
Carotenoid Nostoxanthin Production by Sphingomonas sp. SG73 Isolated from Deep Sea Sediment

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
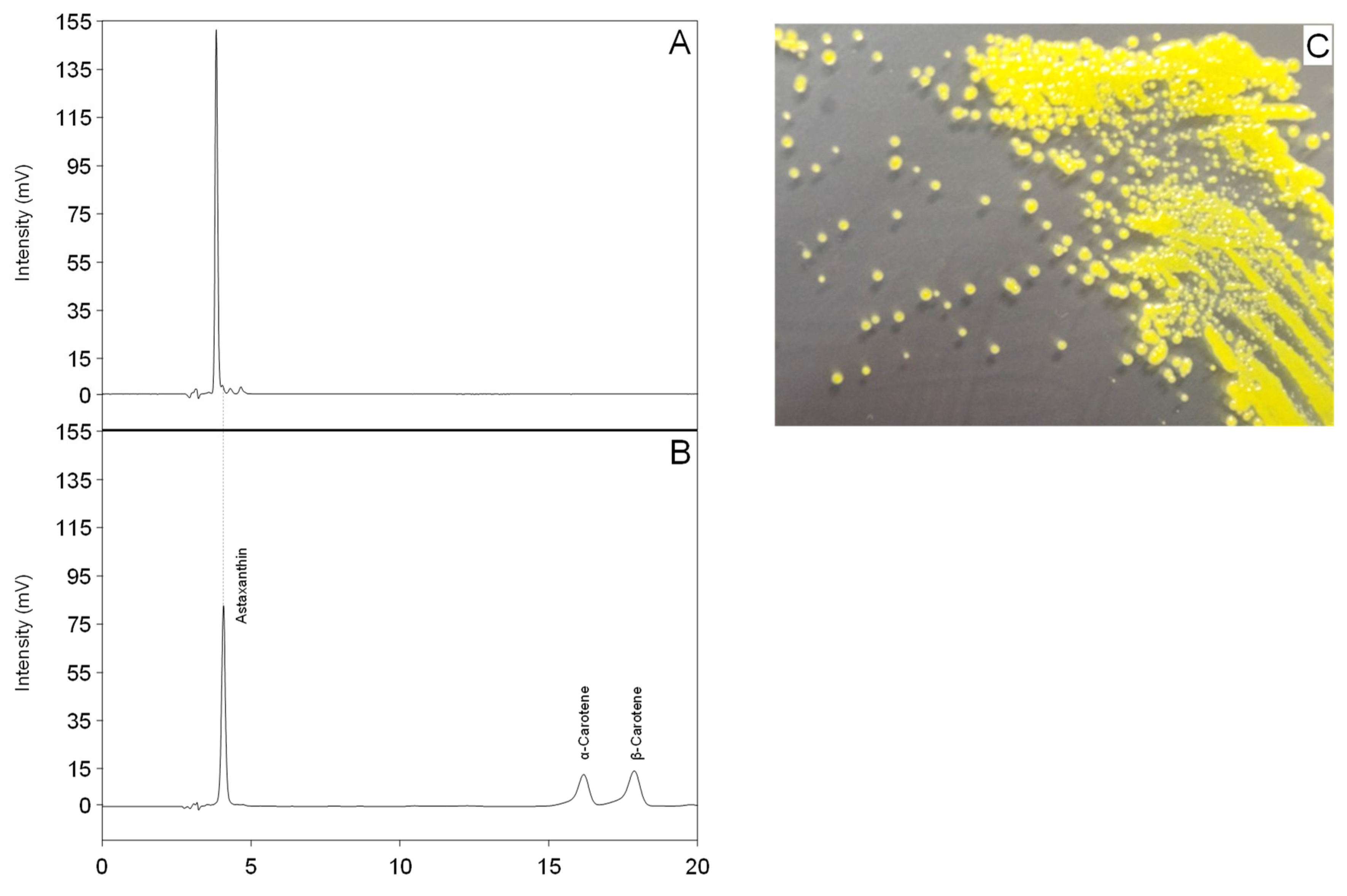
2.1. Isolation of a Yellow Microorganism from Deep Sea Sediment
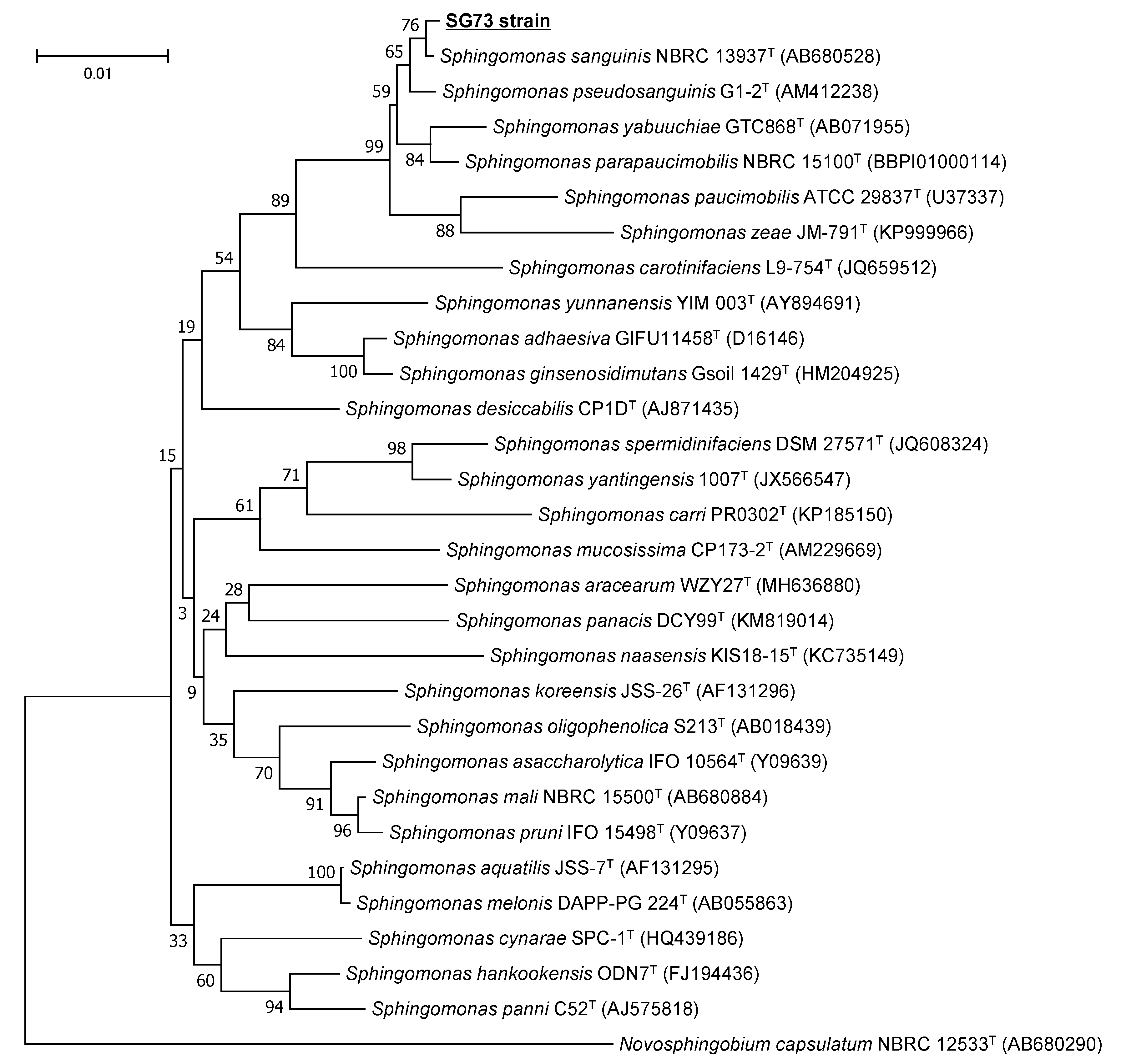
2.2. Identification of the Hydrophilic Carotenoid-Producing Strain SG73
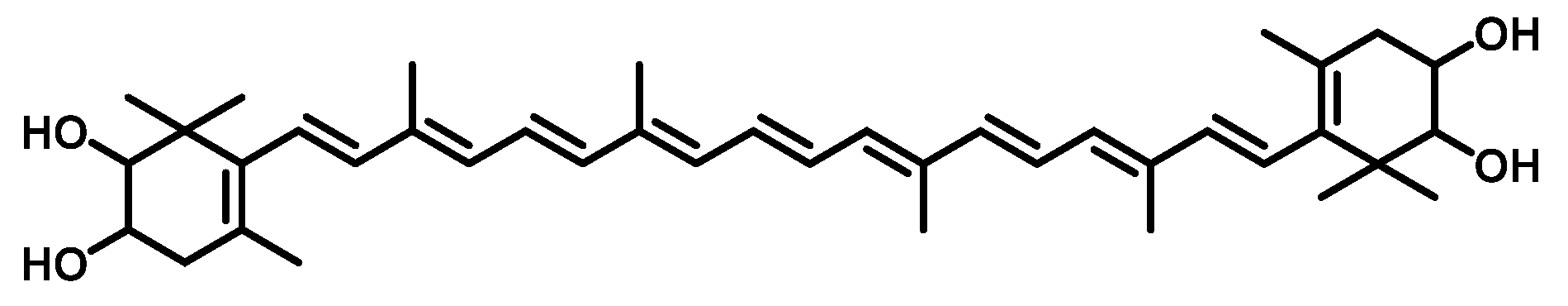
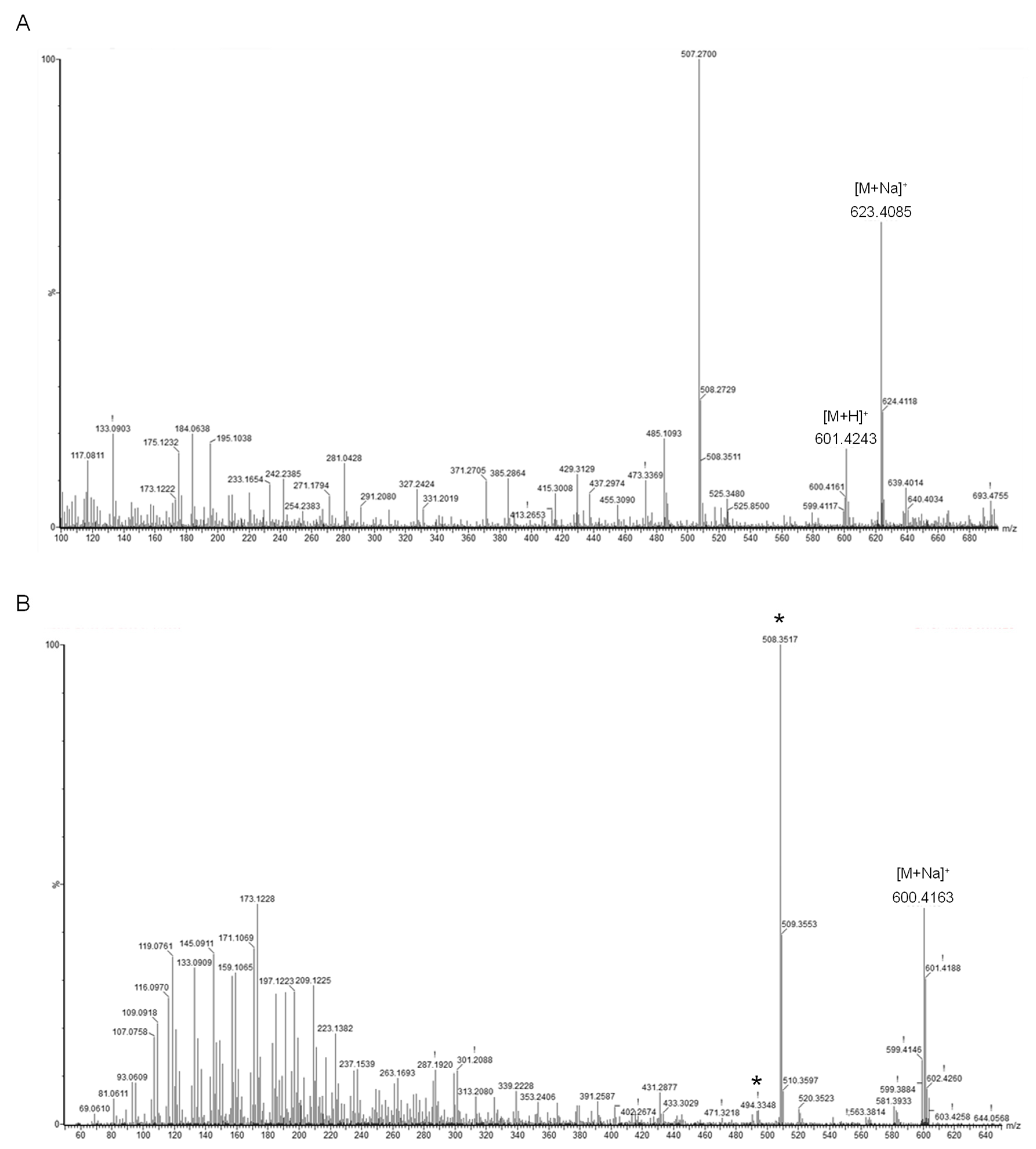
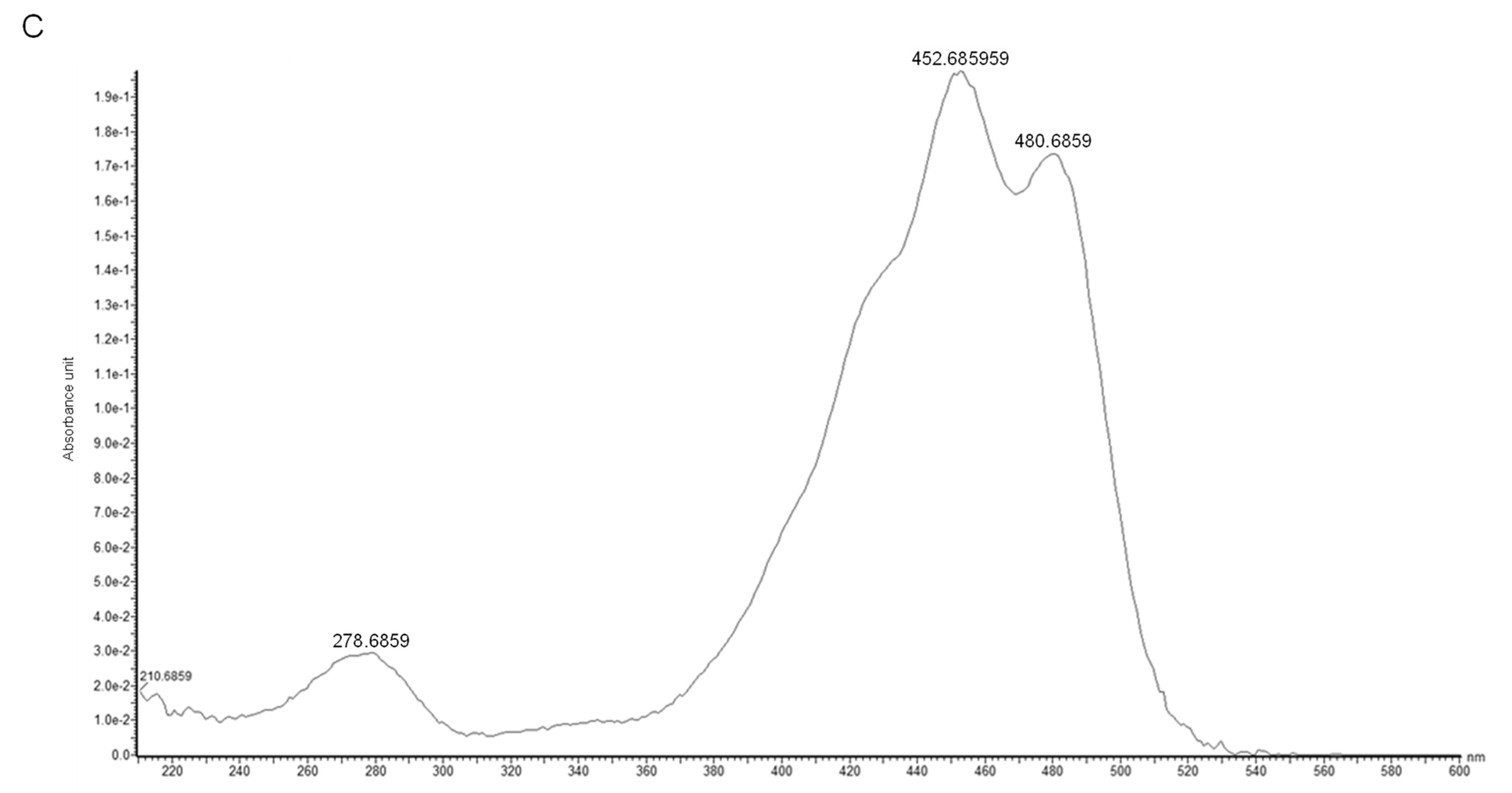
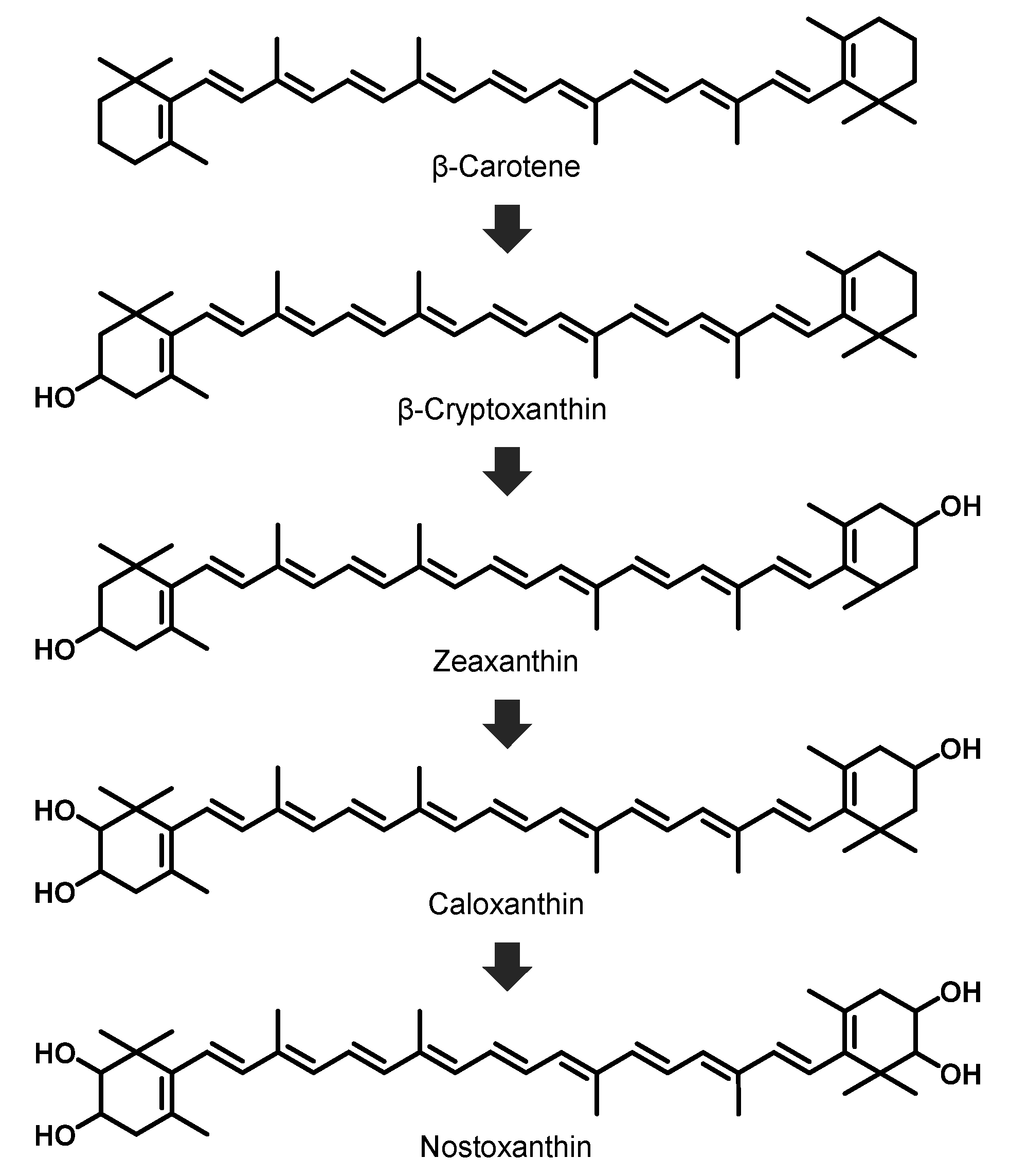
2.3. Identification of the Main Carotenoid Produced by Sphingomonas sp. SG73
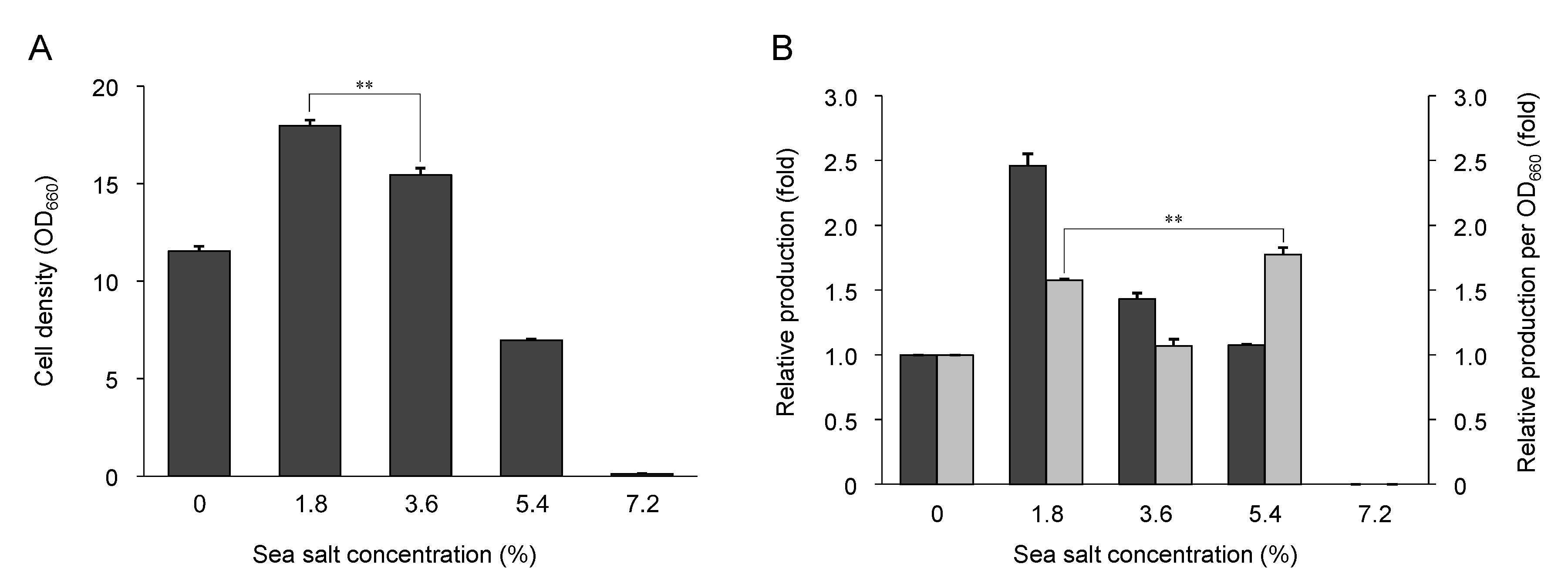
2.4. Effect of Sea Salt on Growth and Nostoxanthin Production
3. Materials and Methods
3.1. Materials
3.2. Strains and Media
3.3. Cultivation of Isolated Strains
3.4. Carotenoid Analysis
3.5. Identification of Carotenoid Structure
3.6. Genetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Britton, G. Carotenoid research: History and new perspectives for chemistry in biological systems. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158699. [Google Scholar] [CrossRef] [PubMed]
- Frank, H.A.; Cogdell, R.J. Carotenoids in Photosynthesis. Photochem. Photobiol. 1996, 63, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other Nutrients from Haematococcus pluvialis—Multifunctional Applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Jena, G. Intervention of astaxanthin against cyclophosphamide-induced oxidative stress and DNA damage: A study in mice. Chem. Interact. 2009, 180, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Hosokawa, M.; Miyashita, K. Fucoxanthin: A Marine Carotenoid Exerting Anti-Cancer Effects by Affecting Multiple Mechanisms. Mar. Drugs 2013, 11, 5130–5147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maoka, T. Carotenoids in Marine Animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in Algae: Distributions, Biosyntheses and Functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Otero, P.; Echave, J.; Carreira-Casais, A.; Chamorro, F.; Collazo, N.; Jaboui, A.; Lourenço-Lopes, C.; Simal-Gandara, J.; Prieto, M. Xanthophylls from the Sea: Algae as Source of Bioactive Carotenoids. Mar. Drugs 2021, 19, 188. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wu, X.; Li, O.; Qian, C.; Gao, H. Cloning and Characterization of Genes Involved in Nostoxanthin Biosynthesis of Sphingomonas elodea ATCC 31461. PLoS ONE 2012, 7, e35099. [Google Scholar] [CrossRef]
- Gich, F.; Overmann, J. Sandarakinorhabdus limnophila gen. nov., sp. nov., a novel bacteriochlorophyll a-containing, obligately aerobic bacterium isolated from freshwater lakes. Int. J. Syst. Evol. Microbiol. 2006, 56, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Hanada, S.; Kawase, Y.; Hiraishi, A.; Takaichi, S.; Matsuura, K.; Shimada, K.; Nagashima, K.V.P. Porphyrobacter tepidarius sp. nov., a Moderately Thermophilic Aerobic Photosynthetic Bacterium Isolated from a Hot Spring. Int. J. Syst. Bacteriol. 1997, 47, 408–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwai, M.; Maoka, T.; Ikeuchi, M.; Takaichi, S. 2,2’-beta-hydroxylase (CrtG) is involved in carotenogenesis of both nostoxanthin and 2-hydroxymyxol 2’-fucoside in Thermosynechococcus elongatus strain BP-1. Plant Cell Physiol. 2008, 49, 1678–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, C.L.; Andrewes, A.G.; McQuade, T.J.; Starr, M.P. The pigment of Pseudomonas paucimobilis is a carotenoid (Nostoxanthin), rather than a brominated aryl-polyene (xanthomonadin). Curr. Microbiol. 1979, 3, 1–4. [Google Scholar] [CrossRef]
- Nishida, Y.; Adachi, K.; Kasai, H.; Shizuri, Y.; Shindo, K.; Sawabe, A.; Komemushi, S.; Miki, W.; Misawa, N. Elucidation of a Carotenoid Biosynthesis Gene Cluster Encoding a Novel Enzyme, 2,2′-β-Hydroxylase, from Brevundimonas sp. Strain SD212 and Combinatorial Biosynthesis of New or Rare Xanthophylls. Appl. Environ. Microbiol. 2005, 71, 4286–4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaichi, S.; Shimada, K.; Ishidsu, J.-I. Carotenoids from the aerobic photosynthetic bacterium, Erythrobacter longus: β-Carotene and its hydroxyl derivatives. Arch. Microbiol. 1990, 153, 118–122. [Google Scholar] [CrossRef]
- Tao, L.; Rouvière, P.E.; Cheng, Q. A carotenoid synthesis gene cluster from a non-marine Brevundimonas that synthesizes hydroxylated astaxanthin. Gene 2006, 379, 101–108. [Google Scholar] [CrossRef]
- Kawahara, K.; Kuraishi, H. Chemical structure and function of glycosphingolipids of Sphingomonas spp. and their distribution among members of the alpha-4 subclass of Proteobacteria. J. Ind. Microbiol. Biotechnol. 1999, 23, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, N. MassBank Record: CA000155. Available online: http://www.massbank.jp/RecordDisplay?id=CA000155&dsn=Kyoto_Univ (accessed on 6 April 2021).
- Maoka, T. Structural studies of carotenoids in plants, animals, and food products. In Carotenoids Nutrition, Analysis and Technology; Kaczor, A., Baranska, M., Eds.; Wiley Blackwell: Oxford, UK, 2016; pp. 103–129. [Google Scholar]
- Britton, G. UV/Visible Spectrometry; Brikhauser Verlag: Basel, Switzerland, 1995; Volume 1B. [Google Scholar]
- Iwata, T.; Shinomura, Y.; Natori, Y.; Igarashi, Y.; Sohrin, R.; Suzuki, Y. Relationship between Salinity and Nutrients in the Subsurface Layer in the Suruga Bay. J. Oceanogr. 2005, 61, 721–732. [Google Scholar] [CrossRef]
- Hara, K.Y.; Kageyama, Y.; Tanzawa, N.; Hirono-Hara, Y.; Kikukawa, H.; Wakabayashi, K. Development of astaxanthin production from citrus peel extract using Xanthophyllomyces dendrorhous. Environ. Sci. Pollut. Res. 2021, 28, 12640–12647. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 33, 1870–1874. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Water Depth | |
|---|---|---|
| Spot 1 | 34°8482′ N, 138°3589′ E | 376 m |
| Spot 2 | 34°8526′ N, 138°3994′ E | 691 m |
| Spot 3 | 34°9149′ N, 138°6507′ E | 1548 m |
| Spot 4 | 34°9152′ N, 138°6534′ E | 1523 m |
| Spot 5 | 34°1872′ N, 138°4220′ E | 3595 m |
| Spot 6 | 34°2736′ N, 138°4813′ E | 3233 m |
| Spot 7 | 34°7847′ N, 138°6345′ E | 1652 m |
| Spot 8 | 34°9867′ N, 138°6967′ E | 1299 m |
| Spot 9 | 34°2409′ N, 138°4803′ E | 3372 m |
| Spot 10 | 34°7835′ N, 138°6171′ E | 1730 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikukawa, H.; Okaya, T.; Maoka, T.; Miyazaki, M.; Murofushi, K.; Kato, T.; Hirono-Hara, Y.; Katsumata, M.; Miyahara, S.; Hara, K.Y. Carotenoid Nostoxanthin Production by Sphingomonas sp. SG73 Isolated from Deep Sea Sediment. Mar. Drugs 2021, 19, 274. https://doi.org/10.3390/md19050274
Kikukawa H, Okaya T, Maoka T, Miyazaki M, Murofushi K, Kato T, Hirono-Hara Y, Katsumata M, Miyahara S, Hara KY. Carotenoid Nostoxanthin Production by Sphingomonas sp. SG73 Isolated from Deep Sea Sediment. Marine Drugs. 2021; 19(5):274. https://doi.org/10.3390/md19050274
Chicago/Turabian StyleKikukawa, Hiroshi, Takuma Okaya, Takashi Maoka, Masayuki Miyazaki, Keita Murofushi, Takanari Kato, Yoko Hirono-Hara, Masahiro Katsumata, Shoichi Miyahara, and Kiyotaka Y. Hara. 2021. "Carotenoid Nostoxanthin Production by Sphingomonas sp. SG73 Isolated from Deep Sea Sediment" Marine Drugs 19, no. 5: 274. https://doi.org/10.3390/md19050274
APA StyleKikukawa, H., Okaya, T., Maoka, T., Miyazaki, M., Murofushi, K., Kato, T., Hirono-Hara, Y., Katsumata, M., Miyahara, S., & Hara, K. Y. (2021). Carotenoid Nostoxanthin Production by Sphingomonas sp. SG73 Isolated from Deep Sea Sediment. Marine Drugs, 19(5), 274. https://doi.org/10.3390/md19050274