
Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action

 ,
,  ,
,
Abstract
:1. Introduction

| Drug Class | Compound/ Organism a | Chemistry | Pharmacologic Activity | IC50 b1 | MMOA b2 | Country c | References |
|---|---|---|---|---|---|---|---|
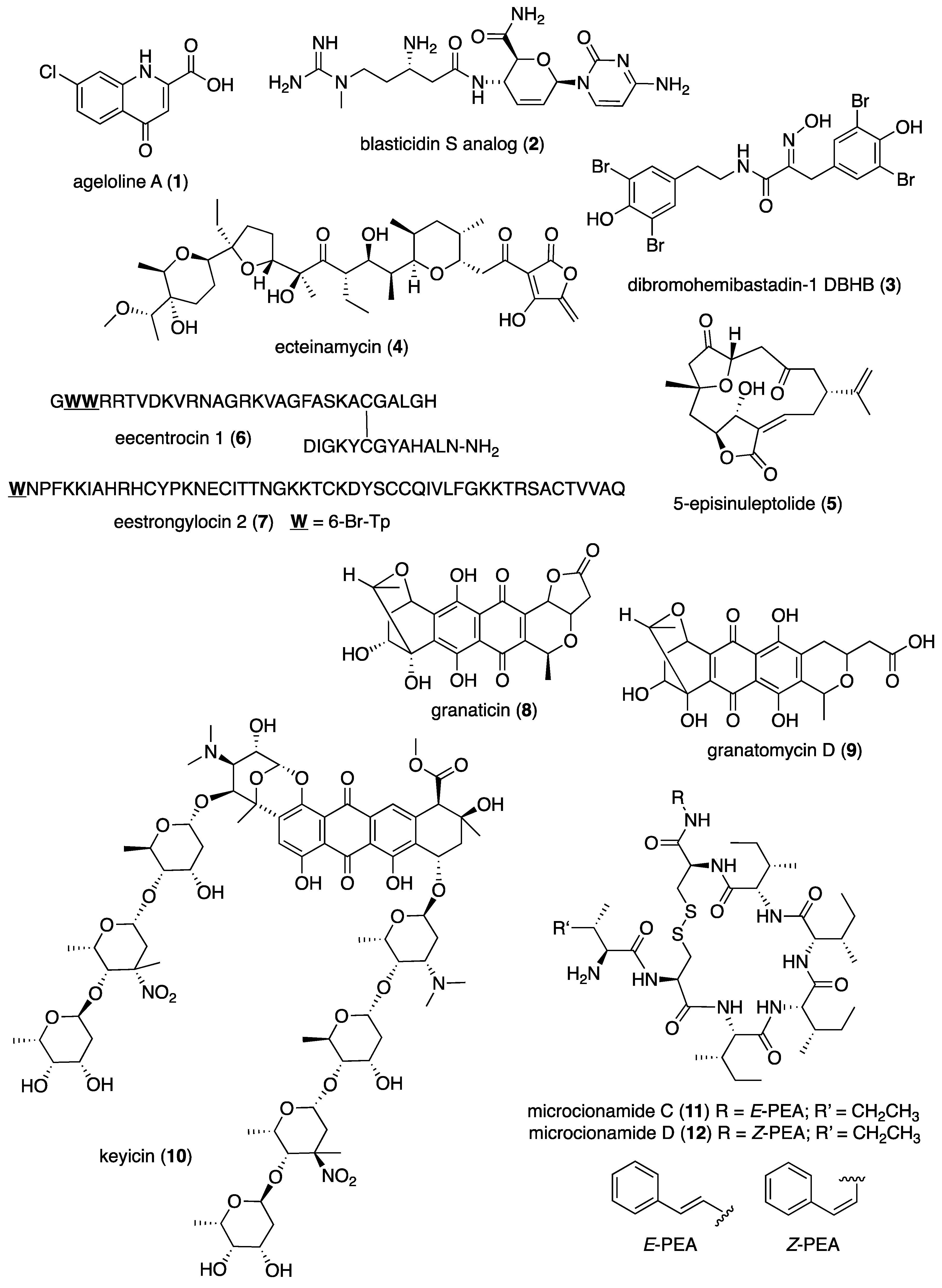
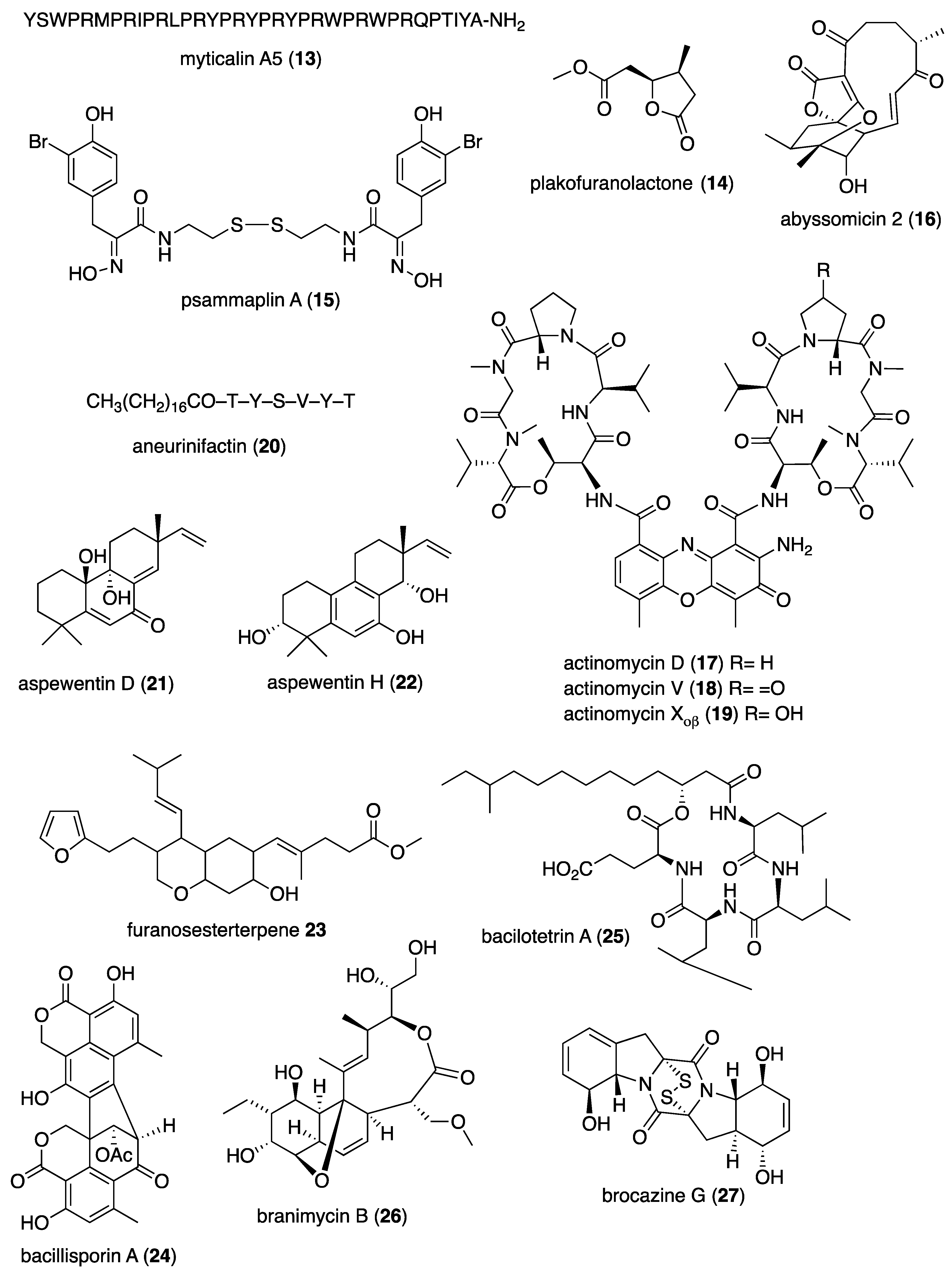
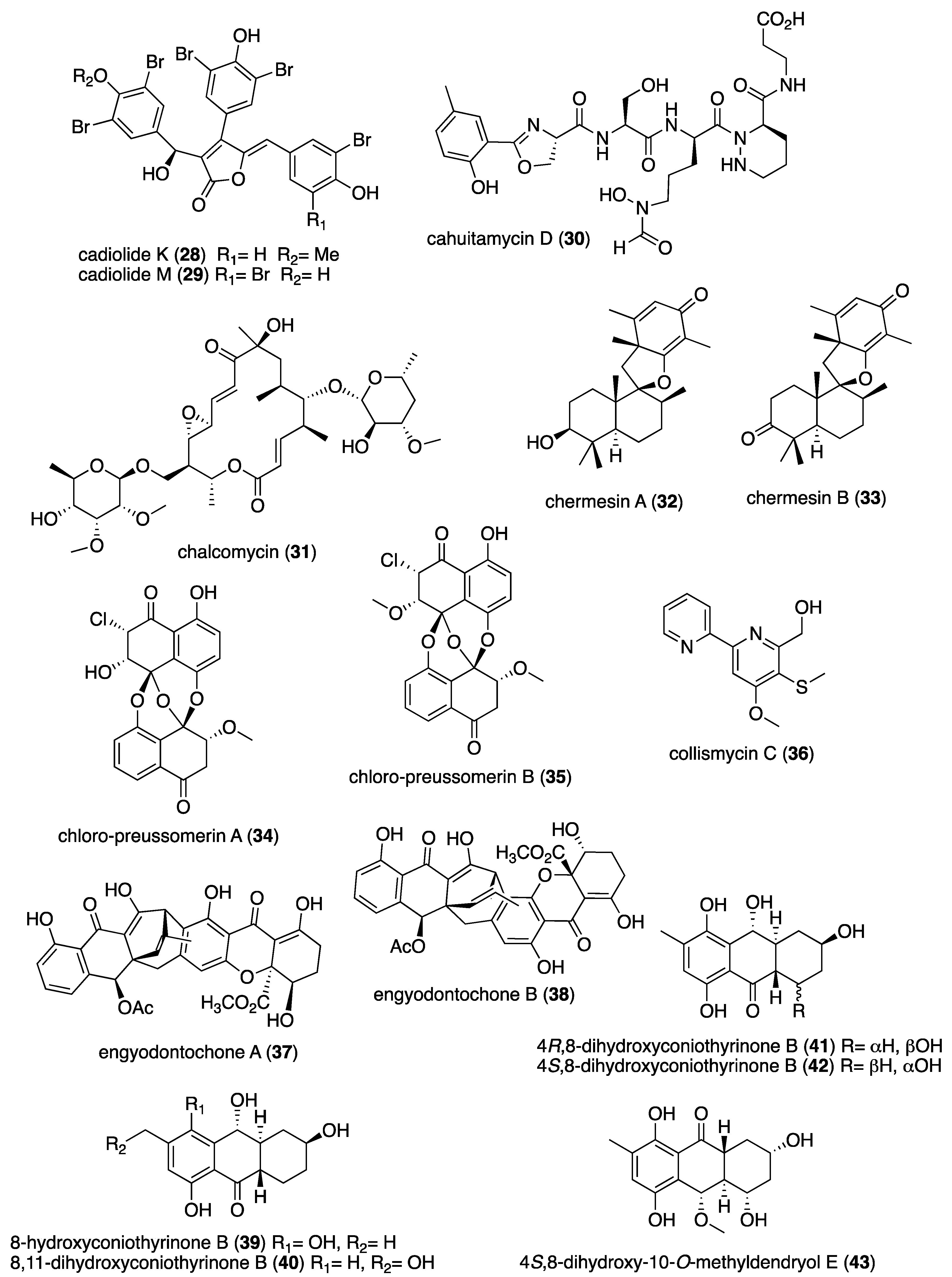
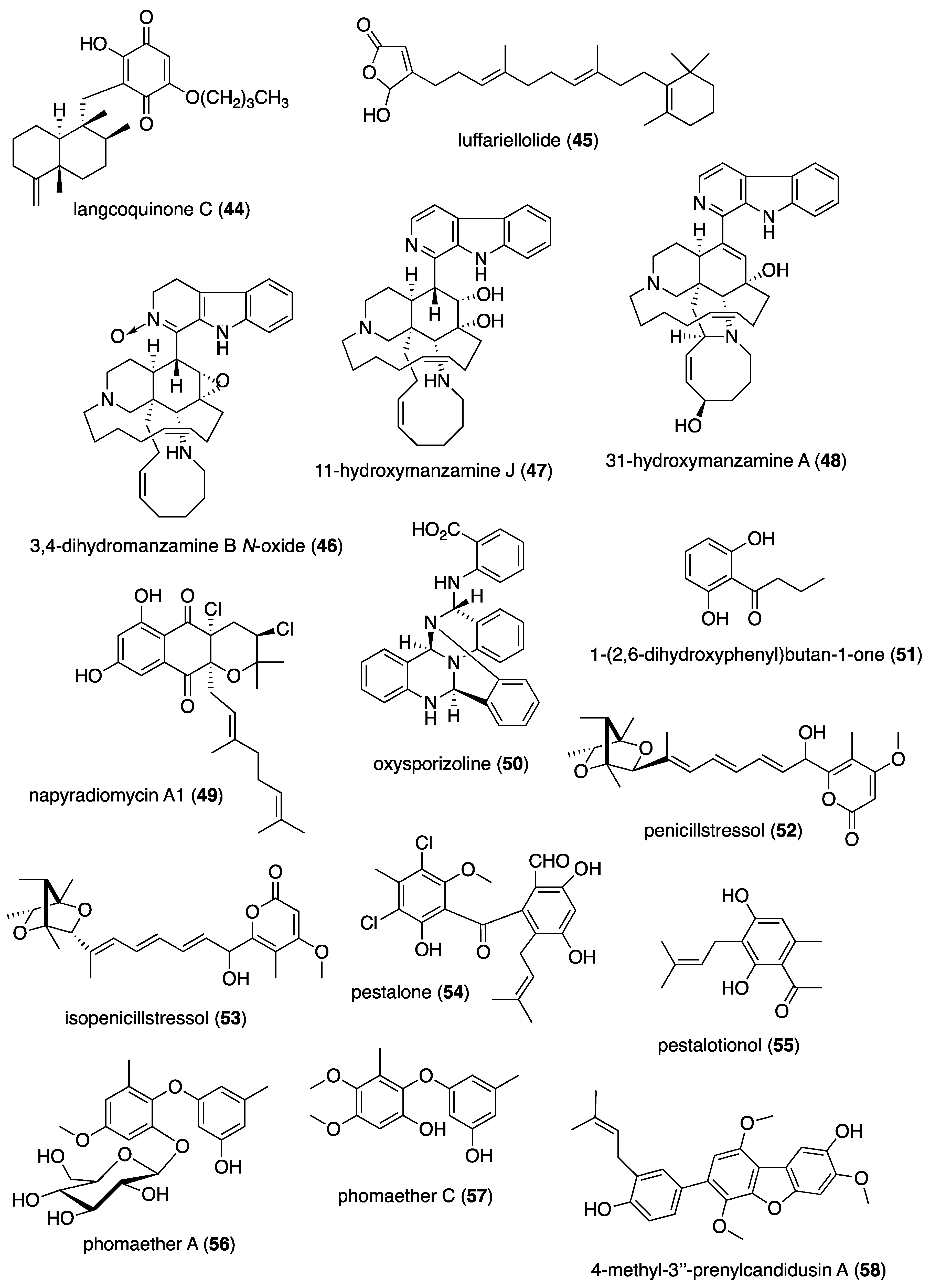
| Antibacterial | ageloline A (1)/bacterium | Alkaloid f | C. trachomatis inhibition | 9.5 μM | Antioxidant activity | DEU | [20] |
| Antibacterial | blasticidin S analog (2)/sponge | Alkaloid f | S. aureus inhibition | 6.2 μg/mL + | norA multidrug trasporter inactivation | CAN, USA | [21] |
| Antibacterial | dibromohemibastadin-1(3)/sponge | Peptide f | P. auruginosa biofilm disruption | 10 μM * | Quorum sensing activity | DEU, FRA, GBR | [22] |
| Antibacterial | ecteinamycin (4)/bacterium | Polyketide d | C. difficile inhibition | 0.059 μM + | K+ transport dysregulation | JPN, USA | [23] |
| Antibacterial | 5-episinuleptolide (5)/soft coral | Terpenoid e | A. baumannii biofilm formation inhibition | 20 μM * | PNAG gene expression inhibition | TWN | [24] |
| Antibacterial | E. esculentus peptides (6,7)/sea urchin | Peptide f | Gram-positive and negative inhibition | 0.1–3.1 μM + | Heavy chains bioactive | NOR, SWE | [25] |
| Antibacterial | granaticin and granatomycin D (8,9)/bacterium | Polyketide d | B. subtilis and MR S. aureus inhibition | 1.6, 6.2 μg/mL + | Co-culture enhanced MIC | USA | [26] |
| Antibacterial | keyicin (10)/bacterium | Polyketide d | B. subtilis and MR S. aureus inhibition | 2.5–9.9 μM + | Fatty acid metabolism modulation | USA | [27] |
| Antibacterial | microcionamides C and D (11, 12)/sponge | Peptide f | S. aureus inhibition | 6.2 μM + | Depolarize cytoplasmic membranes | DEU, IDN, IRN | [28] |
| Antibacterial | myticalin A5 (13)/mussel | Peptide f | Gram-positive and negative inhibition | 2–8 μM + | RNA synthesis inhibition | DEU, ITA | [29] |
| Antibacterial | plakofuranolactone (14)/sponge | Polyketide d | Quorum quenching inhibition | 0.1 μM * | Specificity to QS systems | ITA | [30] |
| Antibacterial | psammaplin A (15)/sponge | Peptide f | V. vulnificus in vivo growth inhibition | 50 μg/mouse ** | Associated in vitro and in vivo pathology suppressed | S. KOR | [31] |
| Antibacterial | abyssomicin 2 (16)/bacterium | Polyketide d | B. thuringiensis and M. luteus inhibition | 3.6, 7.2 μg/mL + | Undetermined | CHN | [32] |
| Antibacterial | actinomycins D, V, and X2 (17–19)/bacterium | Peptide f | MR S. aureus, B. subtilis and E. coli inhibition | 0.08–0.61 μM | Undetermined | CHN, EGY, SAU | [33,34] |
| Antibacterial | aneurinifactin (20)/bacterium | Lipopeptide f | K. pneumoniae and S. aureus inhibition | 4, 8 μg/mL + | Undetermined | IND | [35] |
| Antibacterial | aspewentins D and H (21, 22)/fungus | Terpenoid e | P. aeruginosa and M. luteus inhibition | 4 μg/mL + | Undetermined | CHN | [36] |
| Antibacterial | B. subtilis furanoterpenoid (23)/bacterium | Terpenoid e | V. vulnificus and parahaemolyticus inhibition | 3.12 μg/mL + | Undetermined | IND | [37] |
| Antibacterial | bacillisporin A (24)/fungus | Polyketide d | B. subtilis inhibition | 0.12 μM + | Undetermined | BGD, CHN | [38] |
| Antibacterial | bacilotetrin A (25)/bacterium | Peptide f | MR S. aureus inhibition | 8 μg/mL + | Undetermined | S. KOR | [39] |
| Antibacterial | branimycin B (26)/bacterium | Polyketide d | M. luteus and C. urealyticum inhibition | 1, 8 μg/mL + | Undetermined | ESP | [40] |
| Antibacterial | brocazine G (27)/fungus | Alkaloid f | S. aureus inhibition | 0.25 μg/mL + | Undetermined | CHN, DEU, HUN | [41] |
| Antibacterial | cadiolides K and M (28, 29)/ascidian | Polyketide d | MR S. aureus inhibition | 1–2 μg/mL + | Undetermined | S. KOR | [42] |
| Antibacterial | cahuitamycin D (30)/bacterium | Peptide f | A. baumannii biofilm inhibition | 8.4 μM | Undetermined | CRI, USA | [43] |
| Antibacterial | chalcomycin (31)/bacterium | Polyketide d | S. aureus inhibition | 4 μg/mL + | Undetermined | CHN | [44,45] |
| Antibacterial | chermesins A and B (32, 33)/fungus | Terpenoid e | M. luteus inhibition | 8 μg/mL + | Undetermined | CHN | [46] |
| Antibacterial | chloro-preussomerins A and B (34, 35)/fungus | Polyketide d | S. aureus inhibition | 3.2, 6.2 μg/mL + | Undetermined | CHN | [47] |
| Antibacterial | collismycin C (36)/bacterium | Alkaloid f | MR S. aureus biofilm inhibition | 10 μg/mL * | Undetermined | S. KOR | [48] |
| Antibacterial | engyodontochones A and B (37, 38)/fungus | Polyketide d | MR S. aureus inhibition | 0.17, 0.24 μM | Undetermined | CHN, DEU | [49] |
| Antibacterial | hydroanthraquinones (39–43/fungus | Polyketide d | S. aureus inhibition | 2–8 μg/mL * | Undetermined | CHN, DEU | [50] |
| Antibacterial | langcoquinone C (44)/sponge | Terpenoid e | B. subtilis inhibition | 6.2 μM + | Undetermined | JPN, MMR, VNM | [51] |
| Antibacterial | luffariellolide (45)/sponge | Terpenoid e | S. enterica inhibition | 4 μg/mL + | Undetermined | S. KOR | [52] |
| Antibacterial | manzamine alkaloids (46–48)/sponge | Alkaloid f | Gram-positive and negative inhibition | 2–8 ng/mL + | Undetermined | IDN, S. KOR | [53] |
| Antibacterial | napyradiomycin A1 (49)/bacterium | Terpenoid e | MR S. aureus inhibition | 0.5–1 μg/mL + | Undetermined | ESP | [54] |
| Antibacterial | oxysporizoline (50)/fungus | Alkaloid f | MR S. aureus inhibition | 6.25 μg/mL + | Undetermined | S. KOR | [55] |
| Antibacterial | P. citrinum 1-(2,6-dihydroxyphenyl)butan-1-one (51)/fungus | Polyketide d | S. aureus inhibition | 6.95 μM + | Undetermined | CHN | [56] |
| Antibacterial | penicillstressols (52, 53)/fungus | Polyketide d | MR S. aureus inhibition | 0.5 μg/mL + | Undetermined | CHN | [57] |
| Antibacterial | pestalone (54)/fungus | Polyketide d | MR S. aureus inhibition | 6.25 μM + | Undetermined | CHN | [58] |
| Antibacterial | pestalotionol (55)/fungus | Polyketide d | B. subtilis and S. aureus inhibition | 2, 8 μg/mL + | Undetermined | CHN, TWN | [59] |
| Antibacterial | phomaethers A and C (56, 57)/fungus | Polyketide d | E. coli and S. aureus inhibition | 0.15–1.25 μM + | Undetermined | CHN | [60] |
| Antibacterial | 4-methyl-3”-prenylcandidusin A (58)/fungus | Polyketide d | MR S. aureus and V. vulnificus inhibition | 3.8, 7.8 μg/mL + | Undetermined | CHN | [61] |
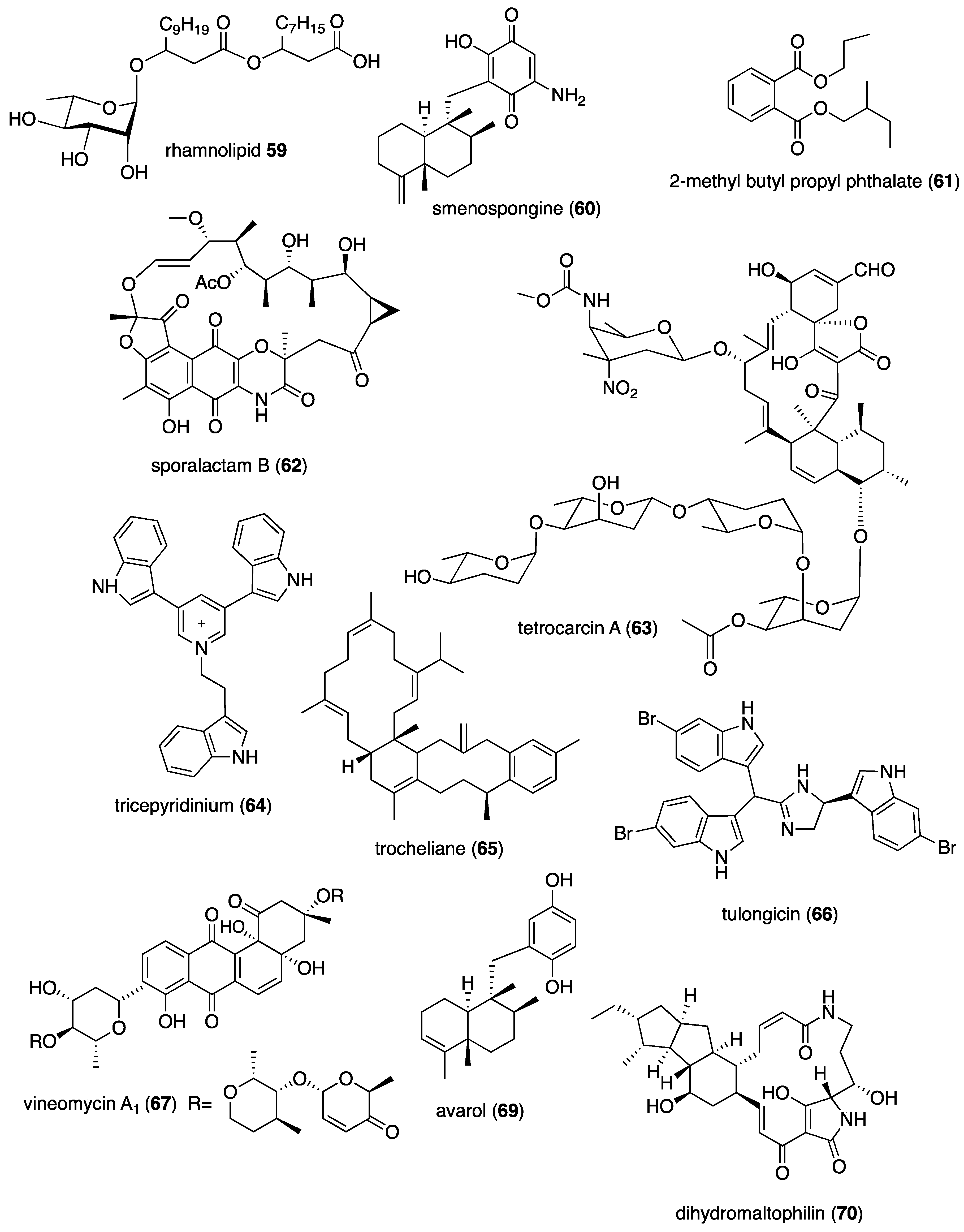
| Antibacterial | Pseudomonas sp. rhamnolipid (59)/bacterium | Lipid e | B. cenocepacia and S. aureus inhibition | 1.6–3.1 μg/mL + | Undetermined | GBR, ITA | [62] |
| Antibacterial | smenospongine (60)/sponge | Terpenoid e | B. cereus and S. aureus inhibition | 3.1 μM + | Undetermined | CHN, USA | [63] |
| Antibacterial | S. cheonanensis phthalate (61)/bacterium | Polyketide d | P. vulgaris inhibition | 4 μg/mL + | Undetermined | IND | [64] |
| Antibacterial | sporalactam B (62)/bacterium | Polyketide d | MR S. aureus and E. coli inhibition | 0.4–1.8 μM ** | Undetermined | CAN, PHL | [65] |
| Antibacterial | tetrocarcin A (63)/fungus | Polyketide d | B. subtilis inhibition | 0.03–0.125 μg/mL + | Undetermined | CHN | [66,67] |
| Antibacterial | tricepyridinium (64)/sponge | Alkaloid f | B. subtilis and S. aureus inhibition | 0.78–1.56 μg/mL + | Undetermined | JPN | [68] |
| Antibacterial | trocheliane (65)/soft coral | Terpenoid e | A. baumannii and S. aureus inhibition | 4–4.2 μM + | Undetermined | EGY, IDN, SAU | [69] |
| Antibacterial | tulongicin (66)/sponge | Alkaloid f | S. aureus inhibition | 1.2 μg/mL + | Undetermined | ITA, NZL, USA | [70] |
| Antibacterial | vineomycin A1 (67)/bacterium | Polyketide d | S. aureus inhibition | 4 μg/mL+ | Undetermined | CHN | [71] |
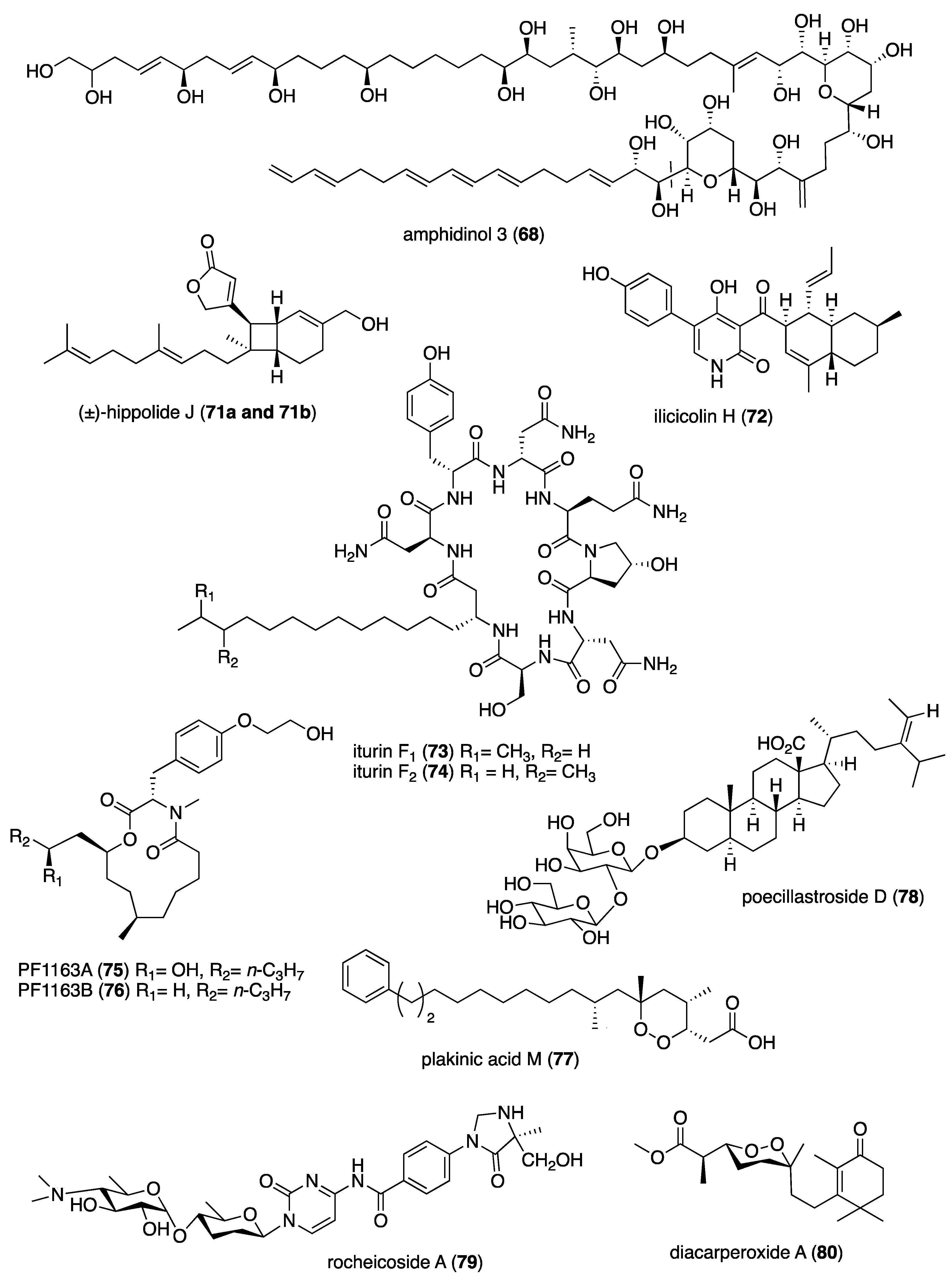
| Antifungal | amphidinol 3 (68)/dinoflagellate | Polyketide d | Pore formation requires cholesterol or ergosterol | 2.0 μM * | Toroidal pore 2.6-4.0 nM | JPN, PHL | [72,73] |
| Antifungal | avarol (69)/sponge | Terpenoid e | C. albicans inhibition | 6–8 µg/mL + | Undetermined | SRB | [74] |
| Antifungal | dihydromaltophilin (70)/bacterium | Polyketide d | C. albicans inhibition | 3 μM | Undetermined | AUS, MEX | [75] |
| Antifungal | hippolide j (71a, 71b)/sponge | Terpenoid e | C. albicans inhibition | 0.1 µg/mL + | Undetermined | CHN, GBR | [76] |
| Antifungal | ilicicolin H (72)/fungus | Polyketide d | C. albicans inhibition | <0.25 µg/mL + | Undetermined | DEU, DNK, ESP | [77] |
| Antifungal | iturin F1 and F2 (73, 74)/bacterium | Peptide f | A. flavus and P. griseofulvum inhibition | 3.1 µg/mL + | Undetermined | JPN, S. KOR | [78] |
| Antifungal | P. meleagrinum macrolides (75, 76)/fungus | Polyketide d | C. albicans inhibition | 1–2 μg/mL ** | Undetermined | JPN | [79] |
| Antifungal | plakinic acid M (77)/sponge | Polyketide d | C. gattii inhibition | 2.4 μM ** | Undetermined | USA | [80] |
| Antifungal | poecillastroside D (78)/sponge | Terpenoid e | A. fumigatus inhibition | 6 μg/mL ** | Undetermined | ESP, FRA, IRL, OMN, SWE | [81] |
| Antifungal | rocheicoside A (79)/ bacterium | Alkaloid f | C. albicans inhibition | 4 μg/mL + | Undetermined | TUR | [82] |
| Antimalarial | diacarperoxide A (80)/sponge | Terpenoid e | P. falciparum D6 and W2 strain inhibition | 1.9–2.0 μM | Undetermined | CHN, USA | [83] |
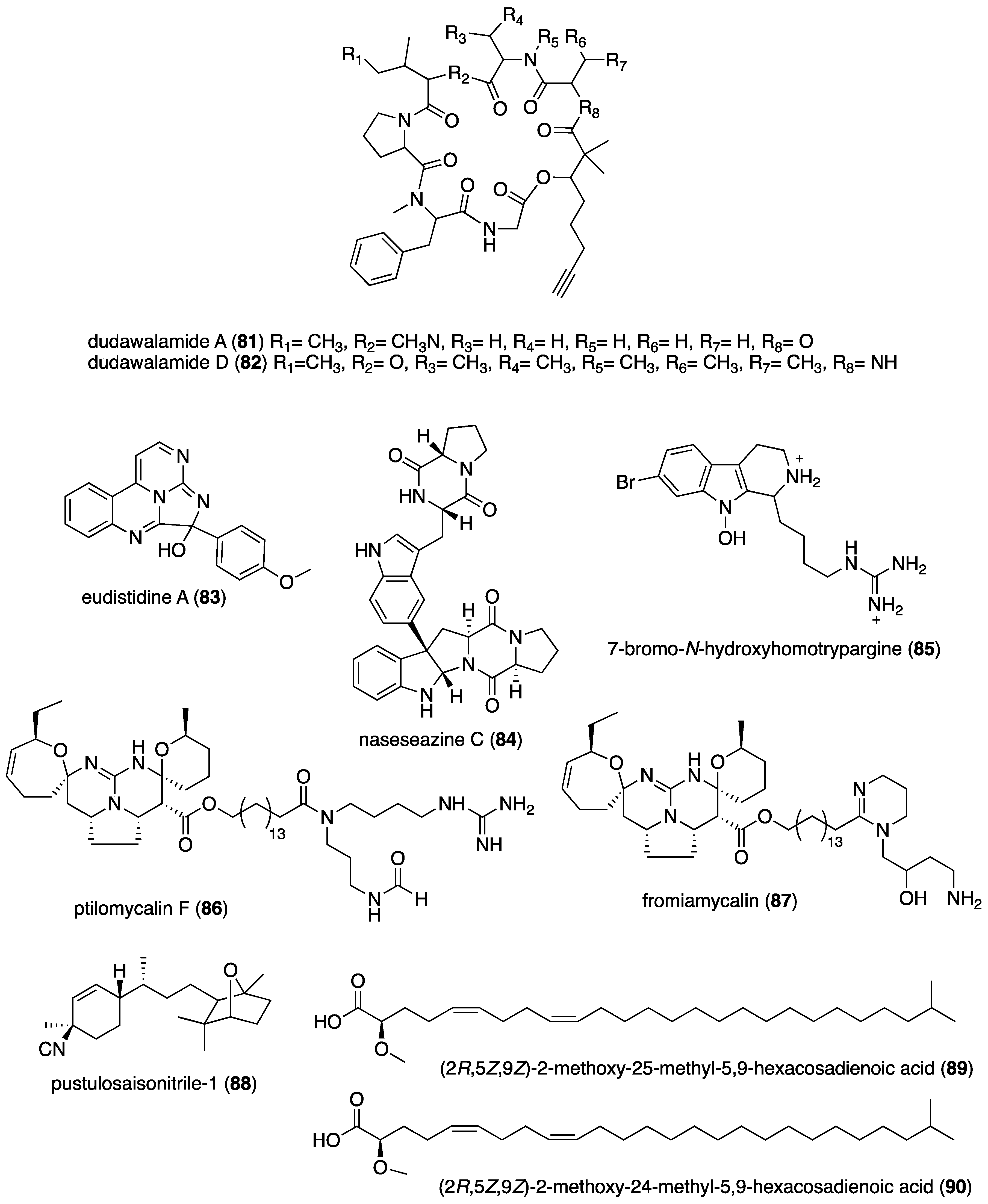
| Antimalarial | dudawalamide A and D (81, 82)/cyanobacterium | Peptide f | P. falciparum W2 strain inhibition | 3.5 μM | Undetermined | JOR, PAN, USA | [84] |
| Antimalarial | eudistidine A (83)/soft coral | Alkaloid f | P. falciparum D6 and W2 strain inhibition | 1.1–1.4 μM | Undetermined | CAN, USA | [85] |
| Antimalarial | naseseazine C (84)/bacterium | Alkaloid f | P. falciparum 3D7 inhibition | 3.5 μM | Undetermined | AUS | [86] |
| Antimalarial | P. opacum β-carboline (85)/ascidian | Alkaloid f | P. falciparum FcB1inhibition | 3.8 μM | Undetermined | FRA, NZL | [87] |
| Antimalarial | ptilomycalin F (86)/sponge | Alkaloid f | P. falciparum 3D7 strain inhibition | 0.23 μM | Undetermined | BEL, FRA, CHE, NLD | [88] |
| Antimalarial | pustulosaisonitrile-1 (88)/nudibranch | Terpenoid e | P. falciparum 3D7 strain inhibition | 1.08 μM | Undetermined | AUS, USA | [89] |
| Antileishmanial | A. Niger fatty acids (89, 90)/sponge | Lipid e | L. infantum inhibition | 0.17, 0.34 mg/mL | TopIB inhibition | ESP, USA | [90] |
| Antileishmanial | dudawalamide D (82)/cyanobacterium | Peptide f | L. donovani inhibition | 2.6 μM | Undetermined | JOR, PAN, USA | [84] |
| Antileishmanial | Gorgonia sp. sterol (91) sponge | Terpenoid e | L. infantum inhibition | >10 μM * | Undetermined | ESP, PAN | [91] |
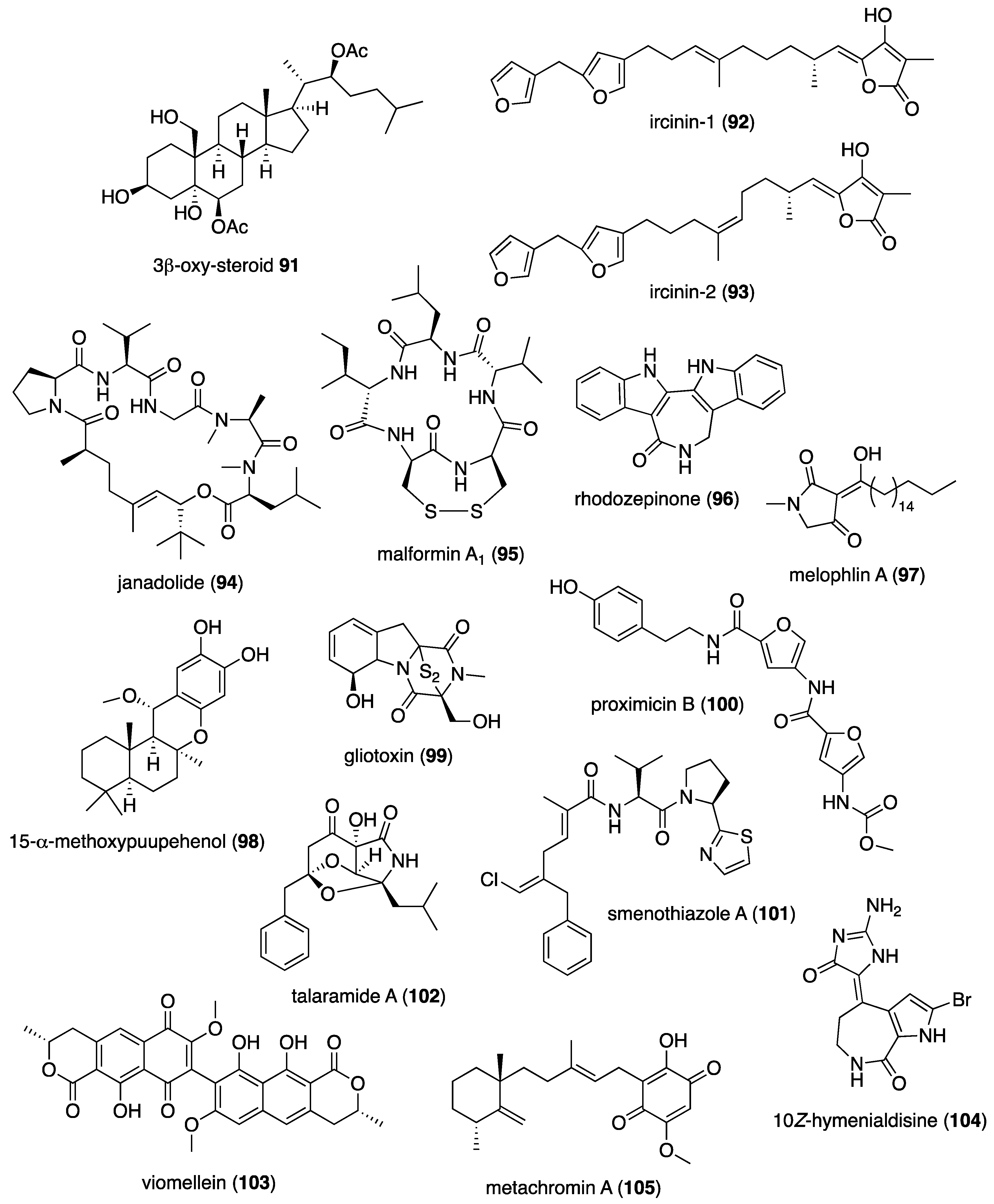
| Antileishmanial | ircinin-1 and 2 (92, 93)/sponge | Terpenoid e | L. donovani inhibition | 28–31 μM | Undetermined | CHE, DEU, ITA, TUR | [92] |
| Antitrypanosomal | janadolide (94)/cyanobacterium | Peptide f | T. b. brucei inhibition | 0.047 μM | Undetermined | JPN | [93] |
| Antitrypanosomal | malformin A1 (95)/fungus | Peptide f | T. congolense inhibition | 0.015 µg/mL | Undetermined | JPN, PHL | [94] |
| Antitrypanosomal | rhodozepinone (96)/bacterium | Alkaloid f | T. b. brucei inhibition | 16.3 µg/mL | Undetermined | DEU, EGY | [95] |
| Antituberculosis | melophlin A (97)/sponge | Alkaloid f | M. smegmatis inhibition | 0.8 μg/mL + | BCG1083 & BCG1321c proteins targeted | IDN, JPN | [96] |
| Antituberculosis | methoxypuupehenol (98)/sponge | Terpenoid e | Dormant M. tuberculosis inhibition | 0.5 μg/mL + | Bactericidal activity | USA | [97] |
| Antituberculosis | gliotoxin (99)/fungus | Alkaloid f | M. tuberculosis inhibition | 0.03 μM + | Undetermined | CHN | [98] |
| Antituberculosis | proximicin B (100)/bacterium | Peptide f | M. bovis Pasteur 1173P2 inhibition | 6.25 μg/mL + | Undetermined | AUS, CHN, EGY, NGA | [99] |
| Antituberculosis | smenothiazole A (101)/sponge | Peptide f | M. tuberculosis H37Rv inhibition | 4.1 μg/mL + | Undetermined | POL, USA | [100] |
| Antituberculosis | sporalactam B (62)/bacterium | Polyketide d | M. tuberculosis inhibition | 0.06 μM ** | Undetermined | CAN, PHL | [65] |
| Antituberculosis | talaramide A (102)/fungus | Alkaloid f | Mycobacterial PknG inhibition | 55 μM | Undetermined | CHN | [101] |
| Antituberculosis | viomellein (103)/fungus | Polyketide d | Dormant M. bovis BCG inhibition | 1.56 μg/mL + | Undetermined | IDN, JPN | [102] |
| Antiviral | hymenialdisine (104)/sponge | Alkaloid f | HIV-1 inhibition | >3.1 μM * | Reverse transcriptase inhibition | DEU, SAU, USA | [103] |
| Antiviral | metachromin A (105)/sponge | Terpenoid e | HBV inhibition | 0.8 μM | Viral promoter inhibition | JPN, NLD | [104] |
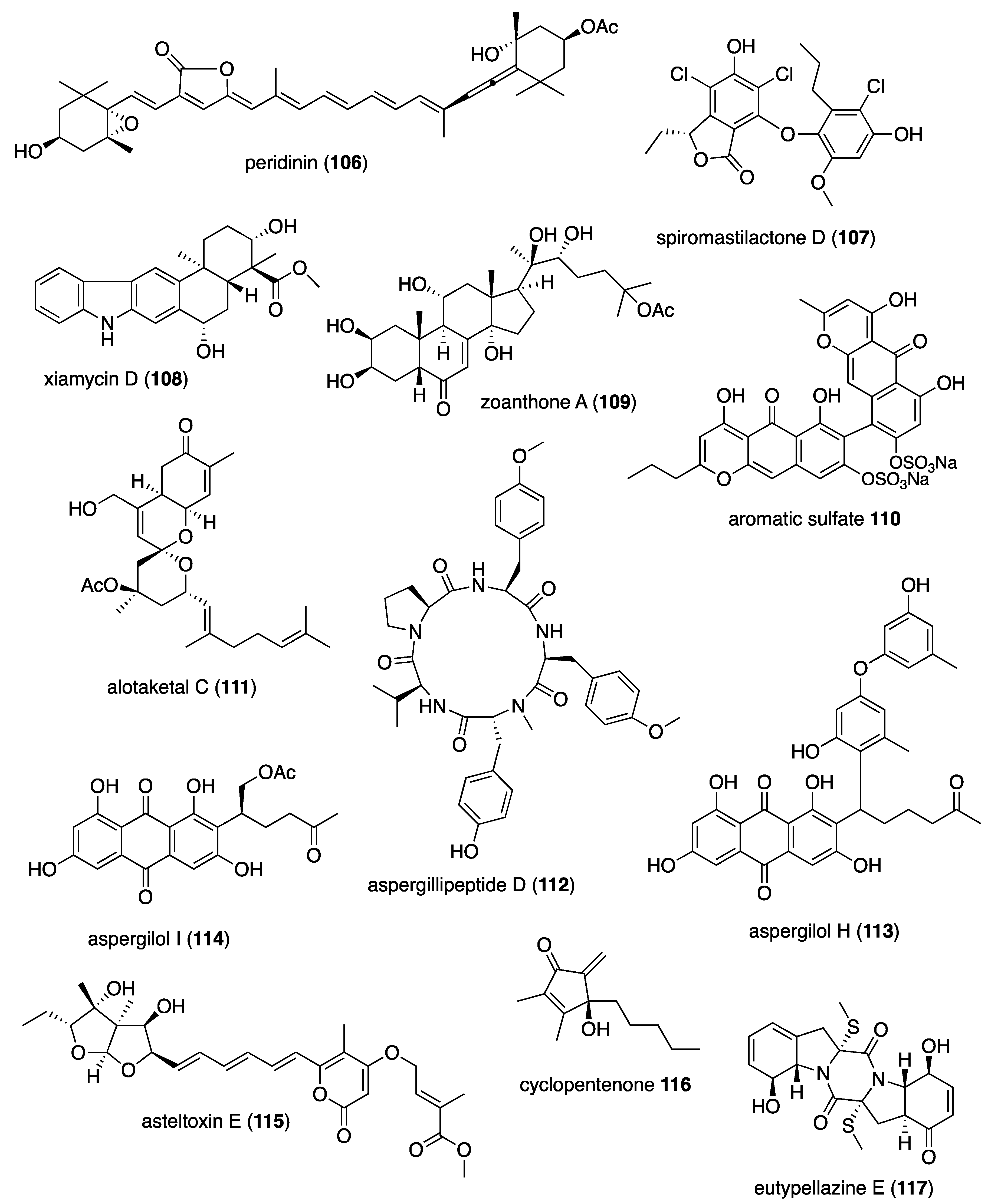
| Antiviral | peridinin (106)/coral | Terpenoid e | HTLV-1 infected T cell inhibition | 0.7-5.4 μM | NF-κB inhibition | JPN | [105] |
| Antiviral | spiromastilactone D (107)/fungus | Polyketide d | H1N1 influenza A virus inhibition | 6.0 μM | HA-sialic acid receptor binding inhibition | CHN, USA | [106] |
| Antiviral | xiamycin D (108)/bacterium | Terpenoid e | PEDV virus inhibition | 0.93 μM | Virion structural proteins inhibition | S. KOR | [107] |
| Antiviral | zoanthone A (109)/sea anemone | Terpenoid e | DENV-2 virus inhibition | 19.6 μM | RNA pocket tunnel binding | TWN | [108] |
| Antiviral | A. polycladia aromatic sulfate (110)/crinoid | Polyketide d | HCV NS3 helicase inhibition | 5 μM | Undetermined | JPN | [109] |
| Antiviral | alotaketal C (111)/sponge | Terpenoid e | HIV expression activation | 1 μM * | Undetermined | CAN | [110] |
| Antiviral | aspergillipeptide D (112)/fungus | Peptide f | HSV-1 inhibition | 9.5 μM | Undetermined | CHN | [111] |
| Antiviral | aspergilols H and I (113, 114)/fungus | Polyketide d | HSV-1 inhibition | 4.7, 6.2 μM | Undetermined | CHN | [112] |
| Antiviral | asteltoxin E (115)/fungus | Polyketide d | H1N1 and H3N2 influenza virus inhibition | 3.5, 6.2 μM | Undetermined | CHN | [113] |
| Antiviral | S. verruca cyclopentenone (116)/soft coral | Polyketide d | HIV infection inhibition | 5.8 μM | Undetermined | CHN, USA | [114] |
| Antiviral | eutypellazine E (117)/fungus | Alkaloid f | HIV-1 inhibition | 3.2 μM | Undetermined | CHN, DEU | [115] |
| Antiviral | ω-hydroxyemodin (118)/fungus | Polyketide d | HCV NS3 protease inhibition | 10.7 μM | Undetermined | EGY, SAU | [116] |
| Antiviral | malformin C (119)/fungus | Peptide f | HIV infection inhibition | 1.4 μM | Undetermined | CHN | [117] |
| Antiviral | manzamine A (120)/sponge | Alkaloid f | HSV-1 inhibition | 1 μM + | Undetermined | USA | [118] |
| Antiviral | peridinin (106)/zoanthid | Terpenoid e | Anti-dengue virus 2 inhibition | 4.5 μM | Undetermined | TWN | [119] |
| Antiviral | stachybonoid A (121)/fungus | Terpenoid e | Dengue virus prM protein expression inhibition | 25 μM | Undetermined | CHN | [120] |
| Antiviral | subergorgols T and U (122, 123)/soft coral | Terpenoid e | H1N1 influenza A virus inhibition | 35–37 μM | Undetermined | CHN, NLD | [121] |
2. Marine Compounds with Antibacterial, Antifungal, Antiprotozoal, Antituberculosis and Antiviral Activities










2.1. Antibacterial Activity
2.2. Antifungal Activity
2.3. Antiprotozoal and Antituberculosis Activity
2.4. Antiviral Activity
3. Marine Compounds with Antidiabetic and Anti-Inflammatory Activity, and Affecting the Immune and Nervous System
| Drug Class | Compound/ Organism a | Chemistry | Pharmacological Activity | IC50 b | MMOA c | Country d | References |
|---|---|---|---|---|---|---|---|
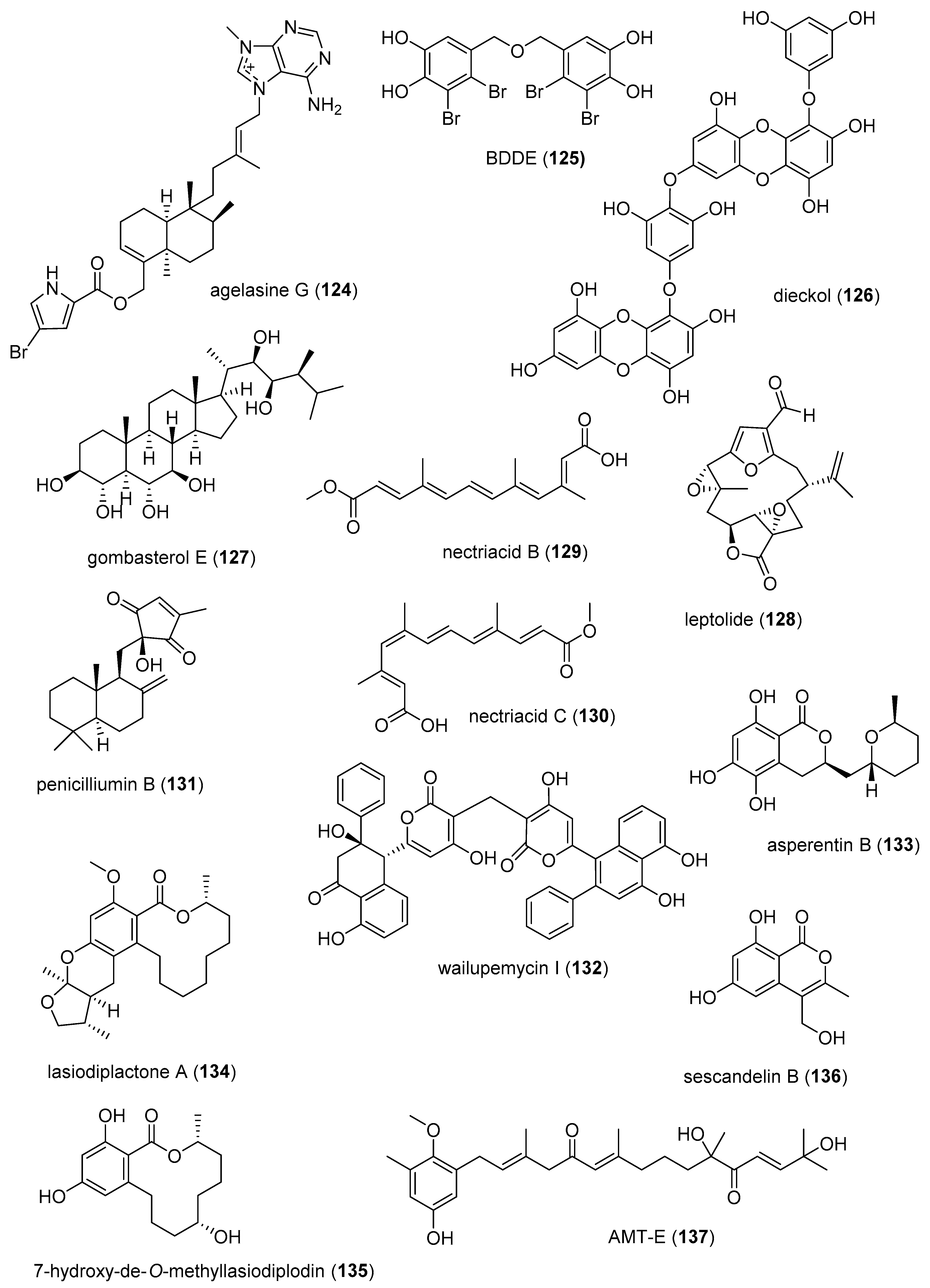
| Antidiabetic | agelasine G (124)/sponge | Alkaloid-terpenoid f | PTP1B inhibition | 15 µM * | Akt insulin pathway increase | IDN, JAP | [151] |
| Antidiabetic | BDDE (125)/alga | Shikimate h | Decrease glucose levels in vivo | 10 mg/kg ** | PTP1B expression inhibition | CHN | [152] |
| Antidiabetic | dieckol (126)/alga | Shikimate h | Decrease in glucose levels | 1 µg/g ** | Akt insulin pathway increase | S. KOR | [153] |
| Antidiabetic | gombasterol E (127)/sponge | Terpenoid f | Enhanced glucose uptake in vitro | 20 µM * | AMPK phosphorylation increase | S. KOR | [154] |
| Antidiabetic | leptolide (128)/soft coral | Terpenoid f | Murine glucose tolerance and insulin sensitivity increased | 0.1 mg/kg ** | PKB phosphorylation | ESP | [155] |
| Antidiabetic | nectriacids B and C (129, 130)/fungus | Polyketides d | α-glucosidase inhibition | 23.5, 42.3 µM | C-12 carboxyl esterification required | CHN | [156] |
| Antidiabetic | penicilliumin B (131)/fungus | Terpenoid f | Glomerular mesangial cells fibrogenic inhibition | 0.5 µM * | NADPH oxidase inhibition | CHN | [157] |
| Antidiabetic | wailupemycin I (132)/bacterium | Polyketide d | α-glucosidase inhibition | 8.3 µM | Competitive inhibition | CHN | [158] |
| Antidiabetic | asperentin B (133)/fungus | Polyketide d | PTP1B inhibition | 2 μM | Undetermined | DEU | [159] |
| Antidiabetic | lasiodiplactone A (134)/fungus | Polyketide d | α-glucosidase inhibition | 29.4 μM | Undetermined | CHN | [160] |
| Antidiabetic | 7-hydroxy-de-O-methyllasiodiplodin (135)/bacterium | Polyketide d | α-glucosidase inhibition | 25.8 μM | Undetermined | CHN | [161] |
| Antidiabetic | sescandelin B (136)/fungus | Polyketide d | α-glucosidase inhibition | 17.2 μM | Undetermined | CHN | [162] |
| Anti-inflammatory | AMT-E (127)/alga | Terpenoid f | Murine colitis inhibition | 10 mg/kg ** | Inhibition of TNF-α, IL-6 | ESP, MAR | [163] |
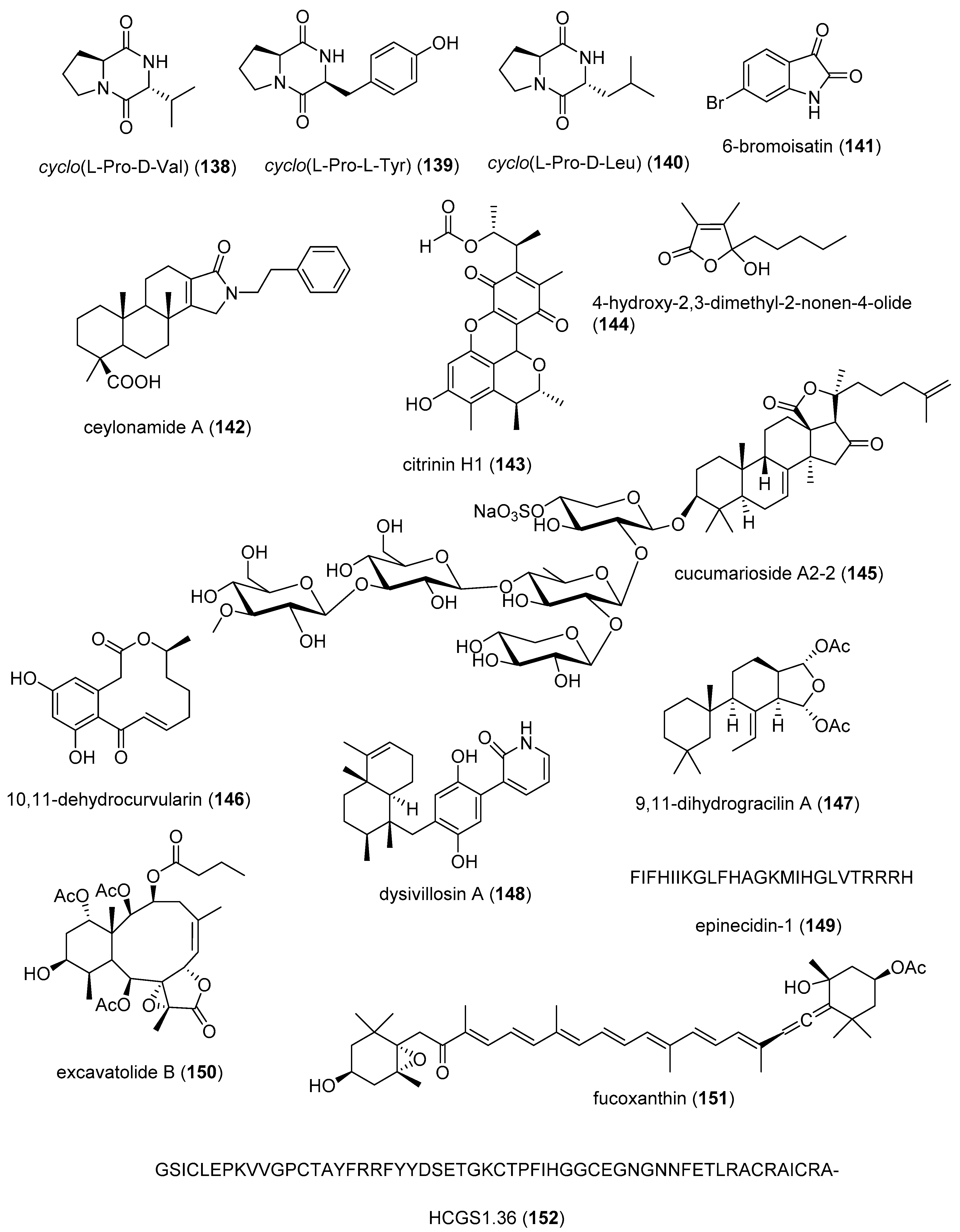
| Anti-inflammatory | Bacillus sp. diketopiperazines (138–140)/bacterium | Peptide g | TGFBIp inhibition in vivo | 5 μM ** | Septic responses inhibition | S. KOR | [164,165] |
| Anti-inflammatory | 6-bromoisatin (141)/mollusc | Alkaloid g | Lung inflammation inhibition in vivo | 0.05 mg/g ** | Inhibition of TNF-α, IL-6 | AUS | [166] |
| Anti-inflammatory | ceylonamide A (142)/sponge | Terpenoid f | Macrophage RANKL inhibition | 13 μM * | SAR completed | IDN, JPN, NLD | [167] |
| Anti-inflammatory | citrinin H1 (143)/fungus | Polyketide e | Microglia NO and PGE2 release inhibition | 8 μM | NF-κB inhibition | S. KOR, VNM | [168] |
| Anti-inflammatory | nonenolide derivative (144)/alga | Polyketide e | BMDC cytokine release inhibition | 7.6–10.9 μM | JNK, ERK, AP-1, NF-κB inhibition | S. KOR | [169] |
| Anti-inflammatory | cucumarioside A2-2 (145)/sea cucumber | Terpenoid f | Binding of macrophage P2X purinergic receptors | 0.02 μM * | Induction Ca2+ oscillations | RUS | [170] |
| Anti-inflammatory | curvularin derivative (146)/fungus | Polyketide e | Macrophage PGE2 and NO release inhibition | 1.9–2.7 μM | NF-κB signaling inhibition | S. KOR | [171] |
| Anti-inflammatory | 9,11-dihydrogracilin A (147)/sponge | Terpenoid f | PBMC proliferation inhibition | 3 μM * | IL-6 and IL-10 inhibition | ITA | [172] |
| Anti-inflammatory | dysivillosin A (148)/sponge | Terpenoid f | Basophil β-hexosaminidase inhibition | 8.2 μM | IL-4 and LTB4 inhibition | CHN | [173] |
| Anti-inflammatory | epinecidin-1 (149)/fish | Peptide g | Inhibition of MyD88 protein levels | 6 µg/mL * | Proteasome degradation required | TWN | [174] |
| Anti-inflammatory | excavatolide B (150)/soft coral | Terpenoid f | Attenuation of rat arthritis [175] | 2.5, 5 mg/kg ** | Decreased MMP-2, MMP-9, CD11b in tissues | CHN, TWN | [175] |
| Anti-inflammatory | fucoxanthin (151)/alga | Terpenoid f | Decreased mice paw edema, adipogenesis and ear inflammation | 4 mg/kg ** | Modulation of iNOS, PLA2, COX-2, ACC, IL-6 and Nrf2 expression | JPN, S. KOR, MEX | [176,177,178,179] |
| Anti-inflammatory | H. crispa peptide (152)/sea anemone | Peptide g | Macrophage histamine receptor inhibition | 10 μM * | Intracellular Ca2+ increase inhibition | RUS | [180] |
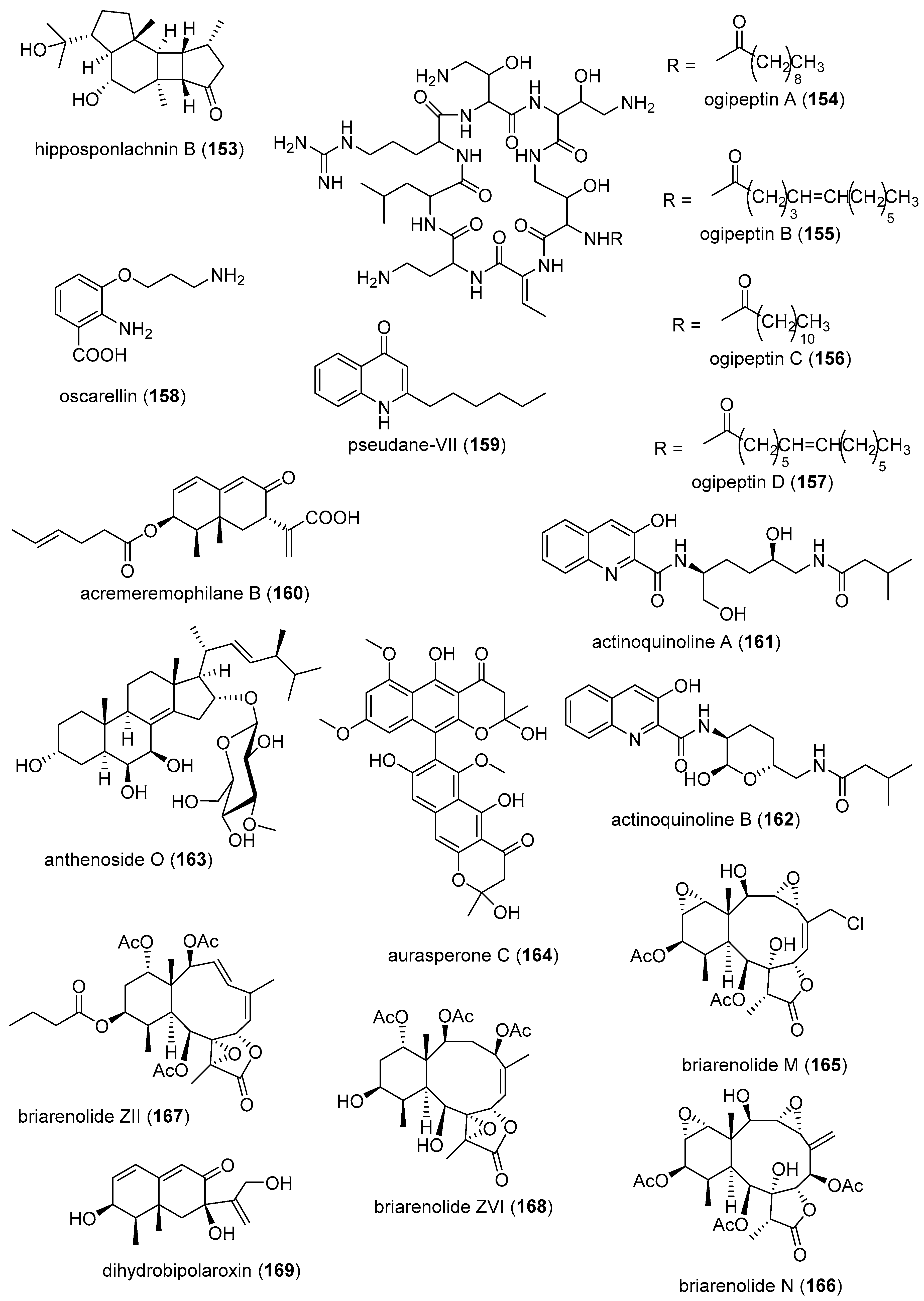
| Anti-inflammatory | hipposponlachnin B (153)/sponge | Terpenoid f | Basophil β-hexosaminidase inhibition | 24 μM | IL-4 and LTB4 inhibition | CHN | [181] |
| Anti-inflammatory | ogipeptins A-D (154–157)/bacterium | Peptide g | Macrophage TNF-α production inhibition | 1 µM * | Block LPS binding to CD14 | JPN | [182] |
| Anti-inflammatory | oscarellin (158)/sponge | Alkaloid g | Macrophage TNF-α and IL-6 expression inhibition | >10 μM | JNK, ERK, AP-1, NF-κB inhibition | S. KOR, USA | [183] |
| Anti-inflammatory | pseudane-VIII (159)/bacterium | Alkaloid g | Macrophage NO release inhibition | 6 µM * | iNOS and IL-1β inhibition | S. KOR | [184] |
| Anti-inflammatory | acremeremophilane B (160)/fungus | Terpenoid f | Macrophage NO release inhibition | 8 µM | Undetermined | CHN, DEU | [185] |
| Anti-inflammatory | actinoquinolines A and B (161, 162)/bacterium | Alkaloid g | COX-1 and -2 inhibition | 1.4–7.6 μM | Undetermined | EGY, USA | [186] |
| Anti-inflammatory | anthenoside O (163)/starfish | Terpenoid f | Macrophage SOX inhibition | >10 µM * | Undetermined | RUS, VNM | [187] |
| Anti-inflammatory | aurasperone C (164)/fungus | Polyketide e | COX-2 inhibition | 4.2 μM | Undetermined | CHN | [188] |
| Anti-inflammatory | briarenolides M and N (165, 166)/soft coral | Terpenoid f | Macrophage iNOS expression inhibition | 10 μM * | Undetermined | TWN | [189] |
| Anti-inflammatory | briarenolides ZII and ZVI (167, 168)/soft coral | Terpenoid f | Macrophage iNOS and COX-2 expression inhibition | 10 μM * | Undetermined | TWN | [190] |
| Anti-inflammatory | dihydrobipolaroxin (169)/fungus | Terpenoid f | Macrophage NO release inhibition | >12.5 μM * | Undetermined | CHN | [191] |
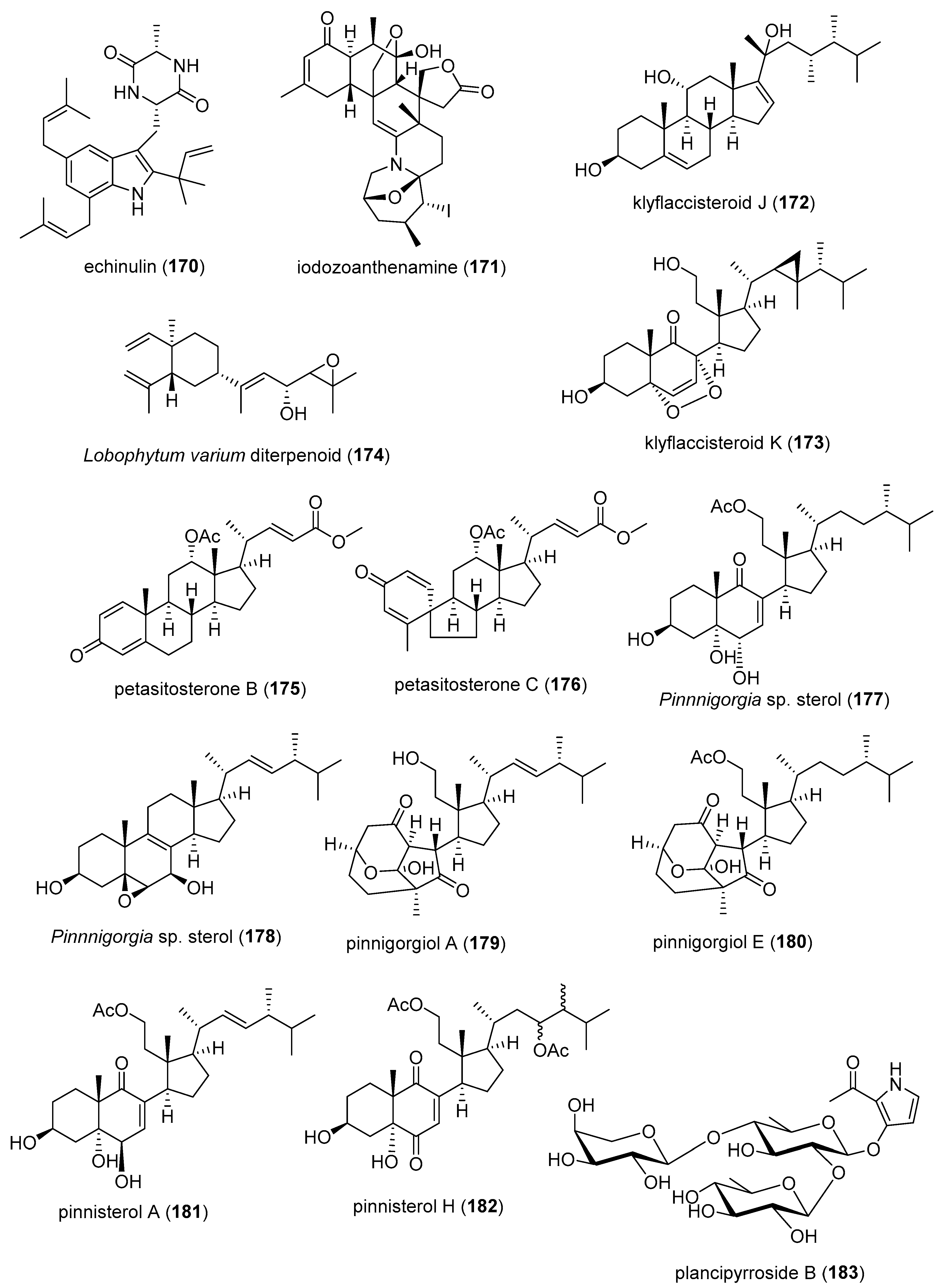
| Anti-inflammatory | echinulin (170)/fungus | Alkaloid g | Microglia NO release inhibition | 4.6 μM | Undetermined | CHN, S. KOR | [192] |
| Anti-inflammatory | 5α-iodozoanthenamine (171)/zoanthid | Alkaloid g | Neutrophil SOX and elastase inhibition | >10 μM * | Undetermined | TWN | [193] |
| Anti-inflammatory | klyflaccisteroid J and K (172, 173)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 1.5–5.8 μM | Undetermined | TWN | [194,195] |
| Anti-inflammatory | L. varium diterpenoid (174)/soft coral | Terpenoid f | Neutrophil elastase inhibition | >10 μM * | Undetermined | EGY, SAU, TWN | [196] |
| Anti-inflammatory | petasitosterones B and C (175, 176)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 2.7–4.4 µM | Undetermined | TWN | [197] |
| Anti-inflammatory | Pinnnigorgia sp. sterols (177, 178)/soft coral | Terpenoid f | Macrophage COX-2 and iNOS expression inhibition | 10 µM * | Undetermined | TWN | [198] |
| Anti-inflammatory | pinnigorgiol A (179)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 4, 5 µM | Undetermined | TWN | [199] |
| Anti-inflammatory | pinnigorgiol E (180)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 1.6, 3.9 µM | Undetermined | TWN | [200] |
| Anti-inflammatory | pinnisterols A and H (181, 182)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 2.3–3.3 µM | Undetermined | TWN | [201,202] |
| Anti-inflammatory | plancipyrroside B (183)/starfish | Terpenoid f | Macrophage iNOS expression inhibition | 5.9 µM | Undetermined | RUS, VNM | [203] |
| Anti-inflammatory | protolinckioside A (184)/starfish | Terpenoid f | Macrophage SOX inhibition | 10 µM | Undetermined | IND, RUS | [204] |
| Anti-inflammatory | sarcophytonolide O (185)/soft coral | Terpenoid f | Macrophage iNOS expression inhibition | 8 μM | Undetermined | CHN, USA | [205] |
| Anti-inflammatory | sinularectols A and B (186, 187)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 0.9–8.5 μM | Undetermined | TWN | [206] |
| Anti-inflammatory | sinubrasolides A and D (188, 189)/soft coral | Terpenoid f | Neutrophil SOX and elastase inhibition | 1.4–8 μM | Undetermined | CAN, SAU, TWN | [207,208] |
| Anti-inflammatory | uprolides N, O and P (190–192)/soft coral | Terpenoid f | Macrophage TNF-α and IL-6 release inhibition | 1.4–4.2 μM | Undetermined | IND, PAN, USA | [209] |
| Immune system | cucumarioside A2-2 (145)/sea cucumber | Terpenoid f | Increase in spleen white pulp and macrophage activation | 3 mg/kg ** | Increased B cell PCNA and M1 macrophages | RUS, TWN | [210,211] |
| Immune system | gracilins A, H and L (193–195)/sponge | Terpenoid f | CD147 receptor modulation and T-cell IL-2 release inhibition | 1 μM * | Hypersensitivity and NFATc inhibition | ESP, GBR | [212,213] |
| Immune system | shinorine and porphyra-334 (196, 197)/alga | Peptide g | NF-κB stimulation | 50 µg/mL * | Tryptophan metabolism modulation | AUT | [214] |
| Immune system | sinulariolide (198)/soft coral | Terpenoid f | Dendritic cell maturation suppression | 25 µg/mL * | IL-6, IL-12 and NO inhibition | TWN | [215] |
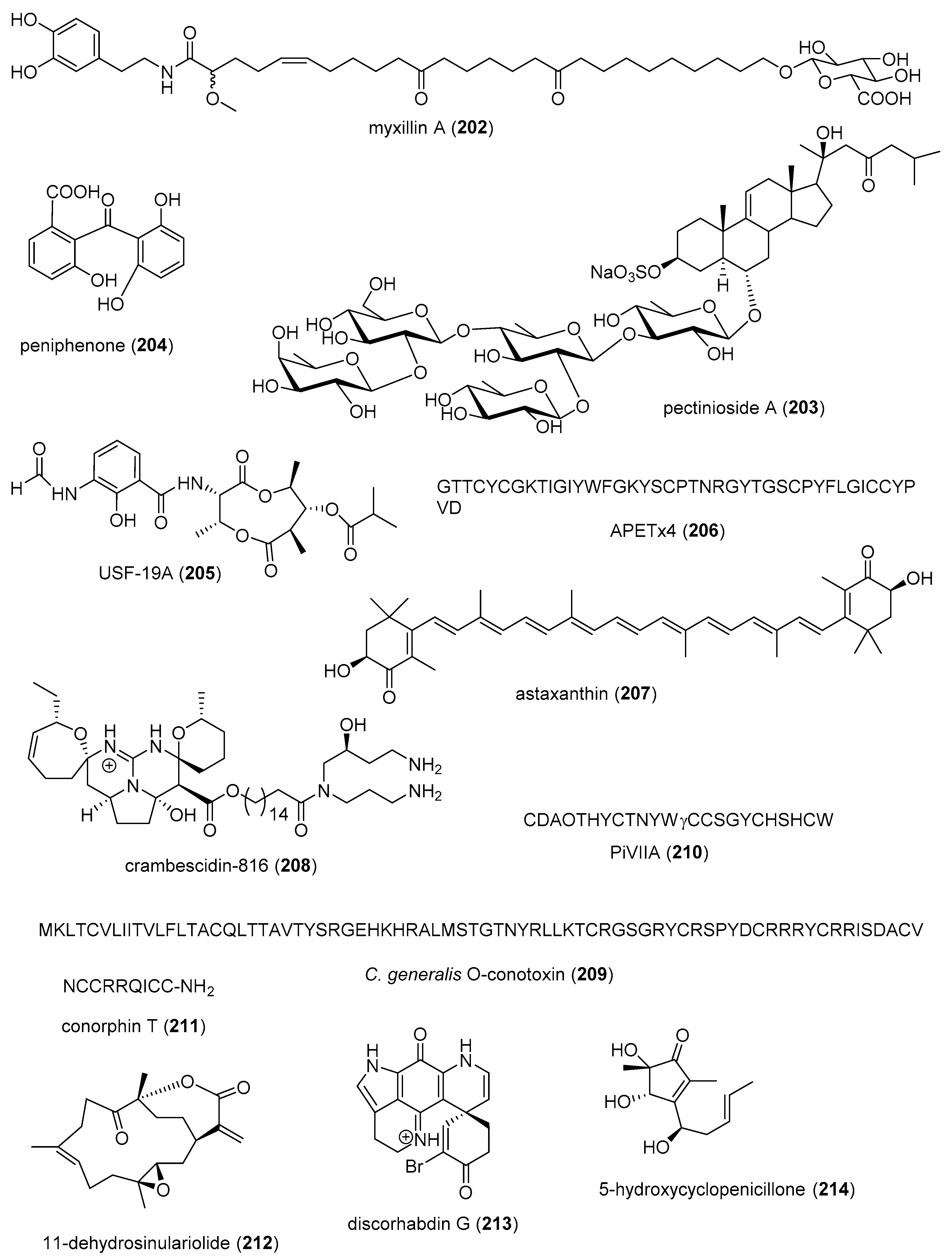
| Immune system | CDMW-3 (199)/fungus | Peptide g | PCA inhibition in vivo | 20 mg/kg ** | Mast cell histamine and cytokine release inhibition | CHN | [216] |
| Immune system | chrysamide C (200)/fungus | Alkaloid g | IL-17 inhibition | >1 μM * | Undetermined | CHN | [217] |
| Immune system | cocosolide (201)/cyanobacterium | Polyketide d | IL-2 inhibition | 2.5 μM * | Undetermined | CHN, USA | [218] |
| Immune system | myxillin A (202)/sponge | Alkaloid g | IL-12p40 release inhibition | 10 µg/mL * | Undetermined | DNK, ISL | [219] |
| Immune system | pectinioside A (203)/starfish | Terpenoid f | Increase OVA-specific IgG1 in vivo | 25 µg * | Undetermined | JPN | [220] |
| Immune system | peniphenone (204)/fungus | Polyketide d | Lymphocyte immune suppression | 8.1–9.3 µg/mL | Undetermined | CHN | [221] |
| Immune system | USF-19A (205)/bacterium | Peptide g | Splenocyte IL-5 release inhibition | 0.57 μM | Undetermined | CHN | [222] |
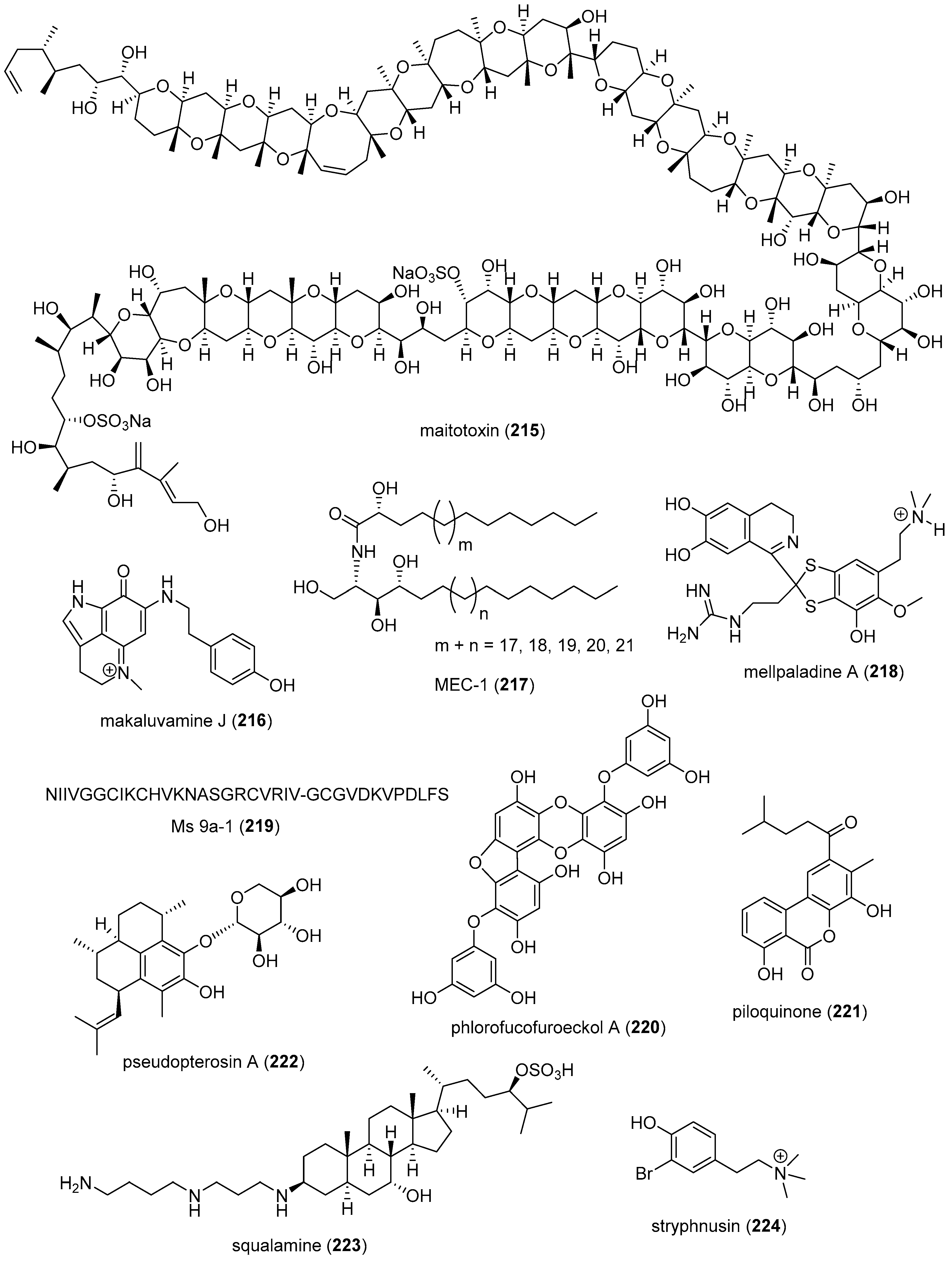
| Nervous system | APETx4 (206)/sea anemone | Peptide g | Kv10.1 potassium channel inhibition | 1.1 µM | Binds channel in closed state | BEL, DEU | [223] |
| Nervous system | astaxanthin (207)/shrimp | Terpenoid f | Penitrem A toxicity reversal | 20 µM * | Block BK channel | EGY, USA | [224] |
| Nervous system | crambescidin 816 (208)/sponge | Alkaloid g | Cortical neurons cytosolic Ca2+ increase | 10 µM * | AMPA and NMDA receptors involved | ESP, FRA, IRL | [225] |
| Nervous system | C. generalis O-conotoxin (209)/cone snail | Peptide g | Α9α10 nACh receptor inhibition | 16.2 nM | Non-competitive inhibition | AUS, CHN | [226] |
| Nervous system | C. princeps PiVIIA peptide (210)/cone snail | Peptide g | Neuronal Ca2+ current increase | 3 µM * | Potentiates two types Ca2+ channels | CUB, MEX | [227] |
| Nervous system | conorphin T (211)/cone snail | Peptide g | KOR agonist | 9.8 µM | In vivo colonic receptor inhibition | AUS | [228] |
| Nervous system | 11-dehydrosinulariolide (212)/soft coral | Terpenoid f | Amelioration PD and spinal cord injury attenuation | 5 µg/rat ** | DJ-1 expression upregulation and microglia activation | TWN | [229,230] |
| Nervous system | discorhabdin G (213)/sponge | Alkaloid g | Eel and human AChE inhibition | 1.3 µM | Reversible competitive inhibition | DEU, ITA, SVN | [231] |
| Nervous system | fucoxanthin (151)/alga | Terpenoid f | BACE1 inhibition | 5.3 µM | Mixed inhibition | GBR, S. KOR | [232] |
| Nervous system | fucoxanthin (151)/alga | Terpenoid f | Reversal BDNF expression | 50 mg/kg ** | Reversed AChE activity | CHN | [233] |
| Nervous system | fucoxanthin (151)/alga | Terpenoid f | Neuroprotection after TBI-induced brain injury | 100 mg/kg ** | Nrf2-ARE pathway modulation | CHN | [234] |
| Nervous system | 5-hydroxycyclopenicillone (214)/fungus | Polyketide d | H2O2-induced neuronal death protection | 30 µM * | DHHP free radical inhibition | CHN, USA | [235] |
| Nervous system | maitotoxin (215)/alga | Polyketide d | Activation of NSCC | 10 pM * | TRPC1 inhibition | MEX | [236] |
| Nervous system | makaluvamine J (216)/sponge | Alkaloid g | Reduction of mitochondrial damage | 0.1–1 µM * | Nrf2 activation | ESP, FJI, GBR | [237] |
| Nervous system | MEC-1 (217)/sponge | Polyketide d | AChE inhibition | 20.9 µM | Docking studies | EGY | [238] |
| Nervous system | mellpaladine A (218)/ascidian | Alkaloid g | In vivo behavior modulation | 8 nM/mouse ** | Serotonin receptor affinity | JPN | [239] |
| Nervous system | Ms 9a-1 peptide (219)/sea anemone | Peptide g | Decrease in nociceptive and inflammatory response in vivo | 0.3 mg/kg ** | TRPA1 modulation | NOR, RUS | [240] |
| Nervous tissue | phlorofucofuroeckol-A (220)/alga | Polyketide d | Glutamate-induced neurotoxicity inhibition | 10 µM * | Intracellular and mitochondrial ROS inhibtion | S. KOR | [241] |
| Nervous system | piloquinone (221)/bacterium | Polyketide d | MAO-B inhibition | 1.2 µM | Reversible competitive inhibition | S. KOR, USA | [242] |
| Nervous system | pseudopterosin A (222)/soft coral | Terpenoid f | Synaptic transmission alteration | 1 µM * | Extensive brain distribution | USA | [243] |
| Nervous system | squalamine (223)/shark | Terpenoid f | Reduction of α-synuclein aggregation in vivo | 50 µM ** | α-synuclein displaced from lipid membranes | ESP, GBR, ITA, NLD, USA | [244] |
| Nervous system | stryphnusin (224)/sponge | Alkaloid g | Eel AChR inhibition | 232 µM | Reversible competitive inhibition | HRV, NOR, SVN, SWE | [245] |
| Nervous system | xyloketal B (225)/fungus | Polyketide d | Cerebral infarction modulation | 50 mg/kg ** | Decreased ROS and cytokines | CAN, CHN, USA | [246] |
| Nervous system | araplysillin X (226)/sponge | Alkaloid g | BACE1 inhibition | 31.4 µM | Undetermined | NZL, USA | [247] |
| Nervous system | caracolamide A (227)/cyanobacterium | Alkaloid g | Ca2+ channel modulation | 10 pM * | Undetermined | BRA, JOR, PAN, USA, | [248] |
| Nervous system | conorfamide-Sr3 (228)/snail | Peptide g | Blocks volatage-gated K+ channel | 2.7 µM *** | Shaker channel specific | MEX | [249] |
| Nervous system | contryphan-Bt (229)/cone snail | Peptide g | Stiff-tail syndrome in vivo | 5 ng/mouse ** | Undetermined | CHN | [250] |
| Nervous system | dehydroaustin (230)/fungus | Meroterpenoid f | AChE inhibition | 0.4 µM | Undetermined | CHN | [251] |
| Nervous system | hymenidin (231)/sponge | Alkaloid g | Kv1.3- Kv1.6 K+ channel inhibition | 2.5–7.6 µM | Undetermined | BEL, GBR, SVN | [252] |
| Nervous system | psammaplysene A (232)/sponge | Alkaloid g | Binding to RNA-binding protein HNRNPK | 86.2 µM *** | Undetermined | USA | [253] |
| Nervous system | terreulactone C (233)/fungus | Meroterpenoid f | AChE inhibition | 28 nM | Undetermined | CHN | [254] |
| Nervous system | turripeptide (234)/turrid snail | Peptide g | Α9α10 nAChR inhibition | 10.2 µM | Undetermined | AUS, KAS, MEX, PHL, USA | [255] |








3.1. Antidiabetic Activity
3.2. Anti-Inflammatory Activity
3.3. Marine Compounds with Activity on the Immune System
3.4. Marine Compounds Affecting the Nervous System
4. Marine Compounds with Miscellaneous Mechanisms of Action
| Compound/Organism a | Chemistry | Pharmacological Activity | IC50 b | MMOA c | Country d | References |
|---|---|---|---|---|---|---|
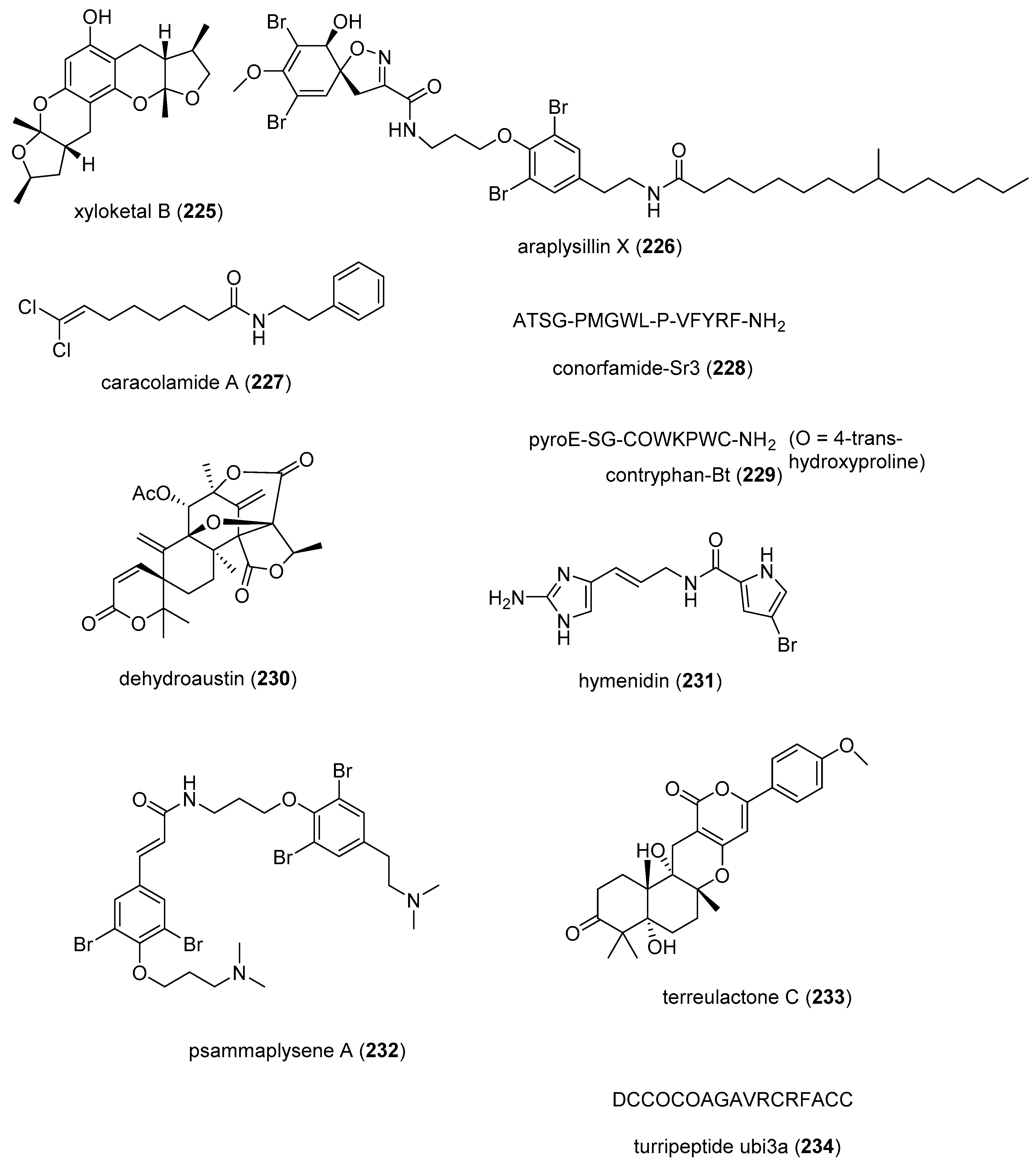
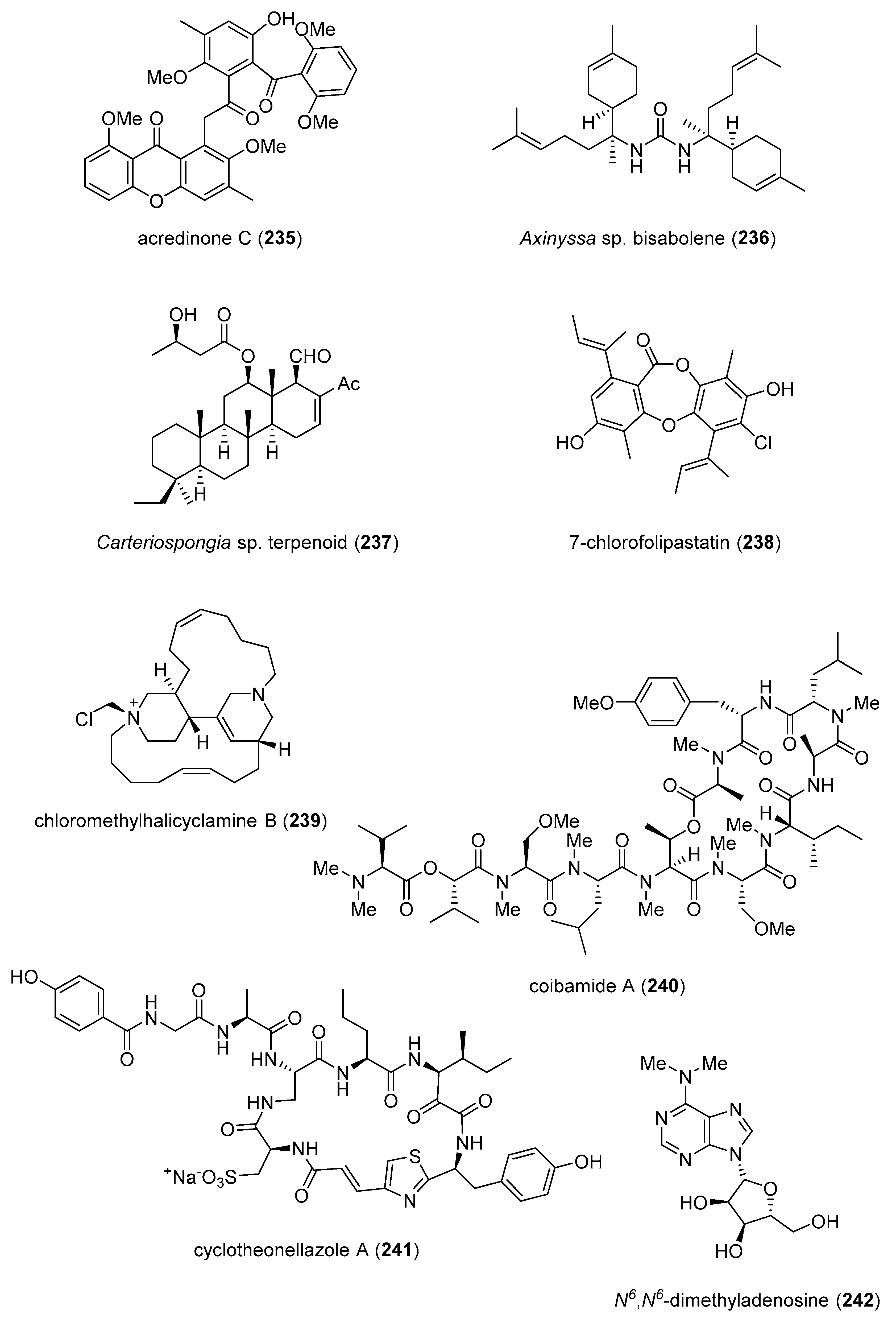
| acredinone C (235)/fungus | Polyketide e | Osteoclast differentiation induction inhibition | 10 μM * | NFATc1 transcription inhibition | S. KOR | [256] |
| astaxanthin (207)/shrimp | Terpenoid f | Hepatic stellate cell activation inhibition | 10 μM * | Decreased ROS and NOX2 expression reduction | USA | [257] |
| Axinyssa sp. bisabolene (236)/sponge | Terpenoid f | PTP1B inhibition | 1.9 μM | Akt phosphorylation | JPN | [258] |
| Carteriospongia sp. terpenoid (237)/sponge | Terpenoid f | Apoptosis induction | 0.06 μg/mL * | Topoisomerase IIα and Hsp90 inhibition | EGY, SWE, TWN | [259] |
| 7-chlorofolipastatin (238)/fungus | Polyketide e | Macrophage SOAT 1 inhibition | 6.8 μM | SOAT 1 and 2 inhibition in vitro | JPN | [260] |
| chloromethylhalicyclamine B (239)/sponge | Alkaloid g | Protein kinase CK1δ/ε inhibition | 6 μM | ATP-binding site docking | FRA, ITA, NLD | [261] |
| coibamide A (240)/cyanobacterium | Peptide f | VEGFA secretion inhibition | <5 nM | Antiangiogenic properties | USA | [262] |
| cyclotheonellazole A (241)/sponge | Peptide f | chymotrypsin and elastase inhibition | 0.034–0.62 nM | Enzyme S2 subsite binding | BEL, ISR, NLD | [263] |
| N6,N6-dimethyladenosine (242)/bacterium | Alkaloid g | AKT phosphorylation inhibition | 5 μM * | S473 site inhibition | USA | [264] |
| excavatolide B (150)/soft coral | Terpenoid f | Modulation of atrial myocytes | 10 μM * | Ca2+ homeostasis modulation | TWN | [265] |
| fucoxanthin (51)/alga | Terpenoid f | Lung fibrosis attenuation | 10 mg/kg *** | Type 1 collagen expression decrease | S. KOR | [266] |
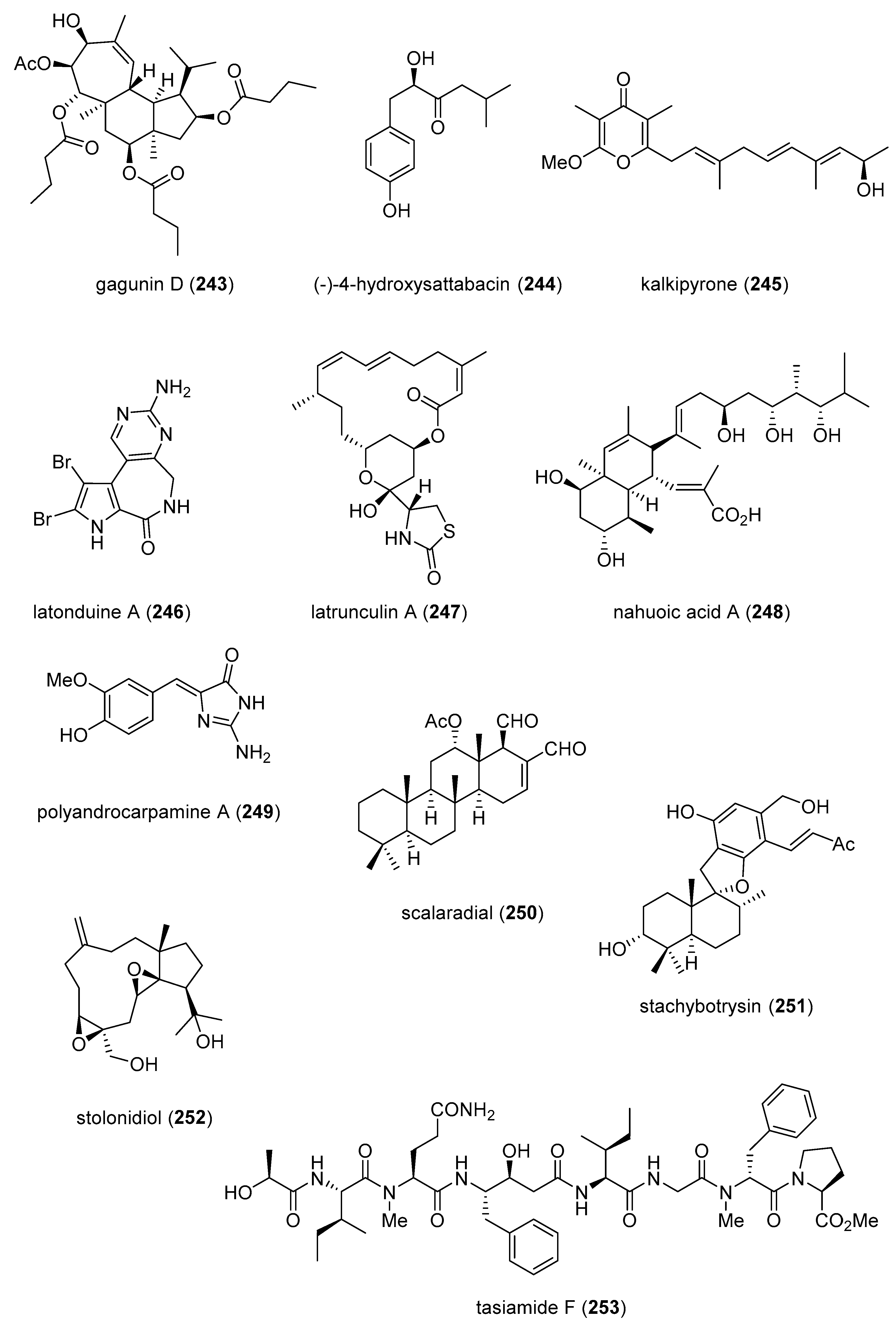
| gagunin D (243)/sponge | Terpenoid f | Melanin synthesis inhibition | 12.7 µM | Tyrosinase expression inhibition | S. KOR | [267] |
| (−)-4-hydroxysattabacin (244)/bacterium | Polyketide e | Melanin synthesis inhibition | 25 μg/mL * | Tyrosinase, TRP-1 and TRP-2 expression inhibition | S. KOR | [268] |
| kalkipyrone (245)/cyanobacterium | Polyketide e | Adipose tissue suppression | 5 mg/kg *** | Enhance LA plasma levels | JPN | [269] |
| latonduine A (246)/sponge | Alkaloid g | CTFR inhibition | 62 nM | PARP isozymes inhibition | CAN, GBR | [270] |
| latrunculin A (247)/sponge | Alkaloid g | ECFC tube inhibition | 0.043 µM | Specific kinases inhibition | USA | [271] |
| nahuoic acid (248)/bacterium | Polyketide e | SETD8 inhibition | 6.5 µM | Competitive inhibition of SAM binding | CAN, PNG | [272] |
| napyradiomycin A1 (49)/bacterium | Terpenoid f | Angiogenesis inhibition | 10 µM | VE-cadherin inhibition | S. KOR | [273] |
| polyandrocarpamine A (249)/sponge | Alkaloid g | DYRK and CLK selective inhibition | 0.17–0.93 µM | Cyclin D1 phosphorylation inhibition | AUS, BRA, DEU, FRA | [274] |
| scalaradial (250)/sponge | Terpenoid f | TRPM2 ion channel inhibition | 0.2 µM | Lack of PLA2 inhibition | JPN, NZL, USA | [275] |
| stachybotrysin (251)/fungus | Terpenoid f | Osteoclast differentiation inhibition | 5 μg/mL * | MAPK kinase pathway inhibition | JPN, S. KOR | [276] |
| stolonidiol (252)/soft coral | Terpenoid f | PKCα membrane translocation | 5 µM * | Increased ChAT activity | USA | [277] |
| tasiamide F (253)/cyanobacterium | Peptide g | Cathepsin D and E inhibition | 23–57 nM | Docking studies completed | USA | [278] |
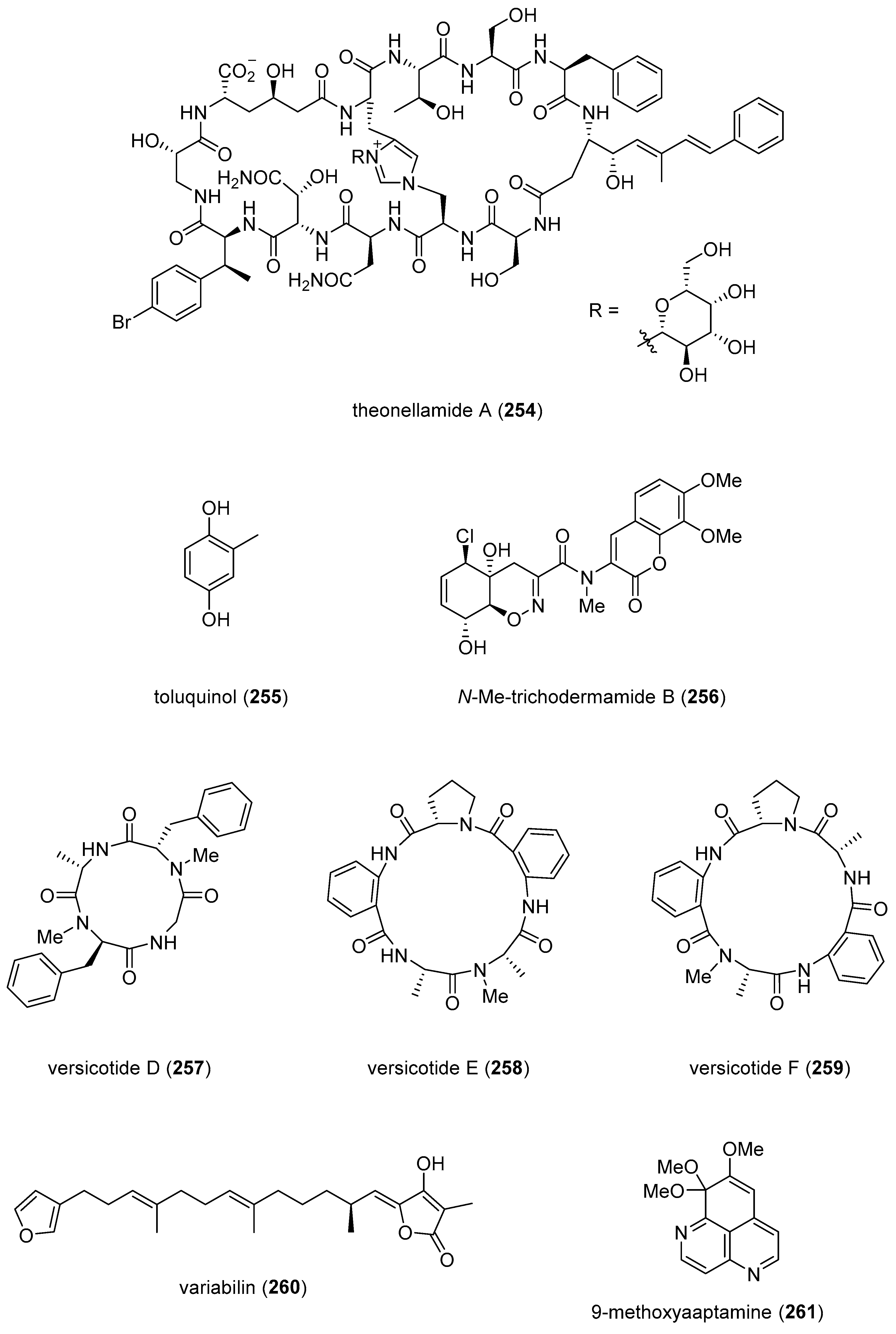
| theonellamide A (254)/sponge | Peptide g | Bilipid membrane disruption | 20 µM * | Binding to sterols | JPN | [279] |
| toluquinol (255)/fungus | Polyketide e | Lymphangiogenesis inhibition | 6.2 µM | Suppression of Akt and ERK ½ phosphorylation | BEL | [280] |
| N-Me-trichodermamide B (256)/fungus | Alkaloid g | H2O2 oxidative damage inhibition | 5 µM * | Nrf2-signaling regulation | CHN | [281] |
| versicotides D–F (257–259)/fungus | Peptide g | Foam cell formation inhibition | 10 µM * | Cholesterol influx inhibition | CHN | [282] |
| variabilin (260)/sponge | Terpenoid f | PTP1B inhibition | 1.5 μM | TCPTP inhibition | IND, JPN | [283] |
| xyloketal B (225)/fungus | Terpenoid f | NAFLD attenuation | 5 mg/kg *** | SREBP-1c expression inhibition | CHN | [284] |
| 9-methoxyaaptamine (261)/sponge | Alkaloid g | PPRE activation | 0.039 μg/mL * | Undetermined | IDN, MYS | [285] |
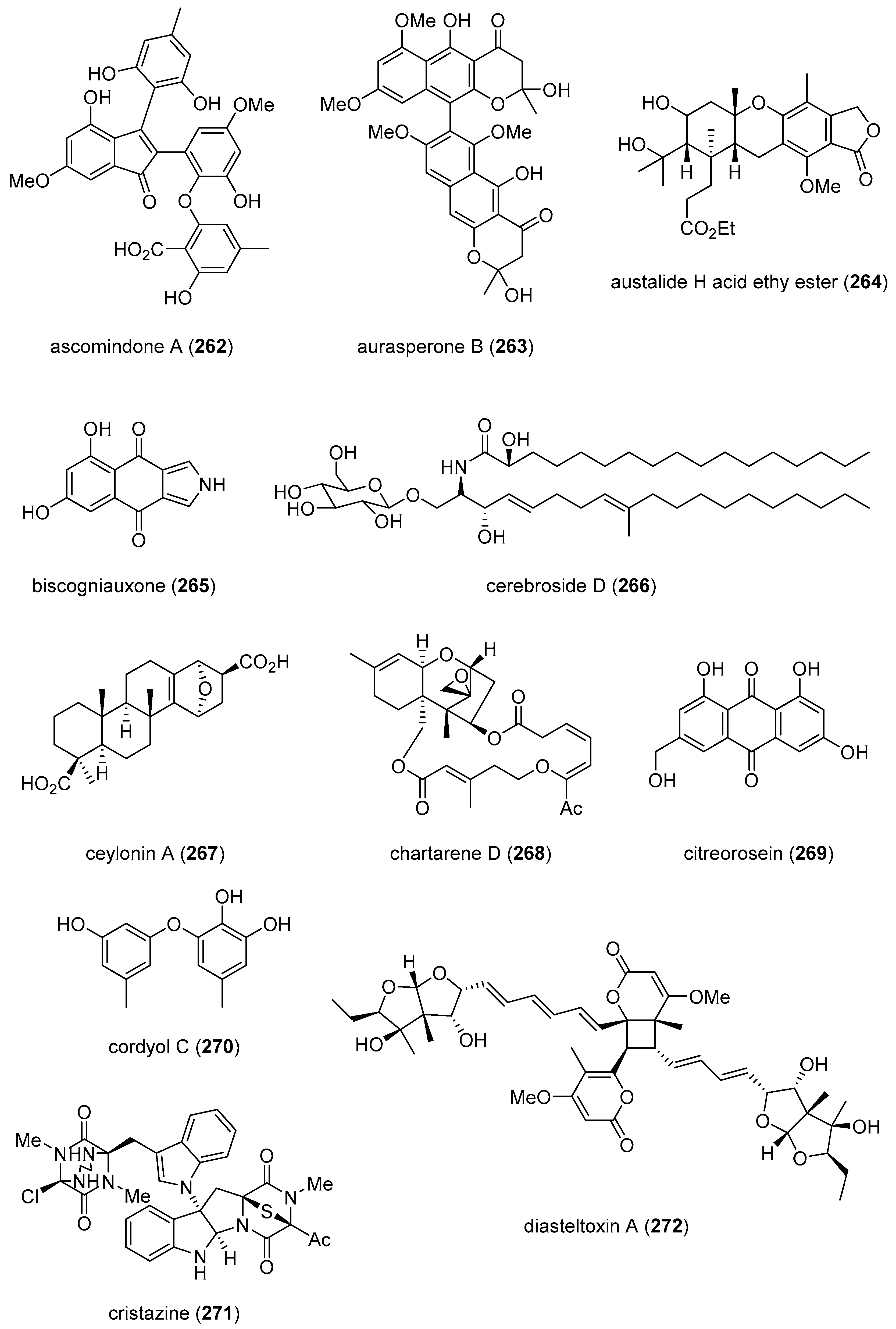
| ascomindone A (262)/fungus | Polyketide e | DPPH radical scavenging inhibition | 18.1 μM | Undetermined | CHN | [286] |
| aurasperone B (263)/fungus | Polyketide e | DPPH radical scavenging inhibition | 0.01 μM | Undetermined | S. KOR | [287] |
| austalide H acid ethy ester (264)/ fungus | Terpenoid f | Endo-1,3-β-D-glucanase inhibition | 0.2 μM | Undetermined | RUS | [288] |
| B. mediterranea cyclopentapeptide (265)/fungus | Peptide g | GSK-3β inhibition | 8.04 μM | Undetermined | CHN, DEU | [289] |
| cerebroside (266)/fungus | Polyketide e | Spermatozoa inhibition | 8 μM | Undetermined | RUS | [290] |
| ceylonin A (267)/sponge | Terpenoid f | Osteoclast inhibition | <50 μM * | Undetermined | NLD, JPN | |
| chartarene D (268)/fungus | Terpenoid f | Tyrosine kinases inhibition | 0.1–0.8 μM | Undetermined | CHN, DEU | [292] |
| citreorosein (269)/fungus | Polyketide e | Anti-obesity activity | 0.17 μM | Undetermined | GBR, PRT, THAI | [293] |
| cordyol C (270)/fungus | Polyketide e | Erythrocyte biomembrane protection | 4.9 μM | Undetermined | CHN | [294] |
| cristazine (271)/fungus | Alkaloid g | DHHP radical scavenging | 19 μM | Undetermined | S. KOR | [295] |
| diasteltoxins A–C (272–274)/fungus | Polyketide e | Thioredoxin reductase inhibition | 7.2–12.8 μM | Undetermined | CHN, DEU | [296] |
| discorhabdin L (275)/sponge | Alkaloid g | HIF-1α transcription inhibition | 0.73 μM | Undetermined | NZL, USA | [297] |
| Dysidea sp. diphenyl ether (276)/sponge | Polyketide e | Mitochondrial complex II inhibition | 6.4 nM | Undetermined | JPN | [298] |
| dysiherbol A (277)/sponge | Terpenoid f | NF-κB inhibition | 0.49 μM | Undetermined | AUS, CHN | [299] |
| echinamine B (278)/sea urchin | Polyketide e | DHHP radical scavenging | 6.5 μM | Undetermined | RUS | [300] |
| endolide B (279)/fungus | Peptide g | Serotonin receptor 5HT2b inhibition | 0.77 μM ** | Undetermined | DEU | [301] |
| europhenol A (280)/fungus | Polyketide e | DHHP radical scavenging | 1.23 μg/mL | Undetermined | CHN, HUN | [302] |
| fascaplysin (281)/sponge | Alkaloid g | P-glycoprotein induction | 1 μM * | Undetermined | IDN | [303] |
| fennebricin A (282)/nudibranch | Alkaloid g | NF-κB inhibition | 1 μM | Undetermined | CHN, HUN | [304] |
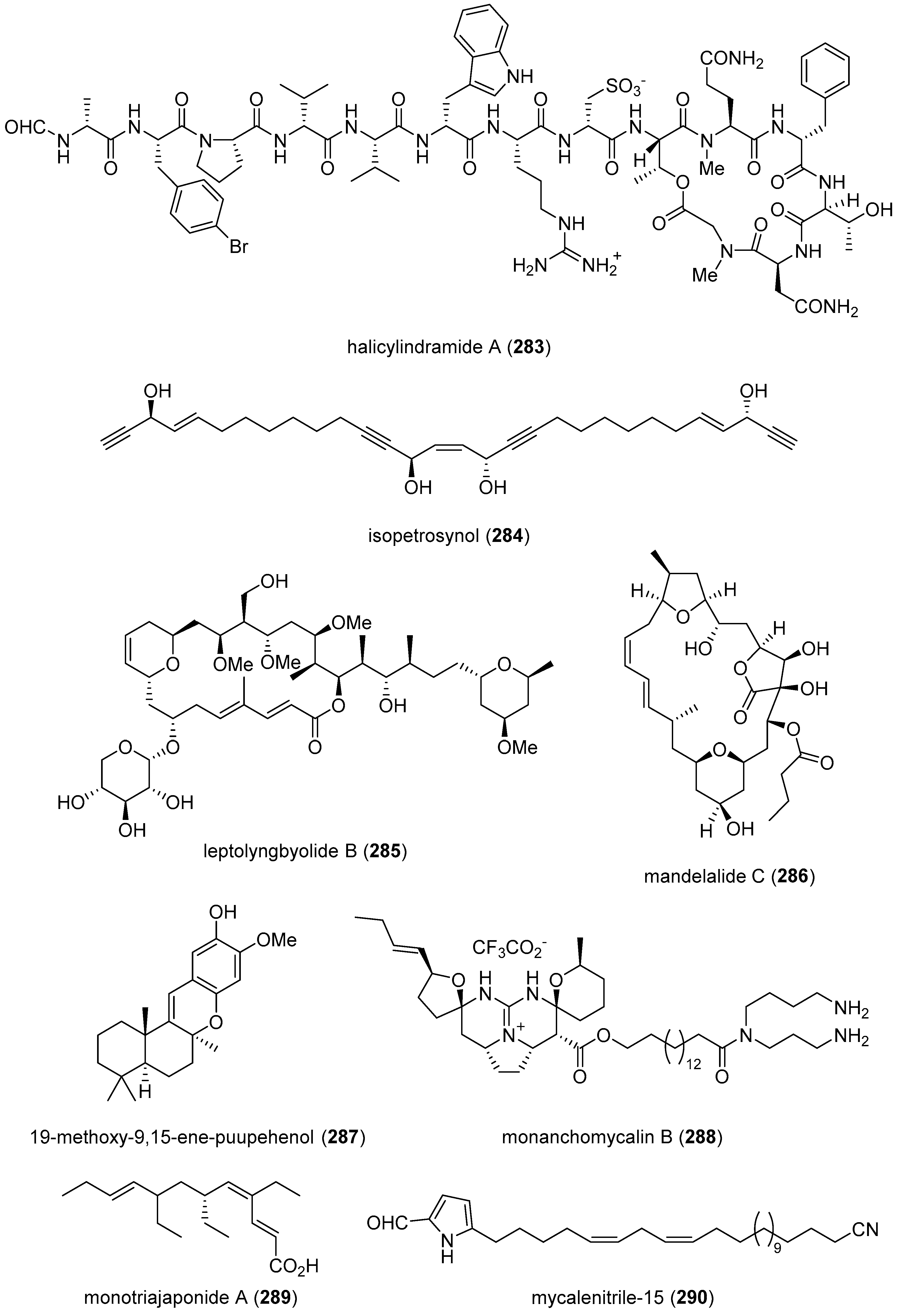
| halicylindramide A(283)/sponge | Peptide g | FXR receptor inhibition | 0.5 μM | Undetermined | AUS, S.KOR | [305] |
| isopetrosynol (284)/sponge | Polyketide e | PTP1B inhibition | 8.2 μM | Undetermined | IDN, JPN | [306] |
| leptolyngbyolide B (285)/ cyanobacterium | Polyketide e | F-actin depolymerization | 11.6 μM | Undetermined | JPN | [307] |
| mandelalide C (286)/ascidian | Polyketide e | Mitochondrial complex V inhibition | 3.4 μM | Undetermined | USA | [308] |
| 19-methoxy-9,15-ene-puupehenol (287)/sponge | Terpenoid f | SR-B1 receptor activation | 1.78 μM | Undetermined | AUS, MYS | [309] |
| monanchomycalin B (288)/sponge | Alkaloid g | TRPV1, 2 and 3 receptor inhibition | 2.8–6.0 μM | Undetermined | RUS, S. KOR | [310] |
| monotriajaponide A (289)/sponge | Polyketide e | PPAR-α and -β activation | 12.5 μM * | Undetermined | CHN, ITA, USA | [311] |
| mycalenitrile-15 (290)/sponge | Terpenoid f | PTP1B inhibition | 8.6 μM * | Undetermined | CHN, ITA | [312] |
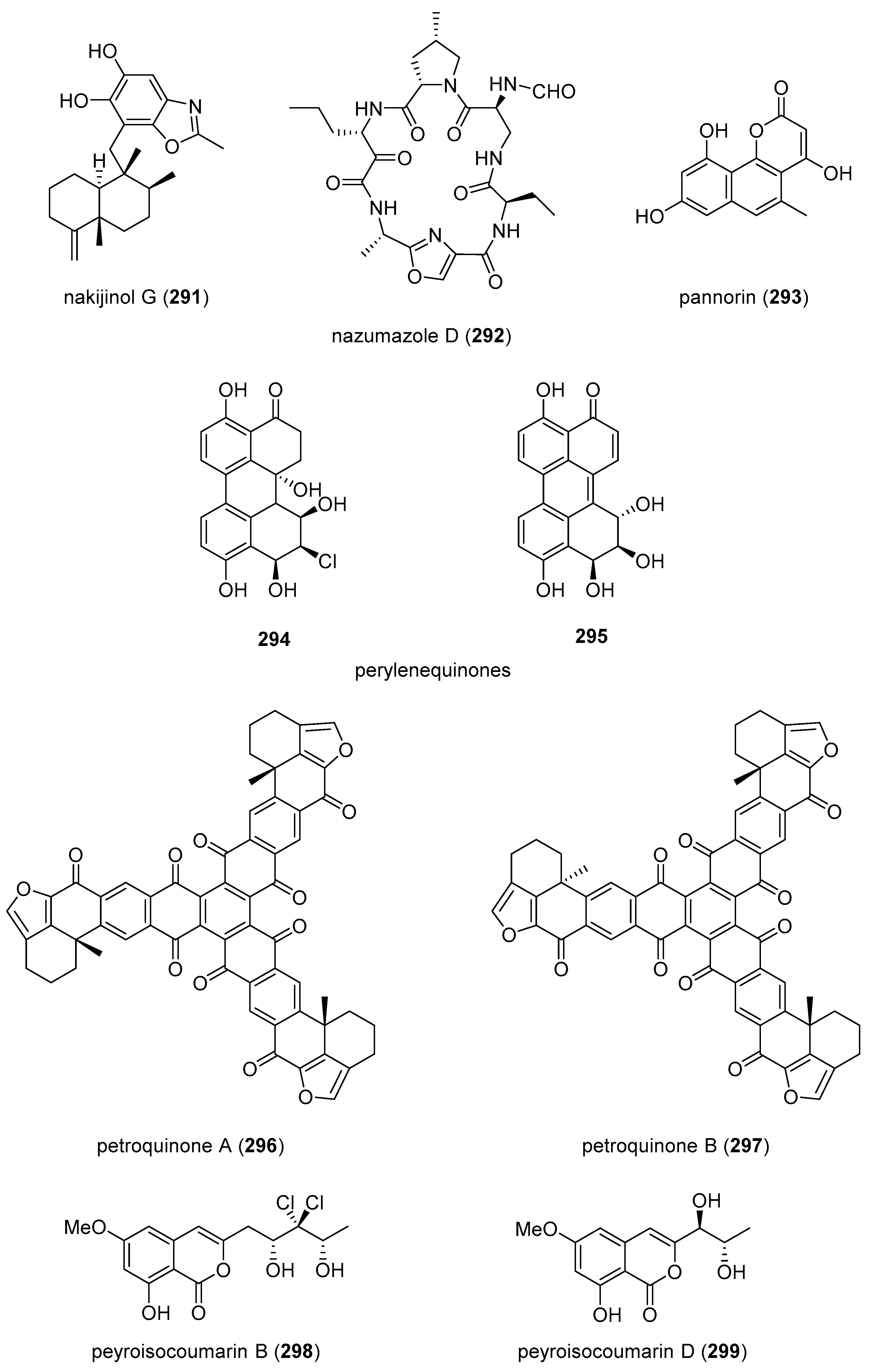
| nakijinol G (291)/sponge | Terpenoid f | PTP1B inhibition | 4.8 μM | Undetermined | CHN | [313] |
| nazumazole D (292)/sponge | Peptide g | chymotrypsin activity inhibition | 2 μM | Undetermined | JPN | [314] |
| pannorin (293)/fungus | Polyketide e | GSK-3β inhibition | 0.35 µM | Undetermined | DEU | [315] |
| perylenequinones (294, 295)/fungus | Polyketide e | BRD4 protein inhibition | 10 μM * | Undetermined | CHN | [316] |
| petroquinones A and B (296, 297)/sponge | Polyketide e | USP7 inhibition | 0.13–2.0 μM | Undetermined | IND, JPN, NLD | [317] |
| peyroisocoumarins B and D (298, 299)/fungus | Polyketide e | ARE expression induction | 10 μM * | Undetermined | CHN, DEU | [318] |
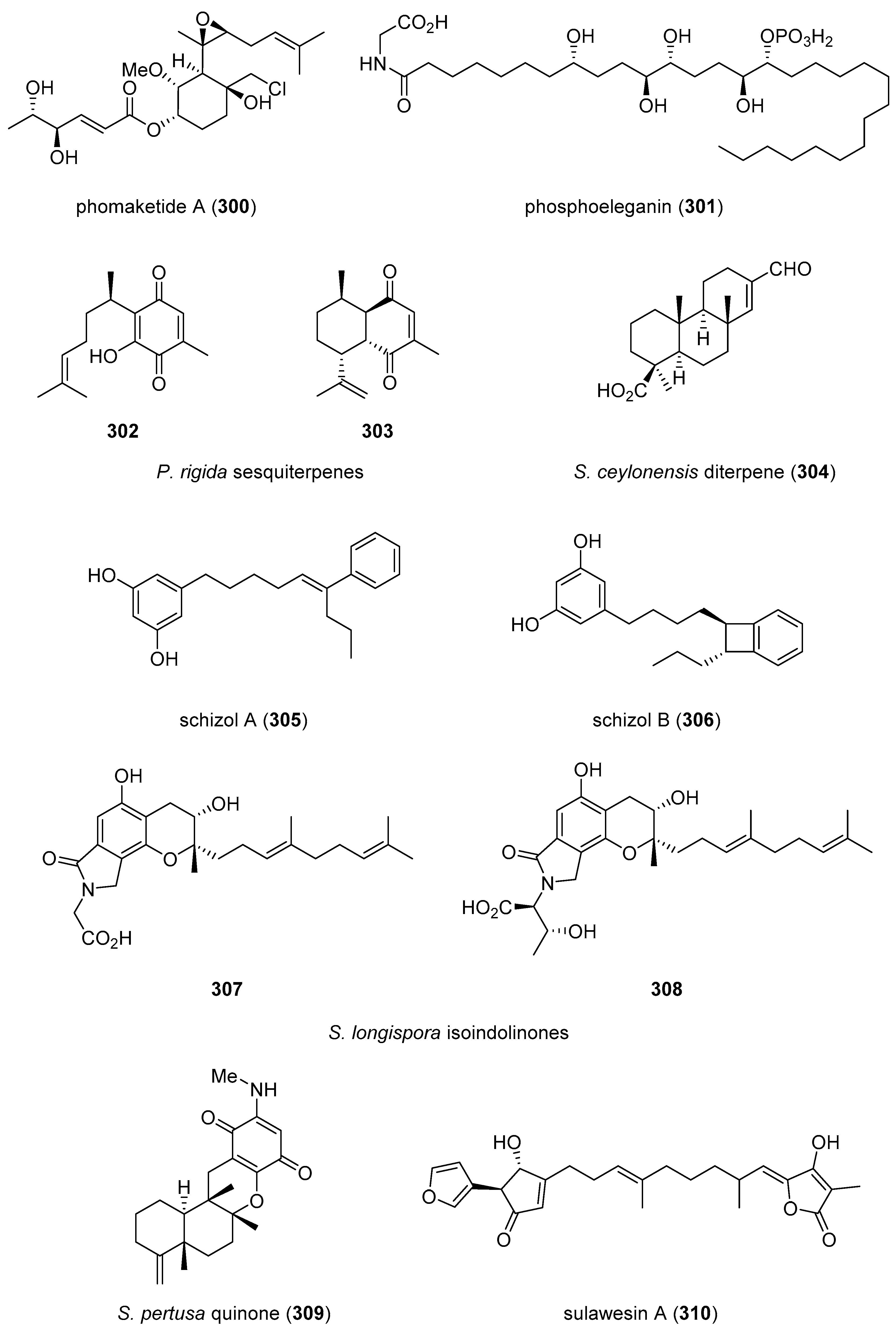
| phomaketide A (300)/fungus | Polyketide e | Angiogenesis inhibition | 8.1 μM | Undetermined | TWN | [319] |
| phosphoeleganin (301)/ascidian | Polyketide e | PTP1B inhibition | 11 μM | Undetermined | CHN, ITA | [320] |
| P. rigida sesquiterpenes (302, 303)/soft coral | Terpenoid f | CDC25 phosphatases | 12–3.4 μM | Undetermined | GRC, FRA | [321] |
| S. ceylonensis diterpene (304)/sponge | Terpenoid f | USP7 inhibition | 8.2 µM | Undetermined | EGY, JPN | [322] |
| schizols A and B (305, 306)/bryozoa | Polyketide e | ABTS cation radical inhibition | 6.2–7.6 µM | Undetermined | ESP | [323] |
| S. longispora isoindolinones (307, 308)/fungus | Alkaloid g | Fibrinolytic activity | 25 μg/mL * | Undetermined | CHN | [324] |
| S. pertusa quinone (309)/sponge | Terpenoid f | CDK-2 inhibition | 4.8 µM ** | Undetermined | CHN | [325] |
| sulawesin A (310)/sponge | Terpenoid f | USP7 inhibition | 2.8 μM | Undetermined | EGY, IDN, JPN, NLD | [326] |
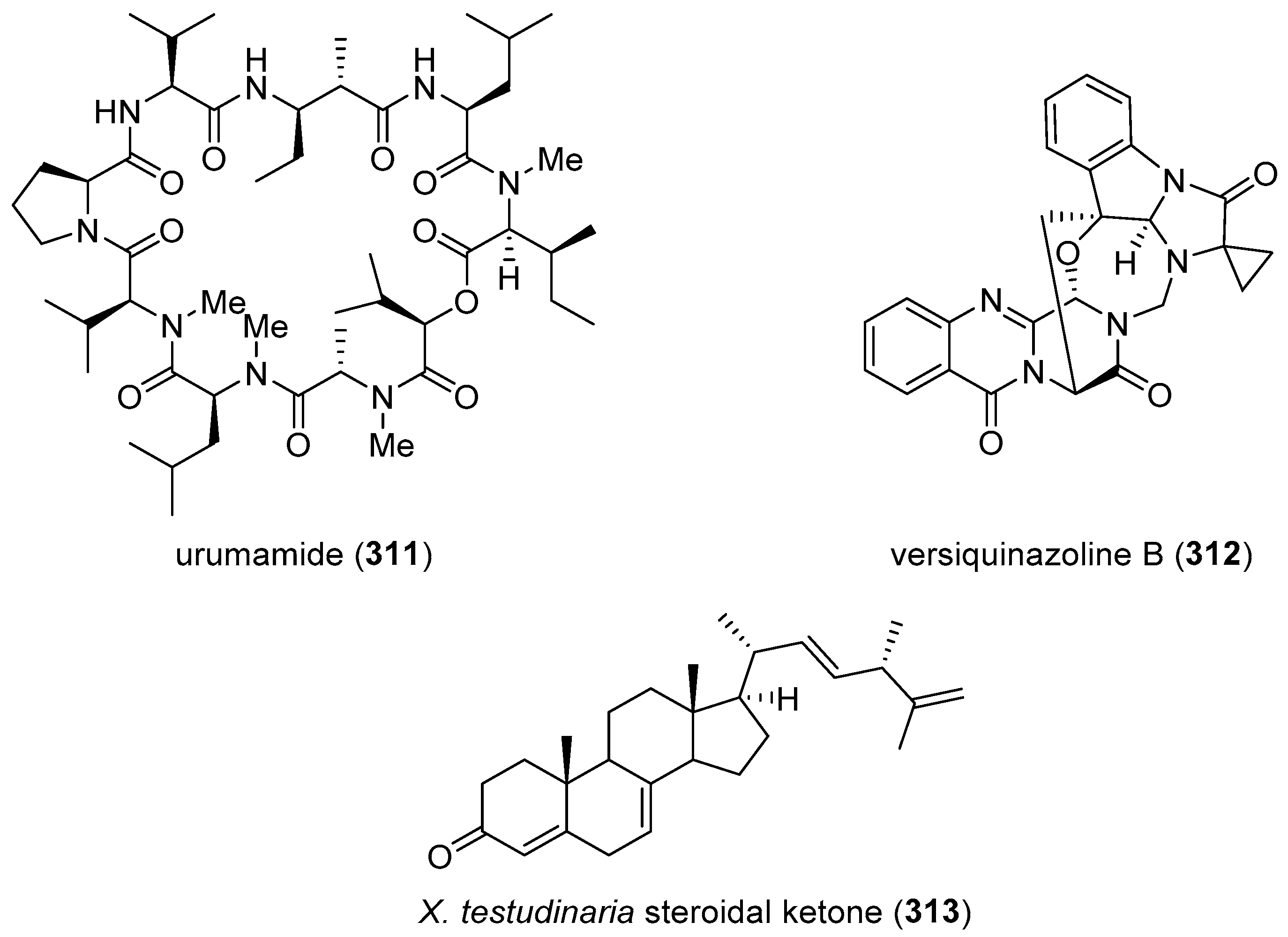
| urumamide (311)/cyanobacterium | Peptide g | Chymotrypsin inhibition | 33 μM | Undetermined | JPN | [327] |
| versiquinazoline B (312)/fungus | Alkaloid g | Thioredoxin reductase inhibition | 12 µM | Undetermined | CHN, DEU | [328] |
| X. testudinaria steroidal ketone (313)/ sponge | Terpenoid f | PTP1B inhibition | 4.27 μM | Undetermined | CHN | [329] |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
5. Reviews on Marine Pharmacology and Pharmaceuticals
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mayer, A.M.S.; Lehmann, V.K.B. Marine pharmacology in 1998: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, anthelmintic, antiplatelet, antiprotozoal, and antiviral activities; with actions on the cardiovascular, endocrine, immune, and nervous systems; and other miscellaneous mechanisms of action. Pharmacologist 2000, 42, 62–69. [Google Scholar]
- Mayer, A.M.; Hamann, M.T. Marine pharmacology in 1999: Compounds with antibacterial, anticoagulant, antifungal, anthelmintic, anti-inflammatory, antiplatelet, antiprotozoal and antiviral activities affecting the cardiovascular, endocrine, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 2002, 132, 315–339. [Google Scholar]
- Mayer, A.M.S.; Hamann, M.T. Marine pharmacology in 2000: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antituberculosis, and antiviral activities; affecting the cardiovascular, immune, and nervous systems and other miscellaneous mechanisms of action. Mar. Biotechnol. 2004, 6, 37–52. [Google Scholar]
- Mayer, A.M.; Hamann, M.T. Marine pharmacology in 2001–2002: Marine compounds with anthelmintic, antibacterial, anticoagulant, antidiabetic, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems and other miscellaneous mechanisms of action. Comp. Biochem. Phys. C Toxicol. Pharmacol. 2005, 140, 265–286. [Google Scholar]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2003–2004: Marine compounds with anthelmintic antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Phys. C Toxicol. Pharmacol. 2007, 145, 553–581. [Google Scholar]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2005–2006: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta 2009, 1790, 283–308. [Google Scholar]
- Mayer, A.M.; Rodriguez, A.D.; Berlinck, R.G.; Fusetani, N. Marine pharmacology in 2007–2008: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Comp. Biochem. Phys. C Toxicol. Pharmacol. 2011, 153, 191–222. [Google Scholar]
- Mayer, A.M.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2009–2011: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and other Miscellaneous Mechanisms of Action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar]
- Mayer, A.M.S.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2012–2013: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2017, 15, 273. [Google Scholar]
- Mayer, A.M.S.; Guerrero, A.J.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2014–2015: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, Antiviral, and Anthelmintic Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2019, 18, 5. [Google Scholar]
- Schmitz, F.J.; Bowden, B.F.; Toth, S.I. Antitumor and Cytotoxic Compounds from Marine Organisms. In Marine Biotechnology, Pharmaceutical and Bioactive Natural Products; Attaway, D.H., Zaborsky, O.R., Eds.; Plenum Press: New York, NY, USA; London, UK, 1993; Volume 1, pp. 197–308. [Google Scholar]
- Beesoo, R.; Bhagooli, R.; Neergheen-Bhujun, V.S.; Li, W.W.; Kagansky, A.; Bahorun, T. Antibacterial and antibiotic potentiating activities of tropical marine sponge extracts. Comp. Biochem. Phys. C Toxicol. Pharmacol. 2017, 196, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Kim, H.S.; Kim, S.Y.; Lee, S.H.; Lee, W.W.; Jeon, Y.J. Identification of sterols from the soft coral Dendronephthya gigantea and their anti-inflammatory potential. Environ. Toxicol. Pharmacol. 2017, 55, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Joseph, F.R.S.; Iniyan, A.M.; Vincent, S.G.P. HR-LC-MS based analysis of two antibacterial metabolites from a marine sponge symbiont Streptomyces pharmamarensis ICN40. Microb. Pathog. 2017, 111, 450–457. [Google Scholar] [CrossRef]
- Al-Amoudi, S.; Essack, M.; Simoes, M.F.; Bougouffa, S.; Soloviev, I.; Archer, J.A.; Lafi, F.F.; Bajic, V.B. Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential. Mar. Drugs 2016, 14, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iniguez-Martinez, A.M.; Cardoso-Martinez, F.; de la Rosa, J.M.; Cueto, M.; Diaz-Marrero, A.R.; Darias, J.; Becerril-Espinosa, A.; Plata-Rosas, L.; Soria-Mercado, I.E. Compounds isolated from Salinispora arenicola of the Gulf of California, Mexico. Rev. Biol. Mar. Oceanogr. 2016, 51, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.M.; Murphy, J.; MacAdam, D.; Osterbauer, C.; Baseer, I.; Hall, M.L.; Feher, D.; Williams, P. Classical and Alternative Activation of Cyanobacterium Oscillatoria sp. Lipopolysaccharide-Treated Rat Microglia in vitro. Toxicol. Sci. 2016, 149, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Swanson-Mungerson, M.; Incrocci, R.; Subramaniam, V.; Williams, P.; Hall, M.L.; Mayer, A.M.S. Effects of cyanobacteria Oscillatoria sp. lipopolysaccharide on B cell activation and Toll-like receptor 4 signaling. Toxicol. Lett. 2017, 275, 101–107. [Google Scholar] [CrossRef]
- Di, X.; Oskarsson, J.T.; Omarsdottir, S.; Freysdottir, J.; Hardardottir, I. Lipophilic fractions from the marine sponge Halichondria sitiens decrease secretion of pro-inflammatory cytokines by dendritic cells and decrease their ability to induce a Th1 type response by allogeneic CD4+ T cells. Pharm. Biol. 2017, 55, 2116–2122. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Othman, E.M.; Reimer, A.; Grune, M.; Kozjak-Pavlovic, V.; Stopper, H.; Hentschel, U.; Abdelmohsen, U.R. Ageloline A, new antioxidant and antichlamydial quinolone from the marine sponge-derived bacterium Streptomyces sp. SBT345. Tetrahedrom Lett. 2016, 57, 2786–2789. [Google Scholar] [CrossRef]
- Davison, J.R.; Lohith, K.M.; Wang, X.; Bobyk, K.; Mandadapu, S.R.; Lee, S.L.; Cencic, R.; Nelson, J.; Simpkins, S.; Frank, K.M.; et al. A New Natural Product Analog of Blasticidin S Reveals Cellular Uptake Facilitated by the NorA Multidrug Transporter. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Le, N.T.; Niemann, H.; Proksch, P.; Tait, K.; Linossier, I.; Rehel, K.; Hellio, C.; Fay, F. Sponge-Inspired Dibromohemibastadin Prevents and Disrupts Bacterial Biofilms without Toxicity. Mar. Drugs 2017, 15. [Google Scholar] [CrossRef] [Green Version]
- Wyche, T.P.; Alvarenga, R.F.R.; Piotrowski, J.S.; Duster, M.N.; Warrack, S.R.; Cornilescu, G.; De Wolfe, T.J.; Hou, Y.; Braun, D.R.; Ellis, G.A.; et al. Chemical Genomics, Structure Elucidation, and in Vivo Studies of the Marine-Derived Anticlostridial Ecteinamycin. ACS Chem. Biol. 2017, 12, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, S.P.; Hung, W.C.; Huang, C.Y.; Lin, Y.S.; Chan, M.Y.; Lu, P.L.; Lin, L.; Sheu, J.H. 5-Episinuleptolide Decreases the Expression of the Extracellular Matrix in Early Biofilm Formation of Multi-Drug Resistant Acinetobacter baumannii. Mar. Drugs 2016, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Solstad, R.G.; Li, C.; Isaksson, J.; Johansen, J.; Svenson, J.; Stensvag, K.; Haug, T. Novel Antimicrobial Peptides EeCentrocins 1, 2 and EeStrongylocin 2 from the Edible Sea Urchin Echinus esculentus Have 6-Br-Trp Post-Translational Modifications. PLoS ONE 2016, 11, e0151820. [Google Scholar] [CrossRef] [Green Version]
- Sung, A.A.; Gromek, S.M.; Balunas, M.J. Upregulation and Identification of Antibiotic Activity of a Marine-Derived Streptomyces sp. via Co-Cultures with Human Pathogens. Mar. Drugs 2017, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Adnani, N.; Chevrette, M.G.; Adibhatla, S.N.; Zhang, F.; Yu, Q.; Braun, D.R.; Nelson, J.; Simpkins, S.W.; McDonald, B.R.; Myers, C.L.; et al. Coculture of Marine Invertebrate-Associated Bacteria and Interdisciplinary Technologies Enable Biosynthesis and Discovery of a New Antibiotic, Keyicin. ACS Chem. Biol. 2017, 12, 3093–3102. [Google Scholar] [CrossRef]
- Mokhlesi, A.; Stuhldreier, F.; Wex, K.W.; Berscheid, A.; Hartmann, R.; Rehberg, N.; Sureechatchaiyan, P.; Chaidir, C.; Kassack, M.U.; Kalscheuer, R.; et al. Cyclic Cystine-Bridged Peptides from the Marine Sponge Clathria basilana Induce Apoptosis in Tumor Cells and Depolarize the Bacterial Cytoplasmic Membrane. J. Nat. Prod. 2017, 80, 2941–2952. [Google Scholar] [CrossRef]
- Leoni, G.; De, P.A.; Mardirossian, M.; Gambato, S.; Florian, F.; Venier, P.; Wilson, D.N.; Tossi, A.; Pallavicini, A.; Gerdol, M. Myticalins: A Novel Multigenic Family of Linear, Cationic Antimicrobial Peptides from Marine Mussels (Mytilus spp.). Mar. Drugs 2017, 15, 261. [Google Scholar] [CrossRef] [Green Version]
- Costantino, V.; Della, S.G.; Saurav, K.; Teta, R.; Bar-Shalom, R.; Mangoni, A.; Steindler, L. Plakofuranolactone as a Quorum Quenching Agent from the Indonesian Sponge Plakortis cf. lita. Mar. Drugs 2017, 15, 59. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Lee, A.; Jung, J.H.; Choi, S.H.; Kim, T.S. In vitro and in vivo anti-Vibrio vulnificus activity of psammaplin A, a natural marine compound. Mol. Med. Rep 2016, 14, 2691–2696. [Google Scholar] [CrossRef]
- Song, Y.; Li, Q.; Qin, F.; Sun, C.; Liang, H.; Wei, X.; Wong, N.K.; Ye, L.; Zhang, Y.; Shao, M.; et al. Neoabyssomicins A–C, polycyclic macrolactones from the deep-sea derived Streptomyces koyangensis SCSIO 5802. Tetrahedron 2017, 73, 5366–5372. [Google Scholar] [CrossRef]
- Zhang, X.; Ye, X.; Chai, W.; Lian, X.Y.; Zhang, Z. New Metabolites and Bioactive Actinomycins from Marine-Derived Streptomyces sp. ZZ338. Mar. Drugs 2016, 14, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Wang, C.; Gui, P.; Liu, H.; Khalaf, S.M.H.; Elsayed, E.A.; Wadaan, M.A.M.; Hozzein, W.N.; Zhu, W. Identification, Bioactivity, and Productivity of Actinomycins from the Marine-Derived Streptomyces heliomycini. Front. Microbiol. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Balan, S.S.; Kumar, C.G.; Jayalakshmi, S. Aneurinifactin, a new lipopeptide biosurfactant produced by a marine Aneurinibacillus aneurinilyticus SBP-11 isolated from Gulf of Mannar: Purification, characterization and its biological evaluation. Microbiol. Res. 2017, 194, 1–9. [Google Scholar] [CrossRef]
- Li, X.D.; Li, X.M.; Li, X.; Xu, G.M.; Liu, Y.; Wang, B.G. Aspewentins D-H, 20-Nor-isopimarane Derivatives from the Deep Sea Sediment-Derived Fungus Aspergillus wentii SD-310. J. Nat. Prod. 2016, 79, 1347–1353. [Google Scholar] [CrossRef]
- Chakraborty, K.; Thilakan, B.; Raola, V.K. Antimicrobial polyketide furanoterpenoids from seaweed-associated heterotrophic bacterium Bacillus subtilis MTCC 10403. Phytochemistry 2017, 142, 112–125. [Google Scholar] [CrossRef]
- Huang, H.; Liu, T.; Wu, X.; Guo, J.; Lan, X.; Zhu, Q.; Zheng, X.; Zhang, K. A new antibacterial chromone derivative from mangrove-derived fungus Penicillium aculeatum (No. 9EB). Nat. Prod. Res. 2017, 31, 2593–2598. [Google Scholar] [CrossRef]
- Tareq, F.S.; Shin, H.J. Bacilotetrins A and B, Anti-Staphylococcal Cyclic-Lipotetrapeptides from a Marine-Derived Bacillus subtilis. J. Nat. Prod. 2017, 80, 2889–2892. [Google Scholar] [CrossRef]
- Brana, A.F.; Sarmiento-Vizcaino, A.; Perez-Victoria, I.; Otero, L.; Fernandez, J.; Palacios, J.J.; Martin, J.; de la Cruz, M.; Diaz, C.; Vicente, F.; et al. Branimycins B and C, Antibiotics Produced by the Abyssal Actinobacterium Pseudonocardia carboxydivorans M-227. J. Nat. Prod. 2017, 80, 569–573. [Google Scholar] [CrossRef]
- Meng, L.H.; Wang, C.Y.; Mandi, A.; Li, X.M.; Hu, X.Y.; Kassack, M.U.; Kurtan, T.; Wang, B.G. Three Diketopiperazine Alkaloids with Spirocyclic Skeletons and One Bisthiodiketopiperazine Derivative from the Mangrove-Derived Endophytic Fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef]
- Wang, W.; Kim, H.; Patil, R.S.; Giri, A.G.; Won, D.H.; Hahn, D.; Sung, Y.; Lee, J.; Choi, H.; Nam, S.J.; et al. Cadiolides J-M, antibacterial polyphenyl butenolides from the Korean tunicate Pseudodistoma antinboja. Bioorg. Med. Chem. Lett. 2017, 27, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Tripathi, A.; Wu, J.; Schultz, P.J.; Yim, I.; McQuade, T.J.; Yu, F.; Arevang, C.J.; Mensah, A.Y.; Tamayo-Castillo, G.; et al. Discovery of cahuitamycins as biofilm inhibitors derived from a convergent biosynthetic pathway. Nat. Commun. 2016, 7, 10710. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Ding, R.; Jiang, S.T.; Tang, J.S.; Hu, D.; Chen, G.D.; Lin, F.; Hong, K.; Yao, X.S.; Gao, H. Aldgamycins J-O, 16-Membered Macrolides with a Branched Octose Unit from Streptomycetes sp. and Their Antibacterial Activities. J. Nat. Prod. 2016, 79, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, L.; Pei, X.; Deng, F.; Hu, D.; Chen, G.; Wang, C.; Hong, K.; Yao, X.; Gao, A.H. Chalcomycins from Marine-Derived Streptomyces sp. and Their Antimicrobial Activities. Mar. Drugs 2017, 15, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Li, X.M.; Liu, Y.; Zhang, P.; Wang, J.N.; Wang, B.G. Chermesins A-D: Meroterpenoids with a Drimane-Type Spirosesquiterpene Skeleton from the Marine Algal-Derived Endophytic Fungus Penicillium chermesinum EN-480. J. Nat. Prod. 2016, 79, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, D.; Cai, R.; Cui, H.; Long, Y.; Lu, Y.; Li, C.; She, Z. Cytotoxic and Antibacterial Preussomerins from the Mangrove Endophytic Fungus Lasiodiplodia theobromae ZJ-HQ1. J. Nat. Prod. 2016, 79, 2397–2402. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, E.; Choi, H.; Lee, J. Collismycin C from the Micronesian Marine Bacterium Streptomyces sp. MC025 Inhibits Staphylococcus aureus Biofilm Formation. Mar. Drugs 2017, 15, 387. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, Antibiotic Polyketides from the Marine Fungus Engyodontium album Strain LF069. Chemistry 2016, 22, 7452–7462. [Google Scholar] [CrossRef]
- Li, H.L.; Li, X.M.; Li, X.; Wang, C.Y.; Liu, H.; Kassack, M.U.; Meng, L.H.; Wang, B.G. Antioxidant Hydroanthraquinones from the Marine Algal-Derived Endophytic Fungus Talaromyces islandicus EN-501. J. Nat. Prod. 2017, 80, 162–168. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Ito, T.; Kurimoto, S.I.; Ogawa, M.; Win, N.N.; Hung, V.Q.; Nguyen, H.T.; Kubota, T.; Kobayashi, J.; Morita, H. New merosesquiterpenes from a Vietnamese marine sponge of Spongia sp. and their biological activities. Bioorg. Med. Chem. Lett. 2017, 27, 3043–3047. [Google Scholar] [CrossRef]
- Lee, J.; Shin, A.; Lee, H. Isolation and synthesis of luffariellolide derivatives and evaluation of antibacterial activities against gram-negative bacteria. Bull. Korean Chem. Soc. 2017, 38, 804–807. [Google Scholar] [CrossRef]
- Kim, C.K.; Riswanto, R.; Won, T.H.; Kim, H.; Elya, B.; Sim, C.J.; Oh, D.C.; Oh, K.B.; Shin, J. Manzamine Alkaloids from an Acanthostrongylophora sp. Sponge. J. Nat. Prod. 2017, 80, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Lacret, R.; Perez-Victoria, I.; Oves-Costales, D.; de la Cruz, M.; Domingo, E.; Martin, J.; Diaz, C.; Vicente, F.; Genilloud, O.; Reyes, F. MDN-0170, a New Napyradiomycin from Streptomyces sp. Strain CA-271078. Mar. Drugs 2016, 14, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nenkep, V.; Yun, K.; Son, B.W. Oxysporizoline, an antibacterial polycyclic quinazoline alkaloid from the marine-mudflat-derived fungus Fusarium oxysporum. J. Antibiot. 2016, 69, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.J.; Huang, G.L.; Xu, Y.; Song, X.M.; Yao, J.; Liu, H.; Wang, R.P.; Sun, X.P. A new benzopyrans derivatives from a mangrove-derived fungus Penicillium citrinum from the South China Sea. Nat. Prod. Res. 2016, 30, 821–825. [Google Scholar] [CrossRef]
- Auckloo, B.N.; Pan, C.; Akhter, N.; Wu, B.; Wu, X.; He, S. Stress-Driven Discovery of Novel Cryptic Antibiotics from a Marine Fungus Penicillium sp. BB1122. Front. Microbiol. 2017, 8, 1450. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.F.; Wang, Y.; Zhang, X.L.; Wei, M.Y.; Wang, C.Y.; Shao, C.L. Two Dichlorinated Benzophenone Derivatives from the Soft Coral-Derived Pestalotiopsis sp. Fungus and Their Antibacterial Activity. Chem. Nat. Compd. 2017, 53, 1174–1176. [Google Scholar] [CrossRef]
- Pan, C.; Shi, Y.; Auckloo, B.N.; Hassan, S.S.U.; Akhter, N.; Wang, K.; Ye, Y.; Arthur Chen, C.T.; Tao, X.; Wu, B. Isolation and Antibiotic Screening of Fungi from a Hydrothermal Vent Site and Characterization of Secondary Metabolites from a Penicillium Isolate. Mar. Biotechnol. 2017, 19, 469–479. [Google Scholar] [CrossRef]
- Shi, T.; Qi, J.; Shao, C.L.; Zhao, D.L.; Hou, X.M.; Wang, C.Y. Bioactive Diphenyl Ethers and Isocoumarin Derivatives from a Gorgonian-Derived Fungus Phoma sp. (TA07-1). Mar. Drugs 2017, 15, 146. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liao, Y.; Tang, C.; Huang, X.; Luo, Z.; Chen, J.; Cai, P. Cytotoxic and Antibacterial Compounds from the Coral-Derived Fungus Aspergillus tritici SP2-8-1. Mar. Drugs 2017, 15, 348. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, P.; Maida, I.; Palma, E.F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial Activity of Monoramnholipids Produced by Bacterial Strains Isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Xu, H.Y.; Huang, A.M.; Wang, L.; Wang, Q.; Cao, P.Y.; Yang, P.M. Antibacterial Meroterpenoids from the South China Sea Sponge Dysidea sp. Chem. Pharm. Bull. 2016, 64, 1036–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangamuri, U.; Muvva, V.; Poda, S.; Naragani, K.; Munaganti, R.K.; Chitturi, B.; Yenamandra, V. Bioactive metabolites produced by Streptomyces Cheonanensis VUK-A from Coringa mangrove sediments: Isolation, structure elucidation and bioactivity. 3 Biotech. 2016, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Dalisay, D.S.; Chen, J.; Polishchuck, E.A.; Patrick, B.O.; Narula, G.; Ko, M.; Av-Gay, Y.; Li, H.; Magarvey, N.; et al. Aminorifamycins and Sporalactams Produced in Culture by a Micromonospora sp. Isolated from a Northeastern-Pacific Marine Sediment Are Potent Antibiotics. Org. Lett. 2017, 19, 766–769. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Hu, Y.; Wang, Q.; Zhou, H.; Wang, Y.; Gan, M. Tetrocarcins N and O, glycosidic spirotetronates from a marine-derived Micromonospora sp. identified by PCR-based screening. RSC Adv. 2016, 6, 91773–91778. [Google Scholar] [CrossRef]
- Gui, C.; Zhang, S.; Zhu, X.; Ding, W.; Huang, H.; Gu, Y.C.; Duan, Y.; Ju, J. Antimicrobial Spirotetronate Metabolites from Marine-Derived Micromonospora harpali SCSIO GJ089. J. Nat. Prod. 2017, 80, 1594–1603. [Google Scholar] [CrossRef]
- Okada, M.; Sugita, T.; Wong, C.P.; Wakimoto, T.; Abe, I. Identification of Pyridinium with Three Indole Moieties as an Antimicrobial Agent. J. Nat. Prod. 2017, 80, 1205–1209. [Google Scholar] [CrossRef]
- Zubair, M.S.; Alarif, W.M.; Al-Footy, K.O.; Ph, M.; Ali, M.; Basaif, S.A.; Al-Lihaibi, S.S.; Ayyad, S.E. New antimicrobial biscembrane hydrocarbon and cembranoid diterpenes from the soft coral Sarcophyton trocheliophorum. Turk. J. Chem. 2016, 40, 385–392. [Google Scholar] [CrossRef]
- Liu, H.B.; Lauro, G.; O’Connor, R.D.; Lohith, K.; Kelly, M.; Colin, P.; Bifulco, G.; Bewley, C.A. Tulongicin, an Antibacterial Tri-Indole Alkaloid from a Deep-Water Topsentia sp. Sponge. J. Nat. Prod. 2017, 80, 2556–2560. [Google Scholar] [CrossRef]
- Hu, Z.; Qin, L.; Wang, Q.; Ding, W.; Chen, Z.; Ma, Z. Angucycline antibiotics and its derivatives from marine-derived actinomycete Streptomyces sp. A6H. Nat. Prod. Res. 2016, 30, 2551–2558. [Google Scholar] [CrossRef]
- Espiritu, R.A. Membrane permeabilizing action of amphidinol 3 and theonellamide A in raft-forming lipid mixtures. Z. Naturforsch. C 2017, 72, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Sumino, A.; Shimada, E.; Kinoshita, M.; Matsumori, N.; Oiki, S. Channel Formation and Membrane Deformation via Sterol-Aided Polymorphism of Amphidinol 3. Sci. Rep 2017, 7, 10782. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Ciric, A.; Markovic, D.; Tommonaro, G.; Sokovic, M. In vitro avarol does affect the growth of Candida sp. Nat. Prod. Res. 2016, 30, 1956–1960. [Google Scholar] [CrossRef] [PubMed]
- Quezada, M.; Licona-Cassani, C.; Cruz-Morales, P.; Salim, A.A.; Marcellin, E.; Capon, R.J.; Barona-Gomez, F. Diverse Cone-Snail Species Harbor Closely Related Streptomyces Species with Conserved Chemical and Genetic Profiles, Including Polycyclic Tetramic Acid Macrolactams. Front. Microbiol. 2017, 8, 2305. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.H.; Hong, L.L.; Sun, J.B.; Piao, S.J.; Chen, G.D.; Deng, H.; Wang, S.P.; Yang, F.; Lin, H.W. (±)-Hippolide J—A pair of unusual antifungal enantiomeric sesterterpenoids from the marine sponge Hippospongia lachne. Eur. J. Org. Chem 2017, 2017, 3421–3426. [Google Scholar] [CrossRef]
- Kildgaard, S.; Subko, K.; Phillips, E.; Goidts, V.; de la Cruz, M.; Diaz, C.; Gotfredsen, C.H.; Andersen, B.; Frisvad, J.C.; Nielsen, K.F.; et al. A Dereplication and Bioguided Discovery Approach to Reveal New Compounds from a Marine-Derived Fungus Stilbella fimetaria. Mar. Drugs 2017, 15, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, S.; Ko, S.K.; Jang, M.; Kim, J.W.; Kim, G.S.; Lee, J.K.; Jeon, E.S.; Futamura, Y.; Ryoo, I.J.; Lee, J.S.; et al. New Cyclic Lipopeptides of the Iturin Class Produced by Saltern-Derived Bacillus sp. KCB14S006. Mar. Drugs 2016, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Okabe, M.; Sugita, T.; Kinoshita, K.; Koyama, K. Macrolides from a Marine-Derived Fungus, Penicillium meleagrinum var. viridiflavum, Showing Synergistic Effects with Fluconazole against Azole-Resistant Candida albicans. J. Nat. Prod. 2016, 79, 1208–1212. [Google Scholar] [CrossRef]
- Jamison, M.T.; Dalisay, D.S.; Molinski, T.F. Peroxide Natural Products from Plakortis zyggompha and the Sponge Association Plakortis halichondrioides-Xestospongia deweerdtae: Antifungal Activity against Cryptococcus gattii. J. Nat. Prod. 2016, 79, 555–563. [Google Scholar] [CrossRef]
- Calabro, K.; Kalahroodi, E.L.; Rodrigues, D.; Diaz, C.; Cruz, M.; Cautain, B.; Laville, R.; Reyes, F.; Perez, T.; Soussi, B.; et al. Poecillastrosides, Steroidal Saponins from the Mediterranean Deep-Sea Sponge Poecillastra compressa (Bowerbank, 1866). Mar. Drugs 2017, 15, 199. [Google Scholar] [CrossRef] [Green Version]
- Aksoy, S.C.; Uzel, A.; Bedir, E. Cytosine-type nucleosides from marine-derived Streptomyces rochei 06CM016. J. Antibiot. 2016, 69, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, R.P.; Xu, B.; Yu, H.B.; Ma, G.Y.; Wang, G.F.; Dai, S.W.; Zhang, W.; Jiao, W.H.; Song, S.J.; et al. New antimalarial norterpene cyclic peroxides from Xisha Islands sponge Diacarnus megaspinorhabdosa. Bioorg. Med. Chem. Lett. 2016, 26, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Almaliti, J.; Malloy, K.L.; Glukhov, E.; Spadafora, C.; Gutierrez, M.; Gerwick, W.H. Dudawalamides A-D, Antiparasitic Cyclic Depsipeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2017, 80, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.T.; Nani, R.R.; Schauer, E.A.; Martin, G.E.; Williamson, R.T.; Sauri, J.; Buevich, A.V.; Schafer, W.A.; Joyce, L.A.; Goey, A.K.; et al. Characterization and Synthesis of Eudistidine C, a Bioactive Marine Alkaloid with an Intriguing Molecular Scaffold. J. Org. Chem. 2016, 81, 10631–10640. [Google Scholar] [CrossRef] [PubMed]
- Buedenbender, L.; Grkovic, T.; Duffy, S.; Kurtboke, D.I.; Avery, V.K.; Carroll, A.R. Naseseazine C, a new anti-plasmodial dimeric diketopiperazine from a marine sediment derived Streptomyces sp. Tetrahedrom Lett. 2016, 57, 5893–5895. [Google Scholar] [CrossRef]
- Wang, J.; Pearce, A.N.; Chan, S.T.; Taylor, R.B.; Page, M.J.; Valentin, A.; Bourguet-Kondracki, M.L.; Dalton, J.P.; Wiles, S.; Copp, B.R. Biologically Active Acetylenic Amino Alcohol and N-Hydroxylated 1,2,3,4-Tetrahydro-beta-carboline Constituents of the New Zealand Ascidian Pseudodistoma opacum. J. Nat. Prod. 2016, 79, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.E.; Wolfender, J.L.; Queiroz, E.F.; Marcourt, L.; Al-Mourabit, A.; Frederich, M.; Bordignon, A.; De Voogd, N.; Illien, B.; Gauvin-Bialecki, A. Unguiculin A and Ptilomycalins E-H, Antimalarial Guanidine Alkaloids from the Marine Sponge Monanchora unguiculata. J. Nat. Prod. 2017, 80, 1404–1410. [Google Scholar] [CrossRef]
- White, A.M.; Dao, K.; Vrubliauskas, D.; Konst, Z.A.; Pierens, G.K.; Mandi, A.; Andrews, K.T.; Skinner-Adams, T.S.; Clarke, M.E.; Narbutas, P.T.; et al. Catalyst-Controlled Stereoselective Synthesis Secures the Structure of the Antimalarial Isocyanoterpene Pustulosaisonitrile-1. J. Org. Chem. 2017, 82, 13313–13323. [Google Scholar] [CrossRef]
- Carballeira, N.M.; Montano, N.; Amador, L.A.; Rodriguez, A.D.; Golovko, M.Y.; Golovko, S.A.; Reguera, R.M.; Alvarez-Velilla, R.; Balana-Fouce, R. Novel Very Long-Chain alpha-Methoxylated Delta5,9 Fatty Acids from the Sponge Asteropus niger Are Effective Inhibitors of Topoisomerases IB. Lipids 2016, 51, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Cardoso-Martinez, F.; de la Rosa, J.M.; Diaz-Marrero, A.R.; Darias, J.; D’Croz, L.; Jimenez-Anton, M.D.; Corral, M.J.; Garcia, R.; Alunda, J.M.; Cueto, M. Oxysterols from an octocoral of the genus Gorgonia from the eastern Pacific of Panama. RSC Adv. 2016, 6, 38579–38591. [Google Scholar] [CrossRef] [Green Version]
- Chianese, G.; Silber, J.; Luciano, P.; Merten, C.; Erpenbeck, D.; Topaloglu, B.; Kaiser, M.; Tasdemir, D. Antiprotozoal Linear Furanosesterterpenoids from the Marine Sponge Ircinia oros. J. Nat. Prod. 2017, 80, 2566–2571. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Iwasaki, A.; Sumimoto, S.; Kanamori, Y.; Ohno, O.; Iwatsuki, M.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Omura, S.; et al. Janadolide, a Cyclic Polyketide-Peptide Hybrid Possessing a tert-Butyl Group from an Okeania sp. Marine Cyanobacterium. J. Nat. Prod. 2016, 79, 1862–1866. [Google Scholar] [CrossRef] [PubMed]
- Notarte, K.; Nakao, Y.; Yaguchi, T.; Bungihan, M.; Suganuma, K.; Delacruz, T. Trypanocidal activity, cytotoxicity and histone modifications induced by malformin A1 isolated from the marine-derived fungus Aspergillus tubingensis IFM 63452. Mycosphere 2017, 8, 111–120. [Google Scholar] [CrossRef]
- Elsayed, Y.; Refaat, J.; Abdelmohsen, U.; Ahmed, S.; Fouad, M. Rhodozepinone, a new antitrypanosomal azepino-diindole alkaloid from the marine sponge-derived bacterium Rhodococcus sp. UA13. Med. Chem. Res. 2017, 26, 2751–2760. [Google Scholar] [CrossRef]
- Arai, M.; Yamano, Y.; Kamiya, K.; Setiawan, A.; Kobayashi, M. Anti-dormant mycobacterial activity and target molecule of melophlins, tetramic acid derivatives isolated from a marine sponge of Melophlus sp. J. Nat. Med. 2016, 70, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.C.; Gupta, R.; Geden, S.; Roberts, J.; Winder, P.; Pomponi, S.A.; Diaz, M.C.; Reed, J.K.; Wright, A.E.; Rohde, K.H. Selective Killing of Dormant Mycobacterium tuberculosis by Marine Natural Products. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Zhou, X.; Lin, X.; Qin, X.; Zhang, T.; Wang, J.; Tu, Z.; Yang, B.; Liao, S.; Tian, Y.; et al. Antituberculosis compounds from a deep-sea-derived fungus Aspergillus sp. SCSIO Ind09F01. Nat. Prod. Res. 2017, 31, 1958–1962. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Xie, F.; Ren, B.; Wang, Q.; Wang, J.; Wang, Q.; Abdel-Mageed, W.M.; Liu, M.; Han, J.; Oyeleye, A.; et al. Anti-MRSA and anti-TB metabolites from marine-derived Verrucosispora sp. MS100047. Appl. Microbiol. Biotechnol. 2016, 100, 7437–7447. [Google Scholar] [CrossRef]
- Jimenez-Romero, C.; Rode, J.E.; Perez, Y.M.; Franzblau, S.G.; Rodriguez, A.D. Exploring the Sponge Consortium Plakortis symbiotica-Xestospongia deweerdtae as a Potential Source of Antimicrobial Compounds and Probing the Pharmacophore for Antituberculosis Activity of Smenothiazole A by Diverted Total Synthesis. J. Nat. Prod. 2017, 80, 2295–2303. [Google Scholar] [CrossRef]
- Chen, S.; He, L.; Chen, D.; Cai, R.; Long, Y.; Lu, Y.; She, Z. Talaramide A, an unusual alkaloid from the mangrove endophytic fungus Talaromyces sp. (HZ-YX1) as an inhibitor of mycobacterial PknG. New J. Chem. 2017, 41, 4273–4276. [Google Scholar] [CrossRef]
- Kamiya, K.A.; Setiawan, M.; Kobayashi, A.M. Anti-dormant mycobacterial activity of viomellein and xanthomegnin, naphthoquinone dimers produced by marine-derived Aspergillus sp. Nat. Prod. Commun. 2017, 12, 579–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, A.; Kremb, S.; Bader, T.M.; Helfer, M.; Schmitt-Kopplin, P.; Gerwick, W.H.; Brack-Werner, R.; Voolstra, C.R. Alkaloids from the Sponge Stylissa carteri Present Prospective Scaffolds for the Inhibition of Human Immunodeficiency Virus 1 (HIV-1). Mar. Drugs 2016, 14, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, A.; Tamaki, M.; Kasai, H.; Tanaka, T.; Otoguro, T.; Ryo, A.; Maekawa, S.; Enomoto, N.; de Voogd, N.J.; Tanaka, J.; et al. Inhibitory effects of metachromin A on hepatitis B virus production via impairment of the viral promoter activity. Antiviral. Res. 2017, 145, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, C.; Jomori, T.; Tanaka, J.; Senba, M.; Mori, N. Peridinin, a carotenoid, inhibits proliferation and survival of HTLV-1-infected T-cell lines. Int. J. Oncol. 2016, 49, 1713–1721. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Si, L.; Liu, D.; Zhou, A.; Zhang, Z.; Shao, Z.; Wang, S.; Zhang, L.; Zhou, D.; Lin, W. Spiromastilactones: A new class of influenza virus inhibitors from deep-sea fungus. Eur. J. Med. Chem. 2016, 108, 229–244. [Google Scholar] [CrossRef]
- Kim, S.H.; Ha, T.K.; Oh, W.K.; Shin, J.; Oh, D.C. Antiviral Indolosesquiterpenoid Xiamycins C-E from a Halophilic Actinomycete. J. Nat. Prod. 2016, 79, 51–58. [Google Scholar] [CrossRef]
- Cheng, Y.B.; Lee, J.C.; Lo, I.W.; Chen, S.R.; Hu, H.C.; Wu, Y.H.; Wu, Y.C.; Chang, F.R. Ecdysones from Zoanthus spp. with inhibitory activity against dengue virus 2. Bioorg. Med. Chem. Lett. 2016, 26, 2344–2348. [Google Scholar] [CrossRef] [Green Version]
- Hermawan, I.; Furuta, A.; Higashi, M.; Fujita, Y.; Akimitsu, N.; Yamashita, A.; Moriishi, K.; Tsuneda, S.; Tani, H.; Nakakoshi, M.; et al. Four Aromatic Sulfates with an Inhibitory Effect against HCV NS3 Helicase from the Crinoid Alloeocomatella polycladia. Mar. Drugs 2017, 15, 117. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Tietjen, I.; Chen, M.; Williams, D.E.; Daoust, J.; Brockman, M.A.; Andersen, R.J. Sesterterpenoids Isolated from the Sponge Phorbas sp. Activate Latent HIV-1 Provirus Expression. J. Org. Chem. 2016, 81, 11324–11334. [Google Scholar] [CrossRef]
- Ma, X.; Nong, X.H.; Ren, Z.; Wang, J.; Liang, X.; Wang, L.; Qi, S.H. Antiviral peptides from marine gorgonian-derived fungus Aspergillus sp. SCSIO 41501. Tetrahedron 2017, 58, 1151–1155. [Google Scholar] [CrossRef]
- Huang, Z.; Nong, X.; Ren, Z.; Wang, J.; Zhang, X.; Qi, S. Anti-HSV-1, antioxidant and antifouling phenolic compounds from the deep-sea-derived fungus Aspergillus versicolor SCSIO 41502. Bioorg. Med. Chem. Lett. 2017, 27, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.Q.; Lin, X.P.; Wang, Z.; Zhou, X.F.; Qin, X.C.; Kaliyaperumal, K.; Zhang, T.Y.; Tu, Z.C.; Liu, Y. Asteltoxins with Antiviral Activities from the Marine Sponge-Derived Fungus Aspergillus sp. SCSIO XWS02F40. Molecules 2016, 21, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, W.; Cheng, S.; Fu, W.; Zhao, M.; Li, X.; Cai, Y.; Dong, J.; Huang, K.; Gustafson, K.R.; Yan, P. Structurally Diverse Metabolites from the Soft Coral Sinularia verruca Collected in the South China Sea. J. Nat. Prod. 2016, 79, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eutypellazines A-M, thiodiketopiperazine-type alkaloids from deep sea derived fungus Eutypella sp. MCCC 3A00281. RSC Adv. 2017, 7, 33580–33590. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; Al-Farawati, R.; Abou El-Kassem, L.T.; Turki, A.J. Different Culture Metabolites of the Red Sea Fungus Fusarium equiseti Optimize the Inhibition of Hepatitis C Virus NS3/4A Protease (HCV PR). Mar. Drugs 2016, 14, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Fang, W.; Tan, S.; Lin, X.; Xun, T.; Yang, B.; Liu, S.; Liu, Y. Aspernigrins with anti-HIV-1 activities from the marine-derived fungus Aspergillus niger SCSIO Jcsw6F30. Bioorg. Med. Chem. Lett. 2016, 26, 361–365. [Google Scholar] [CrossRef]
- Palem, J.R.; Mudit, M.; Hsia, S.V.; Sayed, K.A. Discovery and preliminary structure-activity relationship of the marine natural product manzamines as herpes simplex virus type-1 inhibitors. Z. Naturforsch. C 2017, 72, 49–54. [Google Scholar] [CrossRef]
- Lee, J.C.; Chang, F.R.; Chen, S.R.; Wu, Y.H.; Hu, H.C.; Wu, Y.C.; Backlund, A.; Cheng, Y.B. Anti-Dengue Virus Constituents from Formosan Zoanthid Palythoa mutuki. Mar. Drugs 2016, 14, 151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, Y.; Jia, C.; Lang, J.; Niaz, S.I.; Yuan, J.; Yu, J.; Chen, S.; Liu, L. Antiviral and anti-inflammatory meroterpenoids: Stachybonoids A–F from the crinoid-derived fungus Stachybotrys chartarum 952. RSC Adv. 2017, 7, 49910–49916. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Ren, J.; Huang, Q.; Long, H.; Jin, H.; Zhang, L.; Liu, H.; Van, O.L.; Lin, W. Pregnane steroids from a gorgonian coral Subergorgia suberosa with anti-flu virus effects. Steroids 2016, 108, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Abdjul, D.B.; Yamazaki, H.; Kanno, S.; Takahashi, O.; Kirikoshi, R.; Ukai, K.; Namikoshi, M. Haliclonadiamine Derivatives and 6-epi-Monanchorin from the Marine Sponge Halichondria panicea Collected at Iriomote Island. J. Nat. Prod. 2016, 79, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Al-Footy, K.O.; Alarif, W.M.; Zubair, M.S.; Ghandourah, M.A.; Aly, M.M. Antibacterial and cytotoxic properties of isoprenoids from the red sea soft coral, Lobophytum sp. Trop. J. Pharm. Res. 2016, 15, 1431–1438. [Google Scholar] [CrossRef] [Green Version]
- Angulo-Preckler, C.; Genta-Jouve, G.; Mahajan, N.; de la Cruz, M.; de Pedro, N.; Reyes, F.; Iken, K.; Avila, C.; Thomas, O.P. Gersemiols A-C and Eunicellol A, Diterpenoids from the Arctic Soft Coral Gersemia fruticosa. J. Nat. Prod. 2016, 79, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Phan, C.S.; Tin, H.S.; Vairappan, C.S.; Muhammad, T.S.T. 16-Hydroxycembra-1,3,7,11-tetraene, a new Cembrane Diterpene from Malaysian Soft Coral Genus Sarcophyton. Nat. Prod. Commun. 2016, 11, 1077–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, T.; Nakamura, K.; Sakai, K.; Fromont, J.; Gonoi, T.; Kobayashi, J. Hyrtinadines C and D, New Azepinoindole-Type Alkaloids from a Marine Sponge Hyrtios sp. Chem. Pharm. Bull. 2016, 64, 975–978. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; La, M.P.; Tang, H.; Sun, P.; Liu, B.S.; Zhuang, C.L.; Yi, Y.H.; Zhang, W. Chemistry and Bioactivity of Briaranes from the South China Sea Gorgonian Dichotella gemmacea. Mar. Drugs 2016, 14, 201. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.M.; Ito, T.; Win, N.N.; Kodama, T.; Hung, V.Q.; Nguyen, H.T.; Morita, H. New antibacterial sesquiterpene aminoquinones from a Vietnamese marine sponge of Spongia sp. Phytochem. Lett. 2016, 17, 288–292. [Google Scholar] [CrossRef]
- Phan, C.S.; Ng, S.Y.; Kamada, T.; Vairappan, C.S. Two New Lobane Diterpenes from a Bornean Soft Coral Sinularia sp. Nat. Prod. Commun. 2016, 11, 899–900. [Google Scholar] [CrossRef] [Green Version]
- Von Salm, J.L.; Witowski, C.G.; Fleeman, R.M.; McClintock, J.B.; Amsler, C.D.; Shaw, L.N.; Baker, B.J. Darwinolide, a New Diterpene Scaffold That Inhibits Methicillin-Resistant Staphylococcus aureus Biofilm from the Antarctic Sponge Dendrilla membranosa. Org. Lett. 2016, 18, 2596–2599. [Google Scholar] [CrossRef]
- Brasseur, L.; Hennebert, E.; Fievez, L.; Caulier, G.; Bureau, F.; Tafforeau, L.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. The Roles of Spinochromes in Four Shallow Water Tropical Sea Urchins and Their Potential as Bioactive Pharmacological Agents. Mar. Drugs 2017, 15, 179. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.D.; Ma, Q.Y.; Huang, S.Z.; Wang, P.; Wang, J.F.; Zhou, L.M.; Yuan, J.Z.; Dai, H.F.; Zhao, Y.X. Chrodrimanins K-N and Related Meroterpenoids from the Fungus Penicillium sp. SCS-KFD09 Isolated from a Marine Worm, Sipunculus nudus. J. Nat. Prod. 2017, 80, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, R.; Radwan, M.; Ma, G.; Mohamed, T.A.; Seliem, M.A.; Thabet, M.; ElSohly, M.A. Bioactive sterols and sesquiterpenes from the Red Sea soft coral Sinularia terspilli. Med. Chem. Res. 2017, 26, 1647–1652. [Google Scholar] [CrossRef]
- Liang, X.R.; Miao, F.P.; Song, Y.P.; Liu, X.H.; Ji, N.Y. Citrinovirin with a new norditerpene skeleton from the marine algicolous fungus Trichoderma citrinoviride. Bioorg. Med. Chem. Lett. 2016, 26, 5029–5031. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Dai, H.; Konuklugil, B.; Orfali, R.S.; Lin, W.; Kalscheuer, R.; Liu, Z.; Proksch, P. Phenolic bisabolanes from the sponge-derived fungus Aspergillus sp. Phytochem. Lett. 2016, 18, 187–191. [Google Scholar] [CrossRef]
- Mohamed-Benkada, M.; Francois, P.Y.; Verite, P.; Pagniez, F.; Caroff, N.; Ruiz, N. Identification and Biological Activities of Long-Chain Peptaibols Produced by a Marine-Derived Strain of Trichoderma longibrachiatum. Chem. Biodivers. 2016, 13, 521–530. [Google Scholar] [CrossRef]
- Sun, Y.; Feng, Z.; Tomura, T.; Suzuki, A.; Miyano, S.; Tsuge, T.; Mori, H.; Suh, J.W.; Iizuka, T.; Fudou, R.; et al. Heterologous Production of the Marine Myxobacterial Antibiotic Haliangicin and Its Unnatural Analogues Generated by Engineering of the Biochemical Pathway. Sci. Rep. 2016, 6, 22091. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Anjum, K.; Song, T.; Wang, W.; Yu, S.; Huang, H.; Lian, X.Y.; Zhang, Z. A new curvularin glycoside and its cytotoxic and antibacterial analogues from marine actinomycete Pseudonocardia sp. HS7. Nat. Prod. Res. 2016, 30, 1156–1161. [Google Scholar] [CrossRef]
- Zhang, Y.; Adnani, N.; Braun, D.R.; Ellis, G.A.; Barns, K.J.; Parker-Nance, S.; Guzei, I.A.; Bugni, T.S. Micromonohalimanes A and B: Antibacterial Halimane-Type Diterpenoids from a Marine Micromonospora Species. J. Nat. Prod. 2016, 79, 2968–2972. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.L.; Wang, M.; Zhao, H.G.; Huang, Y.H.; Lin, Y.Y.; Tan, G.H.; Chen, S.L. Penicilazaphilone C, a new antineoplastic and antibacterial azaphilone from the Marine Fungus Penicillium sclerotiorum. Arch. Pharm. Res. 2016, 39, 1621–1627. [Google Scholar] [CrossRef]
- He, K.Y.; Zhang, C.; Duan, Y.R.; Huang, G.L.; Yang, C.Y.; Lu, X.R.; Zheng, C.J.; Chen, G.Y. New chlorinated xanthone and anthraquinone produced by a mangrove-derived fungus Penicillium citrinum HL-5126. J. Antibiot. 2017, 70, 823–827. [Google Scholar] [CrossRef]
- Kim, J.; Shin, D.; Kim, S.H.; Park, W.; Shin, Y.; Kim, W.K.; Lee, S.K.; Oh, K.B.; Shin, J.; Oh, D.C. Borrelidins C-E: New Antibacterial Macrolides from a Saltern-Derived Halophilic Nocardiopsis sp. Mar. Drugs 2017, 15, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.L.; Chen, D.; Huang, L.; Ni, M.; Zhao, Y.; Fan, H.; Bao, X. Antichlamydial Dimeric Indole Derivatives from Marine Actinomycete Rubrobacter radiotolerans. Planta Med. 2017, 83, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Zin, W.W.M.; Buttachon, S.; Dethoup, T.; Pereira, J.A.; Gales, L.; Inacio, A.; Costa, P.M.; Lee, M.; Sekeroglu, N.; Silva, A.M.S.; et al. Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 0006. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eutypellazines N-S, new thiodiketopiperazines from a deep sea sediment derived fungus Eutypella sp. with anti-VRE activities. Tetrahedron Lett. 2017, 58, 3695–3699. [Google Scholar] [CrossRef]
- Teta, R.; Marteinsson, V.T.; Longeon, A.; Klonowski, A.M.; Groben, R.; Bourguet-Kondracki, M.L.; Costantino, V.; Mangoni, A. Thermoactinoamide A, an Antibiotic Lipophilic Cyclopeptide from the Icelandic Thermophilic Bacterium Thermoactinomyces vulgaris. J. Nat. Prod. 2017, 80, 2530–2535. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wang, M.; Tan, Y.; Hu, X.; He, H.; Xiao, C.; You, X.; Wang, Y.; Gan, M. Neo-actinomycins A and B, natural actinomycins bearing the 5H-oxazolo[4,5-b]phenoxazine chromophore, from the marine-derived Streptomyces sp. IMB094. Sci. Rep. 2017, 7, 3591. [Google Scholar] [CrossRef] [Green Version]
- Jamison, M.T.; Macho, J.; Molinski, T.F. Structure-activity of antifungal compounds inspired by aminobisabolenes from the sponge Halichondria sp. Bioorg. Med. Chem. Lett. 2016, 26, 5244–5246. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Gu, B.; Yang, F.; Jiao, W.H.; Hu, G.; Yu, H.B.; Han, B.N.; Zhang, W.; Shen, Y.; et al. Antifungal bromopyrrole alkaloids from the South China Sea sponge Agelas sp. Tetrahedron 2016, 72, 2964–2971. [Google Scholar] [CrossRef]
- Bertin, M.J.; Demirkiran, O.; Navarro, G.; Moss, N.A.; Lee, J.; Goldgof, G.M.; Vigil, E.; Winzeler, E.A.; Valeriote, F.A.; Gerwick, W.H. Kalkipyrone B, a marine cyanobacterial gamma-pyrone possessing cytotoxic and anti-fungal activities. Phytochemistry 2016, 122, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, H.; Kanno, S.I.; Abdjul, D.B.; Namikoshi, M. A bromopyrrole-containing diterpene alkaloid from the Okinawan marine sponge Agelas nakamurai activates the insulin pathway in Huh-7 human hepatoma cells by inhibiting protein tyrosine phosphatase 1B. Bioorg. Med. Chem. Lett. 2017, 27, 2207–2209. [Google Scholar] [CrossRef]
- Xu, F.; Wang, F.; Wang, Z.; Lv, W.; Wang, W.; Wang, Y. Glucose Uptake Activities of Bis (2, 3-Dibromo-4, 5-Dihydroxybenzyl) Ether, a Novel Marine Natural Product from Red Alga Odonthaliacorymbifera with Protein Tyrosine Phosphatase 1B Inhibition, In Vitro and In Vivo. PLoS ONE 2016, 11, e0147748. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.A.; Lee, S.H.; Lee, J.H.; Kang, N.; Oh, J.Y.; Seun-heui, S.; Ahn, G.; Ko, S.C.; Fernando, S.P.; Kim, S.Y.; et al. A marine algal polyphenol, dieckol, attenuates blood glucose levels by Akt pathway in alloxan induced hyperglycemia zebrafish model. RSC Adv. 2016, 6, 78570–78575. [Google Scholar] [CrossRef]
- Woo, J.K.; Ha, T.K.Q.; Oh, D.C.; Oh, W.K.; Oh, K.B.; Shin, J. Polyoxygenated Steroids from the Sponge Clathria gombawuiensis. J. Nat. Prod. 2017, 80, 3224–3233. [Google Scholar] [CrossRef]
- Villa-Perez, P.; Cueto, M.; Diaz-Marrero, A.R.; Lobaton, C.D.; Moreno, A.; Perdomo, G.; Cozar-Castellano, I. Leptolide Improves Insulin Resistance in Diet-Induced Obese Mice. Mar. Drugs 2017, 15, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Liu, Y.; Nie, Y.; Liu, Z.; Chen, S.; Zhang, Z.; Lu, Y.; He, L.; Huang, X.; She, Z. Polyketides from the Mangrove-Derived Endophytic Fungus Nectria sp. HN001 and Their alpha-Glucosidase Inhibitory Activity. Mar. Drugs 2016, 14, 86. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Wu, Q.; Yu, Y.; Liang, Z.; Liu, Y.; Zhou, L.; Tang, L.; Zhou, X. Penicilliumin B, a novel sesquiterpene methylcyclopentenedione from a deep sea-derived Penicillium strain with renoprotective activities. Sci. Rep. 2017, 7, 10757. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Hao, J.; Wang, L.; Wang, Y.; Kong, F.; Zhu, W. New alpha-glucosidase inhibitors from marine algae-derived Streptomyces sp. OUCMDZ-3434. Sci. Rep. 2016, 6, 20004. [Google Scholar] [CrossRef]
- Wiese, J.; Aldemir, H.; Schmaljohann, R.; Gulder, T.A.M.; Imhoff, J.F. Asperentin B, a New Inhibitor of the Protein Tyrosine Phosphatase 1B. Mar. Drugs 2017, 15, 191. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Z.; Liu, H.; Long, Y.; Chen, D.; Lu, Y.; She, Z. Lasiodiplactone A, a novel lactone from the mangrove endophytic fungus Lasiodiplodia theobromae ZJ-HQ1. Org. Biomol. Chem. 2017, 15, 6338–6341. [Google Scholar] [CrossRef]
- Zhang, L.; Niaz, S.I.; Khan, D.; Wang, Z.; Zhu, Y.; Zhou, H.; Lin, Y.; Li, J.; Liu, L. Induction of Diverse Bioactive Secondary Metabolites from the Mangrove Endophytic Fungus Trichoderma sp. (Strain 307) by Co-Cultivation with Acinetobacter johnsonii (Strain B2). Mar. Drugs 2017, 15, 35. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Liu, Y.; Liu, Z.; Cai, R.; Lu, Y.; Huang, X.; She, Z. Isocoumarins and benzofurans from the mangrove endophytic fungus Talaromyces amestolkiae possess a-glucosidase inhibitory and antibacterial activities. RSC Adv. 2016, 6, 26412–26420. [Google Scholar] [CrossRef]
- Zbakh, H.; Talero, E.; Avila, J.; Alcaide, A.; de Los, R.C.; Zubia, E.; Motilva, V. The Algal Meroterpene 11-Hydroxy-1′-O-Methylamentadione Ameloriates Dextran Sulfate Sodium-Induced Colitis in Mice. Mar. Drugs 2016, 14, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, B.; Ku, S.K.; Gao, M.; Kim, K.M.; Han, M.S.; Choi, H.; Bae, J.S. Suppressive effects of three diketopiperazines from marine-derived bacteria on TGFBIp-mediated septic responses in human endothelial cells and mice. Arch. Pharm. Res. 2016, 39, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Ku, S.K.; Choi, H.; Bae, J.S. Three diketopiperazines from marine-derived bacteria inhibit LPS-induced endothelial inflammatory responses. Bioorg. Med. Chem. Lett. 2016, 26, 1873–1876. [Google Scholar] [CrossRef]
- Ahmad, T.B.; Rudd, D.; Benkendorff, K.; Mahdi, L.K.; Pratt, K.A.; Dooley, L.; Wei, C.; Kotiw, M. Brominated indoles from a marine mollusc inhibit inflammation in a murine model of acute lung injury. PLoS ONE 2017, 12, e0186904. [Google Scholar] [CrossRef] [Green Version]
- El-Desoky, A.H.; Kato, H.; Angkouw, E.D.; Mangindaan, R.E.; de Voogd, N.J.; Tsukamoto, S. Ceylonamides A-F, Nitrogenous Spongian Diterpenes That Inhibit RANKL-Induced Osteoclastogenesis, from the Marine Sponge Spongia ceylonensis. J. Nat. Prod. 2016, 79, 1922–1928. [Google Scholar] [CrossRef]
- Ngan, N.T.; Quang, T.H.; Kim, K.W.; Kim, H.J.; Sohn, J.H.; Kang, D.G.; Lee, H.S.; Kim, Y.C.; Oh, H. Anti-inflammatory effects of secondary metabolites isolated from the marine-derived fungal strain Penicillium sp. SF-5629. Arch. Pharm. Res. 2017, 40, 328–337. [Google Scholar] [CrossRef]
- Manzoor, Z.; Koo, J.E.; Ali, I.; Kim, J.E.; Byeon, S.H.; Yoo, E.S.; Kang, H.K.; Hyun, J.W.; Lee, N.H.; Koh, Y.S. 4-Hydroxy-2,3-Dimethyl-2-Nonen-4-Olide Has an Inhibitory Effect on Pro-Inflammatory Cytokine Production in CpG-Stimulated Bone Marrow-Derived Dendritic Cells. Mar. Drugs 2016, 14, 88. [Google Scholar] [CrossRef] [Green Version]
- Aminin, D.; Pislyagin, E.; Astashev, M.; Es’kov, A.; Kozhemyako, V.; Avilov, S.; Zelepuga, E.; Yurchenko, E.; Kaluzhskiy, L.; Kozlovskaya, E.; et al. Glycosides from edible sea cucumbers stimulate macrophages via purinergic receptors. Sci. Rep. 2016, 6, 39683. [Google Scholar] [CrossRef] [Green Version]
- Ha, T.M.; Ko, W.; Lee, S.J.; Kim, Y.C.; Son, J.Y.; Sohn, J.H.; Yim, J.H.; Oh, H. Anti-Inflammatory Effects of Curvularin-Type Metabolites from a Marine-Derived Fungal Strain Penicillium sp. SF-5859 in Lipopolysaccharide-Induced RAW264.7 Macrophages. Mar. Drugs 2017, 15, 282. [Google Scholar] [CrossRef] [Green Version]
- Ciaglia, E.; Malfitano, A.M.; Laezza, C.; Fontana, A.; Nuzzo, G.; Cutignano, A.; Abate, M.; Pelin, M.; Sosa, S.; Bifulco, M.; et al. Immuno-Modulatory and Anti-Inflammatory Effects of Dihydrogracilin A, a Terpene Derived from the Marine Sponge Dendrilla membranosa. Int. J. Mol. Sci. 2017, 18, 1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, W.H.; Cheng, B.H.; Shi, G.H.; Chen, G.D.; Gu, B.B.; Zhou, Y.J.; Hong, L.L.; Yang, F.; Liu, Z.Q.; Qiu, S.Q.; et al. Dysivillosins A-D, Unusual Anti-allergic Meroterpenoids from the Marine Sponge Dysidea villosa. Sci. Rep. 2017, 7, 8947. [Google Scholar] [CrossRef] [PubMed]
- Su, B.C.; Chen, J.Y. Antimicrobial Peptide Epinecidin-1 Modulates MyD88 Protein Levels via the Proteasome Degradation Pathway. Mar. Drugs 2017, 15, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.Y.; Jean, Y.H.; Lee, H.P.; Lin, S.C.; Pan, C.Y.; Chen, W.F.; Wu, S.F.; Su, J.H.; Tsui, K.H.; Sheu, J.H.; et al. Excavatolide B Attenuates Rheumatoid Arthritis through the Inhibition of Osteoclastogenesis. Mar. Drugs 2017, 15, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Kim, N.H.; Kim, S.J.; Lee, H.J.; Kim, S. Fucoxanthin Inhibits the Inflammation Response in Paw Edema Model through Suppressing MAPKs, Akt, and NFkappaB. J. Biochem. Mol. Toxicol. 2016, 30, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Grasa-Lopez, A.; Miliar-Garcia, A.; Quevedo-Corona, L.; Paniagua-Castro, N.; Escalona-Cardoso, G.; Reyes-Maldonado, E.; Jaramillo-Flores, M.E. Undaria pinnatifida and Fucoxanthin Ameliorate Lipogenesis and Markers of Both Inflammation and Cardiovascular Dysfunction in an Animal Model of Diet-Induced Obesity. Mar. Drugs 2016, 14, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, Y.; Kinoshita, Y.; USUI, M.; Tanaka, R.; Matsushita, T.; Masaaki, M. The Suppressive Effect of a Marine Carotenoid, Fucoxanthin, on Mouse Ear Swelling through Regulation of Activities and mRNA Expression of Inflammation-associated Enzymes. Food Sci. Technol. Res. 2016, 22, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Taira, J.; Sonamoto, M.; Uehara, M. Dual Biological Functions of a Cytoprotective Effect and Apoptosis Induction by Bioavailable Marine Carotenoid Fucoxanthinol through Modulation of the Nrf2 Activation in RAW264.7 Macrophage Cells. Mar. Drugs 2017, 15, 305. [Google Scholar] [CrossRef] [Green Version]
- Sinstova, O.V.; Pislygin, E.A.; Gladkikh, I.N.; Monastyrnaya, M.M.; Menchinskaya, E.S.; Lynchenko, E.V.; Aminin, D.L.; Kozlovskaya, E.P. Kunitz-type peptides of the sea anemone Heteractis crispa: Potential anti-inflammatory compounds. Russ. J. Bioorg. Chem. 2017, 43, 91–97. [Google Scholar]
- Hong, L.L.; Yu, H.B.; Wang, J.; Jiao, W.H.; Cheng, B.H.; Yang, F.; Zhou, Y.J.; Gu, B.B.; Song, S.J.; Lin, H.W. Unusual Anti-allergic Diterpenoids from the Marine Sponge Hippospongia lachne. Sci. Rep. 2017, 7, 43138. [Google Scholar] [CrossRef] [Green Version]
- Kozuma, S.; Hirota-Takahata, Y.; Fukuda, D.; Kuraya, N.; Nakajima, M.; Ando, O. Identification and biological activity of ogipeptins, novel LPS inhibitors produced by marine bacterium. J. Antibiot. 2017, 70, 79–83. [Google Scholar] [CrossRef]
- Kwon, I.S.; Kwak, J.H.; Pyo, S.; Lee, H.W.; Kim, A.; Schmitz, F.J. Oscarellin, an Anthranilic Acid Derivative from a Philippine Sponge, Oscarella stillans, as an Inhibitor of Inflammatory Cytokines in Macrophages. J. Nat. Prod. 2017, 80, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.E.; Jung, I.; Lee, J.S.; Na, J.Y.; Kim, W.J.; Kim, Y.O.; Park, Y.D.; Lee, J.S. Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo. Mar. Drugs 2017, 15, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Zhao, J.; Proksch, P.; Zhao, Z.; Lin, W. Eremophilane-Type Sesquiterpenoids from an Acremonium sp. Fungus Isolated from Deep-Sea Sediments. J. Nat. Prod. 2016, 79, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Boonlarppradab, C.; Fenical, W. Actinoquinolines A and B, anti-inflammatory quinoline alkaloids from a marine-derived Streptomyces sp., strain CNP975. J. Antibiot. 2016, 69, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Malyarenko, T.V.; Kharchenko, S.D.; Kicha, A.A.; Ivanchina, N.V.; Dmitrenok, P.S.; Chingizova, E.A.; Pislyagin, E.A.; Evtushenko, E.V.; Antokhina, T.I.; Minh, C.V.; et al. Anthenosides L-U, Steroidal Glycosides with Unusual Structural Features from the Starfish Anthenea aspera. J. Nat. Prod. 2016, 79, 3047–3056. [Google Scholar] [CrossRef]
- Fang, W.; Lin, X.; Wang, J.; Liu, Y.; Tao, H.; Zhou, X. Asperpyrone-Type Bis-Naphtho-gamma-Pyrones with COX-2-Inhibitory Activities from Marine-Derived Fungus Aspergillus niger. Molecules 2016, 21, 941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.D.; Wen, Z.H.; Wu, Y.C.; Fang, L.S.; Chen, Y.H.; Chang, Y.C.; Sheu, J.H.; Sung, P.J. Briarenolides M-T, new briarane diterpenoids from a Formosan octocoral Briareum sp. Tetrahedron 2016, 72, 944–951. [Google Scholar] [CrossRef]
- Su, Y.D.; Sung, C.S.; Wen, Z.H.; Chen, Y.H.; Chang, Y.C.; Chen, J.J.; Fang, L.S.; Wu, Y.C.; Sheu, J.H.; Sung, P.J. New 9-Hydroxybriarane Diterpenoids from a Gorgonian Coral Briareum sp. (Briareidae). Int. J. Mol. Sci. 2016, 17, 79. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, M.; Tang, J.; Li, X. Eremophilane Sesquiterpenes from a Deep Marine-Derived Fungus, Aspergillus sp. SCSIOW2, Cultivated in the Presence of Epigenetic Modifying Agents. Molecules 2016, 21, 473. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.; Lee, H.; Ko, W.; Kim, D.; Kim, K.; Kwon, H.C. Chemical constituents isolated from Antarctic marine-derived Aspergillus sp. SF-5976 and their anti-inflammatory effects in LPS-stimulated RAW 264.7 and BV2 cells. Tetrahedron 2017, 73, 3905–3912. [Google Scholar] [CrossRef]
- Hsu, Y.M.; Chang, F.R.; Lo, I.W.; Lai, K.H.; El-Shazly, M.; Wu, T.Y.; Du, Y.C.; Hwang, T.L.; Cheng, Y.B.; Wu, Y.C. Zoanthamine-Type Alkaloids from the Zoanthid Zoanthus kuroshio Collected in Taiwan and Their Effects on Inflammation. J. Nat. Prod. 2016, 79, 2674–2680. [Google Scholar] [CrossRef] [PubMed]
- Tseng, W.R.; Huang, C.Y.; Tsai, Y.Y.; Lin, Y.S.; Hwang, T.L.; Su, J.H.; Sung, P.J.; Dai, C.F.; Sheu, J.H. New cytotoxic and anti-inflammatory steroids from the soft coral Klyxum flaccidum. Bioorg. Med. Chem. Lett. 2016, 26, 3253–3257. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.Y.; Huang, C.Y.; Tseng, W.R.; Chiang, P.L.; Hwang, T.L.; Su, J.H.; Sung, P.J.; Dai, C.F.; Sheu, J.H. Klyflaccisteroids K-M, bioactive steroidal derivatives from a soft coral Klyxum flaccidum. Bioorg. Med. Chem. Lett. 2017, 27, 1220–1224. [Google Scholar] [CrossRef]
- Ahmed, A.F.; Teng, W.T.; Huang, C.Y.; Dai, C.F.; Hwang, T.L.; Sheu, J.H. Anti-Inflammatory Lobane and Prenyleudesmane Diterpenoids from the Soft Coral Lobophytum varium. Mar. Drugs 2017, 15, 300. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Chang, C.W.; Tseng, Y.J.; Lee, J.; Sung, P.J.; Su, J.H.; Hwang, T.L.; Dai, C.F.; Wang, H.C.; Sheu, J.H. Bioactive Steroids from the Formosan Soft Coral Umbellulifera petasites. Mar. Drugs 2016, 14, 180. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.D.; Cheng, C.H.; Wen, Z.H.; Wu, Y.C.; Sung, P.J. New anti-inflammatory sterols from a gorgonian Pinnigorgia sp. Bioorg. Med. Chem. Lett. 2016, 26, 3060–3063. [Google Scholar] [CrossRef]
- Chang, Y.C.; Kuo, L.M.; Su, J.H.; Hwang, T.L.; Kuo, Y.H.; Lin, C.S.; Wu, Y.C.; Sheu, J.H.; Sung, P.J. Pinnigorgiols A-C, 9,11-secosterols with a rare ring arrangement from a gorgonian coral Pinnigorgia sp. Tetrahedron 2016, 72, 999–1004. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hwang, T.L.; Sheu, J.H.; Wu, Y.C.; Sung, P.J. New Anti-Inflammatory 9,11-Secosterols with a Rare Tricyclo[5,2,1,1]decane Ring from a Formosan Gorgonian Pinnigorgia sp. Mar. Drugs 2016, 14, 218. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Kuo, L.M.; Hwang, T.L.; Yeh, J.; Wen, Z.H.; Fang, L.S.; Wu, Y.C.; Lin, C.S.; Sheu, J.H.; Sung, P.J. Pinnisterols A-C, New 9,11-Secosterols from a Gorgonian Pinnigorgia sp. Mar. Drugs 2016, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Hwang, T.L.; Kuo, L.M.; Sung, P.J. Pinnisterols D-J, New 11-Acetoxy-9,11-secosterols with a 1,4-Quinone Moiety from Formosan Gorgonian Coral Pinnigorgia sp. (Gorgoniidae). Mar. Drugs 2017, 15, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vien, L.T.; Hanh, T.T.; Huong, P.T.; Dang, N.H.; Thanh, N.V.; Lyakhova, E.; Cuong, N.X.; Nam, N.H.; Kiem, P.V.; Kicha, A.; et al. Pyrrole Oligoglycosides from the Starfish Acanthaster planci Suppress Lipopolysaccharide-Induced Nitric Oxide Production in RAW264.7 Macrophages. Chem. Pharm. Bull. 2016, 64, 1654–1657. [Google Scholar] [CrossRef] [Green Version]
- Malyarenko, T.V.; Kicha, A.A.; Kalinovsky, A.I.; Ivanchina, N.V.; Popov, R.S.; Pislyagin, E.A.; Menchinskaya, E.S.; Padmakumar, K.P.; Stonik, V.A. Four New Steroidal Glycosides, Protolinckiosides A-D, from the Starfish Protoreaster lincki. Chem. Biodivers. 2016, 13, 998–1007. [Google Scholar] [CrossRef]
- Zhao, M.; Cheng, S.; Yuan, W.; Xi, Y.; Li, X.; Dong, J.; Huang, K.; Gustafson, K.R.; Yan, P. Cembranoids from a Chinese Collection of the Soft Coral Lobophytum crassum. Mar. Drugs 2016, 14, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.Y.; Tseng, Y.J.; Chokkalingam, U.; Hwang, T.L.; Hsu, C.H.; Dai, C.F.; Sung, P.J.; Sheu, J.H. Bioactive Isoprenoid-Derived Natural Products from a Dongsha Atoll Soft Coral Sinularia erecta. J. Nat. Prod. 2016, 79, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Ahmed, A.F.; Su, J.H.; Sung, P.J.; Hwang, T.L.; Chiang, P.L.; Dai, C.F.; Liaw, C.C.; Sheu, J.H. Bioactive new withanolides from the cultured soft coral Sinularia brassica. Bioorg. Med. Chem. Lett. 2017, 27, 3267–3271. [Google Scholar] [CrossRef]
- Huang, C.Y.; Su, J.H.; Liaw, C.C.; Sung, P.J.; Chiang, P.L.; Hwang, T.L.; Dai, C.F.; Sheu, J.H. Bioactive Steroids with Methyl Ester Group in the Side Chain from a Reef Soft Coral Sinularia brassica Cultured in a Tank. Mar. Drugs 2017, 15, 280. [Google Scholar] [CrossRef] [Green Version]
- Torres-Mendoza, D.; Gonzalez, Y.; Gomez-Reyes, J.F.; Guzman, H.M.; Lopez-Perez, J.L.; Gerwick, W.H.; Fernandez, P.L.; Gutierrez, M. Uprolides N, O and P from the Panamanian Octocoral Eunicea succinea. Molecules 2016, 21, 819. [Google Scholar] [CrossRef] [Green Version]
- Pislyagin, E.A.; Manzhulo, I.V.; Dmitrenok, P.S.; Aminin, D.L. Cucumarioside A2-2 causes changes in the morphology and proliferative activity in mouse spleen. Acta Histochem. 2016, 118, 387–392. [Google Scholar] [CrossRef]
- Pislyagin, E.A.; Manzhulo, I.V.; Gorpenchenko, T.Y.; Dmitrenok, P.S.; Avilov, S.A.; Silchenko, A.S.; Wang, Y.M.; Aminin, D.L. Cucumarioside A(2)-2 Causes Macrophage Activation in Mouse Spleen. Mar. Drugs 2017, 15, 341. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.A.; Alfonso, A.; Rodriguez, I.; Alonso, E.; Cifuentes, J.M.; Bermudez, R.; Rateb, M.E.; Jaspars, M.; Houssen, W.E.; Ebel, R.; et al. Spongionella Secondary Metabolites, Promising Modulators of Immune Response through CD147 Receptor Modulation. Front. Immunol. 2016, 7, 452. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.A.; Alfonso, A.; Leiros, M.; Alonso, E.; Rateb, M.E.; Jaspars, M.; Houssen, W.E.; Ebel, R.; Tabudravu, J.; Botana, L.M. Identification of Spongionella compounds as cyclosporine A mimics. Pharmacol. Res. 2016, 107, 407–414. [Google Scholar] [CrossRef]
- Becker, K.; Hartmann, A.; Ganzera, M.; Fuchs, D.; Gostner, J.M. Immunomodulatory Effects of the Mycosporine-Like Amino Acids Shinorine and Porphyra-334. Mar. Drugs 2016, 14, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, T.W.; Li, Y.R.; Huang, W.Y.; Su, J.H.; Chan, H.L.; Lin, S.H.; Liu, C.S.; Lin, S.C.; Lin, C.C.; Lin, C.H. Sinulariolide suppresses LPSinduced phenotypic and functional maturation of dendritic cells. Mol. Med. Rep. 2017, 16, 6992–7000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.Y.; Liu, Q.M.; Liu, B.; Xie, C.L.; Cao, M.J.; Yang, X.W.; Liu, G.M. Inhibitory Activities of Compounds from the Marine Actinomycete Williamsia sp. MCCC 1A11233 Variant on IgE-Mediated Mast Cells and Passive Cutaneous Anaphylaxis. J. Agric. Food Chem. 2017, 65, 10749–10756. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, J.; Lin, X.; Zhao, B.; Wei, X.; Li, G.; Kaliaperumal, K.; Liao, S.; Yang, B.; Zhou, X.; et al. Chrysamides A-C, Three Dimeric Nitrophenyl trans-Epoxyamides Produced by the Deep-Sea-Derived Fungus Penicillium chrysogenum SCSIO41001. Org. Lett. 2016, 18, 3650–3653. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Li, Y.; Ratnayake, R.; Luo, D.; Lo, J.; Reibenspies, J.H.; Xu, Z.; Clare-Salzler, M.J.; Ye, T.; Paul, V.J.; et al. Discovery, Total Synthesis and Key Structural Elements for the Immunosuppressive Activity of Cocosolide, a Symmetrical Glycosylated Macrolide Dimer from Marine Cyanobacteria. Chemistry 2016, 22, 8158–8166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einarsdottir, E.; Liu, H.B.; Freysdottir, J.; Gotfredsen, C.H.; Omarsdottir, S. Immunomodulatory N-acyl Dopamine Glycosides from the Icelandic Marine Sponge Myxilla incrustans Collected at a Hydrothermal Vent Site. Planta Med. 2016, 82, 903–909. [Google Scholar] [CrossRef] [Green Version]
- Kawase, O.; Ohno, O.; Suenaga, K.; Xuan, X. Immunological Adjuvant Activity of Pectinioside A, the Steroidal Saponin from the Starfish Patiria pectinifera. Nat. Prod. Commun. 2016, 11, 605–606. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Liu, W.; Liu, Y.; Huang, X.; She, Z. Polyketides with Immunosuppressive Activities from Mangrove Endophytic Fungus Penicillium sp. ZJ-SY(2). Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Huang, H.; Hou, L.; Ju, J.; Li, W. Discovery of Antimycin-Type Depsipeptides from a wbl Gene Mutant Strain of Deepsea-Derived Streptomyces somaliensis SCSIO ZH66 and Their Effects on Pro-inflammatory Cytokine Production. Front. Microbiol. 2017, 8, 678. [Google Scholar] [CrossRef]
- Moreels, L.; Peigneur, S.; Galan, D.T.; De, P.E.; Beress, L.; Waelkens, E.; Pardo, L.A.; Quinton, L.; Tytgat, J. APETx4, a Novel Sea Anemone Toxin and a Modulator of the Cancer-Relevant Potassium Channel KV10.1. Mar. Drugs 2017, 15, 287. [Google Scholar] [CrossRef]
- Goda, A.A.; Naguib, K.M.; Mohamed, M.M.; Amra, H.A.; Nada, S.A.; Abdel-Ghaffar, A.B.; Gissendanner, C.R.; El Sayed, K.A. Astaxanthin and Docosahexaenoic Acid Reverse the Toxicity of the Maxi-K (BK) Channel Antagonist Mycotoxin Penitrem A. Mar. Drugs 2016, 14, 208. [Google Scholar] [CrossRef] [Green Version]
- Mendez, A.G.; Juncal, A.B.; Silva, S.B.L.; Thomas, O.P.; Martin, V.V.; Alfonso, A.; Vieytes, M.R.; Vale, C.; Botana, L.M. The Marine Guanidine Alkaloid Crambescidin 816 Induces Calcium Influx and Cytotoxicity in Primary Cultures of Cortical Neurons through Glutamate Receptors. ACS Chem. Neurosci. 2017, 8, 1609–1617. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Tae, H.S.; Xu, S.; Shao, X.; Adams, D.J.; Wang, C. Identification of a Novel O-Conotoxin Reveals an Unusual and Potent Inhibitor of the Human alpha9alpha10 Nicotinic Acetylcholine Receptor. Mar. Drugs 2017, 15, 170. [Google Scholar] [CrossRef] [Green Version]
- Bernaldez, J.; Jimenez, S.; Gonzalez, L.J.; Ferro, J.N.; Soto, E.; Salceda, E.; Chavez, D.; Aguilar, M.B.; Licea-Navarro, A. A New Member of Gamma-Conotoxin Family Isolated from Conus princeps Displays a Novel Molecular Target. Toxins 2016, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Brust, A.; Croker, D.E.; Colless, B.; Ragnarsson, L.; Andersson, A.; Jain, K.; Garcia-Caraballo, S.; Castro, J.; Brierley, S.M.; Alewood, P.F.; et al. Conopeptide-Derived kappa-Opioid Agonists (Conorphins): Potent, Selective, and Metabolic Stable Dynorphin A Mimetics with Antinociceptive Properties. J. Med. Chem. 2016, 59, 2381–2395. [Google Scholar] [CrossRef]
- Feng, C.W.; Hung, H.C.; Huang, S.Y.; Chen, C.H.; Chen, Y.R.; Chen, C.Y.; Yang, S.N.; Wang, H.D.; Sung, P.J.; Sheu, J.H.; et al. Neuroprotective Effect of the Marine-Derived Compound 11-Dehydrosinulariolide through DJ-1-Related Pathway in In Vitro and In Vivo Models of Parkinson’s Disease. Mar. Drugs 2016, 14, 187. [Google Scholar] [CrossRef]
- Chen, C.H.; Chen, N.F.; Feng, C.W.; Cheng, S.Y.; Hung, H.C.; Tsui, K.H.; Hsu, C.H.; Sung, P.J.; Chen, W.F.; Wen, Z.H. A Coral-Derived Compound Improves Functional Recovery after Spinal Cord Injury through Its Antiapoptotic and Anti-Inflammatory Effects. Mar. Drugs 2016, 14, 160. [Google Scholar] [CrossRef] [Green Version]
- Botic, T.; Defant, A.; Zanini, P.; Zuzek, M.C.; Frangez, R.; Janussen, D.; Kersken, D.; Knez, Z.; Mancini, I.; Sepcic, K. Discorhabdin alkaloids from Antarctic Latrunculia spp. sponges as a new class of cholinesterase inhibitors. Eur. J. Med. Chem. 2017, 136, 294–304. [Google Scholar] [CrossRef]
- Jung, H.A.; Ali, M.Y.; Choi, R.J.; Jeong, H.O.; Chung, H.Y.; Choi, J.S. Kinetics and molecular docking studies of fucosterol and fucoxanthin, BACE1 inhibitors from brown algae Undaria pinnatifida and Ecklonia stolonifera. Food Chem. Toxicol. 2016, 89, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, a Marine Carotenoid, Reverses Scopolamine-Induced Cognitive Impairments in Mice and Inhibits Acetylcholinesterase in Vitro. Mar. Drugs 2016, 14, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, H.; Fan, Y.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin provides neuroprotection in models of traumatic brain injury via the Nrf2-ARE and Nrf2-autophagy pathways. Sci. Rep. 2017, 7, 46763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, F.; Zhao, J.; Ding, L.; Huang, C.; Naman, C.B.; He, S.; Wu, B.; Zhu, P.; Luo, Q.; Gerwick, W.H.; et al. 5-Hydroxycyclopenicillone, a New beta-Amyloid Fibrillization Inhibitor from a Sponge-Derived Fungus Trichoderma sp. HPQJ-34. Mar. Drugs 2017, 15, 260. [Google Scholar] [CrossRef] [Green Version]
- Flores, P.L.; Rodriguez, E.; Zapata, E.; Carbo, R.; Farias, J.M.; Martinez, M. Maitotoxin Is a Potential Selective Activator of the Endogenous Transient Receptor Potential Canonical Type 1 Channel in Xenopus laevis Oocytes. Mar. Drugs 2017, 15, 198. [Google Scholar] [CrossRef] [Green Version]
- Alonso, E.; Alvarino, R.; Leiros, M.; Tabudravu, J.N.; Feussner, K.; Dam, M.A.; Rateb, M.E.; Jaspars, M.; Botana, L.M. Evaluation of the Antioxidant Activity of the Marine Pyrroloiminoquinone Makaluvamines. Mar. Drugs 2016, 14, 197. [Google Scholar] [CrossRef] [Green Version]
- Abdelhameed, R.; Elgawish, M.; Mira, A.; Ibrahim, A.; Ahmed, S.; Shimizu, K.; Yamada, K. Anti-choline esterase activity of ceramides from the Red Sea marine sponge Mycale euplectellioides. RSC Adv. 2016, 6, 20422–20430. [Google Scholar] [CrossRef]
- Uchimasu, H.; Matsumura, M.; Tsuda, M.; Kumagai, K.; Akakabe, M.; Fujieda, M.J.; Sakai, R. Mellpaladines and dopargimine, novel neuroactive guanidine alkaloids from a Palauan Didemnidae tunicate. Tetrahedron 2016, 72, 7185–7193. [Google Scholar] [CrossRef]
- Logashina, Y.A.; Mosharova, I.V.; Korolkova, Y.V.; Shelukhina, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Kozlov, S.A.; Stensvag, K.; et al. Peptide from Sea Anemone Metridium senile Affects Transient Receptor Potential Ankyrin-repeat 1 (TRPA1) Function and Produces Analgesic Effect. J. Biol. Chem. 2017, 292, 2992–3004. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Kang, Y.J.; Shin, S.A.; Bak, D.H.; Lee, J.W.; Lee, K.B.; Yoo, Y.C.; Kim, D.K.; Lee, B.H.; Kim, D.W.; et al. Phlorofucofuroeckol Improves Glutamate-Induced Neurotoxicity through Modulation of Oxidative Stress-Mediated Mitochondrial Dysfunction in PC12 Cells. PLoS ONE 2016, 11, e0163433. [Google Scholar] [CrossRef]
- Lee, H.W.; Choi, H.; Nam, S.J.; Fenical, W.; Kim, H. Potent Inhibition of Monoamine Oxidase B by a Piloquinone from Marine-Derived Streptomyces sp. CNQ-027. J. Microbiol. Biotechnol. 2017, 27, 785–790. [Google Scholar] [CrossRef] [Green Version]
- Caplan, S.L.; Zheng, B.; Dawson-Scully, K.; White, C.A.; West, L.M. Pseudopterosin A: Protection of Synaptic Function and Potential as a Neuromodulatory Agent. Mar. Drugs 2016, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Perni, M.; Galvagnion, C.; Maltsev, A.; Meisl, G.; Muller, M.B.; Challa, P.K.; Kirkegaard, J.B.; Flagmeier, P.; Cohen, S.I.; Cascella, R.; et al. A natural product inhibits the initiation of alpha-synuclein aggregation and suppresses its toxicity. Proc. Natl. Acad. Sci. USA 2017, 114, E1009–E1017. [Google Scholar] [CrossRef] [Green Version]
- Moodie, L.W.; Zuzek, M.C.; Frangez, R.; Andersen, J.H.; Hansen, E.; Olsen, E.K.; Cergolj, M.; Sepcic, K.; Hansen, K.O.; Svenson, J. Synthetic analogs of stryphnusin isolated from the marine sponge Stryphnus fortis inhibit acetylcholinesterase with no effect on muscle function or neuromuscular transmission. Org. Biomol. Chem. 2016, 14, 11220–11229. [Google Scholar] [CrossRef] [Green Version]
- Pan, N.; Lu, L.Y.; Li, M.; Wang, G.H.; Sun, F.Y.; Sun, H.S.; Wen, X.J.; Cheng, J.D.; Chen, J.W.; Pang, J.Y.; et al. Xyloketal B alleviates cerebral infarction and neurologic deficits in a mouse stroke model by suppressing the ROS/TLR4/NF-kappaB inflammatory signaling pathway. Acta Pharmacol. Sin. 2017, 38, 1236–1247. [Google Scholar] [CrossRef]
- Dai, J.; Parrish, S.M.; Yoshida, W.Y.; Yip, M.L.R.; Turkson, J.; Kelly, M.; Williams, P. Bromotyrosine-derived metabolites from an Indonesian marine sponge in the family Aplysinellidae (Order Verongiida). Bioorg. Med. Chem. Lett. 2016, 26, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Naman, C.B.; Almaliti, J.; Armstrong, L.; Caro-Diaz, E.J.; Pierce, M.L.; Glukhov, E.; Fenner, A.; Spadafora, C.; Debonsi, H.M.; Dorrestein, P.C.; et al. Discovery and Synthesis of Caracolamide A, an Ion Channel Modulating Dichlorovinylidene Containing Phenethylamide from a Panamanian Marine Cyanobacterium cf. Symploca Species. J. Nat. Prod. 2017, 80, 2328–2334. [Google Scholar] [CrossRef]
- Campos-Lira, E.; Carrillo, E.; Aguilar, M.B.; Gajewiak, J.; Gomez-Lagunas, F.; Lopez-Vera, E. Conorfamide-Sr3, a structurally novel specific inhibitor of the Shaker K(+) channel. Toxicon 2017, 138, 53–58. [Google Scholar] [CrossRef]
- Han, P.; Cao, Y.; Liu, S.; Dai, X.; Yao, G.; Fan, C.; Wu, W.; Chen, J. Contryphan-Bt: A pyroglutamic acid containing conopeptide isolated from the venom of Conus betulinus. Toxicon 2017, 135, 17–23. [Google Scholar] [CrossRef]
- Long, Y.; Cui, H.; Liu, X.; Xiao, Z.; WEN, S.; She, Z.; Huang, X. Acetylcholinesterase Inhibitory Meroterpenoid from a Mangrove Endophytic Fungus Aspergillus sp. 16-5c. Molecules 2017, 22, 727. [Google Scholar] [CrossRef] [Green Version]
- Zidar, N.; Zula, A.; Tomasic, T.; Rogers, M.; Kirby, R.W.; Tytgat, J.; Peigneur, S.; Kikelj, D.; Ilas, J.; Masic, L.P. Clathrodin, hymenidin and oroidin, and their synthetic analogues as inhibitors of the voltage-gated potassium channels. Eur. J. Med. Chem. 2017, 139, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Boccitto, M.; Lee, N.; Sakamoto, S.; Spruce, L.A.; Handa, H.; Clardy, J.; Seeholzer, S.H.; Kalb, R.G. The Neuroprotective Marine Compound Psammaplysene A Binds the RNA-Binding Protein HNRNPK. Mar. Drugs 2017, 15, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, B.; Wang, Z.; Huang, X.; Liu, Y.; Chen, W.; She, Z. Bioactive alpha-pyrone meroterpenoids from mangrove endophytic fungus Penicillium sp. Nat. Prod. Res. 2016, 30, 2805–2812. [Google Scholar] [CrossRef] [PubMed]
- Omaga, C.A.; Carpio, L.D.; Imperial, J.S.; Daly, N.L.; Gajewiak, J.; Flores, M.S.; Espino, S.S.; Christensen, S.; Filchakova, O.M.; Lopez-Vera, E.; et al. Structure and Biological Activity of a Turripeptide from Unedogemmula bisaya Venom. Biochemistry 2017, 56, 6051–6060. [Google Scholar] [CrossRef] [PubMed]
- Yeon, J.T.; Kim, H.; Kim, K.J.; Lee, J.; Won, D.H.; Nam, S.J.; Kim, S.H.; Kang, H.; Son, Y.J. Acredinone C and the Effect of Acredinones on Osteoclastogenic and Osteoblastogenic Activity. J. Nat. Prod. 2016, 79, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bae, M.; Kim, B.; Park, Y.K.; Koo, S.I.; Lee, J.Y. Astaxanthin prevents and reverses the activation of mouse primary hepatic stellate cells. J. Nutr. Biochem. 2016, 29, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Abdjul, D.B.; Kanno, S.I.; Yamazaki, H.; Ukai, K.; Namikoshi, M. A dimeric urea of the bisabolene sesquiterpene from the Okinawan marine sponge Axinyssa sp. inhibits protein tyrosine phosphatase 1B activity in Huh-7 human hepatoma cells. Bioorg. Med. Chem. Lett. 2016, 26, 315–317. [Google Scholar] [CrossRef]
- Lai, K.H.; Liu, Y.C.; Su, J.H.; El-Shazly, M.; Wu, C.F.; Du, Y.C.; Hsu, Y.M.; Yang, J.C.; Weng, M.K.; Chou, C.H.; et al. Antileukemic Scalarane Sesterterpenoids and Meroditerpenoid from Carteriospongia Phyllospongia sp., Induce Apoptosis via Dual Inhibitory Effects on Topoisomerase II and Hsp90. Sci. Rep. 2016, 6, 36170. [Google Scholar] [CrossRef]
- Uchida, R.; Nakajyo, K.; Kobayashi, K.; Ohshiro, T.; Terahara, T.; Imada, C.; Tomoda, H. 7-Chlorofolipastatin, an inhibitor of sterol O-acyltransferase, produced by marine-derived Aspergillus ungui NKH-007. J. Antibiot. 2016, 69, 647–651. [Google Scholar] [CrossRef]
- Esposito, G.; Bourguet-Kondracki, M.L.; Mai, L.H.; Longeon, A.; Teta, R.; Meijer, L.; Van, S.R.; Mangoni, A.; Costantino, V. Chloromethylhalicyclamine B, a Marine-Derived Protein Kinase CK1delta/epsilon Inhibitor. J. Nat. Prod. 2016, 79, 2953–2960. [Google Scholar] [CrossRef]
- Serrill, J.D.; Wan, X.; Hau, A.M.; Jang, H.S.; Coleman, D.J.; Indra, A.K.; Alani, A.W.; McPhail, K.L.; Ishmael, J.E. Coibamide A, a natural lariat depsipeptide, inhibits VEGFA/VEGFR2 expression and suppresses tumor growth in glioblastoma xenografts. Invest. New Drugs 2016, 34, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Issac, M.; Aknin, M.; Gauvin-Bialecki, A.; De Voogd, N.; Ledoux, A.; Frederich, M.; Kashman, Y.; Carmeli, S. Cyclotheonellazoles A-C, Potent Protease Inhibitors from the Marine Sponge Theonella aff. swinhoei. J. Nat. Prod. 2017, 80, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Vaden, R.M.; Oswald, N.W.; Potts, M.B.; MacMillan, J.B.; White, M.A. FUSION-Guided Hypothesis Development Leads to the Identification of N(6),N(6)-Dimethyladenosine, a Marine-Derived AKT Pathway Inhibitor. Mar. Drugs 2017, 15, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.R.; Tai, B.Y.; Cheng, P.Y.; Chen, P.N.; Sung, P.J.; Wen, Z.H.; Hsu, C.H. Excavatolide B Modulates the Electrophysiological Characteristics and Calcium Homeostasis of Atrial Myocytes. Mar. Drugs 2017, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.Y.; Park, W.S.; Lee, D.S.; Choi, G.; Yim, M.J.; Lee, J.M.; Jung, W.K.; Park, S.G.; Seo, S.K.; Park, S.J.; et al. Fucoxanthin inhibits profibrotic protein expression in vitro and attenuates bleomycin-induced lung fibrosis in vivo. Eur. J. Pharmacol. 2017, 811, 199–207. [Google Scholar] [CrossRef]
- Lee, H.Y.; Jang, E.J.; Bae, S.Y.; Jeon, J.E.; Park, H.J.; Shin, J.; Lee, S.K. Anti-Melanogenic Activity of Gagunin D, a Highly Oxygenated Diterpenoid from the Marine Sponge Phorbas sp., via Modulating Tyrosinase Expression and Degradation. Mar. Drugs 2016, 14, 212. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Leutou, A.S.; Jeong, H.; Kim, D.; Seong, C.N.; Nam, S.J.; Lim, K.M. Anti-Pigmentary Effect of (-)-4-Hydroxysattabacin from the Marine-Derived Bacterium Bacillus sp. Mar. Drugs 2017, 15, 138. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Kawazoe, Y.; Iwasaki, A.; Ohno, O.; Suenaga, K.; Uemura, D. Anti-obesity activities of the yoshinone A and the related marine gamma-pyrone compounds. J. Antibiot. 2016, 69, 348–351. [Google Scholar] [CrossRef]
- Carlile, G.W.; Robert, R.; Matthes, E.; Yang, Q.; Solari, R.; Hatley, R.; Edge, C.M.; Hanrahan, J.W.; Andersen, R.; Thomas, D.Y.; et al. Latonduine Analogs Restore F508del-Cystic Fibrosis Transmembrane Conductance Regulator Trafficking through the Modulation of Poly-ADP Ribose Polymerase 3 and Poly-ADP Ribose Polymerase 16 Activity. Mol. Pharmacol. 2016, 90, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Ebrahim, H.Y.; El Sayed, K.A. Discovery of Novel Antiangiogenic Marine Natural Product Scaffolds. Mar. Drugs 2016, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.E.; Izard, F.; Arnould, S.; Dalisay, D.S.; Tantapakul, C.; Maneerat, W.; Matainaho, T.; Julien, E.; Andersen, R.J. Structures of Nahuoic Acids B-E Produced in Culture by a Streptomyces sp. Isolated from a Marine Sediment and Evidence for the Inhibition of the Histone Methyl Transferase SETD8 in Human Cancer Cells by Nahuoic Acid A. J. Org. Chem. 2016, 81, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.S.; Kim, G.J.; Choi, H.G.; Kim, M.C.; Hahn, D.; Nam, J.W.; Nam, S.J.; Kwon, H.C.; Chin, J.; Cho, S.J.; et al. Identification of Antiangiogenic Potential and Cellular Mechanisms of Napyradiomycin A1 Isolated from the Marine-Derived Streptomyces sp. YP127. J. Nat. Prod. 2017, 80, 2269–2275. [Google Scholar] [CrossRef] [PubMed]
- Loaec, N.; Attanasio, E.; Villiers, B.; Durieu, E.; Tahtouh, T.; Cam, M.; Davis, R.A.; Alencar, A.; Roue, M.; Bourguet-Kondracki, M.L.; et al. Marine-Derived 2-Aminoimidazolone Alkaloids. Leucettamine B-Related Polyandrocarpamines Inhibit Mammalian and Protozoan DYRK & CLK Kinases. Mar. Drugs 2017, 15, 316. [Google Scholar]
- Starkus, J.G.; Poerzgen, P.; Layugan, K.; Kawabata, K.G.; Goto, J.I.; Suzuki, S.; Myers, G.; Kelly, M.; Penner, R.; Fleig, A.; et al. Scalaradial Is a Potent Inhibitor of Transient Receptor Potential Melastatin 2 (TRPM2) Ion Channels. J. Nat. Prod. 2017, 80, 2741–2750. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Ko, S.K.; Kim, H.M.; Kim, G.H.; Son, S.; Kim, G.S.; Hwang, G.J.; Jeon, E.S.; Shin, K.S.; Ryoo, I.J.; et al. Stachybotrysin, an Osteoclast Differentiation Inhibitor from the Marine-Derived Fungus Stachybotrys sp. KCB13F013. J. Nat. Prod. 2016, 79, 2703–2708. [Google Scholar] [CrossRef]
- Mason, J.W.; Schmid, C.L.; Bohn, L.M.; Roush, W.R. Stolonidiol: Synthesis, Target Identification, and Mechanism for Choline Acetyltransferase Activation. J. Am. Chem. Soc. 2017, 139, 5865–5869. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Ratnayake, R.; Paul, V.J.; Luesch, H. Tasiamide F, a potent inhibitor of cathepsins D and E from a marine cyanobacterium. Bioorg. Med. Chem. 2016, 24, 3276–3282. [Google Scholar] [CrossRef] [Green Version]
- Espiritu, R.A.; Cornelio, K.; Kinoshita, M.; Matsumori, N.; Murata, M.; Nishimura, S.; Kakeya, H.; Yoshida, M.; Matsunaga, S. Marine sponge cyclic peptide theonellamide A disrupts lipid bilayer integrity without forming distinct membrane pores. Biochim. Biophys. Acta 2016, 1858, 1373–1379. [Google Scholar] [CrossRef]
- Garcia-Caballero, M.; Blacher, S.; Paupert, J.; Quesada, A.R.; Medina, M.A.; Noel, A. Novel application assigned to toluquinol: Inhibition of lymphangiogenesis by interfering with VEGF-C/VEGFR-3 signalling pathway. Br. J. Pharmacol. 2016, 173, 1966–1987. [Google Scholar] [CrossRef]
- Yang, Z.; Zhu, M.L.; Li, D.H.; Zeng, R.; Han, B.N. N-Me-trichodermamide B isolated from Penicillium janthinellum, with antioxidant properties through Nrf2-mediated signaling pathway. Bioorg. Med. Chem. 2017, 25, 6614–6622. [Google Scholar] [CrossRef]
- Chen, R.; Cheng, Z.; Huang, J.; Liu, D.; Wu, C.; Guo, P.; Lin, W. Versicotides D–F, new cyclopeptides with lipid-lowering activities. RSC Adv. 2017, 7, 49235–49243. [Google Scholar] [CrossRef] [Green Version]
- Abdjul, D.B.; Yamazaki, H.; Kanno, S.I.; Wewengkang, D.S.; Rotinsulu, H.; Sumilat, D.A.; Ukai, K.; Kapojos, M.M.; Namikoshi, M. Furanoterpenes, new types of protein tyrosine phosphatase 1B inhibitors, from two Indonesian marine sponges, Ircinia and Spongia spp. Bioorg. Med. Chem. Lett. 2017, 27, 1159–1161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Meng, T.; Zuo, L.; Bei, Y.; Zhang, Q.; Su, Z.; Huang, Y.; Pang, J.; Xiang, Q.; Yang, H. Xyloketal B Attenuates Fatty Acid-Induced Lipid Accumulation via the SREBP-1c Pathway in NAFLD Models. Mar. Drugs 2017, 15, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamad, H.; Muhammad, R.; Andriani, Y.; Bakar, K.; Ismail, N.; Saidin, J.; Latip, J.; Musa, N.; Parenrengi, A. Potential secondary metabolites from marine sponge Aaptos aaptos for atherosclerosis and vibriosis treatments. Nat. Prod. Commun. 2017, 12, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Liu, Z.; Chen, S.; Huang, X.; Cui, H.; Long, Y.; Lu, Y.; She, Z. Antioxidative Polyketones from the Mangrove-Derived Fungus Ascomycota sp. SK2YWS-L. Sci. Rep. 2016, 6, 36609. [Google Scholar] [CrossRef] [Green Version]
- Leutou, A.S.; Yun, K.; Son, B.W. Induced production of 6,9-dibromoflavasperone, a new radical scavenging naphthopyranone in the marine-mudflat-derived fungus Aspergillus niger. Arch. Pharm. Res. 2016, 39, 806–810. [Google Scholar] [CrossRef]
- Sobolevskaya, M.P.; Zhuravleva, O.I.; Leshchenko, E.V.; Zakharenko, A.M.; Denisenko, V.A.; Kirichuk, N.N.; Popov, R.S.; Berdyshev, D.V.; Pislyagin, E.A.; Pivkin, M.V.; et al. New metabolites from the alga-derived fungi Penicillium thomii Maire and Penicillium lividum Westling. Phytochem. Lett. 2016, 15, 7–12. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Schmaljohann, R.; Imhoff, J.F. Biscogniauxone, a New Isopyrrolonaphthoquinone Compound from the Fungus Biscogniauxia mediterranea Isolated from Deep-Sea Sediments. Mar. Drugs 2016, 14, 204. [Google Scholar] [CrossRef] [Green Version]
- Oleinikova, G.K.; Denisenko, V.A.; Berdyshev, D.V.; Pushilin, M.A.; Kirichuk, N.N.; Menzorova, N.I.; Kuzmich, A.S.; Yurchenko, E.A.; Zhuravleva, O.I. Two new sesterterpenoids, terretonins H and I, from the marine-derived fungus Aspergillus ustus. Phytochem. Lett. 2016, 17, 135–139. [Google Scholar] [CrossRef]
- El-Desoky, A.H.; Kato, H.; Kagiyama, I.; Hitora, Y.; Losung, F.; Mangindaan, R.E.; de Voogd, N.J.; Tsukamoto, S. Ceylonins A-F, Spongian Diterpene Derivatives That Inhibit RANKL-Induced Formation of Multinuclear Osteoclasts, from the Marine Sponge Spongia ceylonensis. J. Nat. Prod. 2017, 80, 90–95. [Google Scholar] [CrossRef]
- Li, Y.; Liu, D.; Cheng, Z.; Proksch, P.; Lin, W. Cytotoxic trichothecene-type sesquiterpenes from the sponge-derived fungus Stachybotrys chartarum with tyrosine kinase inhibition. RSC Adv. 2017, 7, 7259–7267. [Google Scholar] [CrossRef] [Green Version]
- Noinart, J.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Urbatzka, R.; Freitas, S.; Lee, M.; Silva, A.M.S.; Pinto, M.M.M.; et al. A New Ergosterol Analog, a New Bis-Anthraquinone and Anti-Obesity Activity of Anthraquinones from the Marine Sponge-Associated Fungus Talaromyces stipitatus KUFA 0207. Mar. Drugs 2017, 15, 139. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xia, Z.; Tang, J.; Wu, J.; Tong, J.; Li, M.; Ju, J.; Chen, H.; Wang, L. Identification and Biological Evaluation of Secondary Metabolites from Marine Derived Fungi-Aspergillus sp. SCSIOW3, Cultivated in the Presence of Epigenetic Modifying Agents. Molecules 2017, 22, 1302. [Google Scholar] [CrossRef] [Green Version]
- Yun, K.; Khong, T.T.; Leutou, A.S.; Kim, G.D.; Hong, J.; Lee, C.H.; Son, B.W. Cristazine, a New Cytotoxic Dioxopiperazine Alkaloid from the Mudflat-Sediment-Derived Fungus Chaetomium cristatum. Chem. Pharm. Bull. 2016, 64, 59–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, H.; Cheng, Z.; Huang, W.; Wu, Q.; Li, X.; Cui, J.; Proksch, P.; Lin, W. Diasteltoxins A-C, Asteltoxin-Based Dimers from a Mutant of the Sponge-Associated Emericella variecolor Fungus. Org. Lett. 2016, 18, 4678–4681. [Google Scholar] [CrossRef] [PubMed]
- Goey, A.K.; Chau, C.H.; Sissung, T.M.; Cook, K.M.; Venzon, D.J.; Castro, A.; Ransom, T.R.; Henrich, C.J.; McKee, T.C.; McMahon, J.B.; et al. Screening and Biological Effects of Marine Pyrroloiminoquinone Alkaloids: Potential Inhibitors of the HIF-1alpha/p300 Interaction. J. Nat. Prod. 2016, 79, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Shin, D.; Kamiya, K.; Ishida, R.; Setiawan, A.; Kotoku, N.; Kobayashi, M. Marine spongean polybrominated diphenyl ethers, selective growth inhibitors against the cancer cells adapted to glucose starvation, inhibits mitochondrial complex II. J. Nat. Med. 2017, 71, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.H.; Shi, G.H.; Xu, T.T.; Chen, G.D.; Gu, B.B.; Wang, Z.; Peng, S.; Wang, S.P.; Li, J.; Han, B.N.; et al. Dysiherbols A-C and Dysideanone E, Cytotoxic and NF-kappaB Inhibitory Tetracyclic Meroterpenes from a Dysidea sp. Marine Sponge. J. Nat. Prod. 2016, 79, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Vasileva, E.A.; Mishchenko, N.P.; Zadorozhny, P.A.; Fedoreyev, S.A. New Aminonaphthoquinone from the Sea Urchins Strongylocentrotus pallidus and Mesocentrotus nudus. Nat. Prod. Commun. 2016, 11, 821–824. [Google Scholar] [CrossRef] [Green Version]
- Almeida, C.; El, M.F.; Kehraus, S.; Schnakenburg, G.; Konig, G.M. Endolides A and B, Vasopressin and Serotonin-Receptor Interacting N-Methylated Peptides from the Sponge-Derived Fungus Stachylidium sp. Org. Lett. 2016, 18, 528–531. [Google Scholar] [CrossRef]
- Meng, L.H.; Mandi, A.; Li, X.M.; Liu, Y.; Kurtan, T.; Wang, B.G. Isolation, Stereochemical Study, and Antioxidant Activity of Benzofuranone Derivatives from a Mangrove-derived Fungus Eurotium rubrum MA-150. Chirality 2016, 28, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Manda, S.; Sharma, S.; Wani, A.; Joshi, P.; Kumar, V.; Guru, S.K.; Bharate, S.S.; Bhushan, S.; Vishwakarma, R.A.; Kumar, A.; et al. Discovery of a marine-derived bis-indole alkaloid fascaplysin, as a new class of potent P-glycoprotein inducer and establishment of its structure-activity relationship. Eur. J. Med. Chem. 2016, 107, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Chen, W.T.; Kurtan, T.; Mandi, A.; Ding, J.; Li, J.; Li, X.W.; Guo, Y.W. Bioactive isoquinolinequinone alkaloids from the South China Sea nudibranch Jorunna funebris and its sponge-prey Xestospongia sp. Future Med. Chem. 2016, 8, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.; Kim, H.; Yang, I.; Chin, J.; Hwang, H.; Won, D.H.; Lee, B.; Nam, S.J.; Ekins, M.; Choi, H.; et al. The Halicylindramides, Farnesoid X Receptor Antagonizing Depsipeptides from a Petrosia sp. Marine Sponge Collected in Korea. J. Nat. Prod. 2016, 79, 499–506. [Google Scholar] [CrossRef]
- Abdjul, D.B.; Yamazaki, H.; Takahashi, O.; Kirikoshi, R.; Ukai, K.; Namikoshi, M. Isopetrosynol, a New Protein Tyrosine Phosphatase 1B Inhibitor, from the Marine Sponge Halichondria cf. panicea Collected at Iriomote Island. Chem. Pharm. Bull. 2016, 64, 733–736. [Google Scholar] [CrossRef]
- Cui, J.; Morita, M.; Ohno, O.; Kimura, T.; Teruya, T.; Watanabe, T.; Suenaga, K.; Shibasaki, M. Leptolyngbyolides, Cytotoxic Macrolides from the Marine Cyanobacterium Leptolyngbya sp.: Isolation, Biological Activity, and Catalytic Asymmetric Total Synthesis. Chemistry 2017, 23, 8500–8509. [Google Scholar] [CrossRef]
- Nazari, M.; Serrill, J.D.; Wan, X.; Nguyen, M.H.; Anklin, C.; Gallegos, D.A.; Smith, A.B., III; Ishmael, J.E.; McPhail, K.L. New Mandelalides Expand a Macrolide Series of Mitochondrial Inhibitors. J. Med. Chem. 2017, 60, 7850–7862. [Google Scholar] [CrossRef]
- Wahab, H.A.; Pham, N.B.; Muhammad, T.S.; Hooper, J.N.; Quinn, R.J. Merosesquiterpene Congeners from the Australian Sponge Hyrtios digitatus as Potential Drug Leads for Atherosclerosis Disease. Mar. Drugs 2016, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Korolkova, Y.; Makarieva, T.; Tabakmakher, K.; Shubina, L.; Kudryashova, E.; Andreev, Y.; Mosharova, I.; Lee, H.S.; Lee, Y.J.; Kozlov, S. Marine Cyclic Guanidine Alkaloids Monanchomycalin B and Urupocidin A Act as Inhibitors of TRPV1, TRPV2 and TRPV3, but not TRPA1 Receptors. Mar. Drugs 2017, 15, 87. [Google Scholar] [CrossRef] [Green Version]
- Chianese, G.; Yu, H.B.; Yang, F.; Sirignano, C.; Luciano, P.; Han, B.N.; Khan, S.; Lin, H.W.; Taglialatela-Scafati, O. PPAR Modulating Polyketides from a Chinese Plakortis simplex and Clues on the Origin of Their Chemodiversity. J. Org. Chem. 2016, 81, 5135–5143. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.Q.; Liu, H.L.; Chen, S.H.; Mollo, E.; Gavagnin, M.; Li, J.; Li, X.W.; Guo, Y.W. 5-Alkylpyrrole-2-carboxaldehyde derivatives from the Chinese sponge Mycale lissochela and their PTP1B inhibitory activities. Chinese Chem. Lett. 2017, 28, 1190–1193. [Google Scholar] [CrossRef]
- Wang, J.; Mu, F.R.; Jiao, W.H.; Huang, J.; Hong, L.L.; Yang, F.; Xu, Y.; Wang, S.P.; Sun, F.; Lin, H.W. Meroterpenoids with Protein Tyrosine Phosphatase 1B Inhibitory Activity from a Hyrtios sp. Marine Sponge. J. Nat. Prod. 2017, 80, 2509–2514. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, K.; Takada, K.; Okada, S.; Matsunaga, S. Nazumazoles D-F, Cyclic Pentapeptides That Inhibit Chymotrypsin, from the Marine Sponge Theonella swinhoei. J. Nat. Prod. 2016, 79, 1694–1697. [Google Scholar] [CrossRef] [PubMed]
- Wiese, J.; Imhoff, J.F.; Gulder, T.A.; Labes, A.; Schmaljohann, R. Marine Fungi as Producers of Benzocoumarins, a New Class of Inhibitors of Glycogen-Synthase-Kinase 3beta. Mar. Drugs 2016, 14, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Zhang, D.; Zhou, B.; Ma, Z. Inhibitors of BRD4 Protein from a Marine-Derived Fungus Alternaria sp. NH-F6. Mar. Drugs 2017, 15, 76. [Google Scholar] [CrossRef] [Green Version]
- Tanokashira, N.; Kukita, S.K.; Kato, H.; Nehira, T.; Angkouw, E.D.; Mangindaan, R.E.; Devoogd, N.J.; Tsukamoto, S. Petroquinones: Trimeric and dimeric xestoquinone derivatives isolated from the marine sponge Petrosia alfiani. Tetrahedron 2016, 72, 5530–5540. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, D.; Proksch, P.; Yu, S.; Lin, W. Isocoumarin Derivatives from the Sponge-Associated Fungus Peyronellaea glomerata with Antioxidant Activities. Chem. Biodivers. 2016, 13, 1186–1193. [Google Scholar] [CrossRef]
- Lee, M.S.; Wang, S.W.; Wang, G.J.; Pang, K.L.; Lee, C.K.; Kuo, Y.H.; Cha, H.J.; Lin, R.K.; Lee, T.H. Angiogenesis Inhibitors and Anti-Inflammatory Agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef]
- Imperatore, C.; Luciano, P.; Aiello, A.; Vitalone, R.; Irace, C.; Santamaria, R.; Li, J.; Guo, Y.W.; Menna, M. Structure and Configuration of Phosphoeleganin, a Protein Tyrosine Phosphatase 1B Inhibitor from the Mediterranean Ascidian Sidnyum elegans. J. Nat. Prod. 2016, 79, 1144–1148. [Google Scholar] [CrossRef]
- Georgantea, P.; Ioannou, E.; Evain-Bana, E.; Bagrel, D.; Martinet, N.; Vagias, C.; Roussis, V. Sesquiterpenes with inhibitory activity against CDC25 phosphatases from the soft coral Pseudopterogorgia rigida. Tetrahedron 2016, 72, 3262–3269. [Google Scholar] [CrossRef]
- El-Desoky, A.H.; Kato, H.; Tsukamoto, S. Ceylonins G-I: Spongian diterpenes from the marine sponge Spongia ceylonensis. J. Nat. Med. 2017, 71, 765–769. [Google Scholar] [CrossRef]
- Ortega, M.J.; Pantoja, J.J.; de Los, R.C.; Zubia, E. 5-Alkylresorcinol Derivatives from the Bryozoan Schizomavella mamillata: Isolation, Synthesis, and Antioxidant Activity. Mar. Drugs 2017, 15, 344. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Fu, Q.; Wu, W.; Cai, M.; Zhou, X.; Zhang, Y. Producing Novel Fibrinolytic Isoindolinone Derivatives in Marine Fungus Stachybotrys longispora FG216 by the Rational Supply of Amino Compounds According to Its Biosynthesis Pathway. Mar. Drugs 2017, 15, 214. [Google Scholar] [CrossRef]
- Li, J.; Gu, B.B.; Sun, F.; Xu, J.R.; Jiao, W.H.; Yu, H.B.; Han, B.N.; Yang, F.; Zhang, X.C.; Lin, H.W. Sesquiterpene Quinones/Hydroquinones from the Marine Sponge Spongia pertusa Esper. J. Nat. Prod. 2017, 80, 1436–1445. [Google Scholar] [CrossRef]
- Afifi, A.H.; Kagiyama, I.; El-Desoky, A.H.; Kato, H.; Mangindaan, R.E.P.; de Voogd, N.J.; Ammar, N.M.; Hifnawy, M.S.; Tsukamoto, S. Sulawesins A-C, Furanosesterterpene Tetronic Acids That Inhibit USP7, from a Psammocinia sp. Marine Sponge. J. Nat. Prod. 2017, 80, 2045–2050. [Google Scholar] [CrossRef]
- Kanamori, Y.; Iwasaki, A.; Sumimoto, S.; Suenaga, K. Urumamide, a novel chymotrypsin inhibitor with a ß-amino acid from a marine cyanobacterium Okeania sp. Tetrahedrom Lett. 2016, 57, 4213–4216. [Google Scholar] [CrossRef]
- Cheng, Z.; Lou, L.; Liu, D.; Li, X.; Proksch, P.; Yin, S.; Lin, W. Versiquinazolines A-K, Fumiquinazoline-Type Alkaloids from the Gorgonian-Derived Fungus Aspergillus versicolor LZD-14-1. J. Nat. Prod. 2016, 79, 2941–2952. [Google Scholar] [CrossRef]
- He, W.F.; Xue, D.Q.; Yao, L.G.; Li, J.; Liu, H.L.; Guo, Y.W. A new bioactive steroidal ketone from the South China Sea sponge Xestospongia testudinaria. J. Asian Nat. Prod. Res 2016, 18, 195–199. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235–294. [Google Scholar] [CrossRef] [Green Version]
- Berlinck, R.G.; Romminger, S. The chemistry and biology of guanidine natural products. Nat. Prod. Rep. 2016, 33, 456–490. [Google Scholar] [CrossRef]
- Shi, Y.; Moazami, Y.; Pierce, J.G. Structure, synthesis and biological properties of the pentacyclic guanidinium alkaloids. Bioorg. Med. Chem. 2017, 25, 2817–2824. [Google Scholar] [CrossRef]
- Pereira, R.B.; Andrade, P.B.; Valentao, P. Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus. Mar. Drugs 2016, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Chai, Q.Y.; Yang, Z.; Lin, H.W.; Han, B.N. Alkynyl-Containing Peptides of Marine Origin: A Review. Mar. Drugs 2016, 14, 216. [Google Scholar] [CrossRef] [Green Version]
- Mazard, S.; Penesyan, A.; Ostrowski, M.; Paulsen, I.T.; Egan, S. Tiny Microbes with a Big Impact: The Role of Cyanobacteria and Their Metabolites in Shaping Our Future. Mar. Drugs 2016, 14, 97. [Google Scholar] [CrossRef]
- Davila-Cespedes, A.; Hufendiek, P.; Crusemann, M.; Schaberle, T.F.; Konig, G.M. Marine-derived myxobacteria of the suborder Nannocystineae: An underexplored source of structurally intriguing and biologically active metabolites. Beilstein. J. Org. Chem 2016, 12, 969–984. [Google Scholar] [CrossRef]
- Offret, C.; Desriac, F.; Le, C.P.; Mounier, J.; Jegou, C.; Fleury, Y. Spotlight on Antimicrobial Metabolites from the Marine Bacteria Pseudoalteromonas: Chemodiversity and Ecological Significance. Mar. Drugs 2016, 14, 129. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.G.; Liu, Q.; Zhu, G.L.; Liu, H.S.; Zhu, W.M. Marine natural products sourced from marine-derived Penicillium fungi. J. Asian Nat. Prod. Res. 2016, 18, 92–115. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.; Wang, B.G. Secondary Metabolites from the Marine Algal-Derived Endophytic Fungi: Chemical Diversity and Biological Activity. Planta Med. 2016, 82, 832–842. [Google Scholar] [CrossRef] [Green Version]
- Abdul, Q.A.; Choi, R.J.; Jung, H.A.; Choi, J.S. Health benefit of fucosterol from marine algae: A review. J. Sci. Food Agric. 2016, 96, 1856–1866. [Google Scholar] [CrossRef]
- Avila, C. Ecological and Pharmacological Activities of Antarctic Marine Natural Products. Planta Med. 2016, 82, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Franco, C.M. Acetylated Triterpene Glycosides and Their Biological Activity from Holothuroidea Reported in the Past Six Decades. Mar. Drugs 2016, 14, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.C.; Sheu, J.H.; Wu, Y.C.; Sung, P.J. Terpenoids from Octocorals of the Genus Pachyclavularia. Mar. Drugs 2017, 15, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shady, N.H.; El-Hossary, E.M.; Fouad, M.A.; Gulder, T.A.M.; Kamel, M.S.; Abdelmohsen, U.R. Bioactive Natural Products of Marine Sponges from the Genus Hyrtios. Molecules 2017, 22, 781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, M.; Wang, H.; Crews, P. Secondary Metabolites from the Marine Sponge Genus Phyllospongia. Mar. Drugs 2017, 15, 12. [Google Scholar] [CrossRef] [Green Version]
- Amoutzias, G.D.; Chaliotis, A.; Mossialos, D. Discovery Strategies of Bioactive Compounds Synthesized by Nonribosomal Peptide Synthetases and Type-I Polyketide Synthases Derived from Marine Microbiomes. Mar. Drugs 2016, 14, 80. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J. Developing natural product drugs: Supply problems and how they have been overcome. Pharmacol. Ther. 2016, 162, 1–9. [Google Scholar] [CrossRef]
- Daletos, G.; Ebrahim, W.; Ancheeva, E.; El-Neketi, M.; Song, W.; Lin, W.; Proksch, P. Natural Products from Deep-Sea-Derived Fungi A New Source of Novel Bioactive Compounds? Curr. Med. Chem. 2018, 25, 186–207. [Google Scholar] [CrossRef]
- Mariottini, G.L.; Grice, I.D. Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy? Mar. Drugs 2016, 14, 48. [Google Scholar] [CrossRef] [Green Version]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2016, 22, 58. [Google Scholar] [CrossRef] [Green Version]
- Sansinenea, E.; Ortiz, A. Antimycobacterial Natural Products from Marine Pseudopterogorgia elisabethae. Curr. Org. Synth. 2016, 13, 556–568. [Google Scholar] [CrossRef]
- Chen, C.Y.; Tsai, Y.F.; Chang, W.Y.; Yang, S.C.; Hwang, T.L. Marine Natural Product Inhibitors of Neutrophil-Associated Inflammation. Mar. Drugs 2016, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, G.; Busnelli, M.; Manzini, S.; Parolini, C. Nutraceuticals and Bioactive Components from Fish for Dyslipidemia and Cardiovascular Risk Reduction. Mar. Drugs 2016, 14, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghazwi, M.; Kan, Y.Q.; Zhang, W.; Gai, W.P.; Yan, X.X. Neuroprotective Activities of Marine Natural Products from Marine Sponges. Curr. Med. Chem. 2016, 23, 360–382. [Google Scholar] [CrossRef]
- Huang, Y.; Peng, C.; Yi, Y.; Gao, B.; Shi, Q. A Transcriptomic Survey of Ion Channel-Based Conotoxins in the Chinese Tubular Cone Snail (Conus betulinus). Mar. Drugs 2017, 15, 228. [Google Scholar] [CrossRef] [PubMed]
- Molgo, J.; Marchot, P.; Araoz, R.; Benoit, E.; Iorga, B.I.; Zakarian, A.; Taylor, P.; Bourne, Y.; Servent, D. Cyclic imine toxins from dinoflagellates: A growing family of potent antagonists of the nicotinic acetylcholine receptors. J. Neurochem. 2017, 142 (Suppl. 2), 41–51. [Google Scholar] [CrossRef]
- Sadeghi, M.; McArthur, J.R.; Finol-Urdaneta, R.K.; Adams, D.J. Analgesic conopeptides targeting G protein-coupled receptors reduce excitability of sensory neurons. Neuropharmacology 2017, 127, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Kong, W.W.; Shao, C.L.; Li, Y.; Liu, Y.Z.; Liu, M.; Guan, F.F.; Wang, C.Y. Zebrafish Embryo Toxicity Microscale Model for Ichthyotoxicity Evaluation of Marine Natural Products. Mar. Biotechnol. 2016, 18, 264–270. [Google Scholar] [CrossRef]
- Agrawal, S.; Adholeya, A.; Deshmukh, S.K. The Pharmacological Potential of Non-ribosomal Peptides from Marine Sponge and Tunicates. Front. Pharmacol. 2016, 7, 333. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vilas, J.A.; Martinez-Poveda, B.; Quesada, A.R.; Medina, M.A. Aeroplysinin-1, a Sponge-Derived Multi-Targeted Bioactive Marine Drug. Mar. Drugs 2016, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Alfonso, A.; Vieytes, M.R.; Botana, L.M. Yessotoxin, a Promising Therapeutic Tool. Mar. Drugs 2016, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Valeur, E.; Gueret, S.; Adihou, H.; Gopalakrishnan, R.; Lemurell, M.; Waldmann, H.; Grossman, T.; Plowright, A. New Modalities for Challenging Targets in Drug Discovery. Angew. Chem. Int. Ed. 2017, 56, 10294–10323. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer, A.M.S.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2021, 19, 49. https://doi.org/10.3390/md19020049
Mayer AMS, Guerrero AJ, Rodríguez AD, Taglialatela-Scafati O, Nakamura F, Fusetani N. Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Marine Drugs. 2021; 19(2):49. https://doi.org/10.3390/md19020049
Chicago/Turabian StyleMayer, Alejandro M. S., Aimee J. Guerrero, Abimael D. Rodríguez, Orazio Taglialatela-Scafati, Fumiaki Nakamura, and Nobuhiro Fusetani. 2021. "Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action" Marine Drugs 19, no. 2: 49. https://doi.org/10.3390/md19020049
APA StyleMayer, A. M. S., Guerrero, A. J., Rodríguez, A. D., Taglialatela-Scafati, O., Nakamura, F., & Fusetani, N. (2021). Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Marine Drugs, 19(2), 49. https://doi.org/10.3390/md19020049