
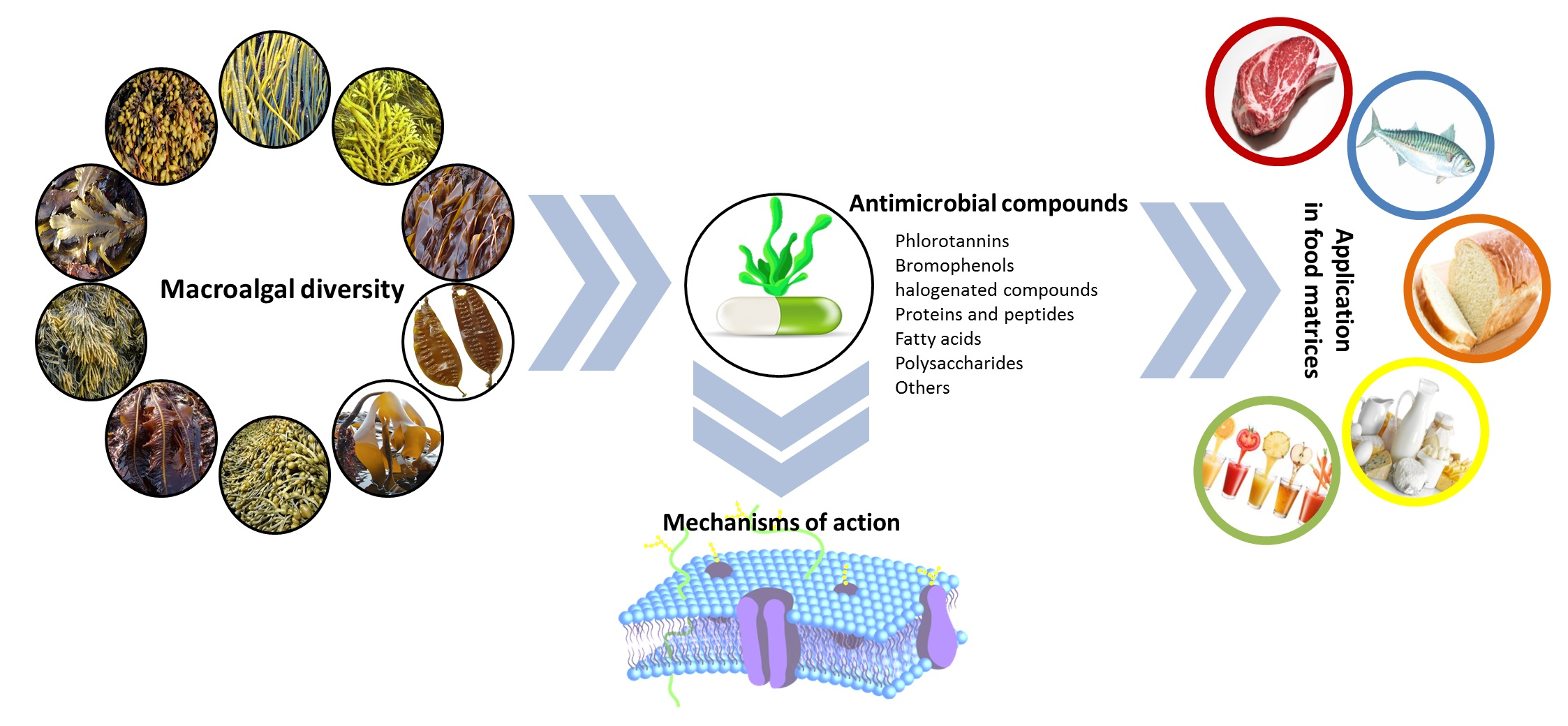
Antimicrobials from Seaweeds for Food Applications

 ,
,  , ,
, , 

Abstract
:
1. Introduction
2. Methods to Test the Antimicrobial Properties of Novel Compounds
2.1. Antimicrobial Susceptibility Methods
2.1.1. Disc Diffusion Method
2.1.2. Dilution Method
2.1.3. Molecular AMR Methods
2.2. In Vivo Assays
3. Macroalgal Compounds with Antimicrobial Properties and Their Mechanisms of Action
3.1. Phenolic Compounds
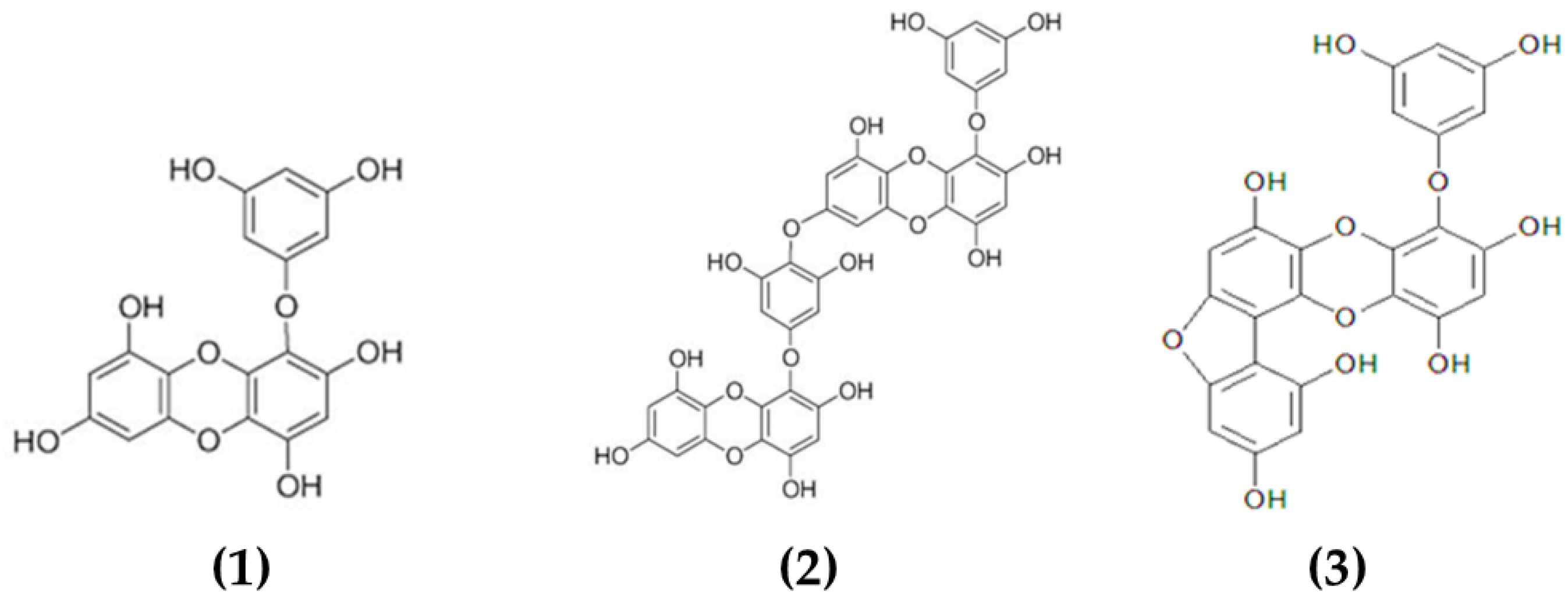
3.1.1. Phlorotannins
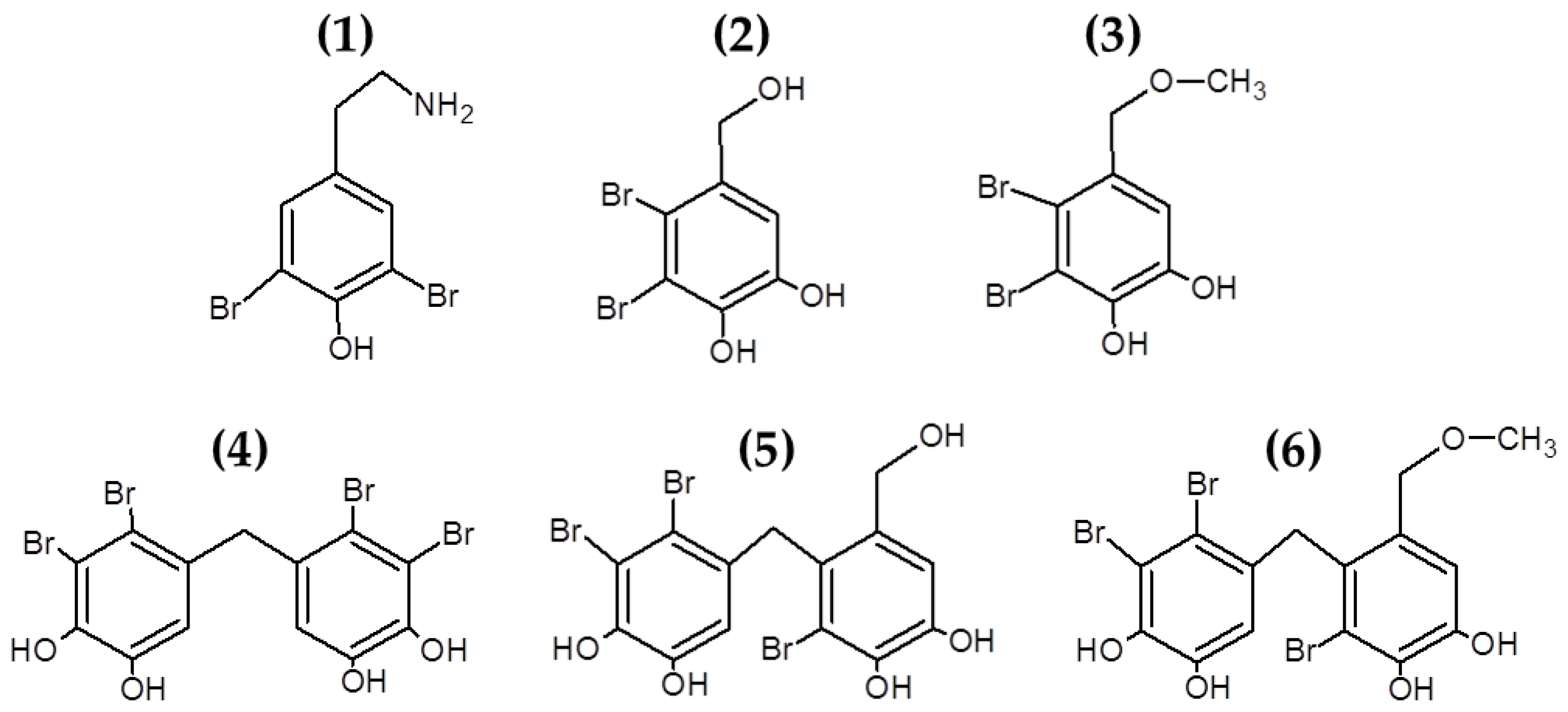
3.1.2. Bromophenols and Other Halogenated Compounds
3.2. Proteins and Peptides
3.3. Fatty Acids
3.4. Polysaccharides
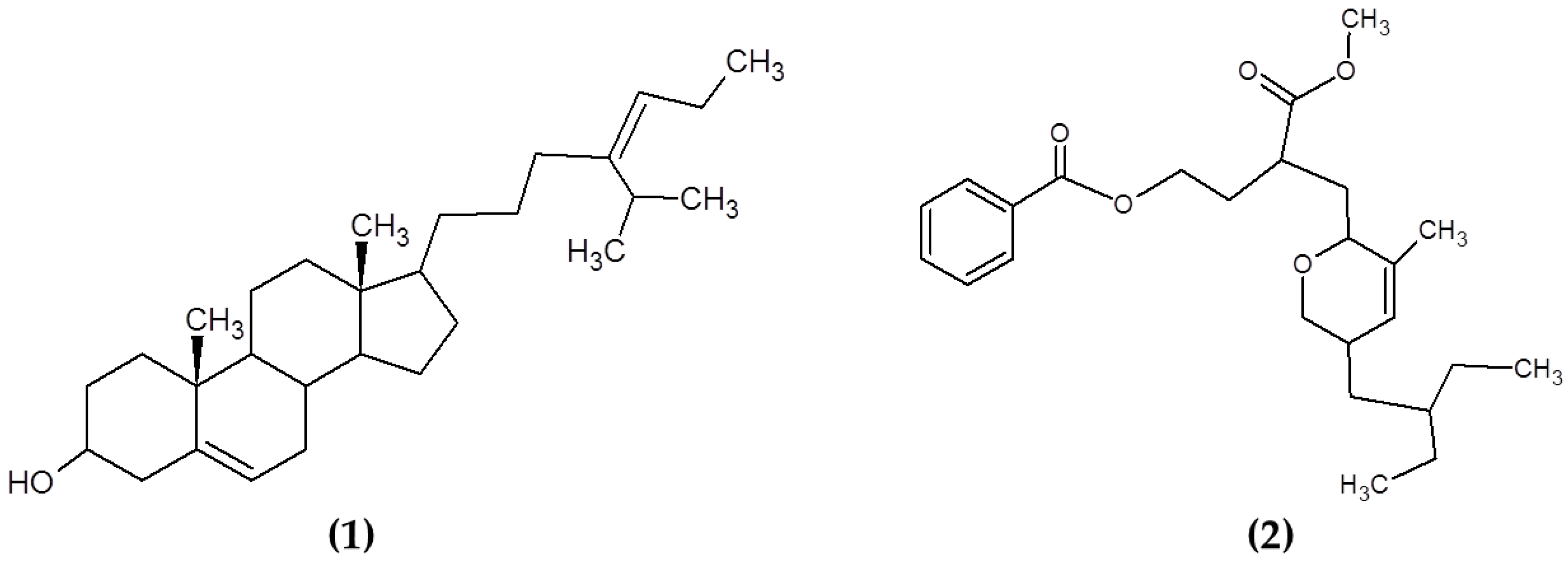
3.5. Other Compounds
4. Incorporation of Antimicrobial Compounds from Macroalgae in Food Matrices
4.1. Meat and Meat Products
4.2. Milk and Dairy Products
4.3. Other Food Products
5. Future Trends and Challenges of the Use of Antimicrobials from Seaweeds for Food Applications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barba, F.J.; de Souza Sant’Ana, A.; Orlien, V.; Koubaa, M. Innovative Technologies for Food Preservation: Inactivation of Spoilage and Pathogenic Microorganisms; Academic Press: London, UK, 2017. [Google Scholar]
- Tiwari, B.K.; Valdramidis, V.P.; O’Donnell, C.P.; Muthukumarappan, K.; Bourke, P.; Cullen, P.J. Application of Natural Antimicrobials for Food Preservation. J. Agric. Food Chem. 2009, 57, 5987–6000. [Google Scholar] [CrossRef] [Green Version]
- Abrahamian, F.M.; Goldstein, E.J. Microbiology of animal bite wound infections. Clin. Microbiol. Rev. 2011, 24, 231–246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.-N.; Tang, G.-Y.; Li, H.-B. Antibacterial and Antifungal Activities of Spices. Int. J. Mol. Sci. 2017, 18, 1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Abadias, M.; Colás-Medà, P.; Usall, J.; Viñas, I. Biopreservative methods to control the growth of foodborne pathogens on fresh-cut lettuce. Int. J. Food Microbiol. 2015, 214, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Vardaka, V.D.; Yehia, H.M.; Savvaidis, I.N. Effects of Citrox and chitosan on the survival of Escherichia coli O157:H7 and Salmonella enterica in vacuum-packaged turkey meat. Food Microbiol. 2016, 58, 128–134. [Google Scholar] [CrossRef]
- Miyashita, K.; Mikami, N.; Hosokawa, M. Chemical and nutritional characteristics of brown seaweed lipids: A review. J. Funct. Foods 2013, 5, 1507–1517. [Google Scholar] [CrossRef]
- Susanto, E.; Fahmi, A.S.; Abe, M.; Hosokawa, M.; Miyashita, K. Lipids, fatty acids, and fucoxanthin content from temperate and tropical brown seaweeds. Aquat. Procedia 2016, 7, 66–75. [Google Scholar] [CrossRef]
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. Globefish Res. Programme 2018, 124, I. [Google Scholar]
- Choudhary, A.; Naughton, L.M.; Montánchez, I.; Dobson, A.D.W.; Rai, D.K. Current Status and Future Prospects of Marine Natural Products (MNPs) as Antimicrobials. Mar. Drugs 2017, 15, 272. [Google Scholar] [CrossRef] [PubMed]
- Roohinejad, S.; Koubaa, M.; Barba, F.J.; Saljoughian, S.; Amid, M.; Greiner, R. Application of seaweeds to develop new food products with enhanced shelf-life, quality and health-related beneficial properties. Food Res. Int. 2017, 99, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Hayes, M. Red and green macroalgae for fish and animal feed and human functional food development. Food Rev. Int. 2016, 32, 15–45. [Google Scholar] [CrossRef]
- Ji, N.-Y.; Li, X.-M.; Li, K.; Ding, L.-P.; Gloer, J.B.; Wang, B.-G. Diterpenes, Sesquiterpenes, and a C15-Acetogenin from the Marine Red Alga Laurencia mariannensis. J. Nat. Prod. 2007, 70, 1901–1905. [Google Scholar] [CrossRef]
- Kamei, Y.; Sueyoshi, M.; Hayashi, K.I.; Terada, R.; Nozaki, H. The novel anti-Propionibacterium acnes compound, Sargafuran, found in the marine brown alga Sargassum macrocarpum. J. Antibiot. 2009, 62, 259–263. [Google Scholar] [CrossRef]
- Lane, A.L.; Mular, L.; Drenkard, E.J.; Shearer, T.L.; Engel, S.; Fredericq, S.; Fairchild, C.R.; Prudhomme, J.; Le Roch, K.; Hay, M.E.; et al. Ecological leads for natural product discovery: Novel sesquiterpene hydroquinones from the red macroalga Peyssonnelia sp. Tetrahedron 2010, 66, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Vairappan, C.S.; Daitoh, M.; Suzuki, M.; Abe, T.; Masuda, M. Antibacterial halogenated metabolites from the Malaysian Laurencia species. Phytochemistry 2001, 58, 291–297. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- van Belkum, A.; Burnham, C.-A.D.; Rossen, J.W.A.; Mallard, F.; Rochas, O.; Dunne, W.M. Innovative and rapid antimicrobial susceptibility testing systems. Nat. Rev. Microbiol. 2020, 18, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Reller, L.B.; Weinstein, M.; Jorgensen, J.H.; Ferraro, M.J. Antimicrobial Susceptibility Testing: A Review of General Principles and Contemporary Practices. Clin. Infect. Dis. 2009, 49, 1749–1755. [Google Scholar]
- Pierre, G.; Sopena, V.; Juin, C.; Mastouri, A.; Graber, M.; Maugard, T. Antibacterial activity of a sulfated galactan extracted from the marine alga Chaetomorpha aerea against Staphylococcus aureus. Biotechnol. Bioprocess Eng. 2011, 16, 937–945. [Google Scholar] [CrossRef]
- Trigui, M.; Gasmi, L.; Zouari, I.; Tounsi, S. Seasonal variation in phenolic composition, antibacterial and antioxidant activities of Ulva rigida (Chlorophyta) and assessment of antiacetylcholinesterase potential. J. Appl. Phycol. 2013, 25, 319–328. [Google Scholar] [CrossRef]
- Vijayabaskar, P.; Shiyamala, V. Antibacterial Activities of Brown Marine Algae (Sargassum wightii and Turbinaria ornata) from the Gulf of Mannar Biosphere Reserve. Adv. Biol. Res. 2011, 5, 99–102. [Google Scholar]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazes, A.; Silkina, A.; Defer, D.; Bernède-Bauduin, C.; Quéméner, E.; Braud, J.-P.; Bourgougnon, N. Active substances from Ceramium botryocarpum used as antifouling products in aquaculture. Aquaculture 2006, 258, 664–674. [Google Scholar] [CrossRef]
- Boisvert, C.; Beaulieu, L.; Bonnet, C.; Pelletier, É. Assessment of the antioxidant and antibacterial activities of three species of edible seaweeds. J. Food Biochem. 2015, 39, 377–387. [Google Scholar] [CrossRef]
- Hellio, C.; De La Broise, D.; Dufossé, L.; Le Gal, Y.; Bourgougnon, N. Inhibition of marine bacteria by extracts of macroalgae: Potential use for environmentally friendly antifouling paints. Mar. Environ. Res. 2001, 52, 231–247. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinteus, S.; Alves, C.; Monteiro, H.; Araújo, E.; Horta, A.; Pedrosa, R. Asparagopsis armata and Sphaerococcus coronopifolius as a natural source of antimicrobial compounds. World J. Microbiol. Biotechnol. 2015, 31, 445–451. [Google Scholar] [CrossRef]
- Dubber, D.; Harder, T. Extracts of Ceramium rubrum, Mastocarpus stellatus and Laminaria digitata inhibit growth of marine and fish pathogenic bacteria at ecologically realistic concentrations. Aquaculture 2008, 274, 196–200. [Google Scholar] [CrossRef]
- Li, L.; Shi, Y.; Su, G.; Le, G. Selectivity for and destruction of Salmonella typhimurium via a membrane damage mechanism of a cell-penetrating peptide ppTG20 analogue. Int. J. Antimicrob. Agents 2012, 40, 337–343. [Google Scholar] [CrossRef]
- Shaheen, B.W.; Wang, C.; Johnson, C.M.; Kaltenboeck, B.; Boothe, D.M. Detection of fluoroquinolone resistance level in clinical canine and feline Escherichia coli pathogens using rapid real-time PCR assay. Vet. Microbiol. 2009, 139, 379–385. [Google Scholar] [CrossRef]
- Volokhov, D.; Chizhikov, V.; Chumakov, K.; Rasooly, A. Microarray analysis of erythromycin resistance determinants. J. Appl. Microbiol. 2003, 95, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-X.; Zhang, Z.-W.; Wang, C.; Yang, H.-W.; Jiang, D.; Zhang, Q.; Mitchelson, K.; Cheng, J. Use of a DNA Microarray for Simultaneous Detection of Antibiotic Resistance Genes among Staphylococcal Clinical Isolates. J. Clin. Microbiol. 2007, 45, 3514–3521. [Google Scholar] [CrossRef] [Green Version]
- Shendure, J.; Balasubramanian, S.; Church, G.M.; Gilbert, W.; Rogers, J.; Schloss, J.A.; Waterston, R.H. DNA sequencing at 40: Past, present and future. Nature 2017, 550, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Sherry, N.L.; Porter, J.L.; Seemann, T.; Watkins, A.; Stinear, T.P.; Howden, B.P. Outbreak investigation using high-throughput genome sequencing within a diagnostic microbiology laboratory. J. Clin. Microbiol. 2013, 51, 1396–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, W.; Westblade, L.; Ford, B. Next-generation and whole-genome sequencing in the diagnostic clinical microbiology laboratory. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1719–1726. [Google Scholar] [CrossRef]
- Capilla, J.; Clemons, K.V.; Stevens, D.A. Animal models: An important tool in mycology. Med. Mycol. 2007, 45, 657–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlemmer, K.B.; Jesus, F.P.K.; Loreto, É.S.; Tondolo, J.S.M.; Ledur, P.C.; Dallabrida, A.; da Silva, T.M.; Kommers, G.D.; Alves, S.H.; Santurio, J.M. An experimental murine model of otitis and dermatitis caused by Malassezia pachydermatis. Mycoses 2018, 61, 954–958. [Google Scholar] [CrossRef]
- Sparber, F.; De Gregorio, C.; Steckholzer, S.; Ferreira, F.M.; Dolowschiak, T.; Ruchti, F.; Kirchner, F.R.; Mertens, S.; Prinz, I.; Joller, N.; et al. The Skin Commensal Yeast Malassezia Triggers a Type 17 Response that Coordinates Anti-fungal Immunity and Exacerbates Skin Inflammation. Cell Host Microbe 2019, 25, 389–403.e6. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, C.; Matty, M.A.; Jurczyszak, D.; Gabor, K.A.; Millard, P.J.; Tobin, D.M.; Kim, C.H. Chapter 4—Infectious disease models in zebrafish. In Methods in Cell Biology; Detrich, H.W., Westerfield, M., Zon, L.I., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 138, pp. 101–136. [Google Scholar]
- Vatsos, I.N.; Rebours, C. Seaweed extracts as antimicrobial agents in aquaculture. J. Appl. Phycol. 2015, 27, 2017–2035. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Tawwab, M.; Ahmad, M.H. Live Spirulina (Arthrospira platensis) as a growth and immunity promoter for Nile tilapia, Oreochromis niloticus (L.), challenged with pathogenic Aeromonas hydrophila. Aquac. Res. 2009, 40, 1037–1046. [Google Scholar] [CrossRef]
- Fadl, S.E.; ElGohary, M.S.; Elsadany, A.Y.; Gad, D.M.; Hanaa, F.F.; El-Habashi, N.M. Contribution of microalgae-enriched fodder for the Nile tilapia to growth and resistance to infection with Aeromonas hydrophila. Algal Res. 2017, 27, 82–88. [Google Scholar] [CrossRef]
- Rajendran, P.; Subramani, P.A.; Michael, D. Polysaccharides from marine macroalga, Padina gymnospora improve the nonspecific and specific immune responses of Cyprinus carpio and protect it from different pathogens. Fish. Shellfish Immunol. 2016, 58, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Yengkhom, O.; Shalini, K.S.; Subramani, P.A.; Michael, R.D. Stimulation of non-specific immunity, gene expression, and disease resistance in Nile Tilapia, Oreochromis niloticus (Linnaeus, 1758), by the methanolic extract of the marine macroalga, Caulerpa scalpelliformis. Vet. World 2019, 12, 271–276. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Vidhya Hindu, S.; Vijayakumar, S.; Mukherjee, A.; Chandrasekaran, N.; Thomas, J. Differential solvent extraction of two seaweeds and their efficacy in controlling Aeromonas salmonicida infection in Oreochromis mossambicus: A novel therapeutic approach. Aquaculture 2015, 443, 56–64. [Google Scholar] [CrossRef]
- Manilal, A.; Selvin, J.; George, S. In vivo therapeutic potentiality of red seaweed, Asparagopsis (Bonnemaisoniales, Rhodophyta) in the treatment of Vibriosis in Penaeus monodon Fabricius. Saudi J. Biol. Sci. 2012, 19, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Thanigaivel, S.; Vijayakumar, S.; Mukherjee, A.; Chandrasekaran, N.; Thomas, J. Antioxidant and antibacterial activity of Chaetomorpha antennina against shrimp pathogen Vibrio parahaemolyticus. Aquaculture 2014, 433, 467–475. [Google Scholar] [CrossRef]
- Holdt, S.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Gómez, I.; Huovinen, P. Brown Algal Phlorotannins: An Overview of Their Functional Roles. In Antarctic Seaweeds; Springer: Berlin/Heidelberg, Germany, 2020; pp. 365–388. [Google Scholar]
- Sudatti, D.B.; Fujii, M.T.; Rodrigues, S.V.; Turra, A.; Pereira, R.C. Prompt induction of chemical defenses in the red seaweed Laurencia dendroidea: The role of herbivory and epibiosis. J. Sea Res. 2018, 138, 48–55. [Google Scholar] [CrossRef]
- Pereira, R.C.; da Silva Costa, E.; Sudatti, D.B.; da Gama, B.A.P. Inducible defenses against herbivory and fouling in seaweeds. J. Sea Res. 2017, 122, 25–33. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Rajauria, G.; O’Doherty, J.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietro, A.S. Biochemical and Photosynthetic Aspects of Energy Production; Elsevier: London, UK, 1980. [Google Scholar]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-S.; Kang, M.-S.; Hwang, H.-J.; Eom, S.-H.; Yang, J.-Y.; Lee, M.-S.; Lee, W.-J.; Jeon, Y.-J.; Choi, J.-S.; Kim, Y.-M. Synergistic effect between dieckol from Ecklonia stolonifera and β-lactams against methicillin-resistant Staphylococcus aureus. Biotechnol. Bioprocess Eng. 2008, 13, 758–764. [Google Scholar] [CrossRef]
- Choi, J.-G.; Kang, O.-H.; Brice, O.-O.; Lee, Y.-S.; Chae, H.-S.; Oh, Y.-C.; Sohn, D.-H.; Park, H.; Choi, H.-G.; Kim, S.-G. Antibacterial activity of Ecklonia cava against methicillin-resistant Staphylococcus aureus and Salmonella spp. Foodborne Pathog. Dis. 2010, 7, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Eom, S.-H.; Lee, E.-H.; Jung, Y.-J.; Kim, H.-J.; Jo, M.-R.; Son, K.-T.; Lee, H.-J.; Kim, J.H.; Lee, M.-S. In vitro antibacterial and synergistic effect of phlorotannins isolated from edible brown seaweed Eisenia bicyclis against acne-related bacteria. Algae 2014, 29, 47. [Google Scholar] [CrossRef]
- Oh, K.-B.; Lee, J.H.; Chung, S.-C.; Shin, J.; Shin, H.J.; Kim, H.-K.; Lee, H.-S. Antimicrobial activities of the bromophenols from the red alga Odonthalia corymbifera and some synthetic derivatives. Bioorg. Med. Chem. Lett. 2008, 18, 104–108. [Google Scholar] [CrossRef]
- Popplewell, W.L.; Northcote, P.T. Colensolide A: A new nitrogenous bromophenol from the New Zealand marine red alga Osmundaria colensoi. Tetrahedron Lett. 2009, 50, 6814–6817. [Google Scholar] [CrossRef]
- Paul, N.A.; de Nys, R.; Steinberg, P.D. Chemical defence against bacteria in the red alga asparagopsis armata: Linking structure with function. Mar. Ecol. Prog. Ser. 2006, 306, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, D.; Alves, C.; Horta, A.; Pinteus, S.; Silva, J.; Culioli, G.; Thomas, O.P.; Pedrosa, R. Antitumor and antimicrobial potential of bromoditerpenes isolated from the red alga, Sphaerococcus coronopifolius. Mar. Drugs 2015, 13, 713–726. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.-E.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Park, N.-H.; Choi, J.-S.; Hwang, S.-Y.; Kim, Y.-C.; Hong, Y.-K.; Cho, K.K.; Choi, I.S. Antimicrobial activities of stearidonic and gamma-linolenic acids from the green seaweed Enteromorpha linza against several oral pathogenic bacteria. Bot. Stud. 2013, 54, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Cecere, E.; Petrocelli, A. Biotechnological potential of the seaweed Cladophora rupestris (Chlorophyta, Cladophorales) lipidic extract. New Biotechnol. 2014, 31, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Kim, S.H.; Baek, K.H. Antioxidant and Free Radical-Scavenging Potential of Essential Oil from E nteromorpha linza L. Prepared by Microwave-Assisted Hydrodistillation. J. Food Biochem. 2015, 39, 80–90. [Google Scholar] [CrossRef]
- Patra, J.; Das, G.; Baek, K.-H. Chemical composition and antioxidant and antibacterial activities of an essential oil extracted from an edible seaweed, Laminaria japonica L. Molecules 2015, 20, 12093–12113. [Google Scholar] [CrossRef] [Green Version]
- Patra, J.K.; Lee, S.W.; Park, J.G.; Baek, K.H. Antioxidant and antibacterial properties of essential oil extracted from an edible seaweed Undaria pinnatifida. J. Food Biochem. 2017, 41, e12278. [Google Scholar] [CrossRef]
- Vijayabaskar, P.; Vaseela, N.; Thirumaran, G. Potential antibacterial and antioxidant properties of a sulfated polysaccharide from the brown marine algae Sargassum swartzii. Chin. J. Nat. Med. 2012, 10, 421–428. [Google Scholar] [CrossRef]
- Marudhupandi, T.; Kumar, T.T.A. Antibacterial effect of fucoidan from Sargassum wightii against the chosen human bacterial pathogens. Int. Curr. Pharm. J. 2013, 2, 156–158. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.U.; Donnell, C.P.; Rai, D.K.; Hossain, M.B.; Burgess, C.M.; Walsh, D.; Tiwari, B.K. Laminarin from Irish Brown Seaweeds Ascophyllum nodosum and Laminaria hyperborea: Ultrasound Assisted Extraction, Characterization and Bioactivity. Mar. Drugs 2015, 13, 4270–4280. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Y.; Cao, M.-J.; Liu, G.-M.; Chen, Q.; Sun, L.; Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohydr. Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. In vitro antioxidant and antibacterial activity of sulfated polysaccharides isolated from Spatoglossum asperum. Carbohydr. Polym. 2017, 170, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.-Y.; Jung, M.-J.; Jeong, I.-H.; Yamazaki, K.; Kawai, Y.; Kim, B.-M. Antimicrobial and Antibiofilm Activities of Sulfated Polysaccharides from Marine Algae against Dental Plaque Bacteria. Mar. Drugs 2018, 16, 301. [Google Scholar] [CrossRef] [Green Version]
- Palanisamy, S.; Vinosha, M.; Rajasekar, P.; Anjali, R.; Sathiyaraj, G.; Marudhupandi, T.; Selvam, S.; Prabhu, N.M.; You, S. Antibacterial efficacy of a fucoidan fraction (Fu-F2) extracted from Sargassum polycystum. Int. J. Biol. Macromol. 2019, 125, 485–495. [Google Scholar] [CrossRef]
- Kavita, K.; Singh, V.K.; Jha, B. 24-Branched Δ5 sterols from Laurencia papillosa red seaweed with antibacterial activity against human pathogenic bacteria. Microbiol. Res. 2014, 169, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Thilakan, B.; Chakraborty, R.D.; Raola, V.K.; Joy, M. O-heterocyclic derivatives with antibacterial properties from marine bacterium Bacillus subtilis associated with seaweed, Sargassum myriocystum. Appl. Microbiol. Biotechnol. 2017, 101, 569–583. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-X.; Wijesekara, I.; Li, Y.; Kim, S.-K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65 Pt 1, 535–543. [Google Scholar] [CrossRef]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can Phlorotannins Purified Extracts Constitute a Novel Pharmacological Alternative for Microbial Infections with Associated Inflammatory Conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Kim, A.-R.; Shin, T.-S.; Lee, M.-S.; Park, J.-Y.; Park, K.-E.; Yoon, N.-Y.; Kim, J.-S.; Choi, J.-S.; Jang, B.-C.; Byun, D.-S. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lee, S.-H.; Le, Q.-T.; Kim, M.-M.; Kim, S.-K. Anti-allergic effects of phlorotannins on histamine release via binding inhibition between IgE and FcεRI. J. Agric. Food Chem. 2008, 56, 12073–12080. [Google Scholar] [CrossRef]
- Shibata, T.; Ishimaru, K.; Kawaguchi, S.; Yoshikawa, H.; Hama, Y. Antioxidant Activities of Phlorotannins Isolated from Japanese Laminariaceae. In Proceedings of the Nineteenth International Seaweed Symposium, Kobe, Japan, 26–31 March 2007; Springer: Amsterdam, The Netherlands, 2007; pp. 255–261. [Google Scholar]
- Hierholtzer, A.; Chatellard, L.; Kierans, M.; Akunna, J.C.; Collier, P.J. The impact and mode of action of phenolic compounds extracted from brown seaweed on mixed anaerobic microbial cultures. J. Appl. Microbiol. 2013, 114, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Shannon, E.; Abu-Ghannam, N. Antibacterial Derivatives of Marine Algae: An Overview of Pharmacological Mechanisms and Applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Bach, S.J.; McAllister, T.A. Sensitivity of Escherichia coli to Seaweed (Ascophyllum nodosum) Phlorotannins and Terrestrial Tannins. Asian-Australas J. Anim. Sci. 2009, 22, 238–245. [Google Scholar] [CrossRef]
- Eom, S.-H.; Lee, D.-S.; Jung, Y.-J.; Park, J.-H.; Choi, J.-I.; Yim, M.-J.; Jeon, J.-M.; Kim, H.-W.; Son, K.-T.; Je, J.-Y. The mechanism of antibacterial activity of phlorofucofuroeckol-A against methicillin-resistant Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2014, 98, 9795–9804. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, Q.; Xu, C.; Yu, J.; Zhao, L.; Guo, Q. Damage to the Membrane Permeability and Cell Death of Vibrio parahaemolyticus Caused by Phlorotannins with Low Molecular Weight from Sargassum thunbergii. J. Aquat. Food Prod. Technol. 2016, 25, 323–333. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in Marine Algae and Their Bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öztaskın, N.; Taslimi, P.; Maraş, A.; Gülcin, İ.; Göksu, S. Novel antioxidant bromophenols with acetylcholinesterase, butyrylcholinesterase and carbonic anhydrase inhibitory actions. Bioorg. Chem. 2017, 74, 104–114. [Google Scholar] [CrossRef]
- Xu, N.; Fan, X.; Yan, X.; Li, X.; Niu, R.; Tseng, C.K. Antibacterial bromophenols from the marine red alga Rhodomela confervoides. Phytochemistry 2003, 62, 1221–1224. [Google Scholar] [CrossRef]
- Barreto, M.; Meyer, J. Isolation and antimicrobial activity of a lanosol derivative from Osmundaria serrata (Rhodophyta) and a visual exploration of its biofilm covering. S. Afr. J. Bot. 2006, 72, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-S.; Lee, T.-H.; Lee, J.H.; Chae, C.-S.; Chung, S.-C.; Shin, D.-S.; Shin, J.; Oh, K.-B. Inhibition of the pathogenicity of Magnaporthe grisea by bromophenols, isocitrate lyase inhibitors, from the red alga Odonthalia corymbifera. J. Agric. Food Chem. 2007, 55, 6923–6928. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, S.R.; Oh, M.-J.; Jung, S.-J.; Kang, S.Y. In vitro antiviral activity of red alga, Polysiphonia morrowii extract and its bromophenols against fish pathogenic infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus. J. Microbiol. 2011, 49, 102–106. [Google Scholar] [CrossRef]
- Park, H.-J.; Kurokawa, M.; Shiraki, K.; Nakamura, N.; Choi, J.-S.; Hattori, M. Antiviral activity of the marine alga Symphyocladia latiuscula against herpes simplex virus (HSV-1) in vitro and its therapeutic efficacy against HSV-1 infection in mice. Biol. Pharm. Bull. 2005, 28, 2258–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleurence, J.; Morançais, M.; Dumay, J. Seaweed proteins. In Proteins in Food Processing, 2nd ed.; Yada, R.Y., Ed.; Woodhead Publishing: Cambridge, UK, 2018; pp. 245–262. [Google Scholar]
- Galland-Irmouli, A.-V.; Fleurence, J.; Lamghari, R.; Luçon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.-P.; Villaume, C.; Guéant, J.-L. Nutritional value of proteins from edible seaweed Palmaria palmata (Dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Lafarga, T.; Acién-Fernández, F.G.; Garcia-Vaquero, M. Bioactive peptides and carbohydrates from seaweed for food applications: Natural occurrence, isolation, purification, and identification. Algal Res. 2020, 48, 101909. [Google Scholar] [CrossRef]
- Singh, R.S.; Walia, A.K. Lectins from red algae and their biomedical potential. J. Appl. Phycol. 2018, 30, 1833–1858. [Google Scholar] [CrossRef] [Green Version]
- The Antimicrobial Database. Available online: http://aps.unmc.edu/AP/ (accessed on 18 July 2020).
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.S.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-Spectrum In Vitro Activity and In Vivo Efficacy of the Antiviral Protein Griffithsin against Emerging Viruses of the Family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Séron, K.; Labitt, R.N.; Danneels, A.; Palmer, K.E.; Whittaker, G.R.; Dubuisson, J.; Belouzard, S. Middle East respiratory syndrome coronavirus infection is inhibited by griffithsin. Antivir. Res. 2016, 133, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Simplicien, M.; Benoist, H.; Van Damme, E.J.M.; Rougé, P. Mannose-Specific Lectins from Marine Algae: Diverse Structural Scaffolds Associated to Common Virucidal and Anti-Cancer Properties. Mar. Drugs 2019, 17, 440. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, T. Physiology and Biochemistry of Algae; Lewin, R.A., Ed.; Academic Press: New York, NY, USA, 1962; pp. 467–473. [Google Scholar]
- Shoubaky, G.; Salem, E. Active ingredients fatty acids as antibacterial agent from the brown algae Padina pavonica and Hormophysa triquetra. J. Coast. Life Med. 2014, 2, 535–542. [Google Scholar]
- Da Costa, E.; Melo, T.; Moreira, A.S.; Bernardo, C.; Helguero, L.; Ferreira, I.; Cruz, M.T.; Rego, A.M.; Domingues, P.; Calado, R. Valorization of lipids from Gracilaria sp. through lipidomics and decoding of antiproliferative and anti-inflammatory activity. Mar. Drugs 2017, 15, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as sources of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Stiger-Pouvreau, V.; Bourgougnon, N.; Deslandes, E. Carbohydrates from seaweeds. In Seaweed in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2016; pp. 223–274. [Google Scholar]
- de Jesus Raposo, M.F.; De Morais, A.M.B.; De Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Yang, F.; Lu, J. Fucoidan Extracted from Undaria pinnatifida: Source for Nutraceuticals/Functional Foods. Mar. Drugs 2018, 16, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zargarzadeh, M.; Amaral, A.J.R.; Custódio, C.A.; Mano, J.F. Biomedical applications of laminarin. Carbohydr. Polym. 2020, 232, 115774. [Google Scholar] [CrossRef]
- He, F.; Yang, Y.; Yang, G.; Yu, L. Studies on antibacterial activity and antibacterial mechanism of a novel polysaccharide from Streptomyces virginia H03. Food Control 2010, 21, 1257–1262. [Google Scholar] [CrossRef]
- Kim, S.-K.; Van Ta, Q. Chapter 14—Potential Beneficial Effects of Marine Algal Sterols on Human Health. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 64, pp. 191–198. [Google Scholar]
- Ikekawa, N.; Morisaki, N.; Tsuda, K.; Yoshida, T. Sterol compositions in some green algae and brown algae. Steroids 1968, 12, 41–48. [Google Scholar] [CrossRef]
- Prakash, S.; Sasikala, S.; Aldous, V.H.J. Isolation and identification of MDR–Mycobacterium tuberculosis and screening of partially characterised antimycobacterial compounds from chosen marine micro algae. Asian Pac. J. Trop. Med. 2010, 3, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Wächter, G.A.; Franzblau, S.G.; Montenegro, G.; Hoffmann, J.J.; Maiese, W.M.; Timmermann, B.N. Inhibition of Mycobacterium t uberculosis Growth by Saringosterol from Lessonia n igrescens. J. Nat. Prod. 2001, 64, 1463–1464. [Google Scholar] [CrossRef]
- Kawakami, L.M.; Yoon, B.K.; Jackman, J.A.; Knoll, W.; Weiss, P.S.; Cho, N.-J. Understanding how sterols regulate membrane remodeling in supported lipid bilayers. Langmuir 2017, 33, 14756–14765. [Google Scholar] [CrossRef] [PubMed]
- Tidgewell, K.; Clark, B.R.; Gerwick, W.H. The Natural Products Chemistry of Cyanobacteria. In Comprehensive Natural Products II; Liu, H.-W., Mander, L., Eds.; Elsevier: Oxford, UK, 2010; pp. 141–188. [Google Scholar]
- Gomez, L.P.; Alvarez, C.; Zhao, M.; Tiwari, U.; Curtin, J.; Garcia-Vaquero, M.; Tiwari, B.K. Innovative processing strategies and technologies to obtain hydrocolloids from macroalgae for food applications. Carbohydr. Polym. 2020, 248, 116784. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, O.G.; Dawczynski, C.; Duelund, L.; Jahreis, G.; Vetter, W.; Schröder, M. On the human consumption of the red seaweed dulse (Palmaria palmata (L.) Weber & Mohr). J. Appl. Phycol. 2013, 25, 1777–1791. [Google Scholar]
- Kadam, S.; Prabhasankar, P. Marine foods as functional ingredients in bakery and pasta products. Food Res. Int. 2010, 43, 1975–1980. [Google Scholar] [CrossRef]
- Cofrades, S.; Benedí, J.; Garcimartin, A.; Sánchez-Muniz, F.; Jimenez-Colmenero, F. A comprehensive approach to formulation of seaweed-enriched meat products: From technological development to assessment of healthy properties. Food Res. Int. 2017, 99, 1084–1094. [Google Scholar] [CrossRef]
- Hall, R.S.; Baxter, A.L.; Fryirs, C.; Johnson, S.K. Liking of health-functional foods containing lupin kernel fibre following repeated consumption in a dietary intervention setting. Appetite 2010, 55, 232–237. [Google Scholar] [CrossRef] [Green Version]
- López-López, I.; Cofrades, S.; Jiménez-Colmenero, F. Low-fat frankfurters enriched with n−3 PUFA and edible seaweed: Effects of olive oil and chilled storage on physicochemical, sensory and microbial characteristics. Meat Sci. 2009, 83, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-López, I.; Cofrades, S.; Yakan, A.; Solas, M.T.; Jiménez-Colmenero, F. Frozen storage characteristics of low-salt and low-fat beef patties as affected by Wakame addition and replacing pork backfat with olive oil-in-water emulsion. Food Res. Int. 2010, 43, 1244–1254. [Google Scholar] [CrossRef]
- Cofrades, S.; López-López, I.; Ruiz-Capillas, C.; Triki, M.; Jiménez-Colmenero, F. Quality characteristics of low-salt restructured poultry with microbial transglutaminase and seaweed. Meat Sci. 2011, 87, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Abu-Ghannam, N. Enhancement of the phytochemical and fibre content of beef patties with Himanthalia elongata seaweed. Int. J. Food Sci. Technol. 2013, 48, 2239–2249. [Google Scholar]
- Moroney, N.C.; O’Grady, M.N.; O’Doherty, J.V.; Kerry, J.P. Effect of a brown seaweed (Laminaria digitata) extract containing laminarin and fucoidan on the quality and shelf-life of fresh and cooked minced pork patties. Meat Sci. 2013, 94, 304–311. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Sineiro, J.; Amado, I.R.; Franco, D. Influence of natural extracts on the shelf life of modified atmosphere-packaged pork patties. Meat Sci. 2014, 96, 526–534. [Google Scholar] [CrossRef]
- Vilar, E.G.; Ouyang, H.; O’Sullivan, M.G.; Kerry, J.P.; Hamill, R.M.; O’Grady, M.N.; Mohammed, H.O.; Kilcawley, K.N. Effect of salt reduction and inclusion of 1% edible seaweeds on the chemical, sensory and volatile component profile of reformulated frankfurters. Meat Sci. 2020, 161, 108001. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Waldron, D.S.; Smyth, T.J.; O’Brien, N.M.; Kerry, J.P. An examination of the potential of seaweed extracts as functional ingredients in milk. Int. J. Dairy Technol. 2014, 67, 182–193. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; O’Grady, M.N.; O’Callaghan, Y.C.; Smyth, T.J.; O’Brien, N.M.; Kerry, J.P. Seaweed extracts as potential functional ingredients in yogurt. Innov. Food Sci. Emerg. Technol. 2016, 37, 293–299. [Google Scholar] [CrossRef]
- del Olmo, A.; Picon, A.; Nuñez, M. Cheese supplementation with five species of edible seaweeds: Effect on microbiota, antioxidant activity, colour, texture and sensory characteristics. Int. Dairy J. 2018, 84, 36–45. [Google Scholar] [CrossRef]
- Lee, C.-J.; Choi, J.-S.; Song, E.-J.; Lee, S.-Y.; Kim, K.-B.-W.-R.; Kim, S.-J.; Yoon, S.-Y.; Lee, S.-J.; Park, N.-B.; Jung, J.-Y. Effect of Myagropsis myagroides extracts on shelf-life and quality of bread. Korean J. Food Sci. Technol. 2010, 42, 50–55. [Google Scholar]
- Jung, K.-I.; Choi, Y.-J.; Cho, E.-K. Effect of Ecklonia cava hot water extracts on shelf-life and quality of muffin. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1672–1677. [Google Scholar] [CrossRef]
- Kim, M.-J.; Kim, K.-B.-W.-R.; Lee, C.-J.; Kwak, J.-H.; Kim, D.-H.; SunWoo, C.; Jung, S.-A.; Kang, J.-Y.; Kim, H.-J.; Choi, J.-S. Effect of Sargassum sagamianum extract on shelf-life and improved quality of morning bread. Korean J. Food Sci. Technol. 2011, 43, 723–728. [Google Scholar] [CrossRef]
- Poveda-Castillo, G.D.C.; Rodrigo, D.; Martínez, A.; Pina-Pérez, M.C. Bioactivity of Fucoidan as an Antimicrobial Agent in a New Functional Beverage. Beverages 2018, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Neetoo, H.; Ye, M.; Chen, H. Bioactive alginate coatings to control Listeria monocytogenes on cold-smoked salmon slices and fillets. Int. J. Food Microbiol. 2010, 136, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Albertos, I.; Martin-Diana, A.B.; Burón, M.; Rico, D. Development of functional bio-based seaweed (Himanthalia elongata and Palmaria palmata) edible films for extending the shelflife of fresh fish burgers. Food Packag. Shelf Life 2019, 22, 100382. [Google Scholar] [CrossRef]
- Barberi, O.N.; Byron, C.J.; Burkholder, K.M.; Gelais, A.T.S.; Williams, A.K. Assessment of bacterial pathogens on edible macroalgae in coastal waters. J. Appl. Phycol. 2020, 32, 683–696. [Google Scholar] [CrossRef] [Green Version]
- Banach, J.L.; Hoek-van den Hil, E.F.; van der Fels-Klerx, H.J. Food safety hazards in the european seaweed chain. Compr. Rev. Food Sci. Food Saf. 2020, 19, 332–364. [Google Scholar] [CrossRef]
- FSAI. Safety Considerations of Seaweed and Seaweed-Derived Foods Available on the Irish Market; FSAI: Dublin, Ireland, 2020. [Google Scholar]
- Commission regulation (EC) 1881/2006 of 19 december 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 49, 5–24.
- Directive 2002/32/EC of the European Parliament and of the Council of 7 may 2002 on undesirable substances in animal feed. Off. J. Eur. Union 2002, 45, 10–21.
- Recommendation (EU) 2018/464 of 19 march 2018 on the monitoring of metals and iodine in seaweed, halophytes and products based on seaweed. Off. J. Eur. Union 2018, 61, 16–18.
- Hanaoka, K.I.; Yosida, K.; Tamano, M.; Kuroiwa, T.; Kaise, T.; Maeda, S. Arsenic in the prepared edible brown alga hijiki, hizikia fusiforme. Appl. Organomet. Chem. 2001, 15, 561–565. [Google Scholar] [CrossRef]
- Ichikawa, S.; Nozawa, S.; Hanaoka, K.I.; Kaise, T. Ingestion and excretion of arsenic compounds present in edible brown algae, hijikia fusiforme, by mice. Food Chem. Toxicol. 2010, 48, 465–469. [Google Scholar] [CrossRef]
- Wright, G.D. Opportunities for natural products in 21 st century antibiotic discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Compounds | Source | Activity Against Bacterial Pathogens | References |
|---|---|---|---|
| Phlorotannins | |||
| Dieckol | Ecklonia stolonifera | Methicillin-resistant Staphylcoccus aureus (MRSA) (MIC 64 μg/mL) and methicillin-susceptible S. aureus (MSSA) (MIC 32 μg/mL). | Lee et al. [58] |
| Eckol | Ecklonia cava | Inhibition of 17 strains of S. aureus MRSA (MIC 125-250 μg/mL) and 16 strains of Salmonella spp. (MIC 250 μg/mL). | Choi et al. [59] |
| Fucofuroeckol-A | Eisenia bicyclis | Propionibacterium acnes (KCTC 3314) (MIC 32 µg/mL), P. acnes isolate 2875 (MIC 32 µg/mL), P. acnes isolate 2876 (MIC 64 µg/mL), S aureus (KCTC 1927) (MIC 128 µg/mL) and Staphylococcus epidermidis (KCTC 1370) (MIC 64 µg/mL). | Lee et al. [60] |
| Bromophenols | |||
| 2,20,3,30-tetrabromo-4,40,5,50- tetrahydroxydiphenylmethane | Odonthalia corymbifera | S. aureus ATCC6538p (MIC 25 μg/mL), Bacillus subtilis ATCC 6633 (MIC 25 μg/mL), Micrococcus luteus IFC 12708 (MIC 25 μg/mL), Proteus vulgaris ATCC3851 (MIC 50 μg/mL), S. Typhimurium ATCC 14028 (MIC 50 μg/mL) and E. coli ATCC 25922 (MIC > 100 μg/mL). | Oh et al. [61] |
| 2,20,3-tribromo-30,4,40,5-tetrahydroxy-60 -hydroxymethyl diphenylmethane | O. corymbifera | S. aureus ATCC6538p (MIC 50 μg/mL), B. subtilis ATCC 6633 (MIC 100 μg/mL), M. luteus IFC 12708 (MIC 50 μg/mL), P. vulgaris ATCC3851 (MIC 100 μg/mL), S. Typhimurium ATCC 14028 (MIC 100 μg/mL) and E. coli ATCC 25922 (MIC > 100 μg/mL). | Oh et al. [61] |
| 3-bromo-4-(2,3-dibromo-4,5-dihydroxybenzyl)-5- methoxymethylpyrocatechol | O. corymbifera | S. aureus ATCC6538p (MIC 25 μg/mL), B. subtilis ATCC 6633 (MIC 25 μg/mL), M. luteus IFC 12708 (MIC 25 μg/mL), P. vulgaris ATCC3851 (MIC 25 μg/mL), S. Typhimurium ATCC 14028 (MIC 25 μg/mL) and E. coli ATCC 25922 (MIC > 100 μg/mL). | Oh et al. [61] |
| Lanosol methyl ether | Osmundaria colensoi | Mycobacterium smegmatis (IC50 7.8 μM). | Popplewell and Northcote [62] |
| Lanosol butanone | O. colensoi | M. smegmatis (IC50 26.2 μM). | Popplewell and Northcote [62] |
| Rhodomelol | O. colensoi | M. smegmatis (IC50 28.1 μM). | Popplewell and Northcote [62] |
| Other halogenated compounds | |||
| Bromoform and dibromoacetic acid | Asparagopsis armata | Both methanol and dichloromethane extracts from A. armata were active against 6 strains of bacteria (Vibrio harveyii, Vibrio alginolyticus, Pseudomonas aeruginosa, S. aureus, S. epidermis and E. coli). In general, E. coli and P. aeruginosa were most susceptible to the dichloromethane extracts. | Paul et al. [63] |
| Sphaerodactylomelol | Sphaerococcus coronopifolius | S. aureus (MIC 96 µM). | Rodrigues et al. [64] |
| Bromosphaerol | S. coronopifolius | S. aureus (MIC 22 µM). | Rodrigues et al. [64] |
| 12R-hydroxybromosphaerol | S. coronopifolius | S. aureus (MIC 6 µM). | Rodrigues et al. [64] |
| Proteins and peptides | |||
| Protein hydrolysate >10 kDa containing 9 peptides (TITLDVEPSDTIDGVK, ISGLIYEETR, MALSSLPR, ILVLQSNQIR, ISAILPSR, IGNGGELPR, LPDAALNR, EAESSLTGGNGCAK and QVHPDTGISK) | Saccharina longicruris | Decrease the maximum specific growth rate of S. aureus at concentrations ranging from 0.31 to 2.5 mg/mL. | Beaulieu et al. [65] |
| Fatty acids | |||
| Stearidonic and gamma-linolenic acids | Enteromorpha linza | Prevotella intermedia (MIC 39.06 μg/mL) and Porphyromonas gingivalis (MIC 9.76 μg/mL). Both compounds were also active against Aggregatibacter actinomycetemcomitans, Candida albicans, Fusobacterium nucleatum subsp. vincentii and Streptococcus mutans. | Park et al. [66] |
| Lipid extract containing palmitic, myristic, oleic, α linolenic, palmitoleic and linoleic acids | Cladophora rupestris | Antibacterial activity against Enterococcus sp., Streptococcus agalactiae and Vibrio cholera non-O1. | Stabili et al. [67] |
| Essential oil composed of acids (54.6%), alkenes (21.1%), alcohols (4.5%), aldehydes (3.7%) and ketones (2.8%) | E. linza | E. coli ATCC 43889 and 43890 (MIC 12.5 mg/mL; MBC 25 mg/mL), S. Typhimurium ATCC 19586 (MIC 25 mg/mL; MBC 25 mg/mL) and S. Typhimurium ATCC 43174 (MIC 12.5 mg/mL; MBC 25 mg/mL). | Patra et al. [68] |
| Essential oil composed of tetradeconoic acid (51.75%), hexadecanoic acid (16.57%), (9Z,12Z)-9,12-Octadecadienoic acid (12.09%) and (9Z)-hexadec-9-enoic acid (9.25%) | Laminaria japonica | S. aureus (11.5 ± 0.58 mm inhibition zone at 25 mg/disc) and Bacillus cereus (10.5 ± 0.57 mm inhibition zone at 25 mg/disc), but no inhibition of E. coli O157:H7. | Patra et al. [69] |
| Essential oil composed of tetradecanoic acid (31.32%) and hexadecanoic acid (22.39%) was present in the highest amount, followed by 3-hexen-1-ol (5.67%), erythritol (4.73%), 4-imidazolidinone (4.40%) and (9Z)-hexadec-9-enoic acid (4.37%) | Undaria Pinnatifida | S. aureus ATCC 12600 (MIC 12.5 mg/mL; MBC 25 mg/mL) and S. Typhimurium ATCC 43174 (MIC 25 mg/mL; MBC 25 mg/mL). | Patra et al. [70] |
| Polysaccharides | |||
| Crude extracts containing fucoidan (sulphate content: 5.3 ± 1.54%) | Sargassum swartzii | 10 µg extract inhibited: S. aureus (9 ± 0.67 mm inhibition), Proteus vulgaris (7 ± 0.72 mm inhibition), E. coli (15 ± 0.28 mm inhibition), B. subtilis (16 ± 0.52 mm inhibition), P. aeruginosa (11 ± 0.48 mm inhibition), Salmonella Typhi (10 ± 0.62 mm inhibition), Shigella flexineri (6 ± 0.78 mm inhibition), Enterococcus faecalis (10 ± 0.68 mm inhibition) and Aeromonas hydrophilla (2 ± 0.32 mm inhibition). | Vijayabaskar et al. [71] |
| Fucoidan (sulphate content: 29.26 ± 0.83%) | Sargassum wightii | P. aeruginosa (MIC 62.5 μg/mL; MBC 150 μg/mL) and E. coli (MIC 125 μg/mL; MBC 250 μg/mL). | Marudhupandi and Kumar [72] |
| Laminarin-rich extracts (no purity reported) | Laminaria hyperborea and Ascophyllum nodosum | E. coli (MIC non detected (nd)-596.8 mg/mL), S. Typhimurium (13.1–33.4 mg/mL) S. aureus (nd-66.8 mg/mL) and L. monocytogenes (nd-66.8 mg/mL). | Kadam et al. [73] |
| Fucoidan | L. japonica | Unprocessed fucoidans did not show obvious antibacterial activity against E. coli and S. aureus even at 10 mg/mL. Depolymerized fucoidans effectively inhibit the proliferation of both bacteria. | Liu et al. [74] |
| Fucoidan (21.35 ± 0.81% sulphate) | Spatoglossum asperum | A. hydrophila (MIC 100 μg/mL). | Palanisamy et al. [75] |
| Fucoidan (sulphate content: 14 ± 2.7%) | Fucus vesiculosus | L. monocytogenes KCTC 13064 (MIC 250 µg/mL), S. aureus KCTC 3881 (MIC 500 µg/mL), E. faecalis KCTC 5289 (MIC 1000 µg/mL), S. mutans KCTC 5458 (MIC 125 µg/mL), S. mutans KCCM 40105 (MIC 250 µg/mL), Streptococcus oralis KCCM 41567 (MIC 500 µg/mL), Streptococcus sobrinus KCTC 5809 (MIC 250 µg/mL), S. sobrinus KCCM 11898 (MIC 250 µg/mL), Streptococcus sanguinis KCTC 5643 (MIC 500 µg/mL), Lactobacillus acidophilus KCTC 3164 (MIC 500 µg/mL) and Streptococcus thermophilus KCTC 3658 (MIC 500 µg/mL). | Jun et al. [76] |
| Fucoidan (sulphate content: 20.41 ± 0.91%) | Sargassum polycystum | S. mutans (MIC 100 μg/mL; MBC 300 μg/mL), P. aeruginosa (MIC 50 μg/mL; MBC 200 μg/mL), S. aureus (MIC 200 μg/mL; MBC 300 μg/mL) and E. coli (MIC 200 μg/mL; MBC 300 μg/mL). | Palanisamy et al. [77] |
| Other compounds | |||
| Sterol (24-propylidene cholest-5-en-3β-ol) | Laurencia papillosa | Antibacterial activity against E. coli, P. aerugenosa, Klebsiella pneumonia and S. flexineri. MIC ranging from 1.2 to 1.7 μg/mL (IC50). | Kavita et al. [78] |
| Polyketide (3-(methoxycarbonyl)-4-(5- (2-ethylbutyl)-5,6-dihydro-3-methyl2H-pyran-2-yl)-butyl benzoate) | Sargassum myriocystum | Vibrio parahemolyticus (zone of inhibition 7 mm at 10 µg/disk), Vibrio vulnificus (zone of inhibition 7 mm at 10 µg/disk) and A. hydrophilla (zone of inhibition 8 mm at 10 µg/disk). | Chakraborty et al. [79] |
| Polyketide (2-(8-butyl-3-ethyl-3,4,4a,5,6, 8ahexahydro-2H-chromen-6-yl)-ethyl benzoate) | S. myriocystum | V. parahemolyticus (zone of inhibition 9 mm at 10 µg/disk), V. vulnificus (zone of inhibition 8 mm at 10 µg/disk) and A. hydrophilla (zone of inhibition 7 mm at 10 µg/disk). | Chakraborty et al. [79] |
| Food Product | Macroalgae or Etract | Microorganisms Tested | Antimicrobial Effect | Effects on the Quality Attributes of Food | References |
|---|---|---|---|---|---|
| Meat products | |||||
| Sausages (frankfurter) | Himanthalia elongata dried. Addition to food: 5%. | Total viable count, lactic acid bacteria and Enterobacteriaceae counts. | Sausages containing seaweed had high total viable count from day 14 of storage, with lactic acid bacteria becoming the predominant microflora. Enterobacteriaceae count was always below 2 log CFU/g. | Seaweed allowed the development of low-fat frankfurters with favorable sensory properties (hard and chewy) with good water and fat binding properties compared to non-supplemented sausages. Macroalgae are useful in the production of products with reduced salt contents. | López-López et al. [129] |
| Frozen beef patties | Undaria pinnatifida dried. Addition to food: 3%. | Viable aerobic microorganism and Enterobacteriaceae counts. | Microbial populations generally increased with the addition of macroalgae, although no significant differences were appreciated in microbial populations with respect to control patties over time. Total viable counts were in the range of 6–6.4 log CFU/g and Enterobacteriaceae did not exceed 4.3 log CFU/g. | Patties with macroalgae had less thawing and cooking losses and were softer compared to control patties, while having a higher mineral content. All the products were considered acceptable by a sensory panel. The addition of macroalgae did not affect the properties of the patties in the course of frozen storage. | López-López et al. [130] |
| Frozen restructured poultry steak | H. elongata dried. Addition to food: 3%. | Total viable count, lactic acid bacteria, Enterobacteriaceae, β-glucuronidase-positive E. coli and Salmonella spp. | Products with macroalgae had higher levels of total viable counts and lactic acid bacteria compared to control. | The incorporation of seaweed caused an increase in purge loss and a reduced cooking loss. All the steaks were judged acceptable by a sensory panel. | Cofrades et al. [131] |
| Frozen beef patties | H. elongata powder. Addition to food: 10–40%. | Total viable counts. | Microbiological counts were low in patties containing macroalgae. No bacterial growth was appreciated after day 30 of storage in patties containing ≥20% of macroalgae. | Patties with seaweed showed reduced cooking losses and were nearly 50% more tender as compared to patties without seaweed. The lipid oxidation, dietary fiber and antioxidant properties of patties were improved by the addition of macroalgae. Sensory analysis (aroma, appearance, texture and taste) indicated that the seaweed patties were accepted by consumers, the patties containing 40% seaweed being preferred overall due to improvements in texture and mouth feel. | Cox and Abu-Ghannam [132] |
| Minced pork patties (fresh and cooked) | Macroalgal extracts (laminarin (9.3%) and fucoidan (7.8%)). Addition to food: 0.01–0.5%. | Total viable counts. | No effect on the microbial population studied. | The addition of macroalgal extracts had no effect on pH, water holding capacity and cook loss of patties. The patties containing 0.01% of macroalgal extract were preferred by the sensory panelists. | Moroney et al. [133] |
| Pork patties | Ulva spp. extract (acidic extract). Addition to food: 1000 mg extract per kg. | Total viable counts, lactic acid bacteria and Pseudomonas. | Macroalgal extracts did not affect the microbial growth in pork patties compared to control. Lactic acid bacteria and Pseudomonas increased during storage in patties with macroalgae. | The addition of macroalgal extract was effective against lipid oxidation, although not enough to maintain color stability during the storage of the patties. | Lorenzo et al. [134] |
| Sausages (frankfurters) | Porphyra umbilicalis, Palmaria palmata, H. elongata and U. pinnatifida. Addition to food: 1%. | Total viable counts. | Sausages containing macroalgae had higher total viable counts at day 15 compared to control with bacterial counts exceeded by day 55 of storage. The frankfurters containing U. pinnatifida had higher counts compared to the other macroalgae used. | Significant differences in sensory attributes (color, external appearance, aroma, flavor and texture) were different depending on the macroalgae used. Reformulated frankfurters containing H. elongata were better accepted compared to the other macroalgae. | Vilar et al. [135] |
| Milk and dairy products | |||||
| Raw whole cow’s milk | Ascophyllum nodosum and Fucus vesiculosus extracts (aqueous and methanolic extracts). Addition to food products: 0.25 and 0.5%. | Total plate count, total coliforms, yeasts and molds. | No effect on the studied microorganisms. | Macroalgal extracts were stable in milk and their antioxidant activities were high before and after an in vitro digestion. Macroalgal extracts improved milk quality and shelf life characteristics. | O’Sullivan et al. [136] |
| Yogurt | A. nodosum and F. vesiculosus extracts (aqueous and methanolic extracts). Addition to food: 0.25 and 0.5%. | S. thermophilus and Lactobacillus delbrueckii subsp. bulgaricus. | No effect on the studied microorganisms. | No effect on pH, whey separation or negative effects on shelf-life of yogurt. Increased yellowness and reduced levels of lipid oxidation in product containing macroalgal extracts. The addition of macroalgal extracts had no effect on the antioxidant properties of the yogurt. Yogurts containing A. nodosum aqueous extract were preferred by the sensory panelists. | O’Sullivan et al. [137] |
| Cheese | Dried macroalgae (H. elongata, Laminaria ochroleuca, P. umbilicalis, Ulva lactuca and U. pinnatifida). Addition to food: 0.1%. | Lactic acid bacteria, lactobacilli, enterococci, Gram-negative bacteria, coliforms, yeasts and molds. | Levels of all bacteria were similar in all cheeses. The levels of Gram-negative and coliforms were below the limit of quantification from day 40 of ripening onwards in cheeses containing P. umbilicalis and U. lactuca. | Cheese with macroalgae had increased whey retention and moisture content and a lower pH compared to control. The supplementation with H. elongata was preferred to the other macroalgae analyzed due to the improved sensory and antioxidant characteristics of cheese. | del Olmo et al. [138] |
| Bakery products | |||||
| Bread | Myagropsis myagroides ethanolic extracts. Addition to food products: 0.5, 1 and 2%. | Total viable counts. | Decreased total microbial count was appreciated in bread containing 2% of macroalgal extract. | Breads containing 0.5% of extract had improved shelf-life, overall quality and acceptance compared to non-supplemented bread. | Lee et al. [139] |
| Muffins | Ecklonia cava hot water extracts. Addition to food products: 0.1, 0.5 and 1%. | Total viable counts. | Muffins containing 1% of extract had a low total microbial count during storage compared to control products. | All supplemented muffins had acceptable sensory attributes (color, flavor, taste, texture and overall acceptability). The antioxidant activities of the products increased with the addition of macroalgal extracts in the products. | Jung et al. [140] |
| Bread | Sargassum sagamianum extract. Addition to food: 0.25, 0.5 and 0.75%. | Total viable counts and mold. | Bread containing extracts had reduced total microbial counts (2 log cycles) and mold counts (3 log cycles) compared to non-supplemented bread. | No changes in moisture content or pH (days 3 to 9). Yellowness of the product increased due to the addition of the macroalgal extract. Breads containing 0.25 and 0.5% of extract had preferable sensory attributes (taste, total preference, inner shape and color) compared to control and highly supplemented breads. | Kim et al. [141] |
| Beverages | |||||
| Ultra-high temperature processed apple beverage | F. vesiculosus (fucoidan extract ≥95% purity). Addition to food: 25, 100 and 1000 μg/mL. | L. monocytogenes and S. Typhimurium (CECT 443). | No effect of fucoidan was detected on L. monocytogenes in beverages at concentrations of 5–25 μg/mL. When the concentration of fucoidan increased to 50 μg/mL, a bacteriostatic effect was appreciated and concentrations of fucoidan of 100–1000 μg/mL were bactericidal. The higher the concentration of fucoidan added, the shorter the exposure time required to completely inactivate S. Typhimurium in the food matrix. In addition, 1000 μg/mL fucoidan reduced the S. Typhimurium initial counts by 4 log cycles after 5 days of exposure incubated at 8 °C. | The organoleptic properties of the apple juice remained intact after the addition of fucoidan (25–1000 μg/mL). | Poveda-Castillo et al. [142] |
| Fish and seafood products | |||||
| Cold smoked salmon slices and fillets | Alginate and carrageenan edible films. | L. monocytogenes (PSU1, PSU9, F5069, ATCC 19115 and Scott A), anaerobic and aerobic bacteria counts. | Alginate coating was the most effective film at inhibiting the growth of L. monocytogenes. Coatings also suppressed the growth of spoilage aerobes and anaerobes, with populations 3.7–4.0 and 2.8–3.0 log CFU/g lower than untreated controls after 30 days of refrigeration. | - | Neetoo et al. [143] |
| Fresh fish burgers | H. elongata and P. palmata edible films containing macroalgae or macroalgal aqueous extracts. | Total aerobic mesophilic and total psychrotrophic bacteria. | Edible films with P. palmata and macroalgal extracts were less effective in reducing the microbial growth of total aerobic mesophilic and total aerobic psychrotrophic bacteria compared to films containing macroalga H. elongata. | Edible films with macroalgae controlled effectively the pH and water activity changes over storage of fish burgers. Reduced lipid oxidation and increased antioxidant capacity of trout burgers over storage when using edible films with macroalgae. | Albertos et al. [144] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabral, E.M.; Oliveira, M.; Mondala, J.R.M.; Curtin, J.; Tiwari, B.K.; Garcia-Vaquero, M. Antimicrobials from Seaweeds for Food Applications. Mar. Drugs 2021, 19, 211. https://doi.org/10.3390/md19040211
Cabral EM, Oliveira M, Mondala JRM, Curtin J, Tiwari BK, Garcia-Vaquero M. Antimicrobials from Seaweeds for Food Applications. Marine Drugs. 2021; 19(4):211. https://doi.org/10.3390/md19040211
Chicago/Turabian StyleCabral, Eduarda M., Márcia Oliveira, Julie R. M. Mondala, James Curtin, Brijesh K. Tiwari, and Marco Garcia-Vaquero. 2021. "Antimicrobials from Seaweeds for Food Applications" Marine Drugs 19, no. 4: 211. https://doi.org/10.3390/md19040211
APA StyleCabral, E. M., Oliveira, M., Mondala, J. R. M., Curtin, J., Tiwari, B. K., & Garcia-Vaquero, M. (2021). Antimicrobials from Seaweeds for Food Applications. Marine Drugs, 19(4), 211. https://doi.org/10.3390/md19040211