Inhibiting the Topoisomerase II | P388; | 5 µg/mL | |
| A549; | 5 µg/mL | ||||
| HT-29; | 5 µg/mL | ||||
| CV1 | 5 µg/mL | ||||
| Virenamides C | The Didemnid ascidian Diplosoma virens | Inhibiting the Topoisomerase II | P388; | 5 µg/mL | |
| A549; | 5 µg/mL | ||||
| HT-29; | 5 µg/mL | ||||
| CV1 | 5 µg/mL | ||||
| SCAP1; (Leu-Ala-Asn-Ala-Lys) | Oyster (Saccostrea cucullata) | Enhancing oxidative DNA damage; Inducing apoptosis | HT-29 | 90.31 mg/mL (24 h); | [43,44] |
| 70.87 mg/mL (48 h); | |||||
| 60.21 mg/mL (72 h) | |||||
| YALPAH | Half-fin anchovy (Setipinna taty) | Inducing apoptosis | PC‑3 | 8.1 mg/mL | [45,46] |
| BCP-A (Trp-Pro-Pro) | Blood clam (Tegillarca granosa) muscle | Inducing apoptosis and inhibiting lipid peroxidation | PC-3; | 1.99 mg/mL; | [47] |
| DU-145; | 2.80 mg/mL; | ||||
| H-1299; | 3.3 mg/mL; | ||||
| HeLa | 2.54 mg/mL | ||||
| BDS-I; (Ala-Ala-Pro-Ala-Phe-Ala-Ser-Gly) | The sea anemone toxin | Blocking KV3.4 currents prevented (the neurotoxic β-amyloid peptide1-42) Aβ1−42-induced caspase-3 activation and apoptotic processes | PC-12 | 75 nM | [49,50] |
| FIMGPY | The skate (R. porosa) cartilage protein hydrolysate | Inducing apoptosis by upregulating the Bax/Bcl-2 ratio and caspase-3 activation | HeLa | 4.81 mg/mL | [51] |
| AAP-H; (Tyr-Val-Pro-Gly-Pro) | The sea anemone Anthopleura anjunae | Inducing apoptosis, decreasing the mitochondrial membrane potential, and increasing Bax/Bcl-2 ratio, cytochrome-C, caspase-3, and caspase-9 | DU-145 | 9.605 mM (24 h); | [52] |
| 7.910 mM (48 h); | |||||
| 2.298 mM (72 h) | |||||
| ILYMP | Cyclina sinensis | Enhancing expression of Bax, cleaved caspase-3/9 as well as suppression of Bcl-2 expression | DU-145 | 11.25 mM | [53] |
| SCH-P9 (Leu-Pro-Gly-Pro) | Sinonovacula constricta hydrolysates | Inducing apoptosis and sub-G1 phase cell cycle arrest | DU‑145; | 1.21 mg/mL (24 h); | [54] |
| PC‑3 | 1.09 mg/mL (24 h) | ||||
| SCH-P10 (Asp-Tyr-Val-Pro) | DU‑145; | 1.41 mg/mL (24 h); | |||
| PC‑3 | 0.91 mg/mL (24 h) | ||||
| SIO | Sepia ink | Inducing apoptosis, and S and G2/M phase cell cycle arrest | DU-145; | <5 mg/mL | [55,56] |
| PC-3; | <5 mg/mL | ||||
| LNCaP | <10 mg/mL | ||||
| Psammaplin A (PsA) | The two Sponges, Jaspis sp.and Poecillastra wondoensis. | Inducing S or S-G2/M phase cell cycle arrest; Inhibting HDAC | P388; HCT-116; A549 | (40 nM) | [58,59] |
| NVP-LAQ824 | Psammaplysilla sp. | Inducing S or S-G2/M phase cell cycle arrest; Inhibting HDAC | H-1299 | 150 nM | [60] |
| HCT-116 | 10 nM | ||||
| Lucentamycins A; | The fermentation broth of a marine-derived actinomycete | Cytotoxicity a | HCT-116 | 0.20 µM; | [62] |
| Lucentamycins B | 11 µM | ||||
| Padanamides A and B | Sediment in the culture of Streptomyces sp. | Cytotoxicity a | Jurkat | 30.9 µM | [63] |
| Tasiamide | Cyanobacterial compound derived from Symploca sp. | Inhibiting the expression of Cath D | KB; | 0.48 μg/mL; | [64,66] |
| LoVo | 3.47 μg/mL | ||||
| Belamide A | Cyanobacterium | Tubulin polymerization inhibition | HCT-116; | 0.74 μM; | [67] |
| A-10 | 20 μM | ||||
| Symplostatin A | Cyanobacterium | Microtubule assembly Inhibiting cell cycle arrest | MDA-MB-435 | 0.15 μM | [68] |
| SK-OV-3; | 0.09 μM | ||||
| NCI/ADR; | 2.90 μM | ||||
| NCI/ADR with Verapamil; | 0.09 μM | ||||
| A-10; | 1.8 μM | ||||
| HUVEC | 0.16 μM | ||||
| Proximicins C | Actinomycetes of the genus Verrucosispora, | Inducing Cell cycle G1 to S phase arrest and inducing apoptotic cell death | U-87 MG; | 12.7 μg/mL | [69] |
| MDA-MD-231 | 11.4 μg/mL | ||||
| Bisebromoamide | Cyanobacterium of the genus Lyngbya sp. | Inhibiting both the Raf/MEK/ERK and PI3K/Akt/mTOR pathways | JFCR39 | (40 nM) | [70,71] |
| HVLSRAPR | Spirulina platensis | Cytotoxicity a | HT-29; | 99.88 µg/mL | [72] |
| Compound | Source | Mechanism | Cell Lines | IC50/(GI50) b | Reference |
|---|---|---|---|---|---|
| Mollamide B | Tunicate Didemnum | Cytotoxicity a | H460; | (>100 µM) | [74] |
| MCF-7; | |||||
| SF-268 | |||||
| Keenamide A | Tunicate Didemnum. | Cytotoxicity a | P-388; | 2.5 µg/mL; | [75] |
| A-549; | 2.5 µg/mL; | ||||
| MEL-20; | 2.5 µg/mL; | ||||
| HT-29 | 5.0 µg/mL | ||||
| Trunkamide A | Didemnid ascidians | Cytotoxicity a | DU-145; | 7.08 nM; | [76] |
| IGROV; | 7.31 nM; | ||||
| SK-BR-3; | 5.44 nM; | ||||
| Hela | 3.90 nM | ||||
| Tamandarin A | Didemnid ascidians | Cytotoxicity a | NCI-60 | 1.4 µM (2.3 µM) | [77] |
| Tamandarin B | NCI-60 | 1.4 µM (2.3 µM) | |||
| Cycloxazoline | Didemnid ascidians | Cell cycle G2/M arrest, Induction of apoptosis | MRC5CV1; T24 | 0.5 μg/mL | [77] |
| Jaspamide (Jasplakinolide, NSC-613009) | Sponge Jaspis johnstoni | Induced apoptosis is associated with caspase-3 activation, increased Bax level, and decreased Bcl-2 protein expression | T24; MCF-7; 15NCI/ADR; A-10 | 60 to 150 µg /mL | [85,86] |
| Geodiamolide A | Sponge Geodia corticostylifera | Induction of apoptosis; Tubulin polymerization inhibition | T47D; | 18.82 nM; | [89] |
| MCF7 | 17.83 nM; | ||||
| Geodiamolides B | T47D; | 113.90 nM; | |||
| MCF7 | 9.82 nM; | ||||
| Phakellistatin 13 | Sponge Phakellia sp. | Induction of both intrinsic and extrinsic apoptosis | BEL-7404 | (10 ng/mL) | [91,92] |
| Phakellistatin 14 | P388 | (5 µg/ mL) | |||
| Microsclerodermin A | Sponge of the genus Amphibleptula | Inhibit NFκB, Induction of apoptosis; | AsPC-1; | 2.3 μM; | [82] |
| BxPC-3; | 0.8 μM; | ||||
| MIA PaCa-2; | 4.3 μM; | ||||
| PANC-1; | 4.0 μM | ||||
| Scleritodermin A | Sponge Scleritoderma nodosum | Tubulin polymerization inhibition | HCT-116; | 1.9 µM; | [83] |
| A2780; | 0.940 µM; | ||||
| SKBR3 | 0.670 µM |
| Compound | Source | Mechanism | Cell Lines | IC50/(GI50) b | Reference |
|---|---|---|---|---|---|
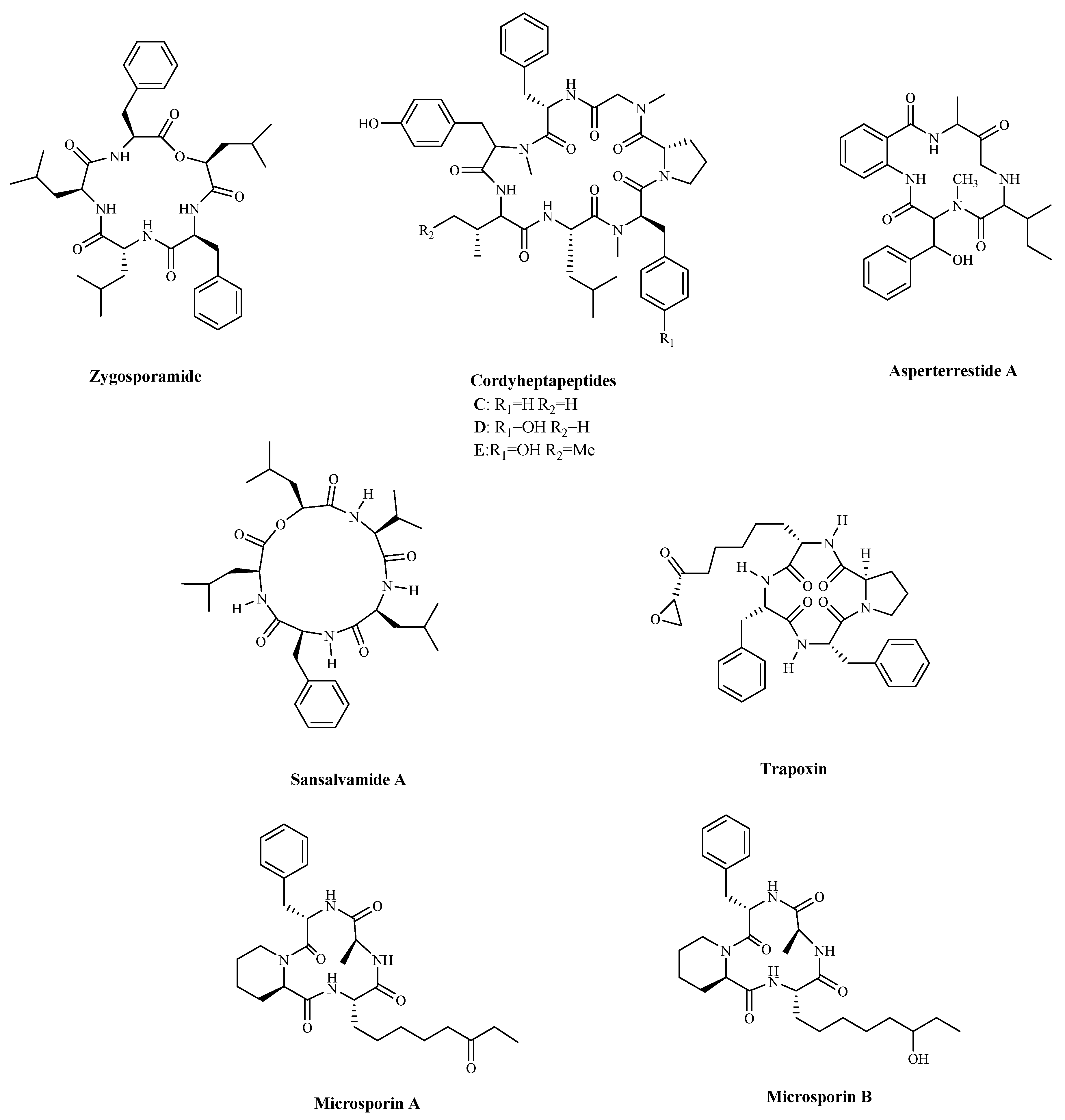
| Zygosporamide | Zygosporium masonii | Cytotoxicity a | SF-268; | (6.5 nM) | [96] |
| RXF 393 | (5.0 nM) | ||||
| Cordyheptapeptide C | Acremonium persicinum | Cytotoxicity a | SF-268; | 3.7 μM; | [97] |
| MCF-7; | 3.0 μM; | ||||
| NCI-H460 | 11.6 μM | ||||
| Cordyheptapeptide D | Acremonium persicinum | Cytotoxicity a | SF-268; | 45.6 μM; | |
| MCF-7; | 82.7 μM; | ||||
| NCI-H460 | >100 μM | ||||
| Cordyheptapeptide E | Acremonium persicinum | Cytotoxicity a | SF-268; | 3.2 μM; | |
| MCF-7; | 2.7 μM; | ||||
| NCI-H460 | 4.5 μM | ||||
| Asperterrestide A | Aspergillus terreus | Cytotoxicity a | U937; | 6.4 μM; | [98] |
| MOLT4 | 6.2 μM | ||||
| Sansalvamide A | Microsporum cf. gypseum | Inhibiting cell growth, and proliferation, and inducing cell apoptosis by regulating the expression of HSP90 | HCT-116; | 1.5 µM; | [99,100] |
| HCT-15 | 1 µM | ||||
| Trapoxin | Fungal product the culture broth of Helicoma ambiens RF-1023 | Inhibiting HDAC | NIH3T3 | 200 ng/mL | [104,105] |
| Microsporin A | Microsporum cf. gypseum | Inhibiting HDAC | HCT-116 | 0.6 mg/mL; | [106] |
| Microsporin B | HCT-116 | 8.5 mg/mL |
| Compound | Source | Mechanism | Cell Lines | IC50 | Reference |
|---|---|---|---|---|---|
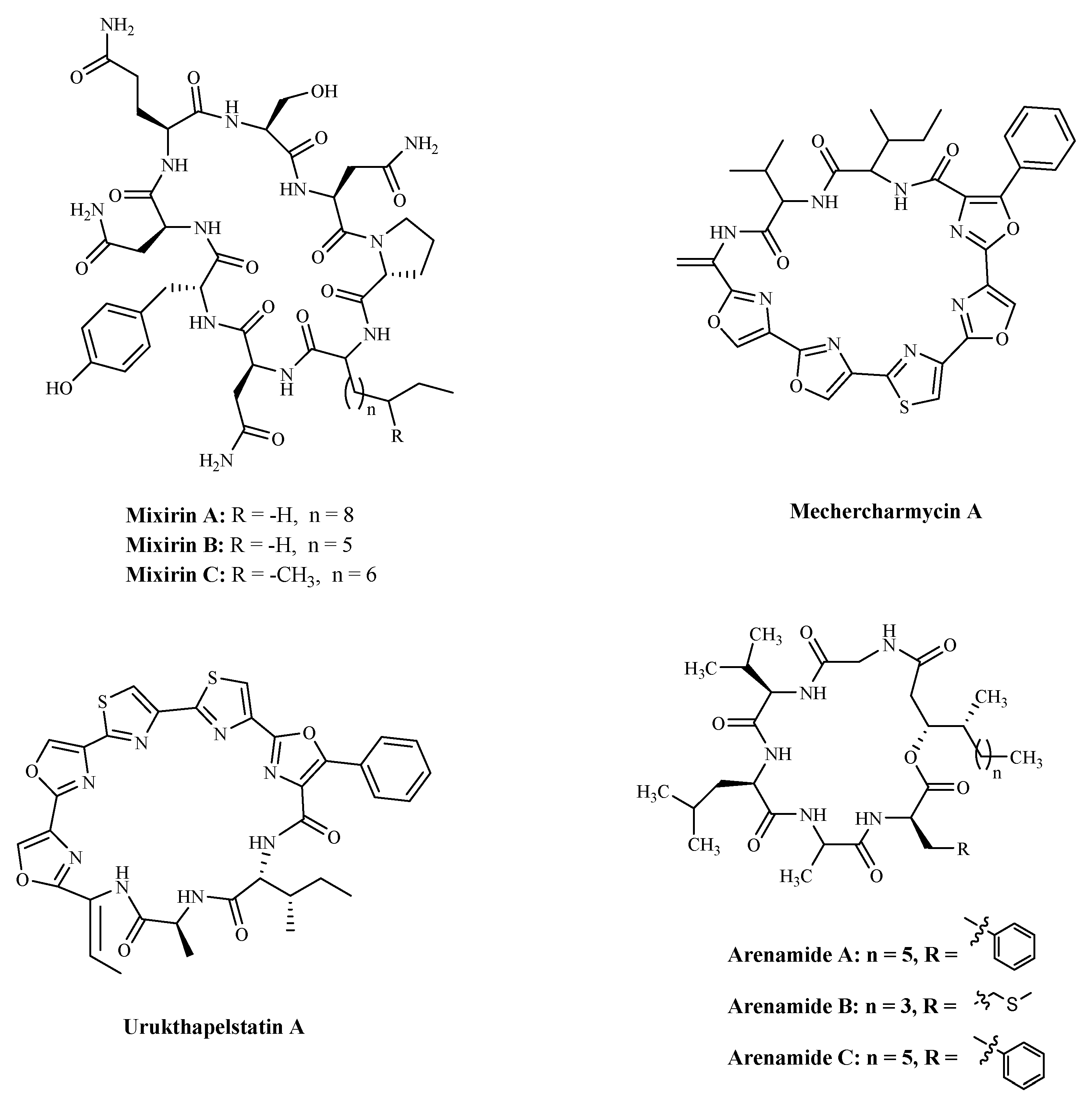
| Mixirin A | Bacillus species. | Cytotoxicity a | HCT-116 | 0.65 µM; | [108] |
| Mixirin B | HCT-116 | 1.6 µM; | |||
| Mixirin C | HCT-116 | 1.26 µM; | |||
| Mechercharmycin A | Thermoactinomyces sp. YM3-251 | Cytotoxicity a | A549; | 40 nM; | [109] |
| Jurkat | 46 nM; | ||||
| Urukthapelstatin A | Thermoactinomycetaceae bacterium Mechercharimyces asporophorigenens YM11-542 | Cytotoxicity a | A549 | 12 nM; | [110] |
| Arenamide A | Salinispora Arenicola. | Inhibiting NF kappa B | 293/NF-κB-Luc | 3.7 μM; | [111] |
| Arenamide B | 293/NF-κB-Luc | 1.7 μM |
| Compound | Source | Mechanism | Cell Lines/Target protein | IC50 | Reference |
|---|---|---|---|---|---|
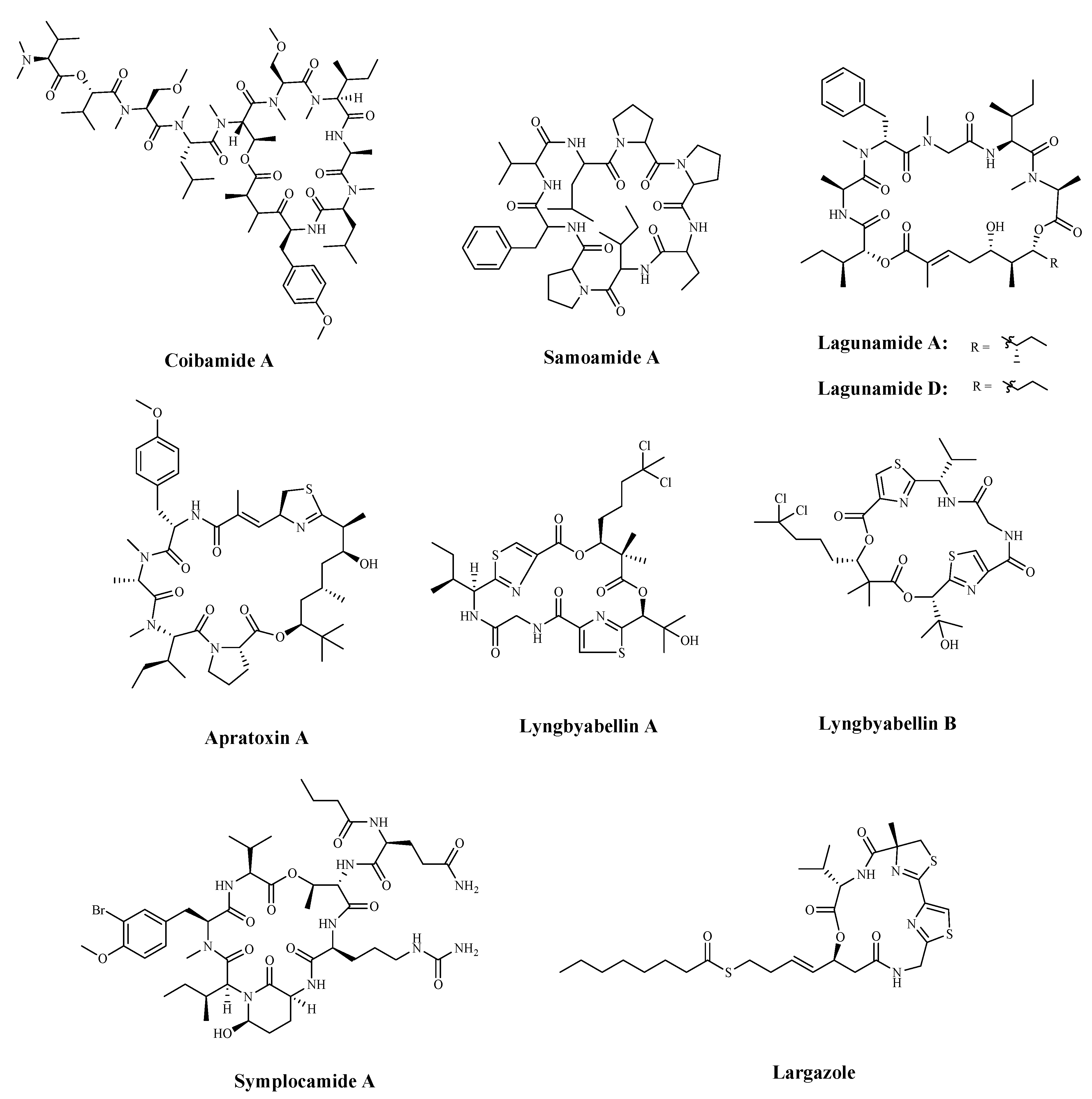
| Coibamide A | Leptolyngbya cyanobacterium | Cytotoxicity a | NCI-H460 | <23 nM | [112] |
| Samoamide A | Symploca sp. | Cytotoxicity a | H460; | 1.1 μM; | [113,114] |
| H116 | 4.5 μM | ||||
| Lagunamide A | cyanobacterium, Lyngbya majuscule, | Cytotoxicity; Caspase-mediated mitochondrial apoptosis | A549 | 7.1 nM; | [116] |
| Lagunamide D | A549 | 6.7 nM | |||
| Apratoxin A | Cyanobacterial metabolite | Down-regulating receptors and growth factor ligands for cancer cells that rely on autocrine loop | HCT-116 | 5.97 nM | [120] |
| Lyngyabellin A | cyanobacterium Lyngbya majuscula | Antiproliferation; Disruption of cellular microfilament network cytokinesis | KB | 0.03 µg/mL; | [122] |
| LoVo | 0.50 µg/mL | ||||
| Chymotrypsin | 0.234 µM | ||||
| Symplocamide A | Cyanobacteria Symploca sp. | Inhibiting Protease | H-460; | 40 nM | [123] |
| Neuro-2a neuroblastoma | 29 nM | ||||
| Largazole | cyanobacteria Symploca sp. | Inhibiting HDAC | HCT-116 | 44 nM | [124,125] |
| Inhibit HDAC1 | 25 nM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.-T.; Liu, Z.-D.; Wang, Z.; Wang, T.; Wang, N.; Wang, N.; Zhang, B.; Zhao, Y.-F. Recent Advances in Small Peptides of Marine Origin in Cancer Therapy. Mar. Drugs 2021, 19, 115. https://doi.org/10.3390/md19020115
Zhang Q-T, Liu Z-D, Wang Z, Wang T, Wang N, Wang N, Zhang B, Zhao Y-F. Recent Advances in Small Peptides of Marine Origin in Cancer Therapy. Marine Drugs. 2021; 19(2):115. https://doi.org/10.3390/md19020115
Chicago/Turabian StyleZhang, Qi-Ting, Ze-Dong Liu, Ze Wang, Tao Wang, Nan Wang, Ning Wang, Bin Zhang, and Yu-Fen Zhao. 2021. "Recent Advances in Small Peptides of Marine Origin in Cancer Therapy" Marine Drugs 19, no. 2: 115. https://doi.org/10.3390/md19020115
APA StyleZhang, Q. -T., Liu, Z. -D., Wang, Z., Wang, T., Wang, N., Wang, N., Zhang, B., & Zhao, Y. -F. (2021). Recent Advances in Small Peptides of Marine Origin in Cancer Therapy. Marine Drugs, 19(2), 115. https://doi.org/10.3390/md19020115