
Affinity Purification of Angiotensin Converting Enzyme Inhibitory Peptides from Wakame (Undaria Pinnatifida) Using Immobilized ACE on Magnetic Metal Organic Frameworks

Abstract
:1. Introduction
2. Results
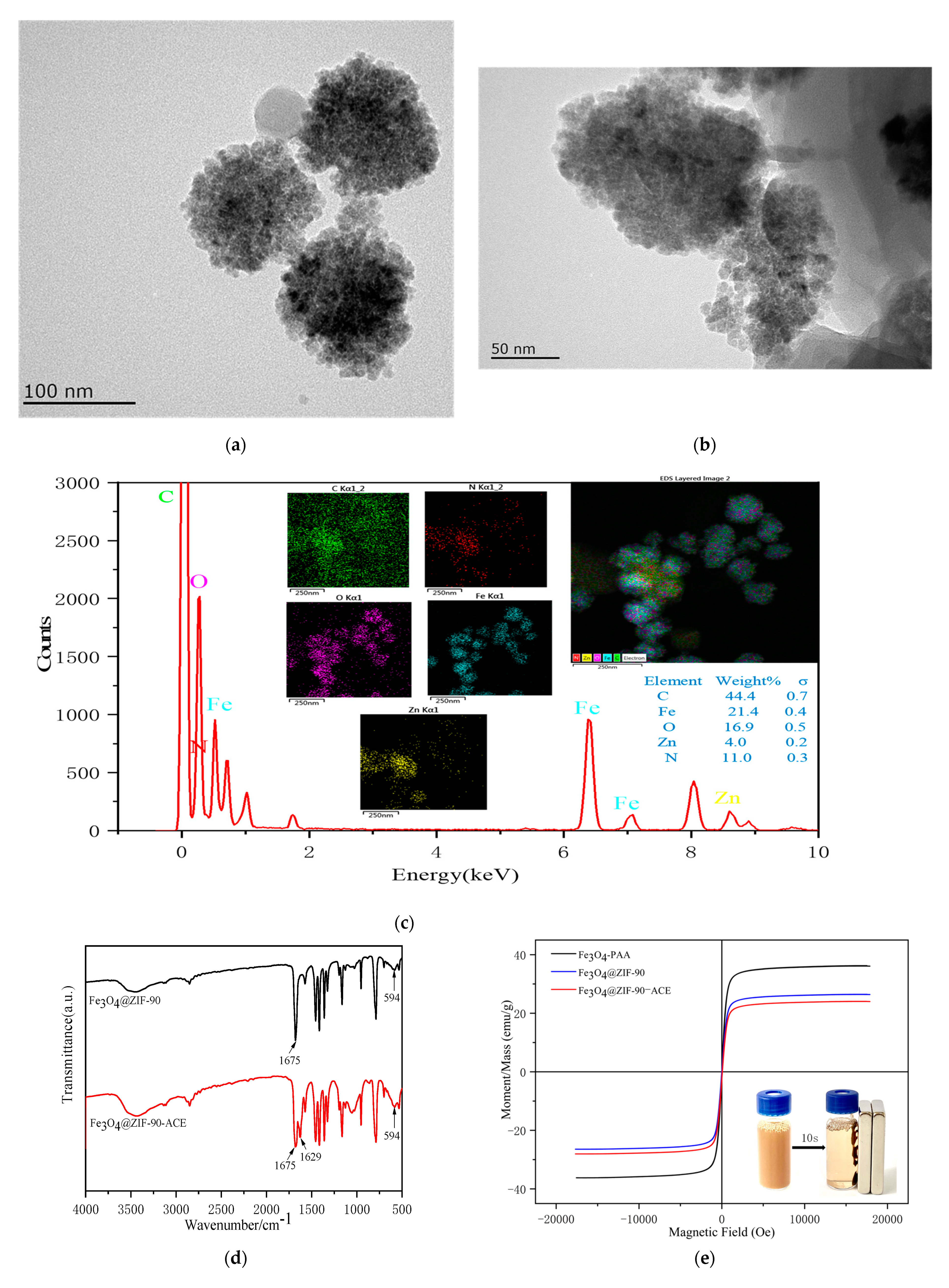
2.1. Characterization of Fe3O4@ZIF-90 and Immobilized ACE
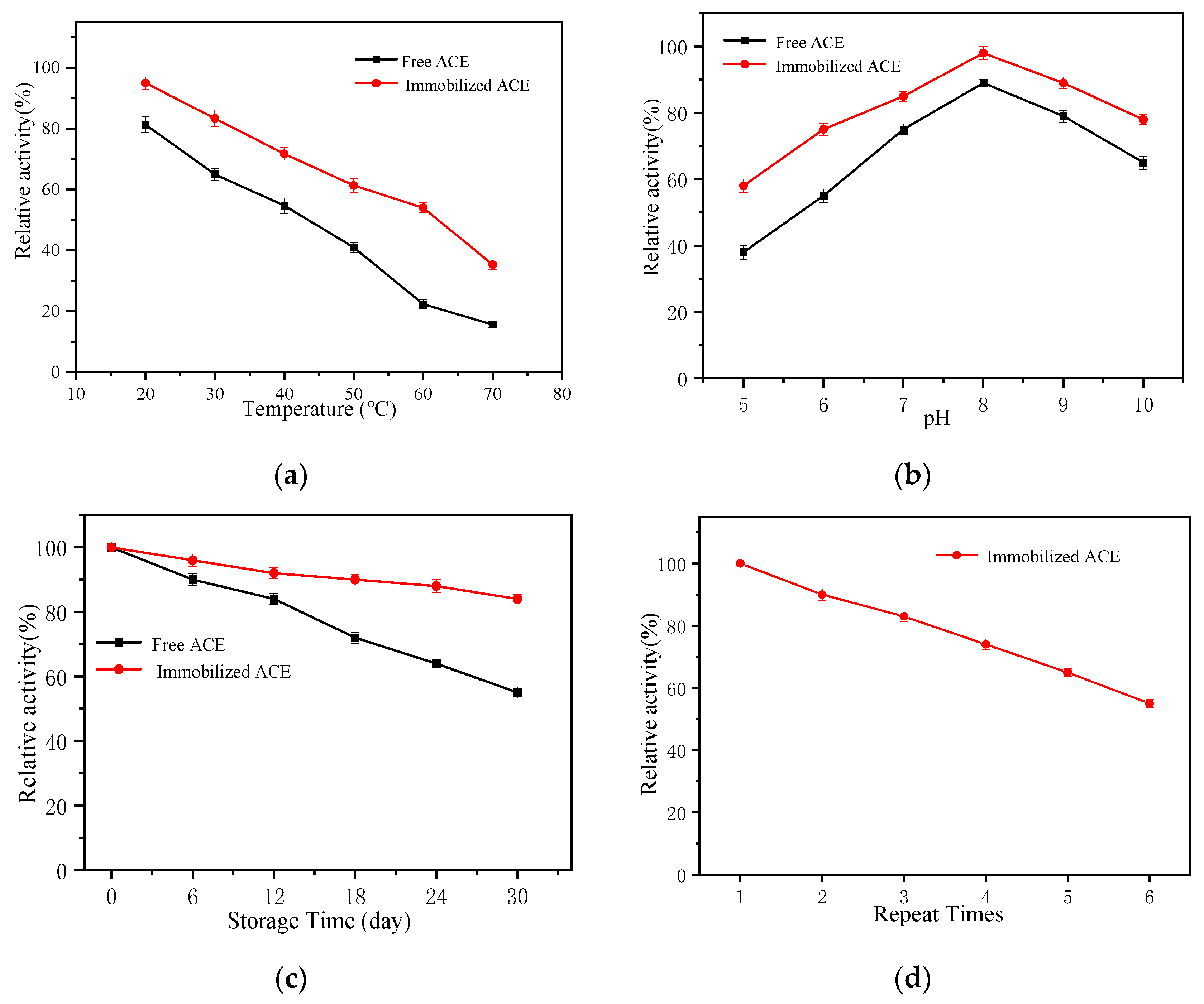
2.2. Stability of Fe3O4@ZIF-90 Immobilized ACE
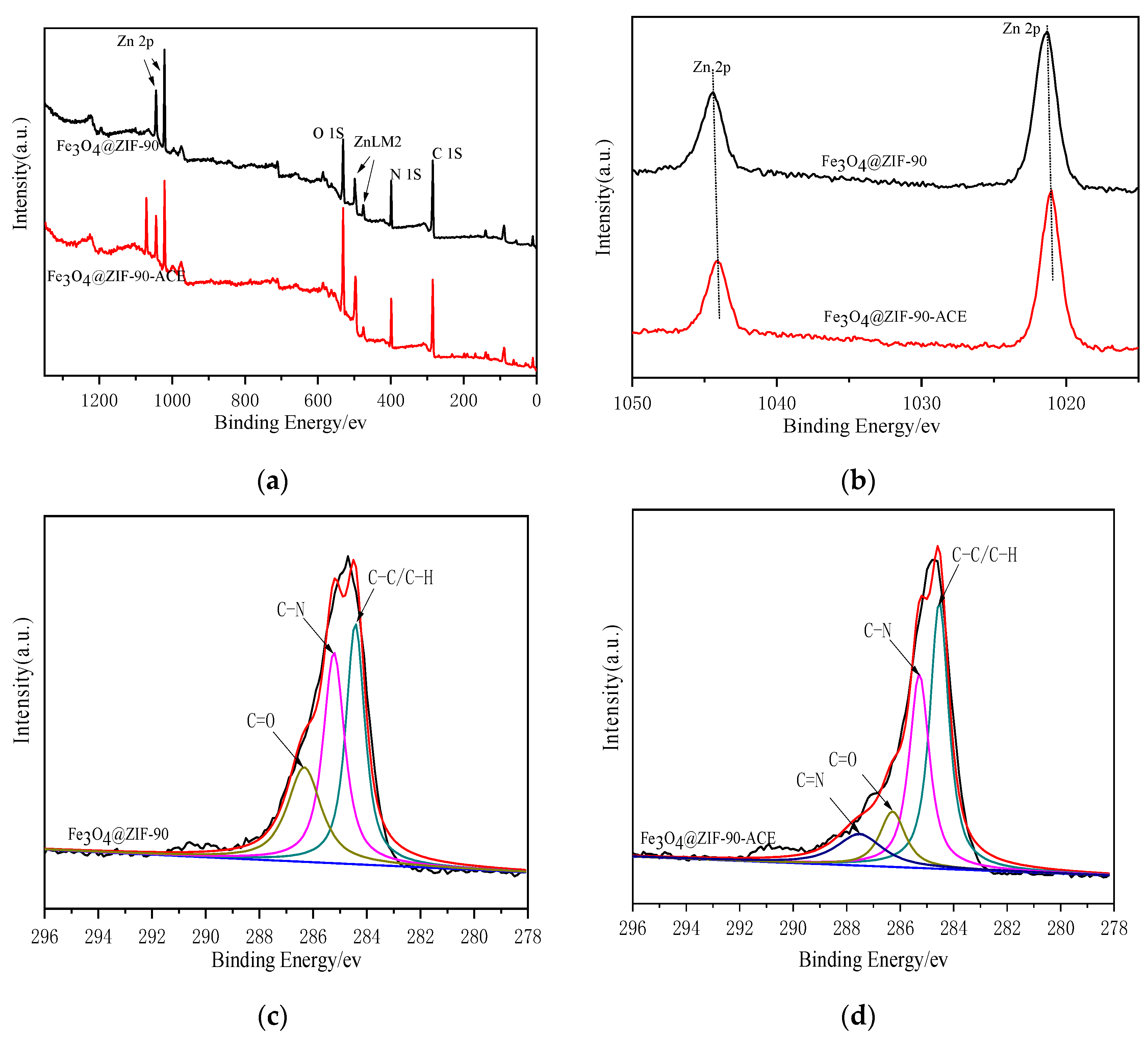
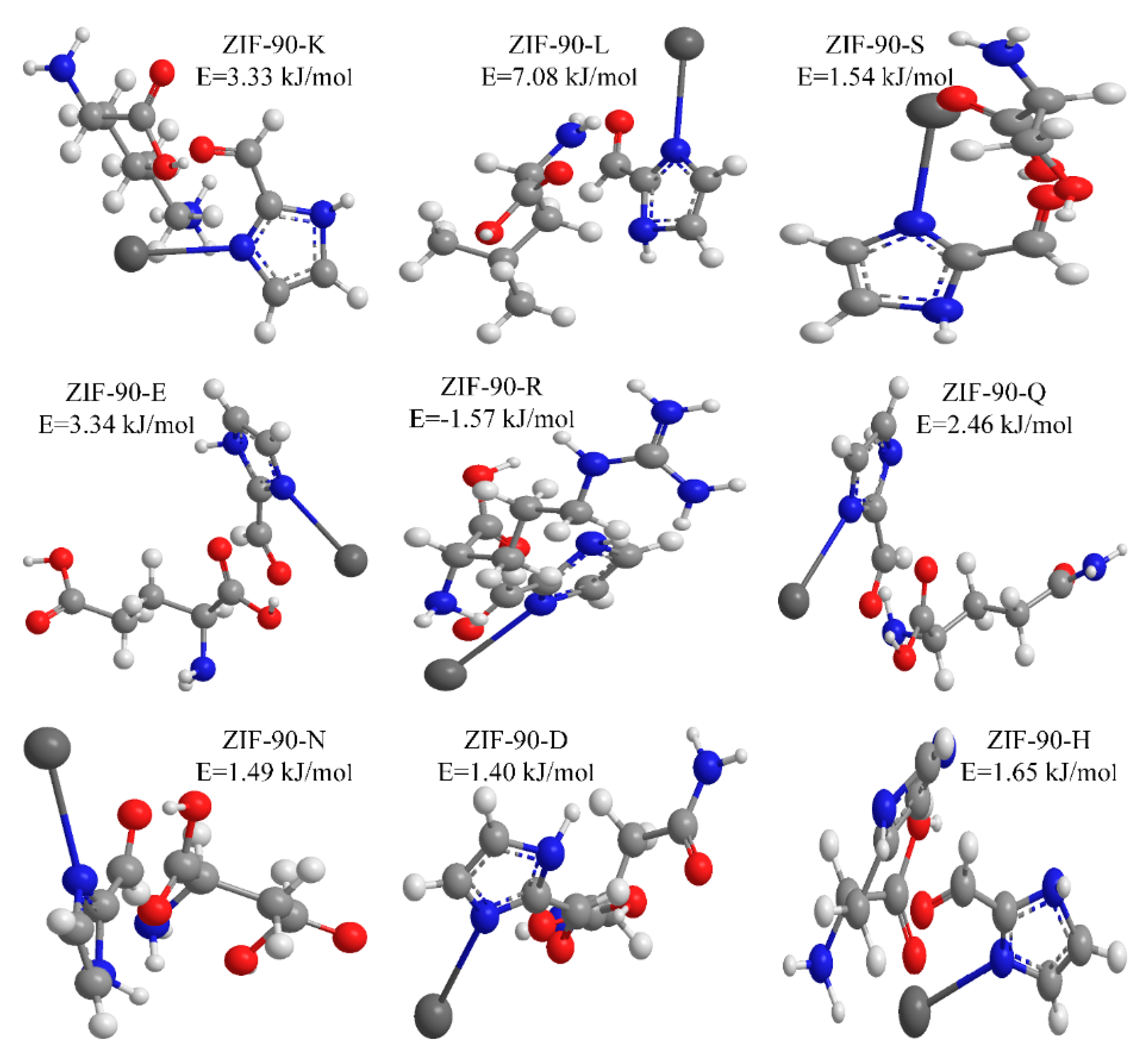
2.3. The Adsorption Mechanism of Fe3O4@ZIF-90-Immobilized ACE
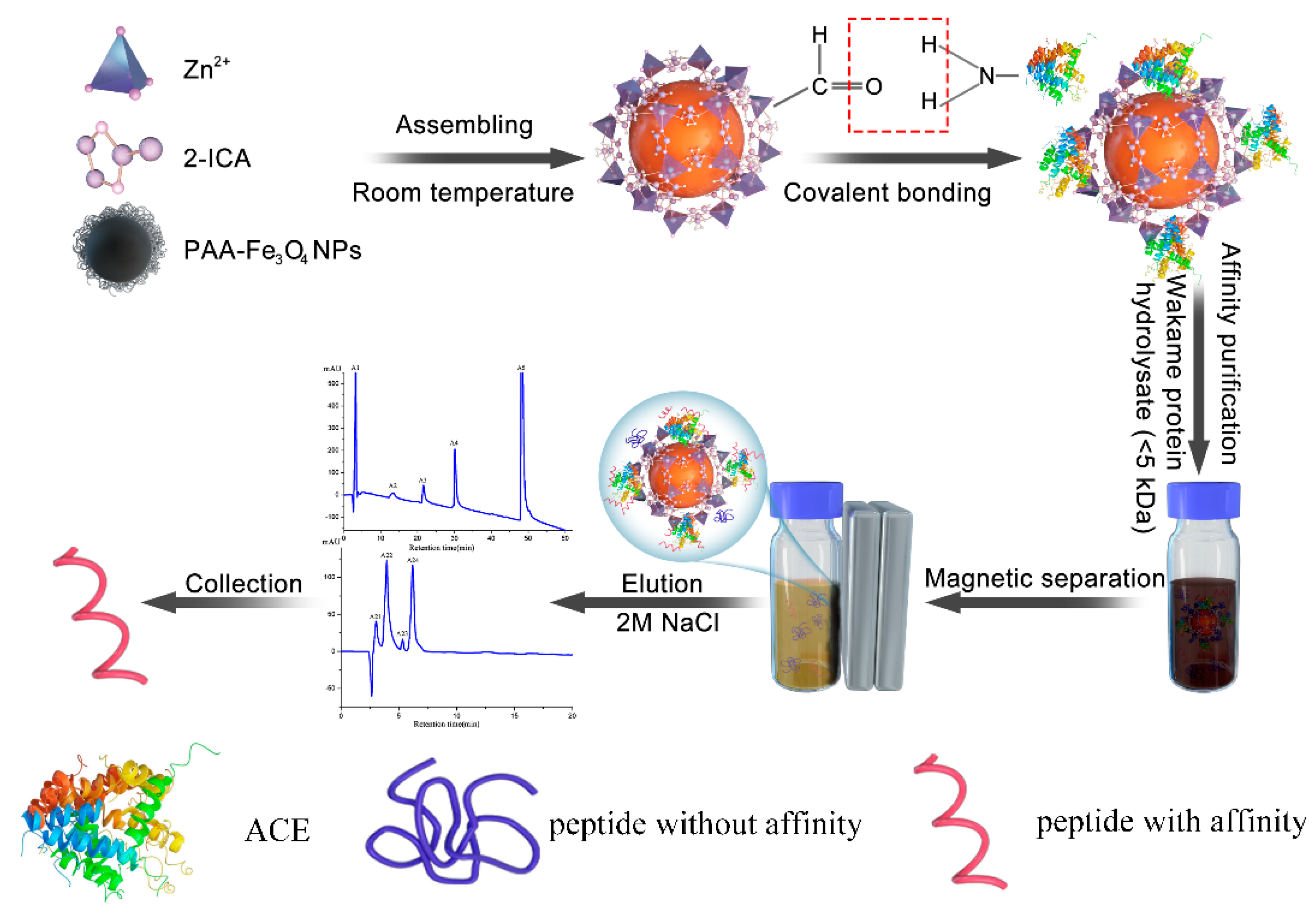
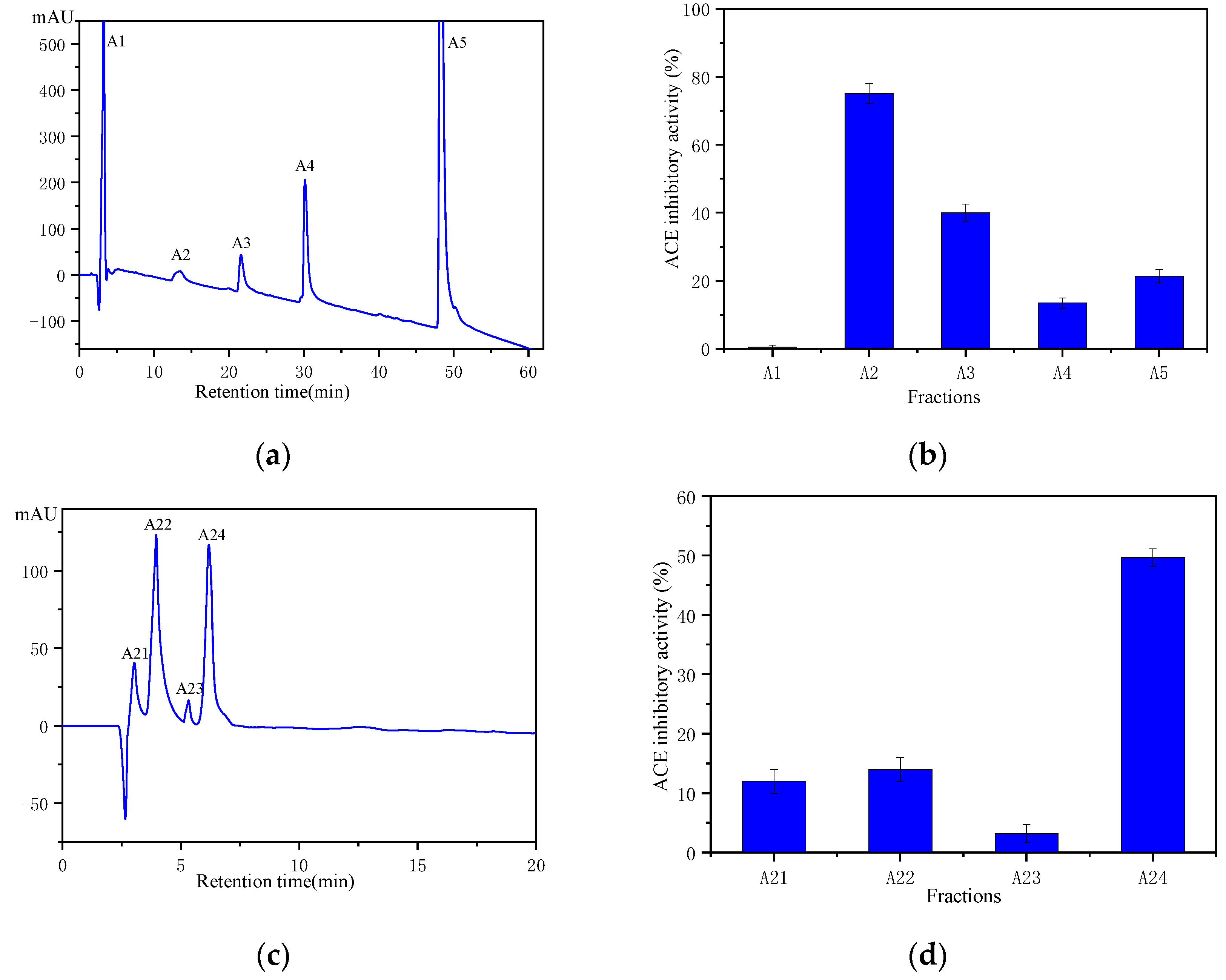
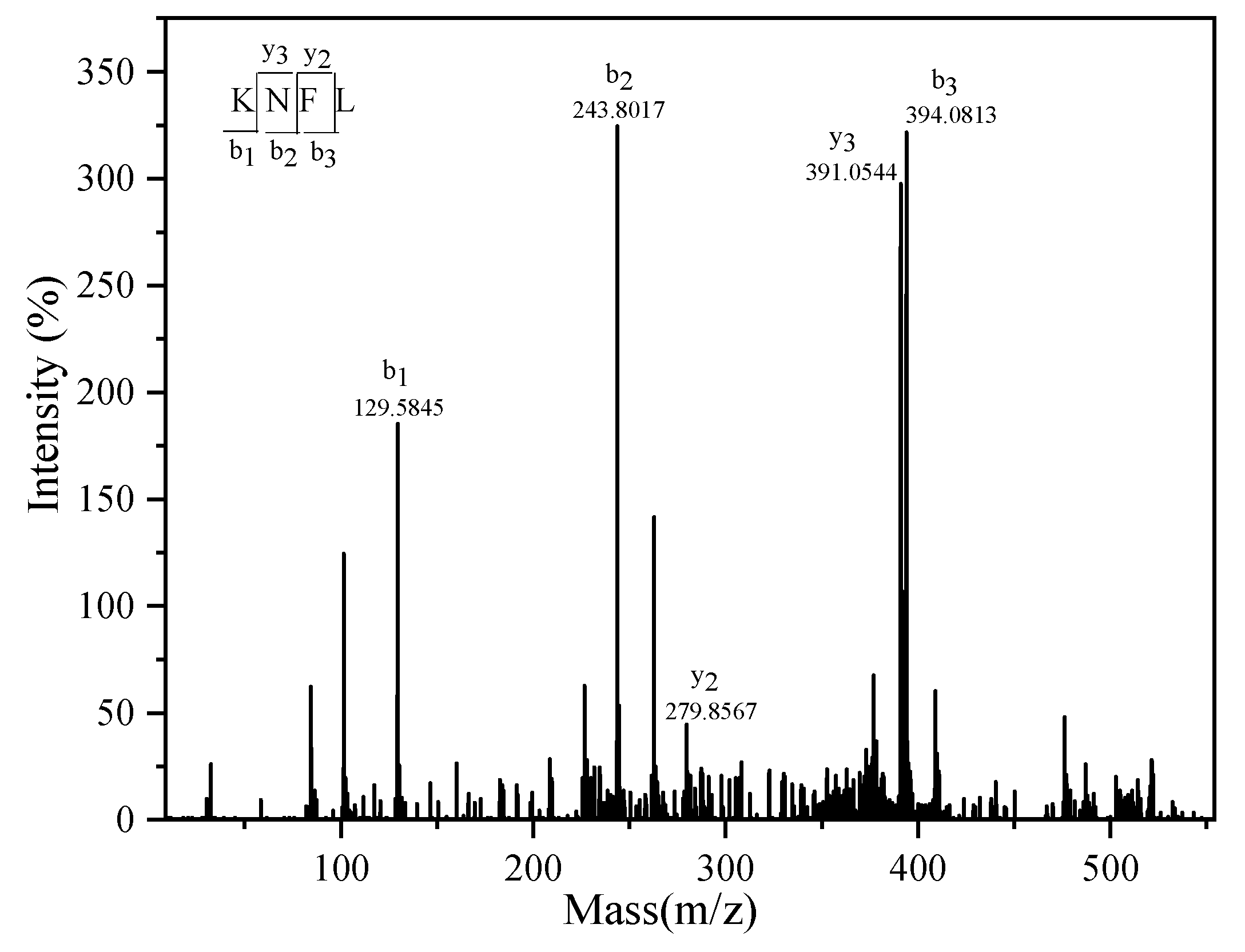
2.4. Purification and Identification of ACE Inhibitor Peptides from Wakame Protein Hydrolysate
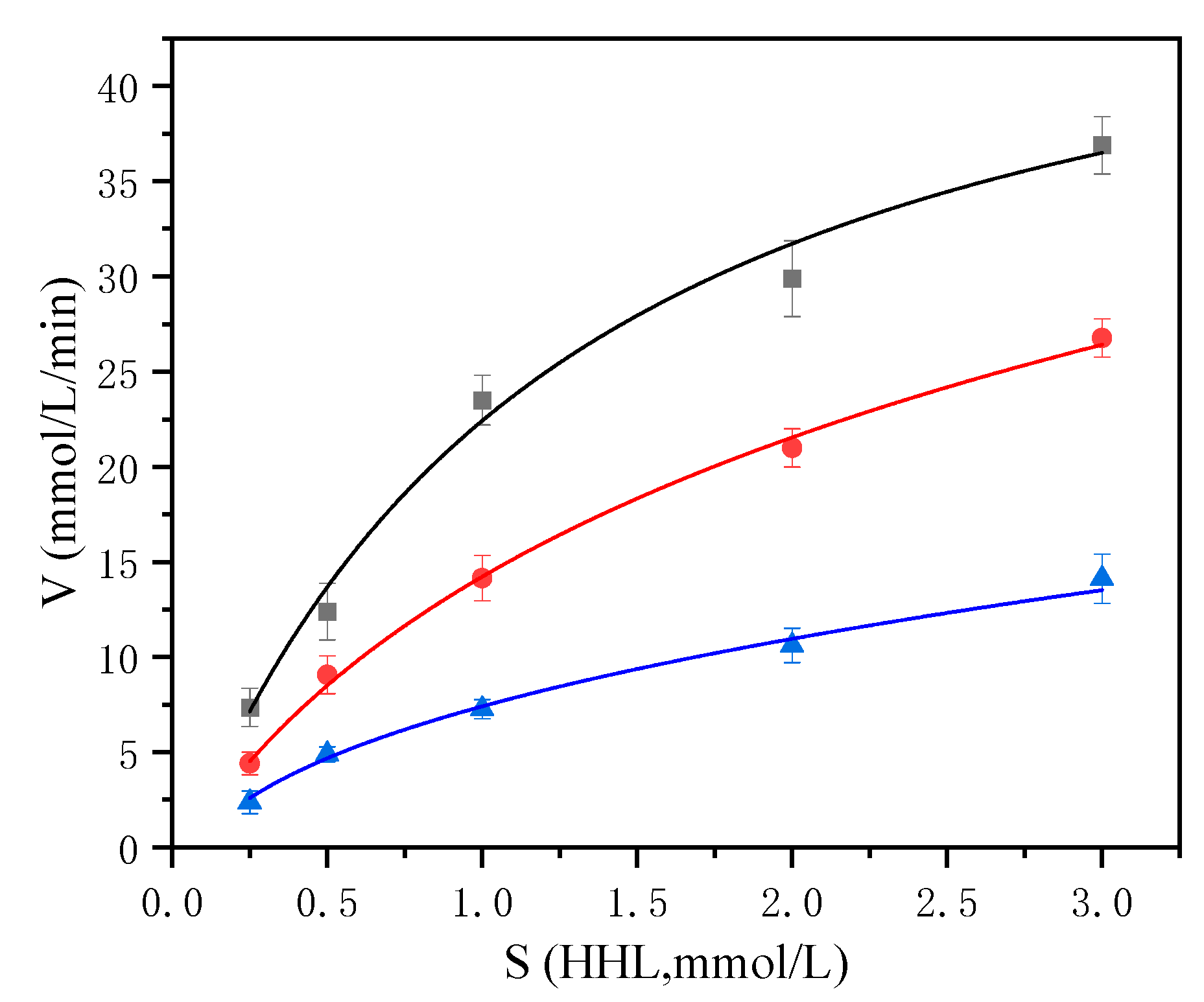
2.5. Inhibition Pattern of ACE Inhibitor Peptide KNFL
2.6. Molecular Interaction between ACE Inhibitors Peptides and ACE
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Synthesis of Fe3O4@ZIF-90
3.3. Characterization of ACE Immobilized onto Fe3O4@ZIF-90
3.3.1. Immobilization of ACE
3.3.2. The Enzymatic Stability of Immobilized ACE
3.3.3. Characterization of Fe3O4@ZIF-90 and Immobilized ACE
3.3.4. The Immobilization Mechanism of Fe3O4@ZIF-90-ACE
3.4. Purification of ACE Inhibitors Peptides from Wakame
3.4.1. Preparation of Wakame Protein Hydrolysate
3.4.2. Magnetic Affinity Purification of ACE Inhibitory Peptide from WPH
3.4.3. Characterization of ACE Inhibitory Peptide from WPH
3.5. Assay of ACE Activity and Inhibitory Activity
3.6. Molecular Docking
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vercruysse, L.; Van Camp, J.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar] [CrossRef]
- Wang, W.; Gonzalez de Mejia, E. A new frontier in soy bioactive peptides that may prevent age-related chronic diseases. Compr. Rev. Food Sci. 2005, 4, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Kuba, M.; Tanaka, K.; Tawata, S.; Takeda, Y.; Yasuda, M. Angiotensin I-converting enzyme inhibitory peptides isolated from tofuyo fermented soybean food. Biosci. Biotech. Biochem. 2003, 67, 1278–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetsuna, K.; Chen, J.R. Identification of antihypertensive peptides from peptic digest of two microalgae, Chlorella vulgaris and Spirulina platensis. Mar. Biotechnol. 2001, 3, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Yukiyoshi, A.; Doi, S.; Sugimoto, H.; Yamada, H.; Matsumoto, K. Gastrointestinal enzyme production of bioactive peptides from royal jelly protein and their antihypertensive ability in SHR. J. Nutr. Biochem. 2002, 13, 80–86. [Google Scholar] [CrossRef]
- Lan, X.D.; Liao, D.K.; Sun, J.H.; Wu, S.G.; Wang, Z.F.; Sun, J.H.; Tong, Z.F. Rapid purification and characterization of angiotensin converting enzyme inhibitory peptides from lizard fish protein hydrolysates with magnetic affinity separation. Food Chem. 2015, 182, 136–142. [Google Scholar] [CrossRef]
- Megías, C.; Pedroche, J.; Yust, M.M.; Alaiz, M.; Girón-Calle, J.; Millán, F. Immobilization of Angiotensin-Converting Enzyme on Glyoxyl-Agarose. J. Agric. Food Chem. 2006, 54, 4641–4645. [Google Scholar] [CrossRef]
- De Almeida, F.G.; Vanzolini, K.L.; Cass, Q.B. Angiotensin converting enzyme immobilized on magnetic beads as a tool for ligand fishing. J. Pharm. Biomed. Anal. 2017, 132, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.R.; Lan, X.D.; Yaseen, M.; Chai, K.G.; Zhou, L.Q.; Sun, J.H.; Lan, P.; Tong, Z.F.; Liao, D.K.; Sun, L.X. Immobilized metal affinity chromatography matrix modified by poly (ethylene glycol) methyl ether for purification of angiotensin I-converting enzyme inhibitory peptide from casein hydrolysate. J. Chromatogr. B 2020, 1143, 122042–122052. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.R.; Lan, X.D.; Yaseen, M.; Wu, S.G.; Feng, X.Z.; Zhou, L.Q.; Sun, J.H.; Liao, A.P.; Liao, D.K.; Sun, L.X. Purification, Characterization and Evaluation of Inhibitory Mechanism of ACE Inhibitory Peptides from Pearl Oyster (Pinctada fucata martensii) Meat Protein Hydrolysate. Mar. Drugs 2019, 17, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, X.D.; Chen, X.H.; Liao, D.K.; SUN, L.X.; He, X.Y.; Lu, S.S.; Wang, Y.L.; Tong, Z.F. Preparation and Properties of Magnetic Agarose Microsphere Immobilized Angiotensin Converting Enzyme. Fine Chem. 2014, 31, 1461–1465. [Google Scholar]
- Sun, M.S.; Zhang, Q.; Ma, Q.; Fu, Y.H.; Jin, W.G.; Zhu, W.B. Affinity purification of angiotensin-converting enzyme inhibitory peptides from Volutharpa ampullacea perryi protein hydrolysate using Zn-SBA-15 immobilized ACE. Eur. Food Res. Technol. 2018, 244, 457–468. [Google Scholar] [CrossRef]
- Megías, C.; Pedroche, J.; Yust, M.M.; Alaiz, M.; N-Calle, J.G.; Millaan, F.; Vioque, J. Affinity purification of angiotensin converting enzyme inhibitory peptides using immobilized ACE. J. Agric. Food. Chem. 2006, 54, 7120–7124. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Jia, B.; Zhou, J.; Liu, J.; Wang, J.C.; Ma, D.Y.; Li, P.; Chen, J. A method using angiotensin converting enzyme immobilized on magnetic beads for inhibitor screening. J. Pharm. Biomed. Anal. 2019, 164, 223–230. [Google Scholar] [CrossRef]
- Liang, J.; Liang, K. Biocatalytic Metal–Organic Frameworks: Prospects Beyond Bioprotective Porous Matrices. Adv. Funct. Mater. 2020, 30, 2001648–2001672. [Google Scholar] [CrossRef]
- Liang, S.; Wu, X.L.; Xiong, J.; Zong, M.H.; Lou, W.Y. Metal-organic frameworks as novel matrices for efficient enzyme immobilization: An update review. Coord. Chem Rev. 2020, 406, 213149–213173. [Google Scholar] [CrossRef]
- Park, K.; Ni, Z.; Cote, A.; Choi, J.Y.; Huang, R.; Uribe-Romo, F.J.; Chae, H.K.; O’Keeffe, M.; Yaghi, O.M. Exceptional chemical and thermal stability of zeolitic imidazolate frameworks. Proc. Natl. Acad. Sci. USA 2006, 103, 10186–10191. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, S.; Feng, F.; Lu, L. Simultaneous purification and immobilization of laccase on magnetic zeolitic imidazolate frameworks: Recyclable biocatalysts with enhanced stability for dye decolorization. Biochem Eng J. 2019, 150, 107285–107292. [Google Scholar] [CrossRef]
- Zhong, C.; Lei, Z.; Huang, H.; Zhang, M.Y.; Cai, Z.W.; Lin, Z.A. One-pot synthesis of trypsin-based magnetic metal–organic frameworks for highly efficient proteolysis. J. Mater. Chem. B. 2020, 8, 4642–4647. [Google Scholar] [CrossRef]
- Abudiab, T.; Beitle, R.R. Preparation of magnetic immobilized metal affinity separation media and its use in the isolation of proteins. J. Chromatogr A. 1998, 795, 211. [Google Scholar] [CrossRef]
- Zou, R.; Gong, Q.; Zheng, J.; Shi, Z.Z.; Zheng, J.P.; Xing, J.; Liu, C.; Jiang, Z.Q.; Wu, A.G. A ZIF-90 Nanoplatform Loading with Enzyme-Responsive Organic Small Molecule Probe for Imaging Hypoxia Status of Tumor Cells. Nanoscale 2020, 12, 14870–14881. [Google Scholar] [CrossRef]
- Nosike, E.I.; Jiang, Z.; Miao, L.; Akakuru, O.U.; Yuan, B.; Wu, S.S.; Zhang, Y.N.; Zhang, Y.J.; Wu, A. A novel hybrid nanoadsorbent for effective Hg2+ adsorption based on zeolitic imidazolate framework (ZIF-90) assembled onto poly acrylic acid capped Fe3O4 nanoparticles and cysteine. J. Hazard Mater. 2020, 392, 122288–122298. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Shi, Y.; Yan, L.; Wu, X.R.; Zhu, J.H.; Zhao, D.B.; Zaved, H.K.; Liu, X.H. Development of an on-line immobilized-glucosidase microreactor coupled to liquid chromatography for screening of α-glucosidase inhibitors. J. Pharm. Biomed. 2019, 180, 113047. [Google Scholar] [CrossRef]
- Zhai, R.; Yuan, Y.; Jiao, F.; Hao, F.R.; Fang, X.; Zhang, Y.J.; Qian, X.H. Facile synthesis of magnetic metal organic frameworks for highly efficient proteolytic digestion used in mass spectrometry-based proteomics. Anal. Chim. Acta. 2017, 994, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Yamori, Y.; Nara, Y.; Tsubouchi, T.; Sogawa, Y.; Ikeda, K.; Horie, R. Dietary prevention of stroke and its mechanisms in stroke-prone spontaneously hypertensive rats preventive effect of dietary fibre and palmitoleic acid. J. Hypertens. Suppl. 1986, 4, 449–452. [Google Scholar]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A.; Nakano, T. Angiotensin I-Converting Enzyme Inhibitory Peptides Derived from Wakame (Undaria pinnatifida) and Their Antihypertensive Effect in Spontaneously Hypertensive Rats. J. Agric. Food. Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef]
- Suetsuna, K.; Maekawa, K.; Chen, J.R. Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J. Nutr. Biochem. 2004, 15, 267–272. [Google Scholar] [CrossRef]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Wu, S.G.; Feng, X.Z.; Lan, X.D.; Xu, Y.J.; Liao, D.K. Purification and identification of Angiotensin-I Converting Enzyme (ACE) inhibitory peptide from lizard fish (Saurida elongata) hydrolysate. J. Funct. Foods 2015, 13, 295–299. [Google Scholar] [CrossRef]
- Kharazmi, S.; Taheri-Kafrani, A.; Soozanipour, A.; Nasrollahzadeh, M.; Varma, R.S. Xylanase immobilization onto trichlorotriazine-functionalized polyethylene glycol grafted magnetic nanoparticles: A thermostable and robust nanobiocatalyst for fruit juice clarification. Int. J. Biol. Macromol. 2020, 163, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.L.; Xu, H.; Lai, L.H.; Gu, W.M.; Xu, P.; Xiong, J.; Yin, H.; Li, X.H.; Ma, Y.Z.; Zhou, J.; et al. Magnetic ZIF-8/cellulose/Fe3O4 nanocomposite: Preparation, characterization, and enzyme immobilization. Bioresour. Bioprocess. 2017, 4, 56–63. [Google Scholar]
- Thanh, M.T.; Thien, T.V.; Du, P.D.; Hung, N.P.; Khieu, D.Q. Iron doped zeolitic imidazolate framework (Fe-ZIF-8): Synthesis and photocatalytic degradation of RDB dye in Fe-ZIF-8. J. Porous Mater. 2018, 25, 857–869. [Google Scholar] [CrossRef]
- Xie, W.; Wan, F. Guanidine post-functionalized crystalline ZIF-90 frameworks as a promising recyclable catalyst for the production of biodiesel via soybean oil transesterification. Energy Convers. Manag. 2019, 198, 111922–111933. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, C.; Wang, S.; Zhou, Y. Experimental and DFT study of selective adsorption mechanisms of Pb(II) by UiO-66-NH2 modified with 1,8-dihydroxy-anthraquinone. J. Ind. Eng. Chem. 2020, 83, 111–122. [Google Scholar] [CrossRef]
- Zhang, F.M.; Dong, H.; Zhang, X.; Sun, X.J.; Sun, X.J.; Liu, M.; Yang, D.D.; Liu, X.; Wei, J.Z. Post synthetic Modification of ZIF-90 for Potential Targeted Codelivery of Two Anticancer Drugs. ACS Appl. Mater. Interfaces 2017, 9, 27332–27337. [Google Scholar] [CrossRef]
- Malinowski, S.; Jaroszyńska-Wolińska, J.; Herbert, P.A.F. Theoretical insight into plasma deposition of laccase bio-coating formation. J. Mater. Sci. 2019, 54, 10746–10763. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Scheicher, R.H.; Pandey, R.; Karna, S.P. Sensitivity of Boron Nitride Nanotubes toward Biomolecules of Different Polarities. J. Phys. Chem. Lett. 2011, 2, 2442–2447. [Google Scholar] [CrossRef]
- Qin, Z.; Lin, S.Y.J.; Chen, Q.M.; Zhang, Y.; Zhou, J.C.; Zhao, L.M. One-step immobilization-purification of enzymes by carbohydrate-binding module family 56 tag fusion. Food Chem. 2019, 299, 125037–125046. [Google Scholar] [CrossRef]
- Sun, L.X.; Wu, S.G.; Zhou, L.Q.; Wang, F.; Lan, X.D.; Sun, J.H.; Tong, Z.F.; Liao, D.K. Separation and Characterization of Angiotensin I Converting Enzyme (ACE) Inhibitory Peptides from Saurida elongata Proteins Hydrolysate by IMAC-Ni2+. Mar. Drugs 2017, 15, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, A.; Sakurai, T.; Ochi, D.; Mitsuyama, E.; Yamauchi, K.; Abe, F. Novel angiotensin I-converting enzyme inhibitory peptide derived from bovine casein. Food Chem. 2013, 141, 3781–3789. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.G.; Feng, X.Z.; Lu, Y.T.; Liu, S.S.; Tian, Y.H. Purification of angiotensin I-converting enzyme (ACE) inhibitory peptides from casein hydrolysate by IMAC-Ni2+. Amino Acids. 2017, 49, 1787–1791. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-Originating ACE Inhibitors, Including Antihypertensive Peptides, as Preventive Food Components in Blood Pressure Reduction. Compr. Rev. Food Sci. Food Safety 2014, 13, 114–134. [Google Scholar]
- Martínez-Maqueda, D.; Miralles, B.; Recio, I.; Hernandez-Ledesma, B. Antihypertensive peptides from food proteins: A review. Food Funct. 2012, 3, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.B.; Wang, J.P.; Liao, W.; Jiang, X.; Wu, J.P. Identification and Characterization of Gastrointestinal-Resistant Angiotensin-Converting Enzyme Inhibitory Peptides from Egg White Proteins. J. Agric. Food Chem. 2019, 67, 7147–7156. [Google Scholar] [CrossRef] [PubMed]
- Matsufuji, H.; Matsui, T.; Seki, E.; Osajima, K.; Nakashima, M. Angiotensin I-converting Enzyme Inhibitory Peptides in an Alkaline Protease Hydrolyzate Derived from Sardine Muscle. Biosci. Biotechnol. Biochem. 1994, 58, 2244. [Google Scholar] [CrossRef]
- Maruyama, S.; Mitachi, H.; Tanaka, H.; Noboru Tomizukaa, N.; Suzuki, H. Studies on the Active Site and Antihypertensive Activity of Angiotensin I-Converting Enzyme Inhibitors Derived from Casein. Agric. Biol. Chem. 1987, 51, 1581–1586. [Google Scholar] [CrossRef] [Green Version]
- Mine, Y.; Shahidi, F. Nutraceutical Proteins and Peptides in Health and Disease; CRC Press: Hoboken, NJ, USA, 2006; Volume 56, pp. 96–108. [Google Scholar]
- Gu, Y.; Wu, J. LC–MS/MS coupled with QSAR modeling in characterising of angiotensin I-converting enzyme inhibitory peptides from soybean proteins. Food Chem. 2013, 141, 2682–2690. [Google Scholar] [CrossRef]
- Balti, R.; Bougatef, A.; Sila, A.; Guillochon, D.; Dhulster, P.; Nedjar-Arroume, N. Nine novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia officinalis) muscle protein hydrolysates and antihypertensive effect of the potent active peptide in spontaneously hypertensive rats. Food Chem. 2015, 170, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.L.; Wang, C.; Chen, C.; Zhang, R.Y.; Liu, H.X.; Lu, W.H.; Jiang, L.Z.; Du, M. Identification of a novel ACE-inhibitory peptide from casein and evaluation of the inhibitory mechanisms. Food Chem. 2018, 256, 98–104. [Google Scholar] [CrossRef]
- Forghani, B.; Zarei, M.; Ebrahimpour, A.; Philip, R.; Bakar, J.; Hamid, A.A.; Saari, N. Purification and characterization of angiotensin converting enzyme-inhibitory peptides derived from Stichopus horrens: Stability study against the ACE and inhibition kinetics. J. Funct. Food 2016, 20, 276–290. [Google Scholar] [CrossRef]
- Qi, C.; Zhang, R.; Liu, F.; Zheng, T.; Wu, W.J. Molecular mechanism of interactions between inhibitory tripeptide GEF and angiotensin-converting enzyme in aqueous solutions by molecular dynamic simulations. J. Mol. Liq. 2017, 249, 389–396. [Google Scholar] [CrossRef]
- Lan, X.D.; Sun, L.X.; Muhammad, Y.; Wang, Z.F.; Liu, H.B.; Sun, J.H.; Zhou, L.Q.; Feng, X.Z.; Liao, D.K.; Wang, S.F. Studies on the Interaction between Angiotensin-Converting Enzyme (ACE) and ACE Inhibitory Peptide from Saurida elongate. J. Agric. Food Chem. 2018, 66, 13414–13422. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Tamires, C.; Adejanildo, S.; Bonomo, R.C.F.; Franco, M.; Finotelli, P.V.; Amarala, P.F.F. Simple physical adsorption technique to immobilize Yarrowia lipolytica lipase purified by different methods on magnetic nanoparticles: Adsorption isotherms and thermodynamic approach. Int. J. Biol. Macromol. 2020, 160, 889–902. [Google Scholar]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from Ulva rigida C. Agardh protein hydrolysate. J. Funct. Foods. 2016, 26, 65–76. [Google Scholar] [CrossRef]
- Liao, P.Y.; Lan, X.D.; Liao, D.K.; Sun, L.X.; Zhou, L.Q.; Sun, J.H. Isolation and Characterization of Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from the Enzymatic Hydrolysate of Carapax Trionycis (the Shell of the Turtle Pelodiscus sinensis). J. Agric. Food Chem. 2018, 66, 7015–7022. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources | Amino Sequence | IC50 | Method of Purification | Reference |
|---|---|---|---|---|
| lizard fish | RVCLP | 175 µM | Conventional approach | [29] |
| RYRP | 52 µM | Affinity purification/ IMAC-Ni2+ | [39] | |
| GMKCAF | 45.7 ± 1.1 µM | Affinity purification/ Magnetic agarose-ACE | [6] | |
| casein | MKP | 0.3 μM | Conventional approach | [40] |
| WYLHYA | 16.2 μM | Affinity purification/ IMAC-Ni2+ | [41] | |
| LLYQEPVLGPVR | 274 ± 5 μM | Affinity purification/ IMAM @ mPEG | [9] | |
| Pinctada fucata martensii | HLHT/GWA | 458.06 ± 3.24 μM/ 109.25 ± 1.45 μM | Affinity purification/ IMAM @ mPEG | [10] |
| Volutharpa ampullaceal perryi | IVTNWDDMGK/ VGPAGRRG | 2.08 mM/4.66 mM | Affinity purification/ Zn-SBA-15-ACE | [12] |
| Wakame | AIYK/YKYY/ KFYG/YNKL | 213 μM/64.2 μM/ 90.5 μM/21 μM/ | Conventional approach | [28] |
| KNFL | 225.87 ± 2.70 µM | Affinity purification/ Fe3O4@ZIF-90-ACE | This study |
| Source | Amino Sequence | IC50 (μM) | Reference |
|---|---|---|---|
| Soy | FFL | 37.00 | [2] |
| Soybean | IFL | 44.67 | [3] |
| Microalgae | AFL | 63.10 | [4] |
| Royal jelly | FNF | 6.92 | [5] |
| Garlic | NF | 46.30 | [47] |
| Soybean | LNF | 511.4 | [48] |
| Cuttlefish | KNGDGY | 51.63 | [49] |
| Wakame | KNFL | 225.87 | this study |
| Kinetics Parameters | Control | KNFL (192 µM) | KNFL (384 µM) |
|---|---|---|---|
| Km (mM/L) | 1.564 | 2.238 | 2.278 |
| Vmax (µM/L·min) | 55.492 | 45.958 | 24.105 |
| Amino Acid | Lipophilicity Parameters | Number of Amino Acids | Combination Mode |
|---|---|---|---|
| L (Leu) | 3.8 | 62 | Covalent Binding /Physical Adsorption |
| K (Lys) | −3.9 | 30 | Covalent Binding |
| R (Arg) | −4.5 | 26 | Covalent Binding |
| S (Ser) | −0.8 | 33 | Physical Adsorption |
| E (Glu) | −3.5 | 40 | Physical Adsorption |
| Q (Gln) | −3.5 | 33 | Physical Adsorption |
| N (Asn) | −3.5 | 31 | Physical Adsorption |
| D (Asp) | −3.5 | 30 | Physical Adsorption |
| H (His) | −3.2 | 21 | Physical Adsorption |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Liao, D.; Sun, L.; Wu, S.; Lan, P.; Wang, Z.; Li, C.; Zhou, Q.; Lu, Y.; Lan, X. Affinity Purification of Angiotensin Converting Enzyme Inhibitory Peptides from Wakame (Undaria Pinnatifida) Using Immobilized ACE on Magnetic Metal Organic Frameworks. Mar. Drugs 2021, 19, 177. https://doi.org/10.3390/md19030177
Feng X, Liao D, Sun L, Wu S, Lan P, Wang Z, Li C, Zhou Q, Lu Y, Lan X. Affinity Purification of Angiotensin Converting Enzyme Inhibitory Peptides from Wakame (Undaria Pinnatifida) Using Immobilized ACE on Magnetic Metal Organic Frameworks. Marine Drugs. 2021; 19(3):177. https://doi.org/10.3390/md19030177
Chicago/Turabian StyleFeng, Xuezhen, Dankui Liao, Lixia Sun, Shanguang Wu, Ping Lan, Zefen Wang, Chunzhi Li, Qian Zhou, Yuan Lu, and Xiongdiao Lan. 2021. "Affinity Purification of Angiotensin Converting Enzyme Inhibitory Peptides from Wakame (Undaria Pinnatifida) Using Immobilized ACE on Magnetic Metal Organic Frameworks" Marine Drugs 19, no. 3: 177. https://doi.org/10.3390/md19030177
APA StyleFeng, X., Liao, D., Sun, L., Wu, S., Lan, P., Wang, Z., Li, C., Zhou, Q., Lu, Y., & Lan, X. (2021). Affinity Purification of Angiotensin Converting Enzyme Inhibitory Peptides from Wakame (Undaria Pinnatifida) Using Immobilized ACE on Magnetic Metal Organic Frameworks. Marine Drugs, 19(3), 177. https://doi.org/10.3390/md19030177