
Fish Oil, Se Yeast, and Micronutrient-Enriched Nutrition as Adjuvant Treatment during Target Therapy in a Murine Model of Lung Cancer



Abstract
:1. Introduction
2. Results
2.1. Cancer Cachexia
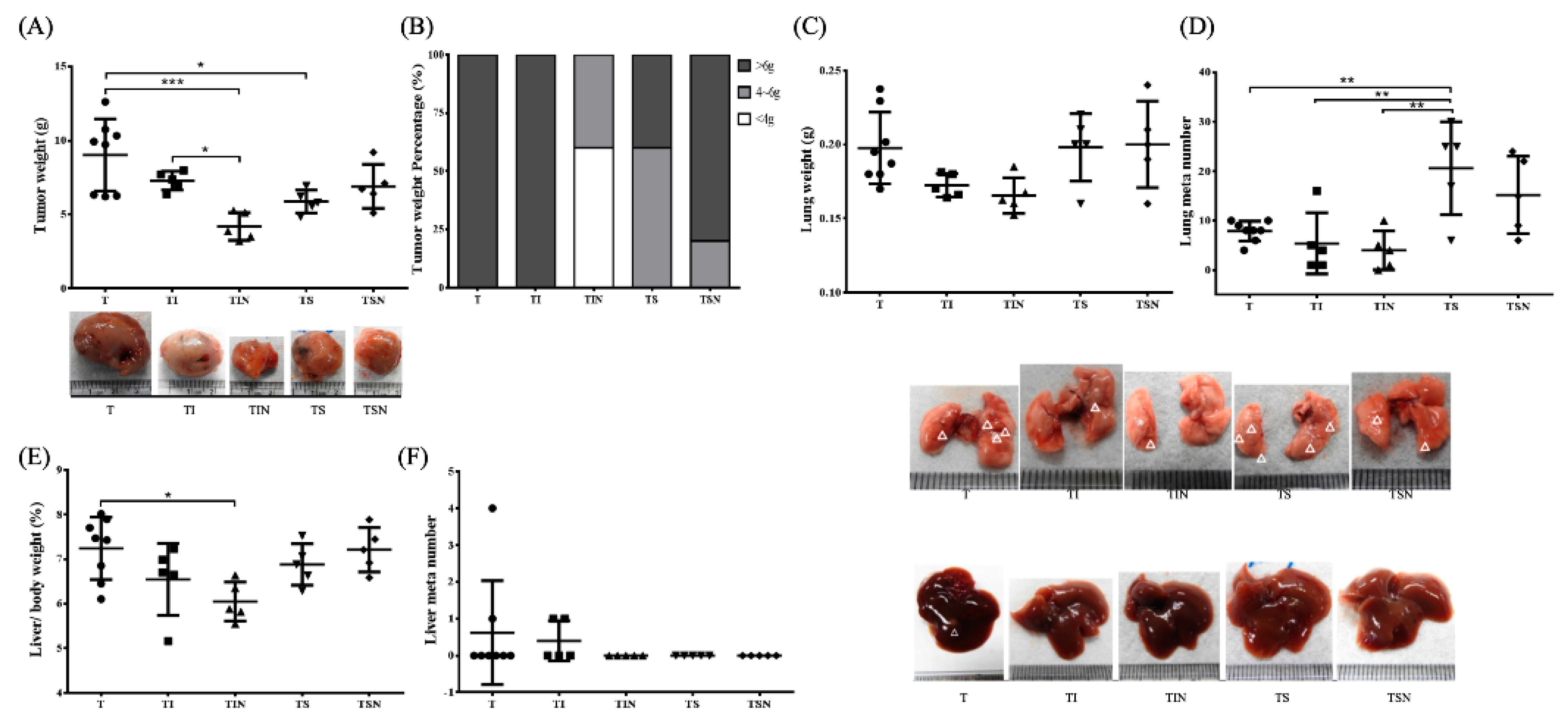
2.2. Tumor Growth and Metastases
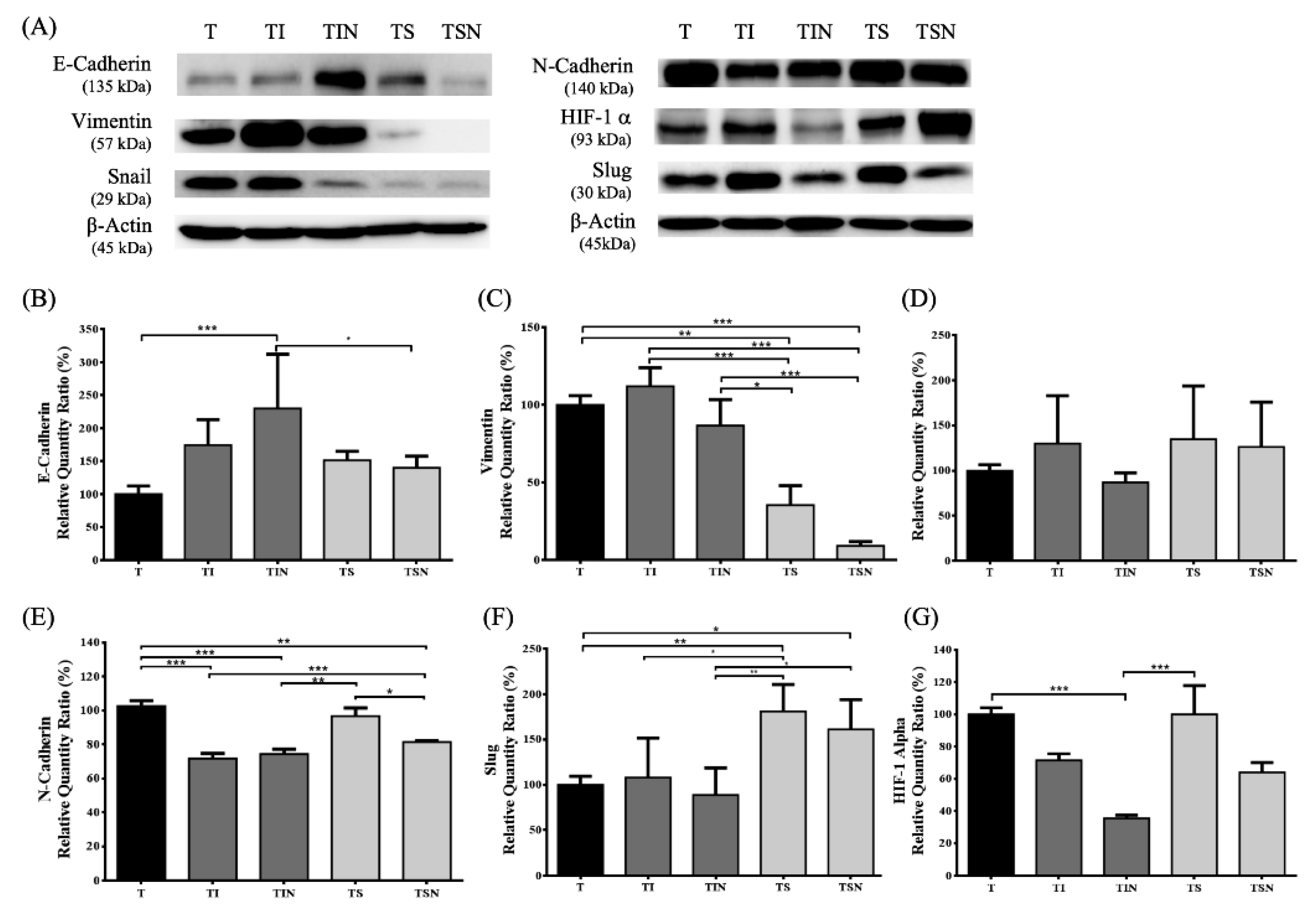
2.3. Inflammation and Oxidation
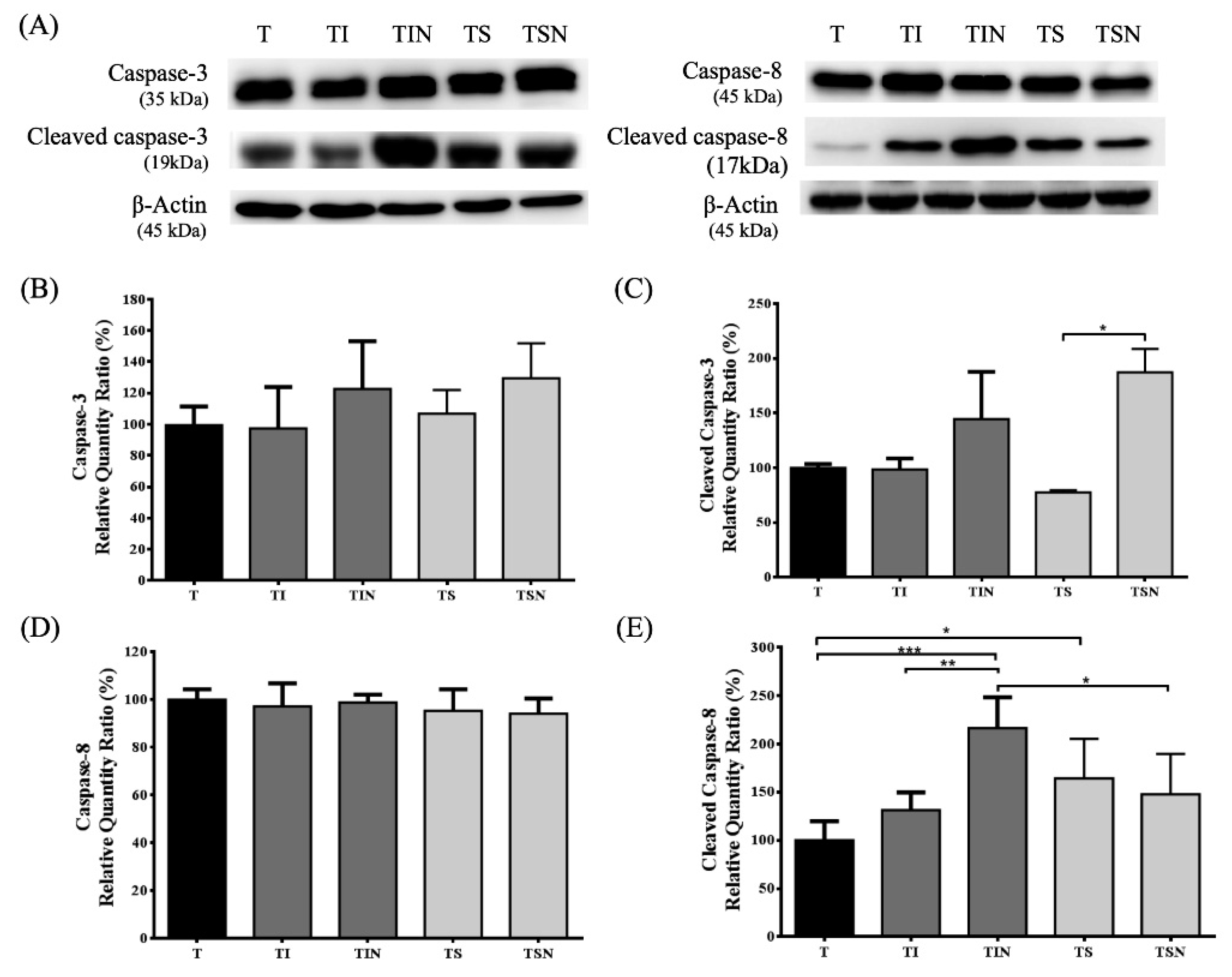
2.4. Tumor Cell Apoptosis
3. Discussion
4. Materials and Methods
4.1. Cell Line and Animals
4.2. Experimental Diet
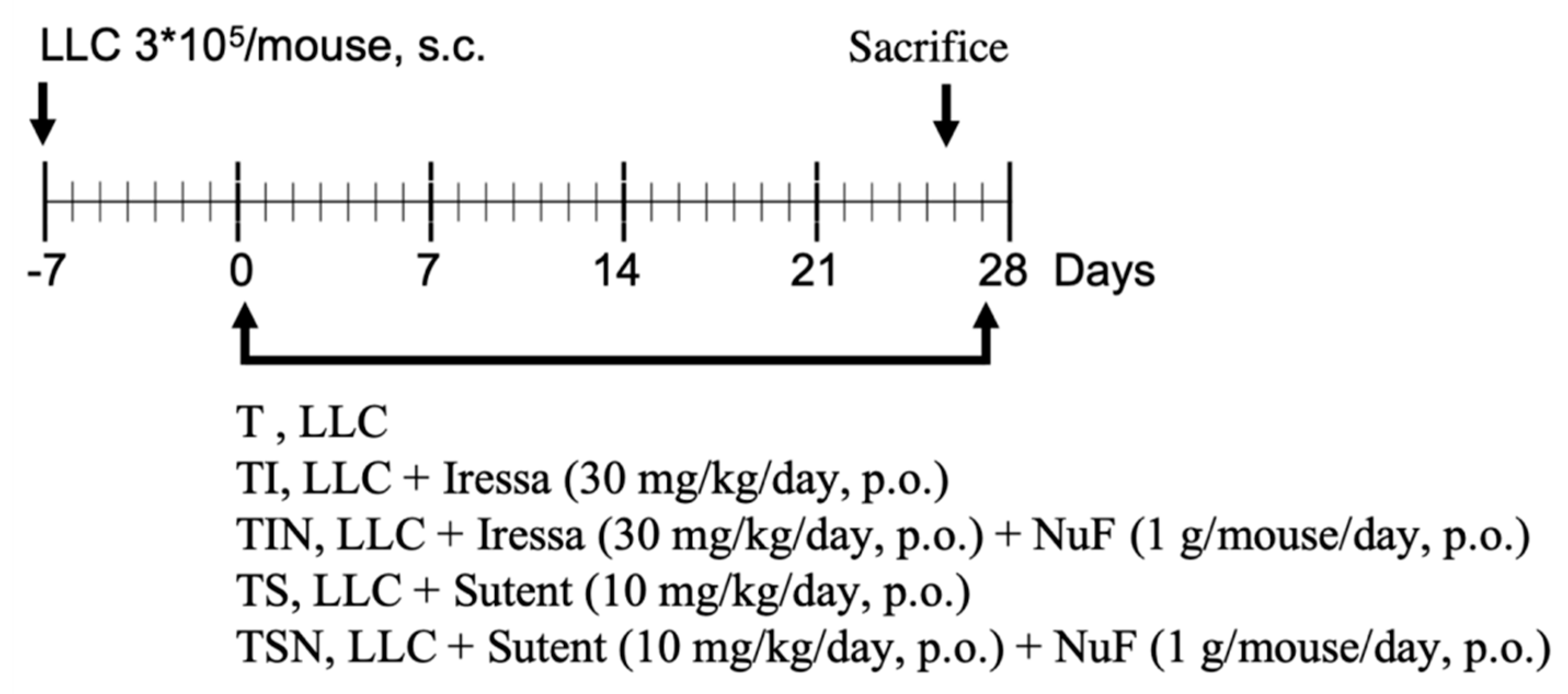
4.3. Experimental Design
4.4. Measurement of Oxidative Stress
4.5. Measurement of Cytokines in Tumor Tissue
4.6. Western Blot
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef]
- Hartmann, J.T.; Haap, M.; Kopp, H.G.; Lipp, H.P. Tyrosine kinase inhibitors—A review on pharmacology, metabolism and side effects. Curr. Drug Metab. 2009, 10, 470–481. [Google Scholar] [CrossRef]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo. Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Argiles, J.M.; Lopez-Soriano, F.J.; Busquets, S. Mechanisms and treatment of cancer cachexia. Nutr. Metab. Cardiovasc. Dis. 2013, 23 (Suppl 1), S19–S24. [Google Scholar] [CrossRef]
- Wallenberg, M.; Misra, S.; Bjornstedt, M. Selenium cytotoxicity in cancer. Basic Clin. Pharmacol. Toxicol. 2014, 114, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berquin, I.M.; Edwards, I.J.; Chen, Y.Q. Multi-targeted therapy of cancer by omega-3 fatty acids. Cancer Lett. 2008, 269, 363–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.J.; Shaw, A.T. Resisting Resistance: Targeted Therapies in Lung Cancer. Trends Cancer 2016, 2, 350–364. [Google Scholar] [CrossRef] [Green Version]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, E.N.; Gao, H.; Anfossi, S.; Mego, M.; Reddy, N.G.; Debeb, B.; Giordano, A.; Tin, S.; Wu, Q.; Garza, R.J.; et al. Inflammation Mediated Metastasis: Immune Induced Epithelial-To-Mesenchymal Transition in Inflammatory Breast Cancer Cells. PLoS ONE 2015, 10, e0132710. [Google Scholar]
- Sosroseno, W.; Barid, I.; Herminajeng, E.; Susilowati, H. Nitric oxide production by a murine macrophage cell line (RAW264.7) stimulated with lipopolysaccharide from Actinobacillus actinomycetemcomitans. Oral Microbiol. Immunol. 2002, 17, 72–78. [Google Scholar] [CrossRef]
- Barrera, G. Oxidative stress and lipid peroxidation products in cancer progression and therapy. ISRN Oncol. 2012, 2012, 137289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenhalgh, J.; Dwan, K.; Boland, A.; Bates, V.; Vecchio, F.; Dundar, Y.; Jain, P.; Green, J.A. First-line treatment of advanced epidermal growth factor receptor (EGFR) mutation positive non-squamous non-small cell lung cancer. Cochrane Database Syst. Rev. 2016, 5, CD010383. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, P.; Cooper, G.M. Activation of the CPP32 apoptotic protease by distinct signaling pathways with differential sensitivity to Bcl-xL. J. Biol. Chem. 1996, 271, 17601–17604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohoe, C.L.; Ryan, A.M.; Reynolds, J.V. Cancer cachexia: Mechanisms and clinical implications. Gastroenterol. Res. Pr. 2011, 2011, 601434. [Google Scholar] [CrossRef] [Green Version]
- Pappalardo, G.; Almeida, A.; Ravasco, P. Eicosapentaenoic acid in cancer improves body composition and modulates metabolism. Nutrition 2015, 31, 549–555. [Google Scholar] [CrossRef]
- Wang, H.; Chan, Y.L.; Li, T.L.; Bauer, B.A.; Hsia, S.; Wang, C.H.; Huang, J.S.; Wang, H.M.; Yeh, K.Y.; Huang, T.H.; et al. Reduction of splenic immunosuppressive cells and enhancement of anti-tumor immunity by synergy of fish oil and selenium yeast. PLoS ONE 2013, 8, e52912. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, T.L.; Hsia, S.; Su, I.L.; Chan, Y.L.; Wu, C.J. Skeletal muscle atrophy is attenuated in tumor-bearing mice under chemotherapy by treatment with fish oil and selenium. Oncotarget 2015, 6, 7758–7773. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.I.; Julliand, S.; Reeds, D.N.; Sinacore, D.R.; Klein, S.; Mittendorfer, B. Fish oil-derived n-3 PUFA therapy increases muscle mass and function in healthy older adults. Am. J. Clin. Nutr. 2015, 102, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Finocchiaro, C.; Segre, O.; Fadda, M.; Monge, T.; Scigliano, M.; Schena, M.; Tinivella, M.; Tiozzo, E.; Catalano, M.G.; Pugliese, M.; et al. Effect of n-3 fatty acids on patients with advanced lung cancer: A double-blind, placebo-controlled study. Br. J. Nutr. 2012, 108, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.Y.; Fattet, L.; Yang, J. Molecular pathways: Linking tumor microenvironment to epithelial-mesenchymal transition in metastasis. Clin. Cancer Res. 2015, 21, 962–968. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Gomez, S.J.; Maziveyi, M.; Alahari, S.K. Regulation of epithelial-mesenchymal transition through epigenetic and post-translational modifications. Mol. Cancer 2016, 15, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1alpha and HIF2alpha: Sibling rivalry in hypoxic tumour growth and progression. Nat. Rev. Cancer 2011, 12, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.J.; Semenza, G.L.; Zhang, H.F. Hypoxia-inducible factor 1 and breast cancer metastasis. J. Zhejiang Univ. Sci. B 2015, 16, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Cabrera, M. Mesenchymal Conversion of Mesothelial Cells Is a Key Event in the Pathophysiology of the Peritoneum during Peritoneal Dialysis. Adv. Med. 2014, 2014, 473134. [Google Scholar] [CrossRef]
- Jiang, F.; Yang, Y.; Xue, L.; Li, B.; Zhang, Z. 1α,25-dihydroxyvitamin D3 Attenuates TGF-β-Induced Pro-Fibrotic Effects in Human Lung Epithelial Cells through Inhibition of Epithelial-Mesenchymal Transition. Nutrients 2017, 9, 980. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, R.; Balaraman, R.; Sen, A.K.; Shukla, D.; Seth, A. Effect of concomitant administration of coenzyme Q10 with sitagliptin on experimentally induced diabetic nephropathy in rats. Ren. Fail. 2017, 39, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Ilag, L.L. Are Long-Chain Polyunsaturated Fatty Acids the Link between the Immune System and the Microbiome towards Modulating Cancer? Medicines 2018, 5, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Dizman, N.; Poroyko, V.; Won, H.; Bergerot, C.D.; Bergerot, P.G.; Maia, M.C.; Hsu, J.; Frankel, P.H.; Jones, J.; et al. Gut microbiome composition and response to sunitinib in metastatic renal cell carcinoma (mRCC). J. Clin. Oncol. 2018, 36, 657. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, J.; Zhou, S.; Yu, L.; Han, F.; Ling, R.; Ling, J. Tentative identification of gefitinib metabolites in non-small-cell lung cancer patient plasma using ultra-performance liquid chromatography coupled with triple quadrupole time-of-flight mass spectrometry. PLoS ONE 2020, 15, e0236523. [Google Scholar] [CrossRef]
- Conley, S.J.; Gheordunescu, E.; Kakarala, P.; Newman, B.; Korkaya, H.; Heath, A.N.; Clouthier, S.G.; Wicha, M.S. Antiangiogenic agents increase breast cancer stem cells via the generation of tumor hypoxia. Proc. Natl. Acad. Sci. USA 2012, 109, 2784–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Lu, G.; Yao, Y.; Gu, W. An Autocrine IL-6/IGF-1R Loop Mediates EMT and Promotes Tumor Growth in Non-small Cell Lung Cancer. Int. J. Biol. Sci. 2019, 15, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Manzi, L.; Costantini, L.; Molinari, R.; Merendino, N. Effect of Dietary omega-3 Polyunsaturated Fatty Acid DHA on Glycolytic Enzymes and Warburg Phenotypes in Cancer. Biomed. Res. Int. 2015, 2015, 137097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conklin, K.A. Chemotherapy-associated oxidative stress: Impact on chemotherapeutic effectiveness. Integr. Cancer Ther. 2004, 3, 294–300. [Google Scholar] [CrossRef]
- Fuchs-Tarlovsky, V. Role of antioxidants in cancer therapy. Nutrition 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.K.; Jain, A.; Jain, A.; Sharma, T.; Singh, B.; Kesharwani, P. Administration of antioxidants in cancer: Debate of the decade. Drug Discov. Today 2018, 23, 763–770. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushmorov, A.; Ratter, F.; Lehmann, V.; Droge, W.; Schirrmacher, V.; Umansky, V. Nitric-oxide-induced apoptosis in human leukemic lines requires mitochondrial lipid degradation and cytochrome C release. Blood 1999, 93, 2342–2352. [Google Scholar] [CrossRef]
- Umansky, V.; Ushmorov, A.; Ratter, F.; Chlichlia, K.; Bucur, M.; Lichtenauer, A.; Rocha, M. Nitric oxide-mediated apoptosis in human breast cancer cells requires changes in mitochondrial functions and is independent of CD95 (APO-1/Fas). Int. J. Oncol. 2000, 16, 109–117. [Google Scholar] [CrossRef]
- Ovadje, P.; Chatterjee, S.; Griffin, C.; Tran, C.; Hamm, C.; Pandey, S. Selective induction of apoptosis through activation of caspase-8 in human leukemia cells (Jurkat) by dandelion root extract. J. Ethnopharmacol. 2011, 133, 86–91. [Google Scholar] [CrossRef]
- Guo, C.H.; Hsia, S.; Chen, P.C. Distribution of selenium and oxidative stress in breast tumor-bearing mice. Nutrients 2013, 5, 594–607. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrawell powder | 1 Serving: 75 g |
|---|---|
| Components (Units) | Amount per Serving |
| Calorie (kcal) | 294 |
| Protein (g) | 19 |
| Fat (g) | 8.7 |
| Saturated fatty acids (g) | 1.1 |
| Monounsaturated fatty acids (g) | 3.7 |
| Polyunsaturated fatty acids (g) | 3.9 |
| Fish oil (Omega-3 fatty acid; n-3) (g) | 1.4 |
| Trans fatty acids (g) | 0 |
| Carbohydrates (g) | 37 |
| Diet fiber (g) | 4.2 |
| Sugar (g) | 3 |
| Sodium (mg) | 360 |
| Cholesterol (mg) | 0 |
| Vitamin A (µg) | 350 (1167 IU) |
| β-Carotene (µg) | 300 (1000 IU) |
| Vitamin B1 (mg) | 1.4 |
| Vitamin B2 (mg) | 1.6 |
| Vitamin B6 (mg) | 1.5 |
| Vitamin B12 (µg) | 2.4 |
| Vitamin C (mg) | 200 |
| Vitamin D (µg) | 5 (200 IU) |
| Vitamin E (mg) | 13, d-α TE (19 IU) |
| Vitamin K (µg) | 37.5 |
| Niacin (mg) | 13.5 |
| Folic acid (µg) | 600 |
| Pantothenic acid (mg) | 5 |
| Biotin (µg) | 100 |
| Choline (mg) | 300 |
| Coenzyme Q10 (mg) | 30 |
| Calcium (mg) | 480 |
| Phosphorous (mg) | 250 |
| Iron (mg) | 4.5 |
| Iodine (µg) | 52 |
| Magnesium (mg) | 117 |
| Zinc (mg) | 7.5 |
| Selenium yeast (µg) | 110 |
| Potassium (mg) | 682 |
| Copper (µg) | 400 |
| Manganese (µg) | 1.5 |
| Chromium (µg) | 90 |
| Molybdenum (µg) | 56.3 |
| (A) Physiological Cachectic Parameters | ||||||||
| Treatment | N | IW (g) | CW (g) | Dela CW (%) | GM (mg) | WAT (mg) | BAT (mg) | Spleen (mg) |
| T | 8 | 21.3 ± 1.3 | 18.80 ± 1.58 | −11.80 ± 9.0 | 86 ± 11 | 13 ± 17 | 20 ± 14 | 480 ± 121 |
| TI | 5 | 20.3 ± 1.4 | 18.28 ± 1.56 | −8.20 ± 10.1 | 88 ± 10 | 36 ± 21 | 43 ± 11 | 354 ± 35 |
| TIN | 5 | 20.9 ± 1.5 | 20.86 ± 2.06 | −0.20 ± 9.8 * | 108 ± 7 | 72 ± 29 **,+ | 57 ± 9 * | 254 ± 50 ** |
| TS | 5 | 21.3 ± 1.2 | 18.17 ± 1.49 | −7.90 ± 7.8 | 86 ± 15 | 39 ± 18 | 55 ± 10* | 403 ± 78 |
| TSN | 5 | 21.9 ± 1.5 | 19.15 ± 1.00 | −4.35 ± 6.6 | 109 ± 38 | 64 ± 21 ** | 63 ± 31 ** | 260 ± 66 ** |
| (B) Immune and Oxidative Parameters | ||||||||
| Treatment | N | Tumor | Serum | |||||
| IL-6 (pg/mL) | TNF-α (pg/mL) | TGF-β (pg/mL) | MDA (uM) | NO (%) | MDA (μM) | NO (%) | ||
| T | 8 | 5.3 ± 1.7 | 51.6 ± 18.2 | 35.9 ± 5.9 | 51.1 ± 28.6 | 100 ± 28.6 | 6.0 ± 4.5 | 100 ± 19.7 |
| TI | 5 | 4.7 ± 2.9 | 47.8 ± 9.9 | 34.6 ± 3.8 | 49.3 ± 28.6 | 111.4 ± 30.7 | 4.2 ± 1.0 * | 78.1 ± 15.2 ** |
| TIN | 5 | 3.2 ± 1.1 # | 45.9 ± 11.4 | 28.6 ± 6.7 * | 40.3 ± 26.0 | 128.4±24.6 *,+ | 2.3 ± 0.8 * | 65.8 ± 20.2 ** |
| TS | 5 | 6.5 ± 2.5 | 49.2 ± 11.8 | 34.6 ± 3.2 | 47.2 ± 30.5 | 99.9 ± 34.5 | 8.5 ± 5.1 | 89.4 ± 18.9 |
| TSN | 5 | 3.0 ± 0.8 # | 44.7 ± 22.2 | 30.0 ± 4.3 | 48.9 ± 35.4 | 121.2 ± 26.2 *,# | 7.1 ± 2.6 | 78.6 ± 13.4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Hsia, S.; Wu, T.-H.; Wu, C.-J. Fish Oil, Se Yeast, and Micronutrient-Enriched Nutrition as Adjuvant Treatment during Target Therapy in a Murine Model of Lung Cancer. Mar. Drugs 2021, 19, 262. https://doi.org/10.3390/md19050262
Wang H, Hsia S, Wu T-H, Wu C-J. Fish Oil, Se Yeast, and Micronutrient-Enriched Nutrition as Adjuvant Treatment during Target Therapy in a Murine Model of Lung Cancer. Marine Drugs. 2021; 19(5):262. https://doi.org/10.3390/md19050262
Chicago/Turabian StyleWang, Hang, Simon Hsia, Tsung-Han Wu, and Chang-Jer Wu. 2021. "Fish Oil, Se Yeast, and Micronutrient-Enriched Nutrition as Adjuvant Treatment during Target Therapy in a Murine Model of Lung Cancer" Marine Drugs 19, no. 5: 262. https://doi.org/10.3390/md19050262
APA StyleWang, H., Hsia, S., Wu, T. -H., & Wu, C. -J. (2021). Fish Oil, Se Yeast, and Micronutrient-Enriched Nutrition as Adjuvant Treatment during Target Therapy in a Murine Model of Lung Cancer. Marine Drugs, 19(5), 262. https://doi.org/10.3390/md19050262