
A Soft Spot for Chemistry–Current Taxonomic and Evolutionary Implications of Sponge Secondary Metabolite Distribution

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Methods
3. Specificity and Phylogenetic Relevance of Sponge Compounds
3.1. General
3.2. Demospongiae
3.2.1. Heteroscleromorpha
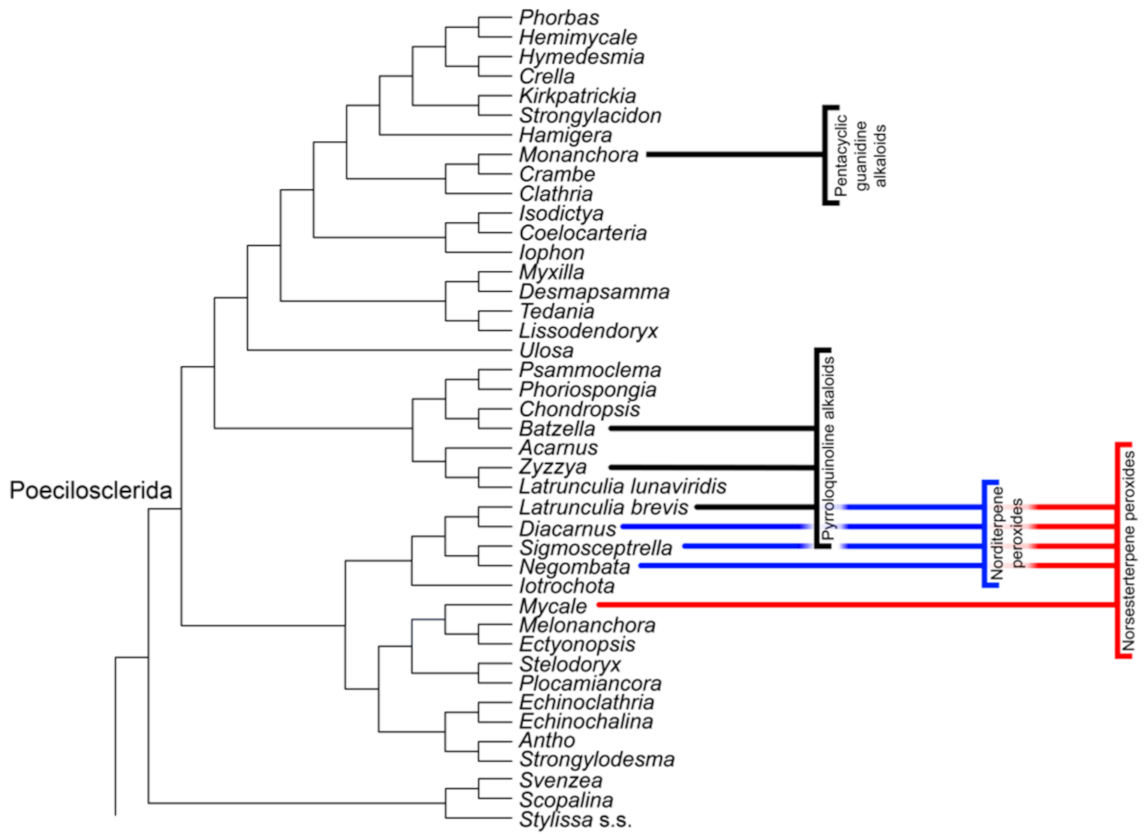
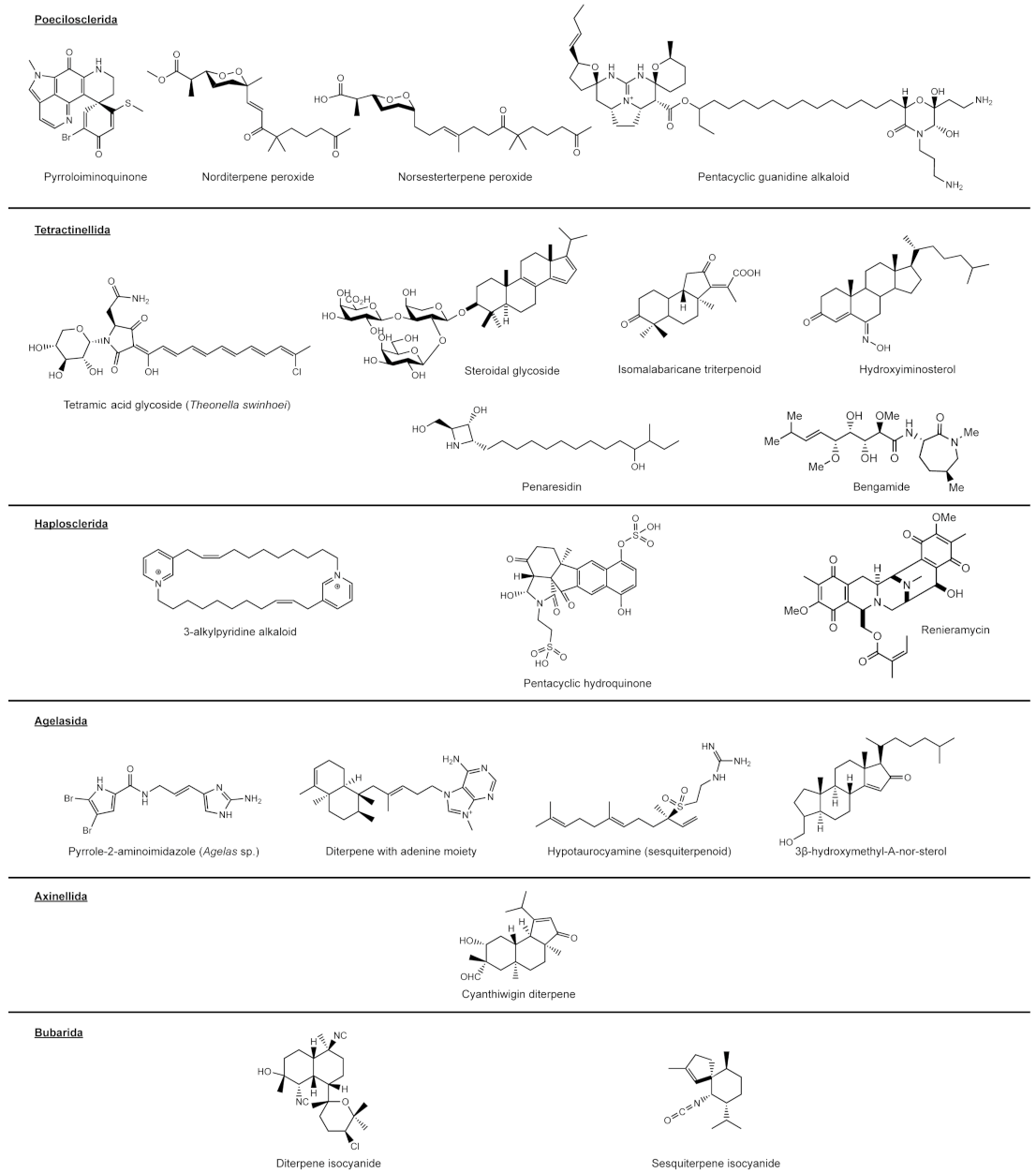
Poecilosclerida
Tetractinellida
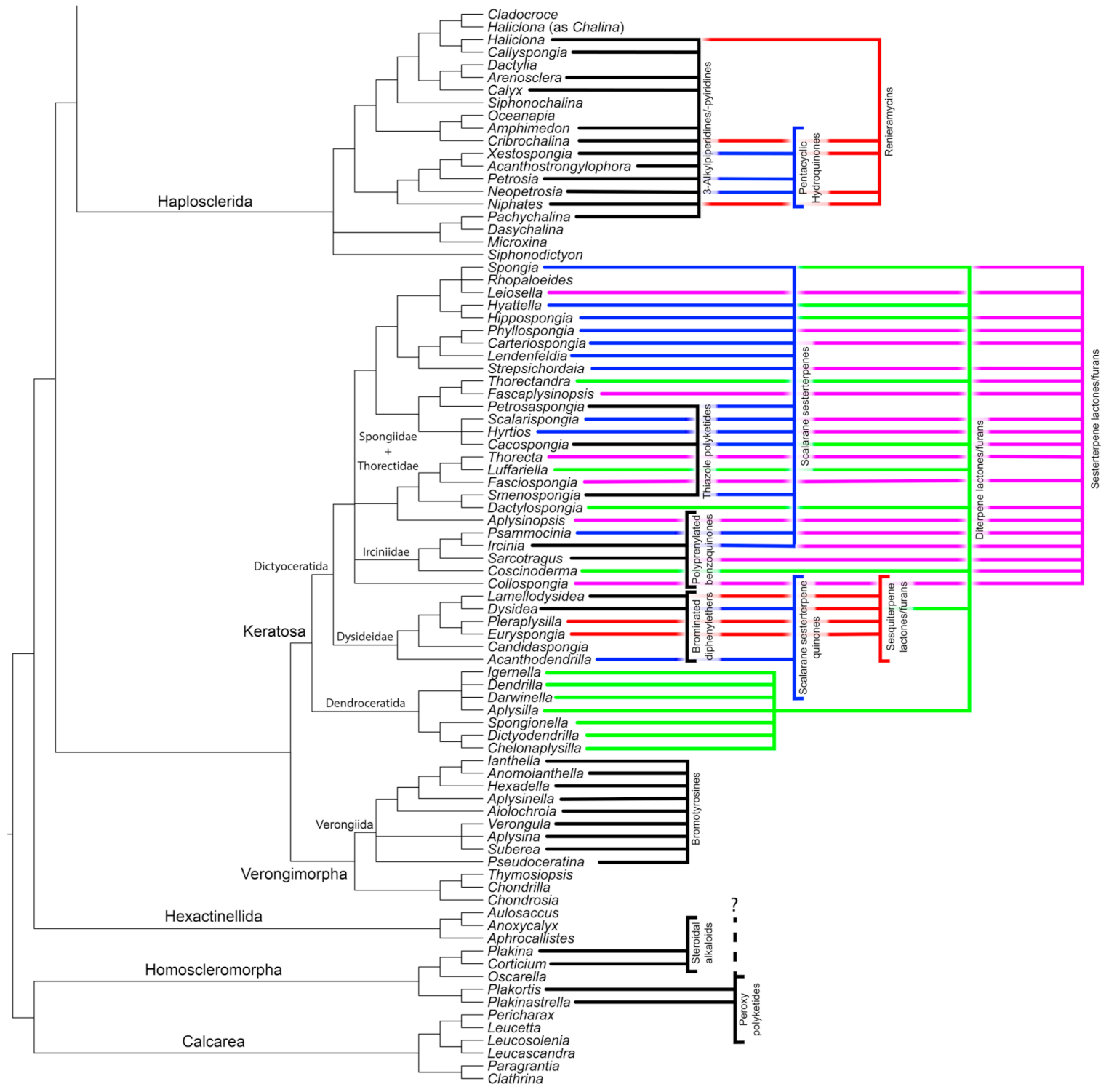
Haplosclerida
On Agelasida, Axinellida, Bubarida, and Suberitida
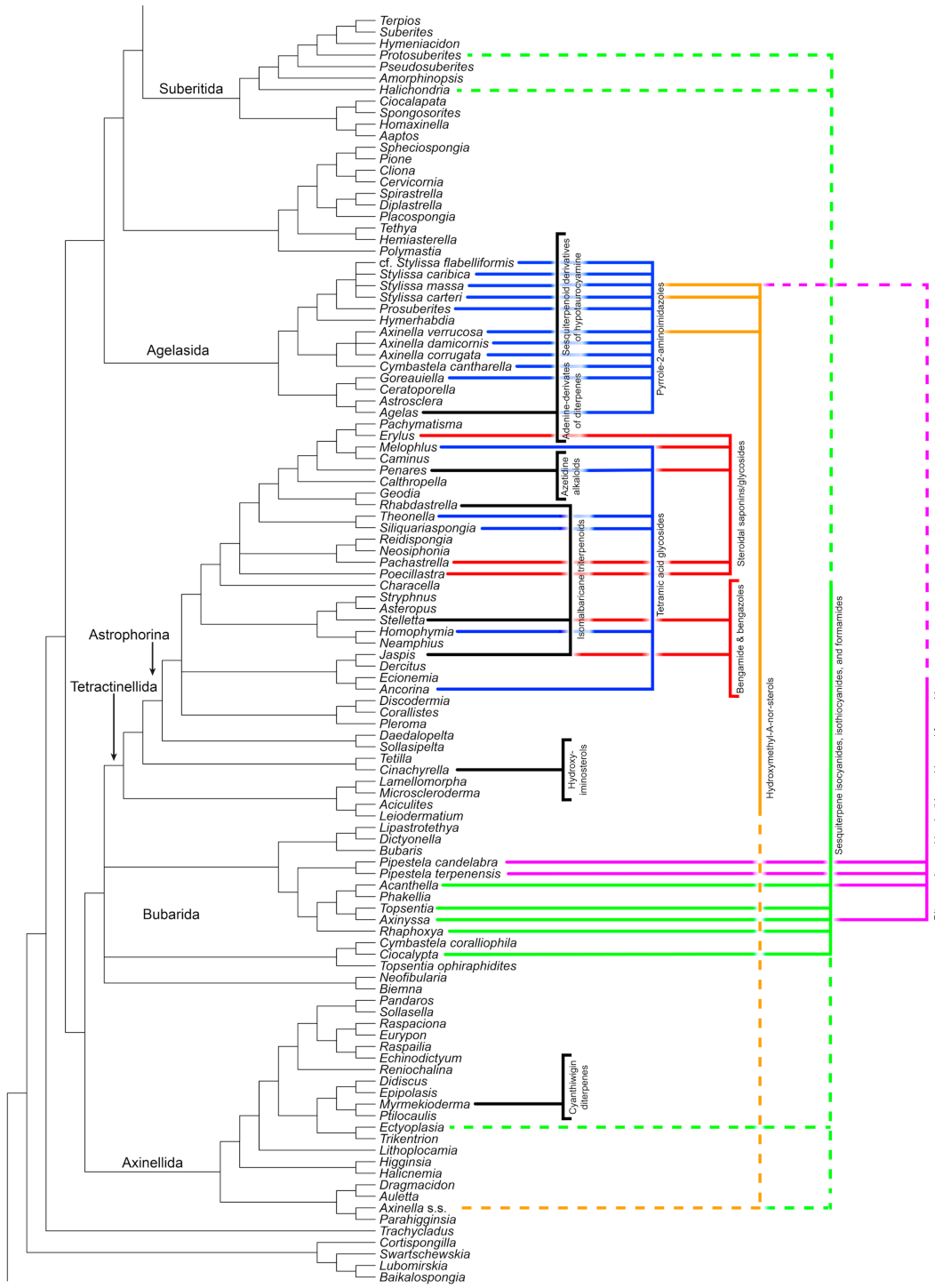
Agelasida
Axinellida
Bubarida and Suberitida “Incertae Sedis”
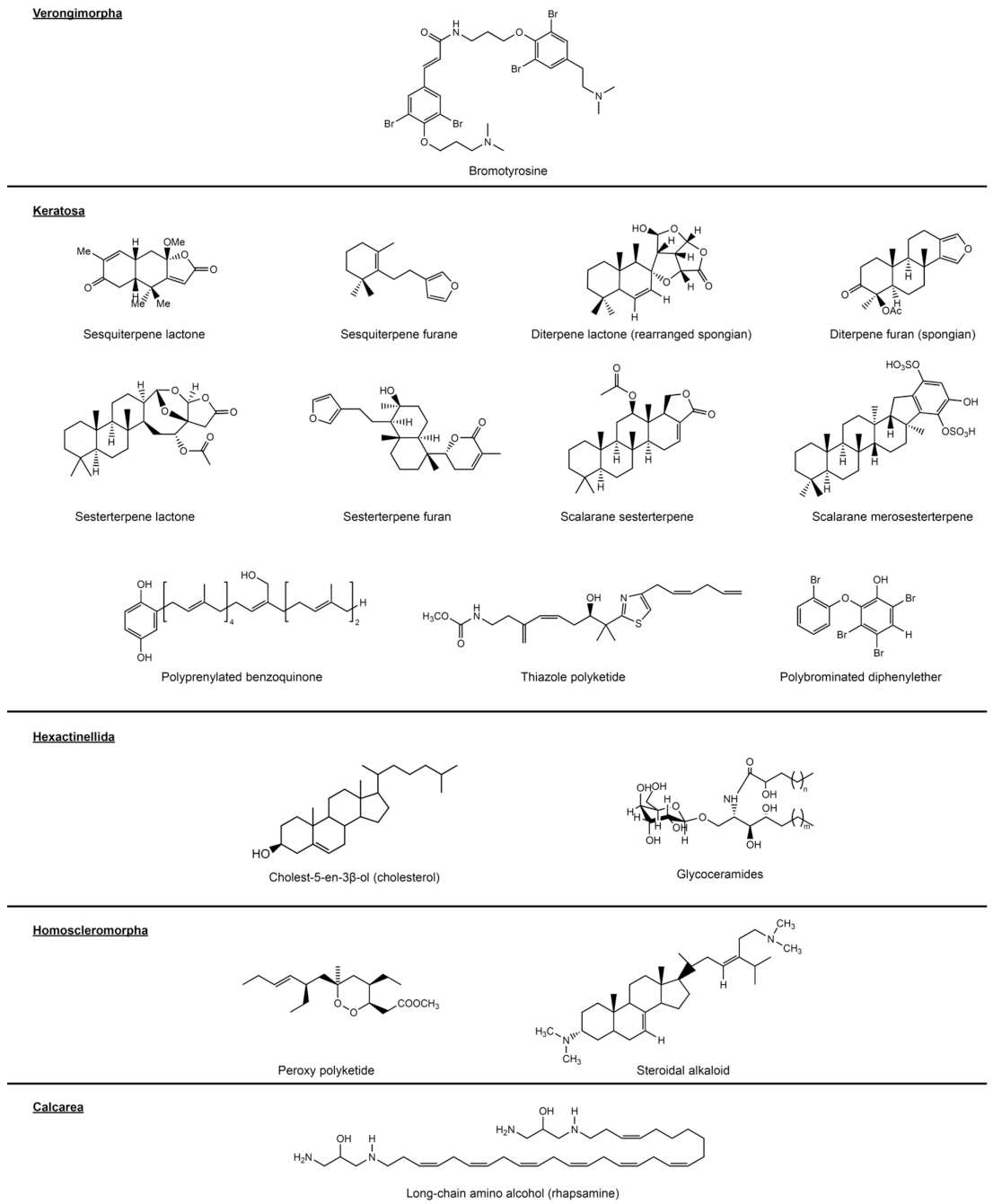
3.2.2. Verongimorpha and Keratosa
Verongiida (Verongimorpha)
3.2.3. Keratosa
3.3. Hexactinellida
3.4. Homoscleromorpha
3.5. Calcarea
4. The Legacy of Chemosystematics–Perspectives on Phylogenetics and Biochemistry
Author Contributions
Funding
Conflicts of Interest
References
- Gehling, J.G.; Rigby, J.K. Long Expected Sponges from the Neoproterozoic Ediacara Fauna of South Australia. J. Paleontol. 1996, 70, 185–195. [Google Scholar] [CrossRef]
- Sperling, E.A.; Robinson, J.M.; Pisani, D.; Peterson, K.J. Where’s the Glass? Biomarkers, Molecular Clocks, and microRNAs Suggest a 200-Myr Missing Precambrian Fossil Record of Siliceous Sponge Spicules. Geobiology 2010, 8, 24–36. [Google Scholar] [CrossRef]
- Schuster, A.; Vargas, S.; Knapp, I.S.; Pomponi, S.A.; Toonen, R.J.; Erpenbeck, D.; Wörheide, G. Divergence Times in Demosponges (Porifera): First Insights from New Mitogenomes and the Inclusion of Fossils in a Birth-Death Clock Model. BMC Evol. Biol. 2018, 18, 114. [Google Scholar] [CrossRef]
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schönberg, C.; et al. World Porifera Database. The World Register of Marine Species (WoRMS). 2021. Available online: http://www.marinespecies.org/porifera (accessed on 12 June 2021).
- Manconi, R.; Pronzato, R. Global Diversity of Sponges (Porifera: Spongillina) in Freshwater. Hydrobiologia 2008, 595, 27–33. [Google Scholar] [CrossRef]
- Woesz, A.; Weaver, J.C.; Kazanci, M.; Dauphin, Y.; Aizenberg, J.; Morse, D.E.; Fratzl, P. Micromechanical Properties of Biological Silica in Skeletons of Deep-Sea Sponges. J. Mater. Res. 2006, 21, 2068–2078. [Google Scholar] [CrossRef] [Green Version]
- Abbas, S.; Kelly, M.; Bowling, J.; Sims, J.; Waters, A.; Hamann, M. Advancement into the Arctic Region for Bioactive Sponge Secondary Metabolites. Mar. Drugs 2011, 9, 2423–2437. [Google Scholar] [CrossRef]
- Wiedenmayer, F. Shallow-Water Sponges of the Western Bahamas; Birkhäuser: Basel, Switzerland, 2013; Volume 28, ISBN 9783034857970. [Google Scholar]
- Krautter, M.; Conway, K.W.; Barrie, J.V.; Neuweiler, M. Discovery of a “Living Dinosaur”: Globally Unique Modern Hexactinellid Sponge Reefs off British Columbia, Canada. Facies 2001, 44, 265–282. [Google Scholar] [CrossRef]
- Leinfelder, R.; Werner, W.; Nose, M.; Schmid, D.; Krautter, M.; Laternser, R.; Takacs, M.; Hartmann, D. Paleoecology, Growth Parameters and Dynamics of Coral, Sponge and Microbolite Reefs from the Late Jurassic; Universität Stuttgart: Stuttgart, Germany, 1996. [Google Scholar]
- Diaz, M.C.; Rützler, K. Sponges: An Essential Component of Caribbean Coral Reefs. Bull. Mar. Sci. 2001, 69, 535–546. [Google Scholar]
- Bell, J.J. The Functional Roles of Marine Sponges. Estuar. Coast. Shelf Sci. 2008, 79, 341–353. [Google Scholar] [CrossRef]
- De Goeij, J.M.; van Oevelen, D.; Vermeij, M.J.A.; Osinga, R.; Middelburg, J.J.; De Goeij, A.F.P.M.; Admiraal, W. Surviving in a Marine Desert: The Sponge Loop Retains Resources within Coral Reefs. Science 2013, 342, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, M.; Pawlik, J.R. Spongivory by Parrotfish in Florida Mangrove and Reef Habitats. Mar. Ecol. 1998, 19, 325–337. [Google Scholar] [CrossRef]
- Furrow, F.B.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Surface Sequestration of Chemical Feeding Deterrents in the Antarctic Sponge Latrunculia apicalis as an Optimal Defense against Sea Star Spongivory. Mar. Biol. 2003, 143, 443–449. [Google Scholar] [CrossRef]
- Loh, T.-L.; Pawlik, J.R. Chemical Defenses and Resource Trade-Offs Structure Sponge Communities on Caribbean Coral Reefs. Proc. Natl. Acad. Sci. USA 2014, 111, 4151–4156. [Google Scholar] [CrossRef] [Green Version]
- López-Victoria, M.; Zea, S.; Weil, E. Competition for Space between Encrusting Excavating Caribbean Sponges and Other Coral Reef Organisms. Mar. Ecol. Prog. Ser. 2006, 312, 113–121. [Google Scholar] [CrossRef]
- Meylan, A. Spongivory in Hawksbill Turtles: A Diet of Glass. Science 1988, 239, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Elardo, S.M.; Kozytska, S.; Bugni, T.S.; Ireland, C.M.; Moll, H.; Hentschel, U. Anti-Parasitic Compounds from Streptomyces sp. Strains Isolated from Mediterranean Sponges. Mar. Drugs 2010, 8, 373–380. [Google Scholar] [CrossRef]
- Thoms, C.; Hentschel, U.; Schmitt, S.; Schupp, P.J. Rapid Tissue Reduction and Recovery in the Sponge Aplysinella sp. Mar. Biol. 2008, 156, 141–153. [Google Scholar] [CrossRef]
- Rohde, S.; Schupp, P.J. Growth and regeneration of the elephant ear sponge Ianthella basta (Porifera). In Ancient Animals, New Challenges: Developments in Sponge Research; Maldonado, M., Turon, X., Becerro, M., Jesús Uriz, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 219–226. ISBN 9789400746886. [Google Scholar]
- Borisenko, I.E.; Adamska, M.; Tokina, D.B.; Ereskovsky, A.V. Transdifferentiation Is a Driving Force of Regeneration in Halisarca dujardini (Demospongiae, Porifera). PeerJ 2015, 3, e1211. [Google Scholar] [CrossRef]
- Burns, E.; Ifrach, I.; Carmeli, S.; Pawlik, J.R.; Ilan, M. Comparison of Anti-Predatory Defenses of Red Sea and Caribbean Sponges. I. Chemical Defense. Mar. Ecol. Prog. Ser. 2003, 252, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.S.; Lopez, N.A.; Young, K.A. Anti-Predator Defenses in Western North Atlantic Sponges with Evidence of Enhanced Defense through Interactions between Spicules and Chemicals. Mar. Ecol. Prog. Ser. 2005, 291, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Unson, M.D.; Holland, N.D.; Faulkner, D.J. A Brominated Secondary Metabolite Synthesized by the Cyanobacterial Symbiont of a Marine Sponge and Accumulation of the Crystalline Metabolite in the Sponge Tissue. Mar. Biol. 1994, 119, 1–11. [Google Scholar] [CrossRef]
- Bewley, C.A.; Holland, N.D.; Faulkner, D.J. Two Classes of Metabolites from Theonella swinhoei Are Localized in Distinct Populations of Bacterial Symbionts. Experientia 1996, 52, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Thoms, C.; Schupp, P.J. Activated Chemical Defense in Marine Sponges—A Case Study on Aplysinella rhax. J. Chem. Ecol. 2008, 34, 1242–1252. [Google Scholar] [CrossRef]
- Rohde, S.; Schupp, P.J. Allocation of Chemical and Structural Defenses in the Sponge Melophlus sarasinorum. J. Exp. Mar. Bio. Ecol. 2011, 399, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, S.; Nietzer, S.; Schupp, P.J. Prevalence and Mechanisms of Dynamic Chemical Defenses in Tropical Sponges. PLoS ONE 2015, 10, e0132236. [Google Scholar] [CrossRef]
- Bergmann, W.; Feeney, R.J. The Isolation of a New Thymine Pentoside from Sponges. J. Am. Chem. Soc. 1950, 72, 2809–2810. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; An, R.; Wang, J.; Sun, N.; Zhang, S.; Hu, J.; Kuai, J. Exploring Novel Bioactive Compounds from Marine Microbes. Curr. Opin. Microbiol. 2005, 8, 276–281. [Google Scholar] [CrossRef]
- Wibowo, J.T.; Kellermann, M.Y.; Köck, M.; Putra, M.Y.; Murniasih, T.; Mohr, K.I.; Wink, J.; Praditya, D.F.; Steinmann, E.; Schupp, P.J. Anti-Infective and Antiviral Activity of Valinomycin and Its Analogues from a Sea Cucumber-Associated Bacterium, Streptomyces sp. SV 21. Mar. Drugs 2021, 19, 81. [Google Scholar] [CrossRef]
- Anjum, K.; Abbas, S.Q.; Shah, S.A.A.; Akhter, N.; Batool, S.; Hassan, S.S.U. Marine Sponges as a Drug Treasure. Biomol. Ther. 2016, 24, 347–362. [Google Scholar] [CrossRef] [Green Version]
- Munro, M.H.G.; Blunt, J.W.; Dumdei, E.J.; Hickford, S.J.H.; Lill, R.E.; Li, S.; Battershill, C.N.; Duckworth, A.R. The discovery and development of marine compounds with pharmaceutical potential. In Progress in Industrial Microbiology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 35, pp. 15–25. [Google Scholar]
- Sipkema, D.; Franssen, M.C.R.; Osinga, R.; Tramper, J.; Wijffels, R.H. Marine Sponges as Pharmacy. Mar. Biotechnol. 2005, 7, 142–162. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Perkins, M.V.; Zhang, W.; Franco, C.M.M. New Marine Natural Products from Sponges (Porifera) of the Order Dictyoceratida (2001 to 2012); a Promising Source for Drug Discovery, Exploration and Future Prospects. Biotechnol. Adv. 2016, 34, 473–491. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M.; Martens, D.; Wijffels, R.H. Towards Commercial Production of Sponge Medicines. Mar. Drugs 2009, 7, 787–802. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Thakur, N.L. Significance of Investigating Allelopathic Interactions of Marine Organisms in the Discovery and Development of Cytotoxic Compounds. Chem. Biol. Interact. 2016, 243, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Berne, S.; Kalauz, M.; Lapat, M.; Savin, L.; Janussen, D.; Kersken, D.; Ambrožič Avguštin, J.; Zemljič Jokhadar, Š.; Jaklič, D.; Gunde-Cimerman, N.; et al. Screening of the Antarctic Marine Sponges (Porifera) as a Source of Bioactive Compounds. Polar Biol. 2016, 39, 947–959. [Google Scholar] [CrossRef]
- Manilal, A.; Sujith, S.; Selvin, J.; Kiran, G.S.; Shakir, C.; Lipton, A.P. Antimicrobial Potential of Marine Organisms Collected from the Southwest Coast of India against Multiresistant Human and Shrimp Pathogens. Sci. Mar. 2010, 74, 287–296. [Google Scholar] [CrossRef]
- Sima, P.; Vetvicka, V. Bioactive Substances with Anti-Neoplastic Efficacy from Marine Invertebrates: Porifera and Coelenterata. World J. Clin. Oncol. 2011, 2, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, P.R. Chemotaxonomy of the Porifera: The Development and Current Status of the Field. Mar. Nat. Prod. Chem. Biol. Perspect. 1983, 5, 1–50. [Google Scholar]
- Van Soest, R.W.M.; Braekman, J.C. Chemosystematics of Porifera: A Review. Mem. Queensl. Mus. 1999, 44, 569–589. [Google Scholar]
- Erpenbeck, D.; van Soest, R.W.M. Status and Perspective of Sponge Chemosystematics. Mar. Biotechnol. 2007, 9, 2–19. [Google Scholar] [CrossRef]
- Wörheide, G.; Dohrmann, M.; Erpenbeck, D.; Larroux, C.; Maldonado, M.; Voigt, O.; Borchiellini, C.; Lavrov, D.V. Deep Phylogeny and Evolution of Sponges (phylum Porifera). Adv. Mar. Biol. 2012, 61, 1–78. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Crisp, M.D.; Driver, F.; Hooper, J.N.A.; Van Soest, R.W.M. Phylogenetic Relationships of the Family Axinellidae (Porifera: Demospongiae) Using Morphological and Molecular Data. Zool. Scr. 2000, 29, 169–198. [Google Scholar] [CrossRef]
- Erpenbeck, D.; Breeuwer, J.A.J.; Parra-Velandia, F.J.; van Soest, R.W.M. Speculation with Spiculation?—Three Independent Gene Fragments and Biochemical Characters versus Morphology in Demosponge Higher Classification. Mol. Phylogenet. Evol. 2006, 38, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, P.; Rapp, H.T. Disrupted Spiculogenesis in Deep-Water Geodiidae (Porifera, Demospongiae) Growing in Shallow Waters. Invertebr. Biol. 2013, 132, 173–194. [Google Scholar] [CrossRef]
- Maldonado, M.; Carmona, M.C.; Uriz, M.J.; Cruzado, A. Decline in Mesozoic Reef-Building Sponges Explained by Silicon Limitation. Nature 1999, 401, 785–788. [Google Scholar] [CrossRef]
- Genta-Jouve, G.; Thomas, O.P. Biosynthesis in Marine Sponges: The Radiolabelling Strikes Back. Phytochem. Rev. 2013, 12, 425–434. [Google Scholar] [CrossRef]
- Jordan, P.A.; Moore, B.S. Biosynthetic Pathway Connects Cryptic Ribosomally Synthesized Posttranslationally Modified Peptide Genes with Pyrroloquinoline Alkaloids. Cell Chem. Biol. 2016, 23, 1504–1514. [Google Scholar] [CrossRef]
- Bergquist, P.R. Sponge Chemistry: A Review. In Proceedings of the Biologie des Spongiaires—Colloques Internationaux du C.N.R.S.; Lévi, C., Boury-Esnault, N., Eds.; CNRS: Paris, France, 1979; pp. 382–393. [Google Scholar]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic Insights into the Marine Sponge Microbiome. Nat. Rev. Microbiol. 2012, 10, 641–654. [Google Scholar] [CrossRef]
- Thacker, R.W.; Freeman, C.J. Sponge–Microbe Symbioses: Recent Advances and New Directions. In Advances in Marine Biology; Academic Press: Waltham, MA, USA, 2012; Volume 62, pp. 57–111. [Google Scholar]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine Drugs from Sponge-Microbe Association—A Review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [Green Version]
- Al-Mourabit, A.; Zancanella, M.A.; Tilvi, S.; Romo, D. Biosynthesis, Asymmetric Synthesis, and Pharmacology, Including Cellular Targets, of the Pyrrole-2-Aminoimidazole Marine Alkaloids. Nat. Prod. Rep. 2011, 28, 1229–1260. [Google Scholar] [CrossRef] [PubMed]
- Tribalat, M.-A.; Marra, M.V.; McCormack, G.P.; Thomas, O.P. Does the Chemical Diversity of the Order Haplosclerida (Phylum Porifera: Class Demospongia) Fit with Current Taxonomic Classification? Planta Med. 2016, 82, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Morrow, C.; Cárdenas, P. Proposal for a Revised Classification of the Demospongiae (Porifera). Front. Zool. 2015, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erpenbeck, D.; Sutcliffe, P.; Cook, S.D.C.; Dietzel, A.; Maldonado, M.; van Soest, R.W.M.; Hooper, J.N.A.; Wörheide, G. Horny Sponges and Their Affairs: On the Phylogenetic Relationships of Keratose Sponges. Mol. Phylogenet. Evol. 2012, 63, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Erpenbeck, D.; Hall, K.; Alvarez, B.; Büttner, G.; Sacher, K.; Schätzle, S.; Schuster, A.; Vargas, S.; Hooper, J.N.A.; Wörheide, G. The Phylogeny of Halichondrid Demosponges: Past and Present Re-Visited with DNA-Barcoding Data. Org. Divers. Evol. 2012, 12, 57–70. [Google Scholar] [CrossRef]
- Gazave, E.; Carteron, S.; Chenuil, A.; Richelle-Maurer, E.; Boury-Esnault, N.; Borchiellini, C. Polyphyly of the Genus Axinella and of the Family Axinellidae (Porifera: Demospongiaep). Mol. Phylogenet. Evol. 2010, 57, 35–47. [Google Scholar] [CrossRef]
- Gutekunst, V.; MÜller, A.U.; Pohl, T.; BrÜmmer, F.; Malik, H.; Fawzi, N.; Erpenbeck, D.; Lehnert, H. A New Fistulose Demosponge Species from the Persian Gulf. Zootaxa 2018, 4450, 565–574. [Google Scholar] [CrossRef]
- Morrow, C.C.; Picton, B.E.; Erpenbeck, D.; Boury-Esnault, N.; Maggs, C.A.; Allcock, A.L. Congruence between Nuclear and Mitochondrial Genes in Demospongiae: A New Hypothesis for Relationships within the G4 Clade (Porifera: Demospongiae). Mol. Phylogenet. Evol. 2012, 62, 174–190. [Google Scholar] [CrossRef]
- Morrow, C.; Cárdenas, P.; Boury-Esnault, N.; Picton, B.; McCormack, G.; Van Soest, R.; Collins, A.; Redmond, N.; Maggs, C.; Sigwart, J.; et al. Integrating Morphological and Molecular Taxonomy with the Revised Concept of Stelligeridae (Porifera: Demospongiae). Zool. J. Linn. Soc. 2019, 187, 31–81. [Google Scholar] [CrossRef]
- Redmond, N.E.; Morrow, C.C.; Thacker, R.W.; Diaz, M.C.; Boury-Esnault, N.; Cárdenas, P.; Hajdu, E.; Lôbo-Hajdu, G.; Picton, B.E.; Pomponi, S.A.; et al. Phylogeny and Systematics of Demospongiae in Light of New Small-Subunit Ribosomal DNA (18S) Sequences. Integr. Comp. Biol. 2013, 53, 388–415. [Google Scholar] [CrossRef] [Green Version]
- Hooper, J.N.A.; Van Soest, R.W.M. Systema Porifera. A Guide to the Classification of Sponges. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; pp. 1–7. ISBN 9781461507475. [Google Scholar]
- Van Soest, R.W.M.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; De Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.A. Global Diversity of Sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef]
- Hooper, J.N.A.; Van Soest, R.W.M. Order Poecilosclerida Topsent, 1928. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; pp. 403–408. ISBN 9781461507475. [Google Scholar]
- Ibrahim, S.R.M.; Ebel, R.; Wray, V.; Müller, W.E.G.; Edrada-Ebel, R.; Proksch, P. Diacarperoxides, Norterpene Cyclic Peroxides from the Sponge Diacarnus megaspinorhabdosa. J. Nat. Prod. 2008, 71, 1358–1364. [Google Scholar] [CrossRef]
- Yang, F.; Zou, Y.; Wang, R.-P.; Hamann, M.T.; Zhang, H.-J.; Jiao, W.-H.; Han, B.-N.; Song, S.-J.; Lin, H.-W. Relative and Absolute Stereochemistry of Diacarperoxides: Antimalarial Norditerpene Endoperoxides from Marine Sponge Diacarnus megaspinorhabdosa. Mar. Drugs 2014, 12, 4399–4416. [Google Scholar] [CrossRef] [Green Version]
- Antunes, E.M.; Copp, B.R.; Davies-Coleman, M.T.; Samaai, T. Pyrroloiminoquinone and Related Metabolites from Marine Sponges. Nat. Prod. Rep. 2005, 22, 62–72. [Google Scholar] [CrossRef]
- Perry, N.B.; Blunt, J.W.; Munro, M.H.G.; Higa, T.; Sakai, R. Discorhabdin D, an Antitumor Alkaloid from the Sponges Latrunculia brevis and Prianos sp. J. Org. Chem. 1988, 53, 4127–4128. [Google Scholar] [CrossRef]
- Botić, T.; Defant, A.; Zanini, P.; Žužek, M.C.; Frangež, R.; Janussen, D.; Kersken, D.; Knez, Ž.; Mancini, I.; Sepčić, K. Discorhabdin Alkaloids from Antarctic Latrunculia spp. Sponges as a New Class of Cholinesterase Inhibitors. Eur. J. Med. Chem. 2017, 136, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Goey, A.K.L.; Chau, C.H.; Sissung, T.M.; Cook, K.M.; Venzon, D.J.; Castro, A.; Ransom, T.R.; Henrich, C.J.; McKee, T.C.; McMahon, J.B.; et al. Screening and Biological Effects of Marine Pyrroloiminoquinone Alkaloids: Potential Inhibitors of the HIF-1α/p300 Interaction. J. Nat. Prod. 2016, 79, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Peifer, C.; Janussen, D.; Tasdemir, D. New Discorhabdin Alkaloids from the Antarctic Deep-Sea Sponge Latrunculia biformis. Mar. Drugs 2019, 17, 439. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Kelly, M.; Tasdemir, D. Chemistry, Chemotaxonomy and Biological Activity of the Latrunculid Sponges (Order Poecilosclerida, Family Latrunculiidae). Mar. Drugs 2021, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Tabakmakher, K.M.; Denisenko, V.A.; Guzii, A.G.; Dmitrenok, P.S.; Dyshlovoy, S.A.; Lee, H.-S.; Makarieva, T.N. Monanchomycalin C, a New Pentacyclic Guanidine Alkaloid from the Far-Eastern Marine Sponge Monanchora pulchra. Nat. Prod. Commun. 2013, 8, 1399–1402. [Google Scholar] [CrossRef] [Green Version]
- Grkovic, T.; Blees, J.S.; Bayer, M.M.; Colburn, N.H.; Thomas, C.L.; Henrich, C.J.; Peach, M.L.; McMahon, J.B.; Schmid, T.; Gustafson, K.R. Tricyclic Guanidine Alkaloids from the Marine Sponge Acanthella cavernosa That Stabilize the Tumor Suppressor PDCD4. Mar. Drugs 2014, 12, 4593–4601. [Google Scholar] [CrossRef] [Green Version]
- Gros, E.; Al-Mourabit, A.; Martin, M.-T.; Sorres, J.; Vacelet, J.; Frederich, M.; Aknin, M.; Kashman, Y.; Gauvin-Bialecki, A. Netamines H–N, Tricyclic Alkaloids from the Marine Sponge Biemna laboutei and Their Antimalarial Activity. J. Nat. Prod. 2014, 77, 818–823. [Google Scholar] [CrossRef]
- Cárdenas, P.; Xavier, J.R.; Reveillaud, J.; Schander, C.; Rapp, H.T. Molecular Phylogeny of the Astrophorida (Porifera, Demospongiae) Reveals an Unexpected High Level of Spicule Homoplasy. PLoS ONE 2011, 6, e18318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, A.; Erpenbeck, D.; Pisera, A.; Hooper, J.; Bryce, M.; Fromont, J.; Wörheide, G. Deceptive Desmas: Molecular Phylogenetics Suggests a New Classification and Uncovers Convergent Evolution of Lithistid Demosponges. PLoS ONE 2015, 10, e116038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putri, S.P.; Kinoshita, H.; Ihara, F.; Igarashi, Y.; Nihira, T. Ophiosetin, a New Tetramic Acid Derivative from the Mycopathogenic Fungus Elaphocordyceps ophioglossoides. J. Antibiot. 2010, 63, 195–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, X.; Li, Q.; Ju, J. Naturally Occurring Tetramic Acid Products: Isolation, Structure Elucidation and Biological Activity. RSC Adv. 2014, 4, 50566–50593. [Google Scholar] [CrossRef]
- Wang, J.-F.; Qin, X.; Xu, F.-Q.; Zhang, T.; Liao, S.; Lin, X.; Yang, B.; Liu, J.; Wang, L.; Tu, Z.; et al. Tetramic Acid Derivatives and Polyphenols from Sponge-Derived Fungus and Their Biological Evaluation. Nat. Prod. Res. 2015, 29, 1761–1765. [Google Scholar] [CrossRef]
- Kamyab, E.; Kellermann, M.Y.; Kunzmann, A.; Schupp, P.J. Chemical Biodiversity and Bioactivities of Saponins in Echinodermata with an Emphasis on Sea Cucumbers (Holothuroidea). In YOUMARES 9-The Oceans: Our Research, Our Future; Springer: Cham, Switzerland, 2020; pp. 121–157. [Google Scholar]
- Kalinin, V.I.; Ivanchina, N.V.; Krasokhin, V.B.; Makarieva, T.N.; Stonik, V.A. Glycosides from Marine Sponges (Porifera, Demospongiae): Structures, Taxonomical Distribution, Biological Activities and Biological Roles. Mar. Drugs 2012, 10, 1671–1710. [Google Scholar] [CrossRef]
- Ivanchina, N.V.; Kicha, A.A.; Stonik, V.A. Steroid Glycosides from Marine Organisms. Steroids 2011, 76, 425–454. [Google Scholar] [CrossRef]
- Yeung, B.K.S.; Hamann, M.T.; Scheuer, P.J.; Kelly-Borges, M. Hapaioside: A 19-Norpregnane Glycoside from the Sponge Cribrochalina olemda. Tetrahedron 1994, 50, 12593–12598. [Google Scholar] [CrossRef]
- Campagnuolo, C.; Fattorusso, E.; Taglialatela-Scafati, O. Feroxosides A-B, Two Norlanostane Tetraglycosides from the Caribbean Sponge Ectyoplasia ferox. Tetrahedron 2001, 57, 4049–4055. [Google Scholar] [CrossRef]
- Regalado, E.L.; Jiménez-Romero, C.; Genta-Jouve, G.; Tasdemir, D.; Amade, P.; Nogueiras, C.; Thomas, O.P. Acanthifoliosides, Minor Steroidal Saponins from the Caribbean Sponge Pandaros acanthifolium. Tetrahedron 2011, 67, 1011–1018. [Google Scholar] [CrossRef]
- Dai, H.-F.; Edrada, R.A.; Ebel, R.; Nimtz, M.; Wray, V.; Proksch, P. Norlanostane Triterpenoidal Saponins from the Marine Sponge Melophlus sarassinorum. J. Nat. Prod. 2005, 68, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Antonov, A.S.; Kalinovsky, A.I.; Afiyatullov, S.S.; Leshchenko, E.V.; Dmitrenok, P.S.; Yurchenko, E.A.; Kalinin, V.I.; Stonik, V.A. Erylosides F8, V1–V3, and W–W2—New Triterpene Oligoglycosides from the Carribean Sponge Erylus goffrilleri. Carbohydr. Res. 2017, 449, 153–159. [Google Scholar] [CrossRef]
- Colorado, J.; Muñoz, D.; Marquez, D.; Marquez, M.; Lopez, J.; Thomas, O.; Martinez, A. Ulososides and Urabosides—Triterpenoid Saponins from the Caribbean Marine Sponge Ectyoplasia ferox. Molecules 2013, 18, 2598–2610. [Google Scholar] [CrossRef]
- Genta-Jouve, G.; Boughanem, C.; Ocaña, O.; Pérez, T.; Thomas, O.P. Eryloside W, a Triterpenoid Saponin from the Sponge Dictyonella marsilii. Phytochem. Lett. 2015, 13, 252–255. [Google Scholar] [CrossRef]
- Tang, S.; Pei, Y.; Fu, H.; Deng, Z.; Li, J.; Proksch, P.; Lin, W. Jaspolides A–F, Six New Isomalabricane-Type Terpenoids from the Sponge Jaspis sp. Chem. Pharm. Bull. 2006, 54, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.; Xu, R.; Lin, W.; Duan, H. Jaspiferin A and B: Two New Secondary Metabolites from the South China Sea Sponge Jaspis stellifera. Rec. Nat. Prod. 2012, 6, 398–401. [Google Scholar]
- García-Ruiz, C.; Sarabia, F. Chemistry and Biology of Bengamides and Bengazoles, Bioactive Natural Products from Jaspis Sponges. Mar. Drugs 2014, 12, 1580–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirirak, T. Chemical Constituents from the Sponge Pachastrissa nux. Ph.D. Thesis, Prince of Songkla University, Hat Yai, Thailand, 2012. [Google Scholar]
- White, K.N.; Tenney, K.; Crews, P. The Bengamides: A Mini-Review of Natural Sources, Analogues, Biological Properties, Biosynthetic Origins, and Future Prospects. J. Nat. Prod. 2017, 80, 740–755. [Google Scholar] [CrossRef]
- Ovenden, S.P.B.; Nielson, J.L.; Liptrot, C.H.; Willis, R.H.; Tapiolas, D.M.; Wright, A.D.; Motti, C.A. A New Diketopiperazine, Cyclo-(4-S-Hydroxy-R-Proline-R-Isoleucine), from an Australian Specimen of the Sponge Stelletta sp. Mar. Drugs 2011, 9, 2469–2478. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, H.; Cutignano, A.; Grauso, L.; Neng, N.; Cachatra, V.; Fontana, A.; Xavier, J.; Cerejo, M.; Vieira, H.; Santos, S. Erylusamides: Novel Atypical Glycolipids from Erylus cf. deficiens. Mar. Drugs 2016, 14, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farokhi, F.; Wielgosz-Collin, G.; Robic, A.; Debitus, C.; Malleter, M.; Roussakis, C.; Kornprobst, J.-M.; Barnathan, G. Antiproliferative Activity against Human Non-Small Cell Lung Cancer of Two O-Alkyl-Diglycosylglycerols from the Marine Sponges Myrmekioderma dendyi and Trikentrion laeve. Eur. J. Med. Chem. 2012, 49, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Wojnar, J.M.; Northcote, P.T. The Agminosides: Naturally Acetylated Glycolipids from the New Zealand Marine Sponge Raspailia agminata. J. Nat. Prod. 2011, 74, 69–73. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; Di Rosa, M.; Ianaro, A. Glycolipids from Sponges. Part 8:1 Plakopolyprenoside from the Marine Sponge Plakortis simplex. An Improved Procedure for Isolation of Glycolipids as Peracetyl Derivatives. Tetrahedron 2000, 56, 1393–1395. [Google Scholar] [CrossRef]
- Xiao, D.J.; Peng, X.D.; Deng, S.Z.; Ma, W.J.; Wu, H.M. Structure Elucidation of (3E)-Cholest-4-En-3, 6-Dione-3-Oxime in Marine Sponge Cinachyrella australiensis from the South China Sea. Chin. J. Org. Chem. 2005, 25, 1606–1609. [Google Scholar]
- Erpenbeck, D.; McCormack, G.P.; Breeuwer, J.A.J.; van Soest, R.W.M. Order Level Differences in the Structure of Partial LSU across Demosponges (Porifera): New Insights into an Old Taxon. Mol. Phylogenet. Evol. 2004, 32, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Nichols, S.A. An Evaluation of Support for Order-Level Monophyly and Interrelationships within the Class Demospongiae Using Partial Data from the Large Subunit rDNA and Cytochrome Oxidase Subunit I. Mol. Phylogenetics Evol. 2005, 34, 81–96. [Google Scholar] [CrossRef]
- McCormack, G.P.; Erpenbeck, D.; Van Soest, R.W.M. Major Discrepancy between Phylogenetic Hypotheses Based on Molecular and Morphological Criteria within the Order Haplosclerida (Phylum Porifera: Class Demospongiae). J. Zoolog. Syst. Evol. Res. 2002, 40, 237–240. [Google Scholar] [CrossRef]
- Andersen, R.J.; Van Soest, R.W.M.; Kong, F. 3-Alkylpiperidine alkaloids isolated from marine sponges in the order Haplosclerida. In Alkaloids: Chemical and Biological Perspectives; Pergamon: New York, NY, USA, 1996; Volume 10, pp. 301–355. ISBN 9780080526997. [Google Scholar]
- Jimenez, J.I.; Goetz, G.; Mau, C.M.S.; Yoshida, W.Y.; Scheuer, P.J.; Thomas Williamson, R.; Kelly, M. Upenamide: An Unprecedented Macrocyclic Alkaloid from the Indonesian Sponge Echinochalina sp. J. Org. Chem. 2000, 65, 8465–8469. [Google Scholar] [CrossRef]
- Mudianta, I.W.; Katavic, P.L.; Lambert, L.K.; Hayes, P.Y.; Banwell, M.G.; Munro, M.H.G.; Bernhardt, P.V.; Garson, M.J. Structure and Absolute Configuration of 3-Alkylpiperidine Alkaloids from an Indonesian Sponge of the Genus Halichondria. Tetrahedron 2010, 66, 2752–2760. [Google Scholar] [CrossRef]
- De Almeida Leone, P.; Carroll, A.R.; Towerzey, L.; King, G.; McArdle, B.M.; Kern, G.; Fisher, S.; Hooper, J.N.A.; Quinn, R.J. Exiguaquinol: A Novel Pentacyclic Hydroquinone from Neopetrosia exigua That Inhibits Helicobacter Pylori MurI. Org. Lett. 2008, 10, 2585–2588. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xu, T.; Yang, X.-W.; Huang, R.; Yang, B.; Tang, L.; Liu, Y. Chemical and Biological Aspects of Marine Sponges of the Genus Xestospongia. Chem. Biodivers. 2010, 7, 2201–2227. [Google Scholar] [CrossRef]
- Chamni, S.; Sirimangkalakitti, N.; Chanvorachote, P.; Saito, N.; Suwanborirux, K. Chemistry of Renieramycins. 17. A New Generation of Renieramycins: Hydroquinone 5-O-Monoester Analogues of Renieramycin M as Potential Cytotoxic Agents against Non-Small-Cell Lung Cancer Cells. J. Nat. 2017, 80, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Tianero, M.D.; Balaich, J.N.; Donia, M.S. Localized Production of Defence Chemicals by Intracellular Symbionts of Haliclona Sponges. Nat. Microbiol. 2019, 4, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-F.; Menna, M.; Cai, Y.-S.; Guo, Y.-W. Polyacetylenes of Marine Origin: Chemistry and Bioactivity. Chem. Rev. 2015, 115, 1543–1596. [Google Scholar] [CrossRef]
- Guella, G.; Mancini, I.; Chiasera, G.; Pietra, F. Rogiolenyne D, the Likely Immediate Precursor of Rogiolenyne A and B, Branched C15 Acetogenins Isolated from the Red Seaweed Laurencia microcladia of II Rogiolo. Conformation and Absolute Configuration in the Whole Series. Helv. Chim. Acta 1992, 75, 303–309. [Google Scholar] [CrossRef]
- Braekman, J.-C.; Daloze, D.; Stoller, C.; Van Soest, R.W.M. Chemotaxonomy of Agelas (Porifera: Demospongiae). Biochem. Syst. Ecol. 1992, 20, 417–431. [Google Scholar] [CrossRef] [Green Version]
- Erpenbeck, D.; Breeuwer, J.A.J.; Soest, R.W.M. Implications from a 28S rRNA Gene Fragment for the Phylogenetic Relationships of Halichondrid Sponges (Porifera: Demospongiae). J. Zoolog. Syst. Evol. Res. 2005, 43, 93–99. [Google Scholar] [CrossRef]
- Alvarez, B.; Hooper, J.N.A. Taxonomic Revision of the Order Halichondrida (Porifera: Demospongiae) of Northern Australia. Family Aictyonellidae. Beagle Rec. Mus. Art Galleries North. Territ. 2010, 26, 13. [Google Scholar]
- Marchais, S.; Al Mourabit, A.; Ahond, A.; Poupat, C.; Potier, P. A Short Synthesis of the Marine Bioactive Metabolite (+/−) Girolline. Tetrahedron Lett. 1998, 39, 8085–8088. [Google Scholar] [CrossRef]
- Williams, D.E.; Patrick, B.O.; Behrisch, H.W.; Van Soest, R.; Roberge, M.; Andersen, R.J. Dominicin, a Cyclic Octapeptide, and Laughine, a Bromopyrrole Alkaloid, Isolated from the Caribbean Marine Sponge Eurypon laughlini. J. Nat. Prod. 2005, 68, 327–330. [Google Scholar] [CrossRef]
- Haber, M.; Carbone, M.; Ilan, M.; Gavagnin, M. Structure of Debromo-Carteramine A, a Novel Bromopyrrole Alkaloid from the Mediterranean Sponge Axinella verrucosa. Arkivoc 2010, 2010, 233. [Google Scholar] [CrossRef] [Green Version]
- Sauleau, P.; Retailleau, P.; Nogues, S.; Carletti, I.; Marcourt, L.; Raux, R.; Mourabit, A.A.; Debitus, C. Dihydrohymenialdisines, New Pyrrole-2-Aminoimidazole Alkaloids from the Marine Sponge Cymbastela cantharella. Tetrahedron Lett. 2011, 52, 2676–2678. [Google Scholar] [CrossRef]
- Sauleau, P.; Moriou, C.; Al Mourabit, A. Metabolomics Approach to Chemical Diversity of the Mediterranean Marine Sponge Agelas oroides. Nat. Prod. Res. 2017, 31, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Morinaka, B.I.; Molinski, T.F. Structures and Solution Conformational Dynamics of Stylissamides G and H from the Bahamian Sponge Stylissa caribica. J. Nat. Prod. 2014, 77, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Hertiani, T.; Edrada-Ebel, R.; Ortlepp, S.; van Soest, R.W.M.; de Voogd, N.J.; Wray, V.; Hentschel, U.; Kozytska, S.; Müller, W.E.G.; Proksch, P. From Anti-Fouling to Biofilm Inhibition: New Cytotoxic Secondary Metabolites from Two Indonesian Agelas Sponges. Bioorg. Med. Chem. 2010, 18, 1297–1311. [Google Scholar] [CrossRef]
- Kubota, T.; Iwai, T.; Takahashi-Nakaguchi, A.; Fromont, J.; Gonoi, T.; Kobayashi, J. ’ichi Agelasines O–U, New Diterpene Alkaloids with a 9-N-Methyladenine Unit from a Marine Sponge Agelas sp. Tetrahedron 2012, 68, 9738–9744. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, M.; Chen, J.; Wang, H.; Tenney, K.; Crews, P. Bioactive Secondary Metabolites from the Marine Sponge Genus Agelas. Mar. Drugs 2017, 15, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erpenbeck, D.; van Soest, R.W.M. A Survey for Biochemical Synapomorphies to Reveal Phylogenetic Relationships of Halichondrid Demosponges (Metazoa: Porifera). Biochem. Syst. Ecol. 2005, 33, 585–616. [Google Scholar] [CrossRef]
- Gallimore, W.A.; Cabral, C.; Kelly, M.; Scheuer, P.J. A Novel D-Ring Unsaturated A-nor Sterol from the Indonesian Sponge, Axinella carteri Dendy. Nat. Prod. Res. 2008, 22, 1339–1343. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, V.; Byju, K.; Emilda, R.; Anu, G.; Nair, S.M.; Chandramohanakumar, N.; Peter, K.J.P.; Kumar, T.R.G.; Vasundhara, G. In Silico Biological Activity of Steroids from the Marine Sponge Axinella carteri. Med. Chem. Res. 2013, 22, 1142–1146. [Google Scholar] [CrossRef]
- Minale, L.; Sodano, G. Marine Sterols: 19-nor-Stanols from the Sponge Axinella polypoides. J. Chem. Soc. Perkin 1974, 1, 1888–1892. [Google Scholar] [CrossRef]
- Malik, S.; Djerassi, C. Minor and Trace Sterols in Marine Invertebrates. 61. Isolation and Structure Elucidation of New A-nor Sterols from the Marine Sponge Phakellia aruensis. Steroids 1989, 53, 271–284. [Google Scholar] [CrossRef]
- Marcos, I.S.; Moro, R.F.; Gil-Mesón, A.; Díez, D. 7-6-5 Tricarbocyclic Diterpenes: Valparanes, Mulinanes, Cyathanes, Homoverrucosanes, and Related Ones. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2016; Volume 48, pp. 137–207. [Google Scholar]
- Green, D.; Goldberg, I.; Stein, Z.; Ilan, M.; Kashman, Y. Cyanthiwigin AD, Novel Cytotoxic Diterpenes from the Sponge Epipolasis reiswigi. Nat. Prod. Lett. 1992, 1, 193–199. [Google Scholar] [CrossRef]
- Garson, M.J.; Simpson, J.S. Marine Isocyanides and Related Natural Products—Structure, Biosynthesis and Ecology. Nat. Prod. Rep. 2004, 21, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Emsermann, J.; Kauhl, U.; Opatz, T. Marine Isonitriles and Their Related Compounds. Mar. Drugs 2016, 14, 16. [Google Scholar] [CrossRef]
- Garson, M.J. Biosynthesis of the Novel Diterpene Isonitrile Diisocyanoadociane by a Marine Sponge of the Amphimedon Genus: Incorporation Studies with Sodium [14C] Cyanide and Sodium [2-14C] Acetate. J. Chem. Soc. Chem. Commun. 1986, 35–36. [Google Scholar] [CrossRef]
- Wright, A.D.; Lang-Unnasch, N. Diterpene Formamides from the Tropical Marine Sponge Cymbastela hooperi and Their Antimalarial Activity in Vitro. J. Nat. Prod. 2009, 72, 492–495. [Google Scholar] [CrossRef]
- Wratten, S.J.; Faulkner, D.J.; Hirotsu, K.; Clardy, J. Diterpenoid Isocyanides from the Marine Sponge Hymeniacidon amphilecta. Tetrahedron Lett. 1978, 19, 4345–4348. [Google Scholar] [CrossRef]
- Avilés, E.; Rodríguez, A.D. Monamphilectine A, a Potent Antimalarial β-Lactam from Marine Sponge Hymeniacidon sp.: Isolation, Structure, Semisynthesis, and Bioactivity. Org. Lett. 2010, 12, 5290–5293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinski, T.F.; Faulkner, D.J.; Van Duyne, G.D.; Clardy, J. Three New Diterpene Isonitriles from a Palauan Sponge of the Genus Halichondria. J. Org. Chem. 1987, 52, 3334–3337. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Murphy, P.T.; Wells, R.J.; Blount, J.F. New Diterpene Isocyanides from a Sponge. Tetrahedron Lett. 1980, 21, 315–318. [Google Scholar] [CrossRef]
- Avilés, E.; Rodríguez, A.D.; Vicente, J. Two Rare-Class Tricyclic Diterpenes with Antitubercular Activity from the Caribbean Sponge Svenzea flava. Application of Vibrational Circular Dichroism Spectroscopy for Determining Absolute Configuration. J. Org. Chem. 2013, 78, 11294–11301. [Google Scholar] [CrossRef] [Green Version]
- Chanthathamrongsiri, N.; Yuenyongsawad, S.; Wattanapiromsakul, C.; Plubrukarn, A. Bifunctionalized Amphilectane Diterpenes from the Sponge Stylissa cf. massa. J. Nat. Prod. 2012, 75, 789–792. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Gavagnin, M.; Manzo, E.; Puliti, R.; Mattia, C.A.; Mazzarella, L.; Cimino, G.; Simpson, J.S.; Garson, M.J. Structural and Stereochemical Revision of Isocyanide and Isothiocyanate Amphilectenes from the Caribbean Marine Sponge Cribochalina sp. Tetrahedron 2005, 61, 8049–8053. [Google Scholar] [CrossRef]
- Jumaryatno, P.; Rands-Trevor, K.; Blanchfield, J.T.; Garson, M.J. Isocyanates in Marine Sponges: Axisocyanate-3, a New Sesquiterpene from Acanthella cavernosa. Arkivoc 2007, 2007, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Sorek, H.; Zelikoff, A.L.; Benayahu, Y.; Kashman, Y. Axiplyns A–E, New Sesquiterpene Isothiocyanates from the Marine Sponge Axinyssa aplysinoides. Tetrahedron Lett. 2008, 49, 2200–2203. [Google Scholar] [CrossRef]
- Fattorusso, E.; Magno, S.; Mayol, L.; Santacroce, C.; Sica, D. New Sesquiterpenoids from the Sponge Axinella cannabina. Tetrahedron 1975, 31, 269–270. [Google Scholar] [CrossRef]
- Ishiyama, H.; Hashimoto, A.; Fromont, J.; Hoshino, Y.; Mikami, Y.; Kobayashi, J. Halichonadins A–D, New Sesquiterpenoids from a Sponge Halichondria sp. Tetrahedron 2005, 61, 1101–1105. [Google Scholar] [CrossRef]
- Karuso, P.; Poiner, A.; Scheuer, P.J. Isocyanoneopupukeanane, a New Tricyclic Sesquiterpene from a Sponge. J. Org. Chem. 1989, 54, 2095–2097. [Google Scholar] [CrossRef]
- Nakamura, H.; Deng, S.; Takamatsu, M.; Kobayashi, J.; Ohizumi, Y.; Hirata, Y. Structure of Halipanicine, a New Sesquiterpene Isothiocyanate from the Okinawan Marine Sponge Halichondria panicea (Pallas). Agric. Biol. Chem. 1991, 55, 581–583. [Google Scholar] [CrossRef]
- Kassuhlke, K.E.; Potts, B.C.M.; Faulkner, D.J. New Nitrogenous Sesquiterpenes from Two Philippine Nudibranchs, Phyllidia pustulosa and P. varicosa, and from a Palauan Sponge, Halichondria cf. lendenfeldi. J. Org. Chem. 1991, 56, 3747–3750. [Google Scholar] [CrossRef]
- Kondempudi, C.M.; Singanaboina, R.; Manchala, N.; Gunda, V.G.; Janapala, V.R.; Yenamandra, V. Chemical Examination of the Sponge Phycopsis sp. Chem. Pharm. Bull. 2009, 57, 990–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, H.; Kobayashi, J.; Ohizumi, Y.; Mitsubishi-Kasei; Hirata, Y. Novel Bisabolene-Type Sesquiterpenoids with a Conjugated Diene Isolated from the Okinawan Sea Sponge Theonella cf. swinhoei. Tetrahedron Lett. 1984, 25, 5401–5404. [Google Scholar] [CrossRef]
- Cui, J.; Shang, R.-Y.; Sun, M.; Li, Y.-X.; Liu, H.-Y.; Lin, H.-W.; Jiao, W.-H. Trichodermaloids A-C, Cadinane Sesquiterpenes from a Marine Sponge Symbiotic Trichoderma sp. SM16 Fungus. Chem. Biodivers. 2020, 17, e2000036. [Google Scholar] [CrossRef] [PubMed]
- Wratten, S.J.; Faulkner, D.J. Carbonimidic Dichlorides from the Marine Sponge Pseudaxinyssa pitys. J. Am. Chem. Soc. 1977, 99, 7367–7368. [Google Scholar] [CrossRef]
- Hirota, H.; Okino, T.; Yoshimura, E.; Fusetani, N. Five New Antifouling Sesquiterpenes from Two Marine Sponges of the Genus Axinyssa and the Nudibranch Phyllidia pustulosa. Tetrahedron 1998, 54, 13971–13980. [Google Scholar] [CrossRef]
- Simpson, J.S.; Raniga, P.; Garson, M.J. Biosynthesis of Dichloroimines in the Tropical Marine Sponge Stylotella aurantium. Tetrahedron Lett. 1997, 38, 7947–7950. [Google Scholar] [CrossRef]
- Kehraus, S.; König, G.M.; Wright, A.D. New Carbonimidic Dichlorides from the Australian Sponge Ulosa spongia and Their Possible Taxonomic Significance. J. Nat. Prod. 2001, 64, 939–941. [Google Scholar] [CrossRef]
- Larghi, E.L.; Bohn, M.L.; Kaufman, T.S. Aaptamine and Related Products. Their Isolation, Chemical Syntheses, and Biological Activity. Tetrahedron 2009, 65, 4257–4282. [Google Scholar] [CrossRef]
- Utkina, N.K.; Denisenko, V.A. N-Demethylaaptanone, A New Congener of Aaptamine Alkaloids from the Vietnamese Marine Sponge Aaptos aaptos. Nat. Prod. Commun. 2016, 11, 1259–1260. [Google Scholar] [CrossRef] [Green Version]
- Gan, J.-H.; Hu, W.-Z.; Yu, H.-B.; Yang, F.; Cao, M.-X.; Shi, H.-J.; Kang, Y.-F.; Han, B.-N. Three New Aaptamine Derivatives from the South China Sea Sponge Aaptos aaptos. J. Asian Nat. Prod. Res. 2015, 17, 1231–1238. [Google Scholar] [CrossRef]
- Díaz-Marrero, A.R.; Brito, I.; Cueto, M.; San-Martín, A.; Darias, J. Suberitane Network, a Taxonomical Marker for Antarctic Sponges of the Genus Suberites? Novel Sesterterpenes from Suberites Caminatus. Tetrahedron Lett. 2004, 45, 4707–4710. [Google Scholar] [CrossRef]
- Solanki, H.; Angulo-Preckler, C.; Calabro, K.; Kaur, N.; Lasserre, P.; Cautain, B.; de la Cruz, M.; Reyes, F.; Avila, C.; Thomas, O.P. Suberitane Sesterterpenoids from the Antarctic Sponge Phorbas areolatus (Thiele, 1905). Tetrahedron Lett. 2018, 59, 3353–3356. [Google Scholar] [CrossRef]
- Daoust, J.; Chen, M.; Wang, M.; Williams, D.E.; Garcia Chavez, M.A.; Wang, Y.A.; Merchant, C.E.; Fontana, A.; Kieffer, T.J.; Andersen, R.J. Sesterterpenoids Isolated from a Northeastern Pacific Phorbas sp. J. Org. Chem. 2013, 78, 8267–8273. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, R.W.M.; Hooper, J.N.A. Order Haplosclerida Topsent, 1928. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; pp. 831–832. ISBN 9781461507475. [Google Scholar]
- Abou-Shoer, M.I.; Shaala, L.A.; Youssef, D.T.A.; Badr, J.M.; Habib, A.-A.M. Bioactive Brominated Metabolites from the Red Sea Sponge Suberea mollis. J. Nat. Prod. 2008, 71, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Davis, R.A.; Shelper, T.; Sykes, M.L.; Avery, V.M.; Elofsson, M.; Sundin, C.; Quinn, R.J. Pseudoceramines A–D, New Antibacterial Bromotyrosine Alkaloids from the Marine Sponge Pseudoceratina sp. Org. Biomol. Chem. 2011, 9, 6755–6760. [Google Scholar] [CrossRef]
- Peng, J.; Li, J.; Hamann, M.T. The Marine Bromotyrosine Derivatives. Alkaloids Chem. Biol. 2005, 61, 59–262. [Google Scholar] [CrossRef] [Green Version]
- Bergquist, P.R. A Revision of the Supraspecific Classification of the Orders Dictyoceratida, Dendroceratida, and Verongida (class Demospongiae). N. Z. J. Zool. 1980, 7, 443–503. [Google Scholar] [CrossRef] [Green Version]
- Erpenbeck, D.; Ekins, M.; Enghuber, N.; Hooper, J.N.A.; Lehnert, H.; Poliseno, A.; Schuster, A.; Setiawan, E.; De Voogd, N.J.; Wörheide, G.; et al. Nothing in (sponge) Biology Makes Sense—Except When Based on Holotypes. J. Mar. Biol. Assoc. UK 2016, 96, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Bergquist, P.R.; de Cook, S.C. Order Dendroceratida Minchin, 1900. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; p. 1067. ISBN 9781461507475. [Google Scholar]
- De Cook, S.C.; Bergquist, P.R. Order Dictyoceratida Minchin, 1900. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; p. 1021. ISBN 9781461507475. [Google Scholar]
- Erpenbeck, D.; Galitz, A.; Ekins, M.; Cook, S.D.C.; Soest, R.W.M.; Hooper, J.N.A.; Wörheide, G. Soft Sponges with Tricky Tree: On the Phylogeny of Dictyoceratid Sponges. J. Zoolog. Syst. Evol. Res. 2020, 58, 27–40. [Google Scholar] [CrossRef] [Green Version]
- De Cook, S.C.; Bergquist, P.R. Family Irciniidae Gray, 1867. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; pp. 1022–1027. ISBN 9781461507475. [Google Scholar]
- Gonzalez, M.A. Spongiane Diterpenoids. Curr. Bioact. Compd. 2007, 3, 1–36. [Google Scholar] [CrossRef]
- Wojnar, J.M.; Dowle, K.O.; Northcote, P.T. The Oxeatamides: Nitrogenous Spongian Diterpenes from the New Zealand Marine Sponge Darwinella oxeata. J. Nat. Prod. 2014, 77, 2288–2295. [Google Scholar] [CrossRef]
- Williams, D.E.; Marques, S.O.; Hajdu, E.; Peixinho, S.; Andersen, R.J.; Berlinck, R.G.S. Pyrodysinoic Acid Derivatives from the Marine Sponge Dysidea robusta. J. Nat. Prod. 2009, 72, 1691–1694. [Google Scholar] [CrossRef]
- Nguyen, X.N.; Nguyen, T.C.; Dan, T.T.H.; Do, T.T.; Nguyen, H.N.; Pham, H.Y.; Do, C.T.; Vu, K.T.; Hoang, L.T.A.; Bui, H.T.; et al. 1H and 13C NMR Assignments of Sesquiterpenes from Dysidea fragilis. Magn. Reson. Chem. 2015, 53, 1057–1060. [Google Scholar] [CrossRef]
- Kato, Y.; Fusetani, N.; Matsunaga, S.; Hashimoto, K. Spongionellin and Dehydrospongionellin, New Furanosesterterpenes Which Inhibit Cell Division of Fertilized Starfish Eggs, from the Marine Sponge Spongionella sp. Chem. Lett. 1985, 14, 1521–1524. [Google Scholar] [CrossRef]
- Kato, Y.; Fusetani, N.; Matsunaga, S.; Hashimoto, K. Okinonellins A and B, Two Novel Furanosesterterpenes, Which Inhibit Cell Division of Fertilized Starfish Eggs, from the Marine sponge Spongionella sp. Experientia 1986, 42, 1299–1300. [Google Scholar] [CrossRef]
- Liu, G.; Pika, J.; Faulkner, D.J. A Sesterterpene from the Palauan Sponge Igernella sp. Nat. Prod. Lett. 1995, 7, 297–301. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Rho, J.-R.; Lee, H.-S.; Santalova, E.A.; Stonik, V.; Shin, J. New Sesterterpene Sulfates from the Sponge Darwinella australensis. J. Nat. Prod. 2003, 66, 1010–1012. [Google Scholar] [CrossRef]
- Cimino, G.; De Luca, P.; De Stefano, S.; Minale, L. Disidein, a Pentacyclic Sesterterpene Condensed with an Hydroxyhydroquinone Moiety, from the Sponge Dysidea pallescens. Tetrahedron 1975, 31, 271–275. [Google Scholar] [CrossRef]
- Cimino, G.; De Rosa, S.; De Stefano, S.; Puliti, R.; Strazzullo, G.; Mattia, C.A.; Mazzarella, L. Absolute Stereochemistry of Disidein and of Two New Related Halogenated Sesterterpenoids. Two-Dimensional Nmr Studies and X-ray Crystal Structure. Tetrahedron 1987, 43, 4777–4784. [Google Scholar] [CrossRef]
- Elyakov, G.B.; Kuznetsova, T.; Mikhailov, V.V.; Maltsev, I.I.; Voinov, V.G.; Fedoreyev, S.A. Brominated Diphenyl Ethers from a Marine Bacterium Associated with the Sponge Dysidea sp. Experientia 1991, 47, 632–633. [Google Scholar] [CrossRef]
- Fan, L.; Reynolds, D.; Liu, M.; Stark, M.; Kjelleberg, S.; Webster, N.S.; Thomas, T. Functional Equivalence and Evolutionary Convergence in Complex Communities of Microbial Sponge Symbionts. Proc. Natl. Acad. Sci. USA 2012, 109, 1878–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Blanton, J.M.; Podell, S.; Taton, A.; Schorn, M.A.; Busch, J.; Lin, Z.; Schmidt, E.W.; Jensen, P.R.; Paul, V.J.; et al. Metagenomic Discovery of Polybrominated Diphenyl Ether Biosynthesis by Marine Sponges. Nat. Chem. Biol. 2017, 13, 537–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.A. Scalarane Sesterterpenoids. Curr. Bioact. Compd. 2010, 6, 178–206. [Google Scholar] [CrossRef] [Green Version]
- Tziveleka, L.-A.; Abatis, D.; Paulus, K.; Bauer, R.; Vagias, C.; Roussis, V. Marine Polyprenylated Hydroquinones, Quinones, and Chromenols with Inhibitory Effects on Leukotriene Formation. Chem. Biodivers. 2005, 2, 901–909. [Google Scholar] [CrossRef]
- Lee, H.-S.; Lee, Y.-J.; Lee, J.W. Identification of New Polyprenyl Hydroquinone Derivatives from Tropical Marine Sponge Ircinia sp. Heterocycles 2012, 85, 1437–1446. [Google Scholar] [CrossRef]
- Esposito, G.; Della Sala, G.; Teta, R.; Caso, A.; Bourguet-Kondracki, M.-L.; Pawlik, J.R.; Mangoni, A.; Costantino, V. Chlorinated Thiazole-Containing Polyketide-Peptides from the Caribbean Sponge Smenospongia conulosa: Structure Elucidation on Microgram Scale. Eur. J. Org. Chem. 2016, 2016, 2871–2875. [Google Scholar] [CrossRef]
- Kotoku, N.; Ishida, R.; Matsumoto, H.; Arai, M.; Toda, K.; Setiawan, A.; Muraoka, O.; Kobayashi, M. Biakamides A–D, Unique Polyketides from a Marine Sponge, Act as Selective Growth Inhibitors of Tumor Cells Adapted to Nutrient Starvation. J. Org. Chem. 2017, 82, 1705–1718. [Google Scholar] [CrossRef]
- Sonnenschein, R.N.; Johnson, T.A.; Tenney, K.; Valeriote, F.A.; Crews, P. A Reassignment of (-)-Mycothiazole and the Isolation of a Related Diol. J. Nat. Prod. 2006, 69, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Reiswig, H.M. Class Hexactinellida Schmidt, 1870. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Willenz, P., Eds.; Springer: Boston, MA, USA, 2002; pp. 1201–1210. ISBN 9781461507475. [Google Scholar]
- Blumenberg, M.; Thiel, V.; Pape, T.; Michaelis, W. The Steroids of Hexactinellid Sponges. Naturwissenschaften 2002, 89, 415–419. [Google Scholar] [CrossRef]
- Núñez-Pons, L.; Carbone, M.; Paris, D.; Melck, D.; Ríos, P.; Cristobo, J.; Castelluccio, F.; Gavagnin, M.; Avila, C. Chemo-Ecological Studies on Hexactinellid Sponges from the Southern Ocean. Naturwissenschaften 2012, 99, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Blumenberg, M.; Hefter, J.; Pape, T.; Pomponi, S.; Reed, J.; Reitner, J.; Wörheide, G.; Michaelis, W. A Chemical View of the Most Ancient Metazoa—Biomarker Chemotaxonomy of Hexactinellid Sponges. Naturwissenschaften 2002, 89, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boute, N.; Exposito, J.Y.; Boury-Esnault, N.; Vacelet, J.; Noro, N.; Miyazaki, K.; Yoshizato, K.; Garrone, R. Type IV Collagen in Sponges, the Missing Link in Basement Membrane Ubiquity. Biol. Cell 1996, 88, 37–44. [Google Scholar] [CrossRef]
- Gazave, E.; Lapébie, P.; Ereskovsky, A.V.; Vacelet, J.; Renard, E.; Cárdenas, P.; Borchiellini, C. No longer Demospongiae: Homoscleromorpha formal nomination as a fourth class of Porifera. In Ancient Animals, New Challenges: Developments in Sponge Research; Maldonado, M., Turon, X., Becerro, M., Jesús Uriz, M., Eds.; Springer: Dordrecht, The Netherland, 2012; pp. 3–10. ISBN 9789400746886. [Google Scholar]
- Uriz, M.-J. Mineral Skeletogenesis in Sponges. Can. J. Zool. 2006, 84, 322–356. [Google Scholar] [CrossRef]
- Sunassee, S.N.; Ransom, T.; Henrich, C.J.; Beutler, J.A.; Covell, D.G.; McMahon, J.B.; Gustafson, K.R. Steroidal Alkaloids from the Marine Sponge Corticium niger That Inhibit Growth of Human Colon Carcinoma Cells. J. Nat. Prod. 2014, 77, 2475–2480. [Google Scholar] [CrossRef] [Green Version]
- Norris, M.D.; Perkins, M.V. Structural Diversity and Chemical Synthesis of Peroxide and Peroxide-Derived Polyketide Metabolites from Marine Sponges. Nat. Prod. Rep. 2016, 33, 861–880. [Google Scholar] [CrossRef] [Green Version]
- Jumaryatno, P.; Lambert, L.K.; Hooper, J.N.A.; Blanchfield, J.T.; Garson, M.J. Cyclic Peroxides from a Two-Sponge Association of Plakortis communis–Agelas mauritiana. Nat. Prod. Commun. 2013, 8, 725–728. [Google Scholar] [CrossRef] [Green Version]
- Hagemann, A.; Voigt, O.; Wörheide, G.; Thiel, V. The Sterols of Calcareous Sponges (Calcarea, Porifera). Chem. Phys. Lipids 2008, 156, 26–32. [Google Scholar] [CrossRef]
- Cheney, K.L.; White, A.; Mudianta, I.W.; Winters, A.E.; Quezada, M.; Capon, R.J.; Mollo, E.; Garson, M.J. Choose Your Weaponry: Selective Storage of a Single Toxic Compound, Latrunculin A, by Closely Related Nudibranch Molluscs. PLoS ONE 2016, 11, e0145134. [Google Scholar] [CrossRef] [Green Version]
- Habener, L.J.; Hooper, J.N.A.; Carroll, A.R. Chemical and Biological Aspects of Marine Sponges from the Family Mycalidae. Planta Med. 2016, 82, 816–831. [Google Scholar] [CrossRef] [Green Version]
- Paul, V.J.; Freeman, C.J.; Agarwal, V. Chemical Ecology of Marine Sponges: New Opportunities through “-Omics”. Integr. Comp. Biol. 2019, 59, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Schupp, P.J.; Dahllöf, I.; Kjelleberg, S.; Steinberg, P.D. Host Specificity in Marine Sponge-Associated Bacteria, and Potential Implications for Marine Microbial Diversity. Environ. Microbiol. 2004, 6, 121–130. [Google Scholar] [CrossRef]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Björk, J.R.; Easson, C.; Astudillo-García, C.; Olson, J.B.; Erwin, P.M.; López-Legentil, S.; Luter, H.; et al. Diversity, Structure and Convergent Evolution of the Global Sponge Microbiome. Nat. Commun. 2016, 7, 11870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moitinho-Silva, L.; Nielsen, S.; Amir, A.; Gonzalez, A.; Ackermann, G.L.; Cerrano, C.; Astudillo-Garcia, C.; Easson, C.; Sipkema, D.; Liu, F.; et al. Erratum to: The Sponge Microbiome Project. Gigascience 2018, 7, giy145. [Google Scholar] [CrossRef] [PubMed]
- Steinert, G.; Wemheuer, B.; Janussen, D.; Erpenbeck, D.; Daniel, R.; Simon, M.; Brinkhoff, T.; Schupp, P.J. Prokaryotic Diversity and Community Patterns in Antarctic Continental Shelf Sponges. Front. Mar. Sci. 2019, 6, 297. [Google Scholar] [CrossRef]
- Kind, T.; Fiehn, O. Strategies for Dereplication of Natural Compounds Using High-Resolution Tandem Mass Spectrometry. Phytochem. Lett. 2017, 21, 313–319. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Quinn, R.A.; Nothias, L.-F.; Vining, O.; Meehan, M.; Esquenazi, E.; Dorrestein, P.C. Molecular Networking As a Drug Discovery, Drug Metabolism, and Precision Medicine Strategy. Trends Pharmacol. Sci. 2017, 38, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.-E.; Kellermann, M.Y.; Schupp, P.J. Secondary Metabolites of Marine Microbes: From Natural Products Chemistry to Chemical Ecology. In YOUMARES 9-The Oceans: Our Research, Our Future; Springer: Cham, Switzerland, 2020; pp. 159–180. [Google Scholar]
- Reverter, M.; Rohde, S.; Parchemin, C.; Tapissier-Bontemps, N.; Schupp, P.J. Metabolomics and Marine Biotechnology: Coupling Metabolite Profiling and Organism Biology for the Discovery of New Compounds. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Class | Taxonomic Group | 1998 | 2004 | 2020 |
|---|---|---|---|---|
| Pyrroloquinoline alkaloids | Poecilosclerida | O | + | + |
| Norditerpene peroxides | Podospongiidae (Poecilosclerida) | n.a. | n.a. | + |
| Norsesterterpene peroxides | Poecilosclerida | O | + | + |
| Pentacyclic guanidine alkaloids | Monanchora (Poecilosclerida) | + | + | + |
| Tetramic acids | Tetractinellida | n.a. | O | + |
| Steroidal saponins/glycosides | Tetractinellida | O | - | O |
| Isomalabaricane triterpenoids | Astrophorina (Tetractinellida) | + | + | + |
| Bengamide and bengazoles | Ancorinidae (Tetractinellida) | n.a. | O | + |
| Hydroxyiminosterols | Cinachyrella (Tetractinellida) | n.a. | + | + |
| Azetidine alkaloids | Penares (Tetractinellida) | + | + | + |
| 3-Alkylpiperidines + 3-Alkylpyridines | Haplosclerida | + | - | + |
| Renieramycins | Haplosclerida | O | O | + |
| Straight-chain polyacetylenes | Haplosclerida | + | O | - |
| Pentacyclic hydroquinones | Petrosiidae (Haplosclerida) | n.a. | n.a. | + |
| 3β-Hydroxymethyl-A-nor-sterols | Axinellida | n.a. | + | + |
| Cyanthiwigin diterpenes | Myrmekioderma (Axinellida) | O | + | + |
| Diterpene iso/thio/cyanides + formamides | Bubarida | O | O | + |
| Sesquiterpene iso/thio/cyanides + formamides | Bubarida | O | O | + |
| Carbonimidic dichlorides | Formerly Halichondrida | n.a. | O | - |
| Aaptamines | Suberitida | + | - | - |
| Suberitane-derived sesterterpenes | Suberitida | n.a. | + | - |
| Pyrrole-2-aminoimidazole alkaloids | Agelasida | + | + | + |
| Adenine-derivatives of diterpenes | Agelas (Agelasida) | n.a. | n.a. | + |
| Hypotaurocyamine (Sesquiterp. derivatives) | Agelas (Agelasida) | + | + | + |
| Bromotyrosines | Verongiida | + | - | + |
| Sesquiterpene lactones/furans | Dysideidae (Dictyoceratida) | O | - | + |
| Diterpene lactones/furans | Dendroceratida + Dictyoceratida | O | + | + |
| Sesterterpene lactones/furans | Spongiidae, Thorectidae, Irciniidae (Dictyoceratida) | O | - | + |
| Scalarane sesterterpenes | Spongiidae, Thorectidae, Irciniidae (Dictyoceratida) | n.a. | + | + |
| Scalarane sesterterpene hydroquinones | Dysidea + Acanthodendrilla (Dictyoceratida) | n.a. | n.a. | + |
| Polyprenylated benzoquinones | Irciniidae (Dictyoceratida) | n.a. | n.a. | + |
| Thiazole polyketides | Thorectidae (Dictyoceratida) | n.a. | n.a. | + |
| Polybrominated diphenyl ethers | Dysideidae (symbiotic origin) (Dictyoceratida) | n.a. | - | + |
| Cholest-5-en-3β-ol/5α(H)-cholestan-3β-ol | Hexactinellida | n.a. | + | + |
| Glycoceramides | Hexactinellida | n.a. | n.a. | + |
| Peroxy-Polyketides | Plakortis + Plakinastrella (Homoscleromorpha) | O | - | O |
| Steroidal alkaloids | Plakina + Corticium (Homoscleromorpha) | + | + | + |
| C27 to C29Δ5,7,22 & C27 to C29Δ5,7,9(11),22 sterols | Calcarea | n.a. | n.a. | - |
| Long-chain aminoalcohols | Clathrinida (Calcarea) | + | O | O |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galitz, A.; Nakao, Y.; Schupp, P.J.; Wörheide, G.; Erpenbeck, D. A Soft Spot for Chemistry–Current Taxonomic and Evolutionary Implications of Sponge Secondary Metabolite Distribution. Mar. Drugs 2021, 19, 448. https://doi.org/10.3390/md19080448
Galitz A, Nakao Y, Schupp PJ, Wörheide G, Erpenbeck D. A Soft Spot for Chemistry–Current Taxonomic and Evolutionary Implications of Sponge Secondary Metabolite Distribution. Marine Drugs. 2021; 19(8):448. https://doi.org/10.3390/md19080448
Chicago/Turabian StyleGalitz, Adrian, Yoichi Nakao, Peter J. Schupp, Gert Wörheide, and Dirk Erpenbeck. 2021. "A Soft Spot for Chemistry–Current Taxonomic and Evolutionary Implications of Sponge Secondary Metabolite Distribution" Marine Drugs 19, no. 8: 448. https://doi.org/10.3390/md19080448
APA StyleGalitz, A., Nakao, Y., Schupp, P. J., Wörheide, G., & Erpenbeck, D. (2021). A Soft Spot for Chemistry–Current Taxonomic and Evolutionary Implications of Sponge Secondary Metabolite Distribution. Marine Drugs, 19(8), 448. https://doi.org/10.3390/md19080448