
Proteases Production and Chitin Preparation from the Liquid Fermentation of Chitinous Fishery By-Products by Paenibacillus elgii


 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussions
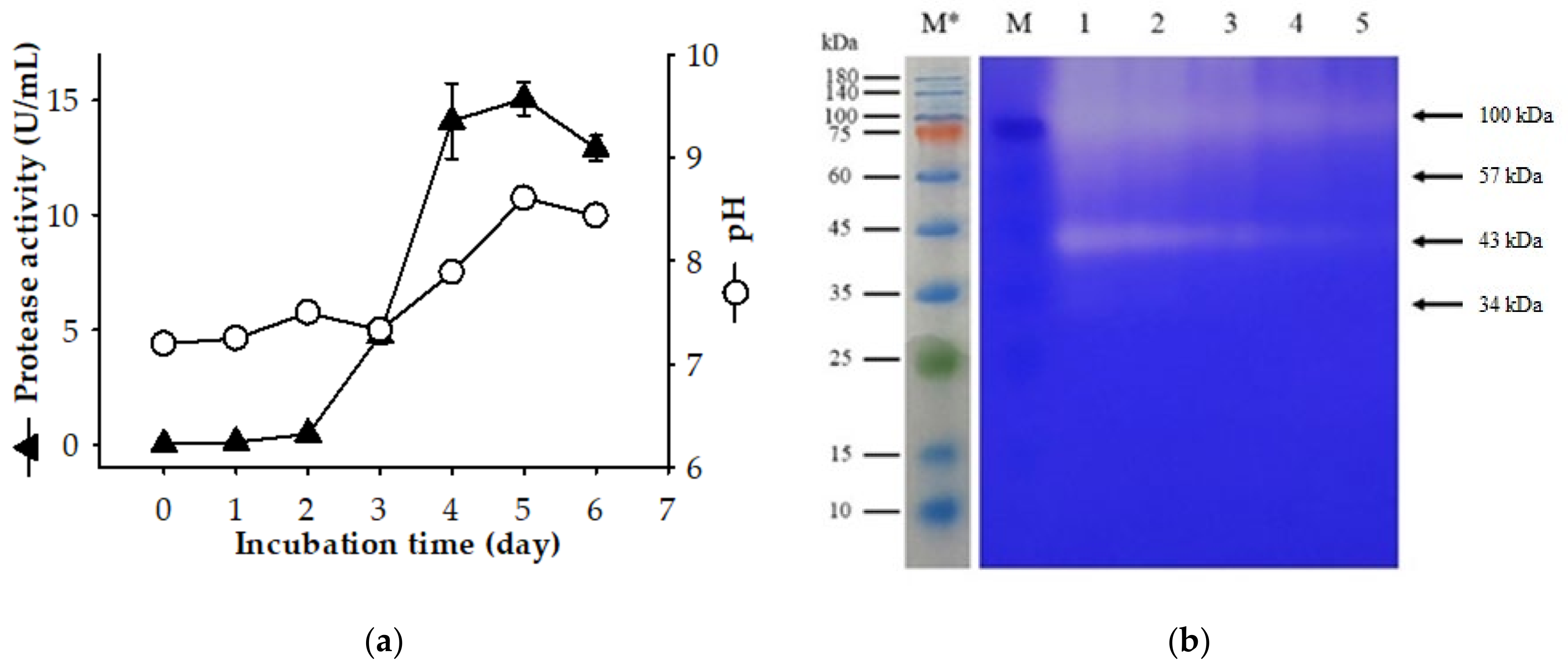
2.1. Screening a Protease-Producing Bacterial Strain and the Production of Extracellular Proteases by P. elgii TKU051
2.2. Biochemical Characterization of P. elgii TKU051 Crude Enzyme Cocktail
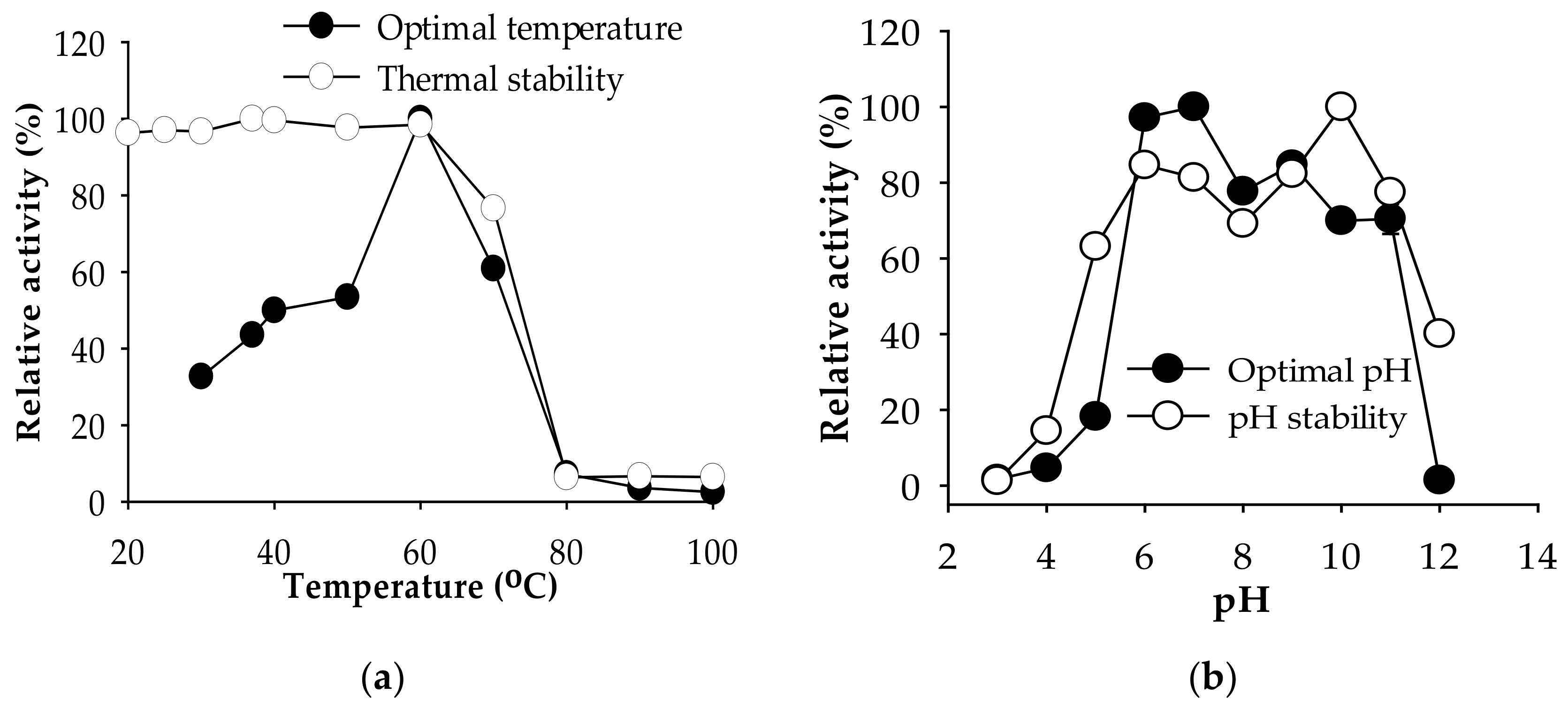
2.2.1. Effect of Temperature and pH
2.2.2. Effect of Metal Ions, Inhibitors, Surfactants, and Reducing Agents
2.2.3. Substrate Specificity
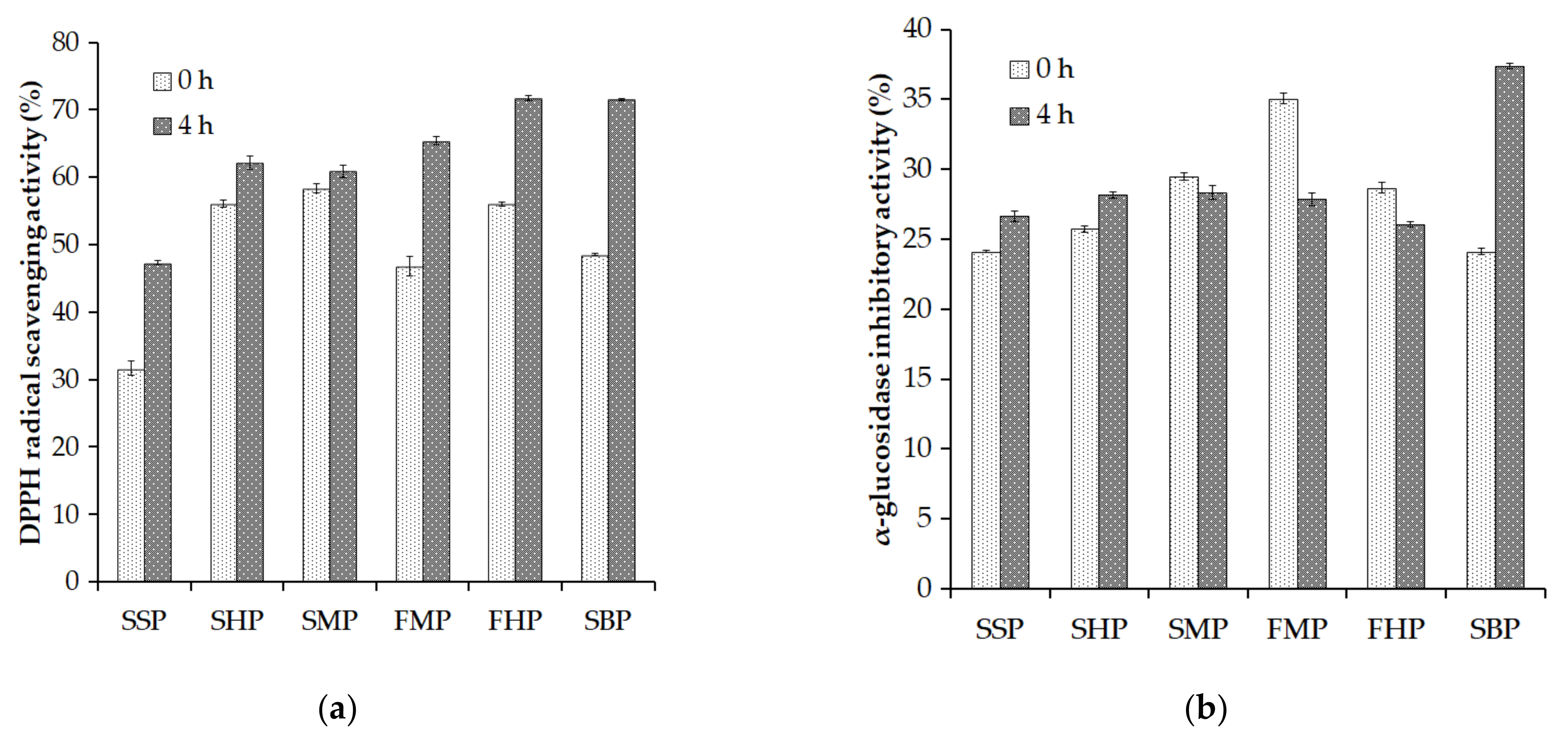
2.3. Evaluation of the 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity and α-Glucosidase Inhibitory Activity of Some Proteinaceous Material Hydrolysates
2.4. Deproteinization Ability of P. elgii TKU051
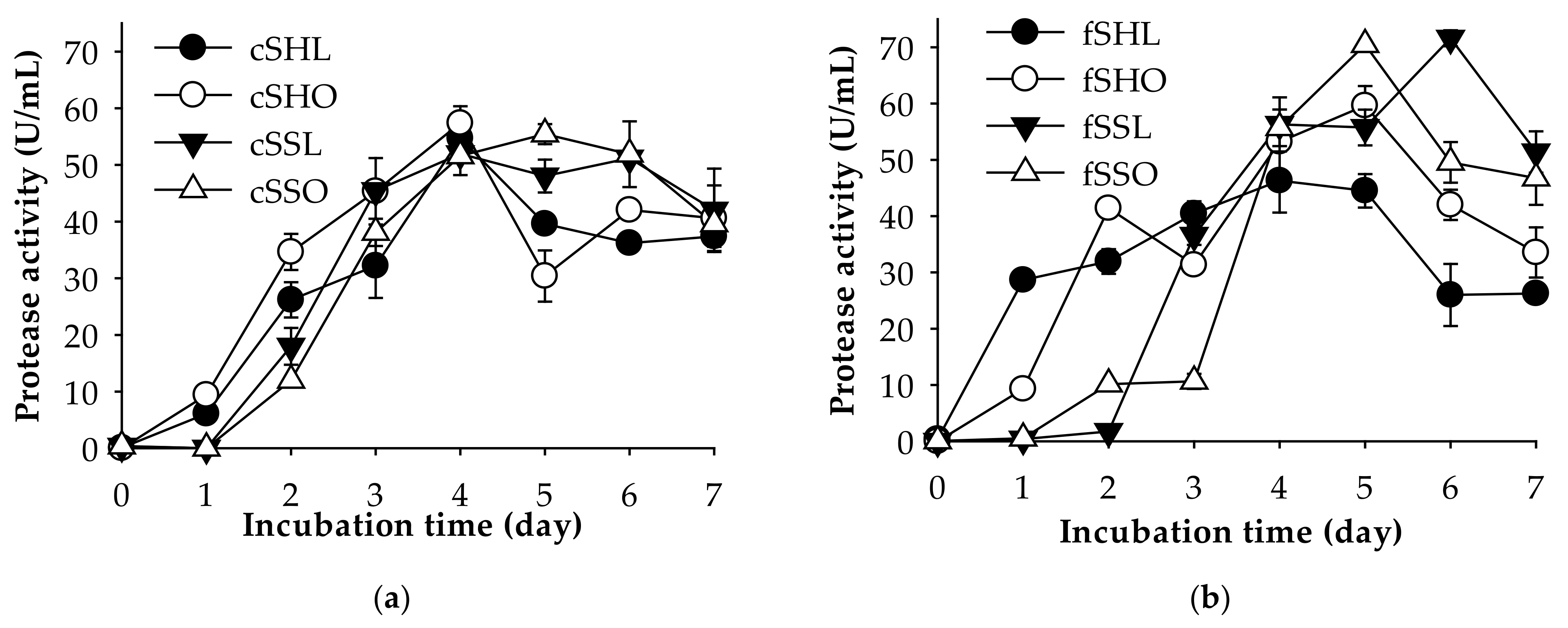
2.4.1. Type of Shrimp Waste
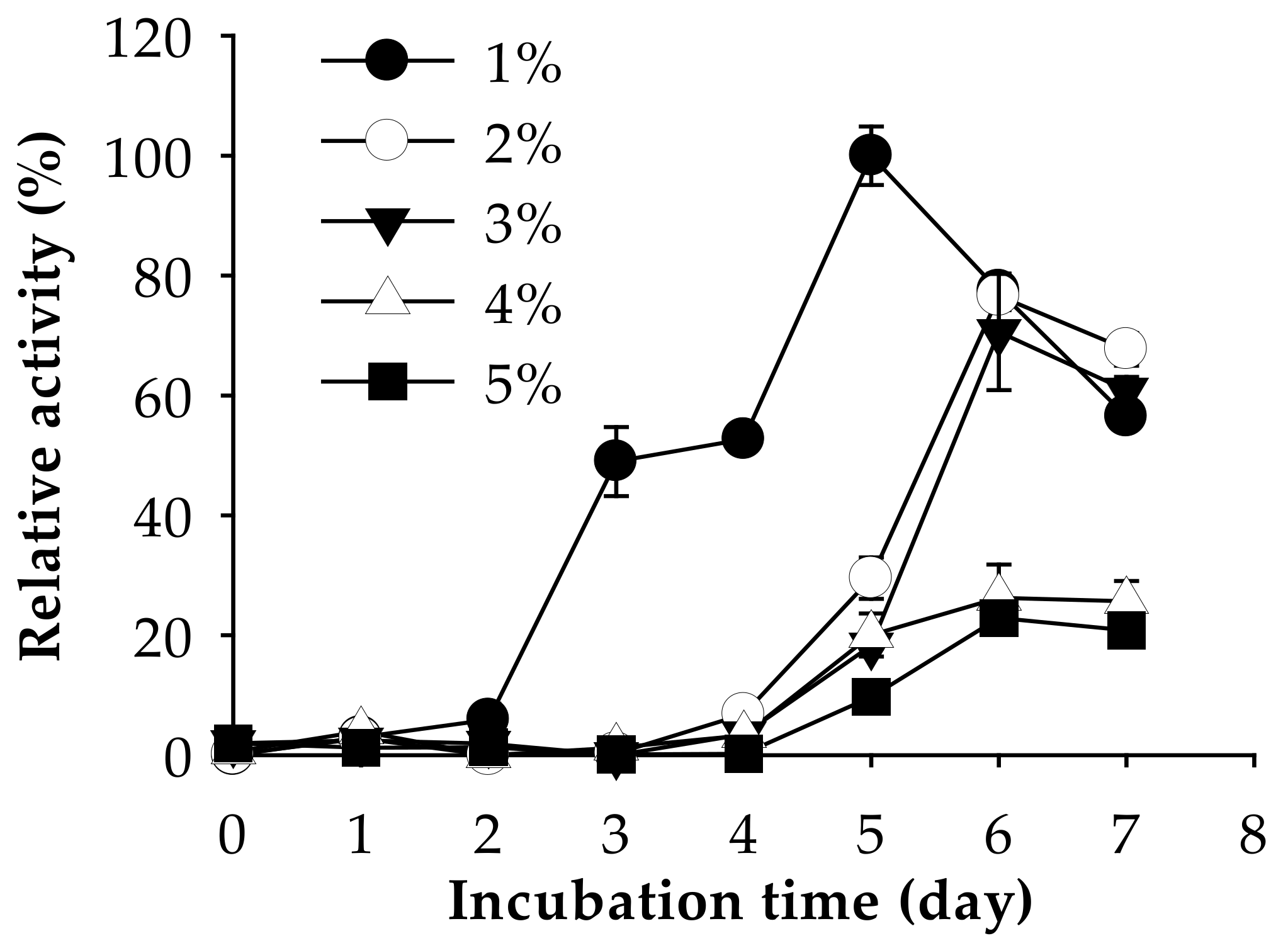
2.4.2. Amount of Shrimp Waste
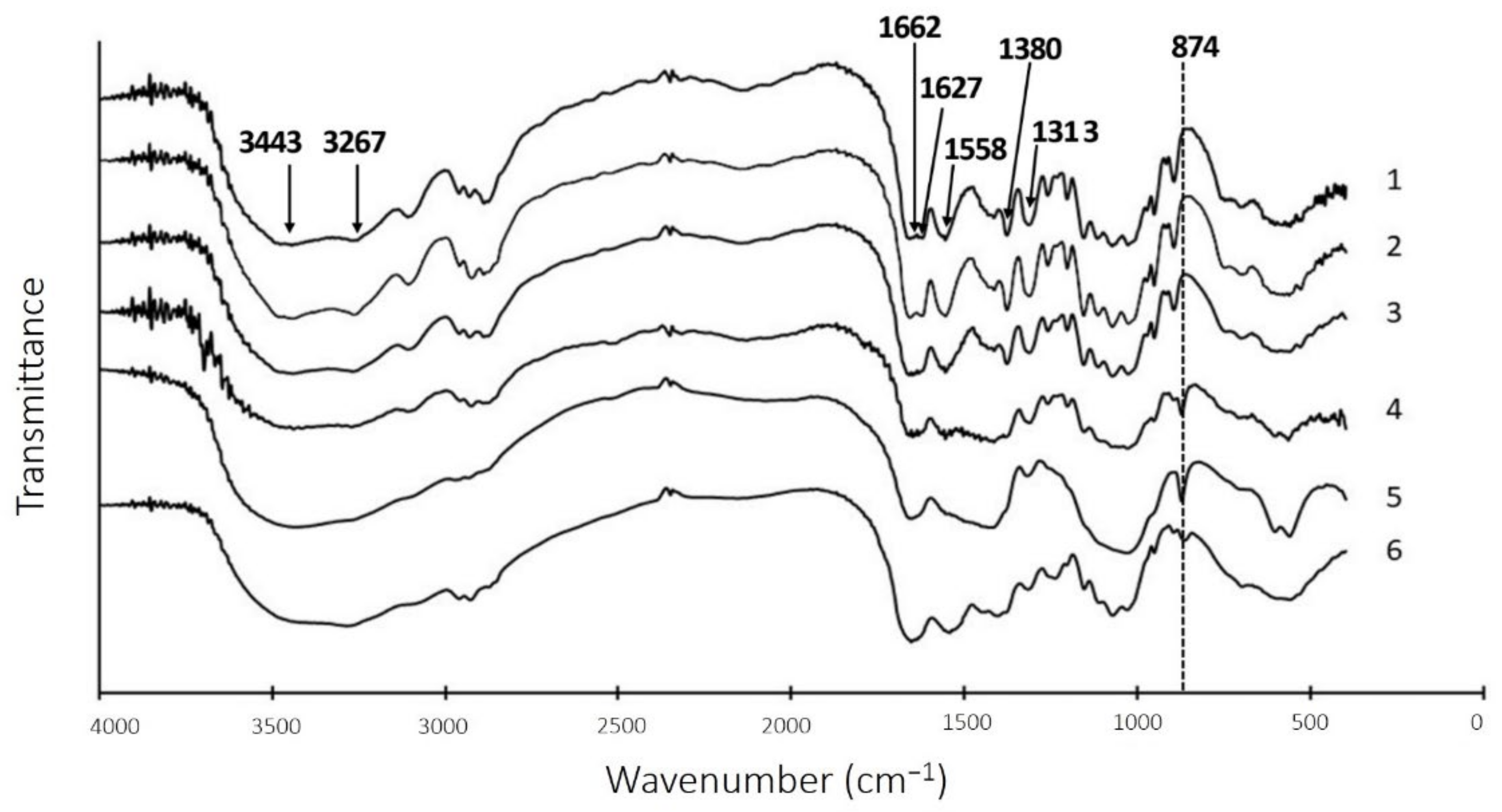
2.5. Chitin Extraction after Liquid Fermentation
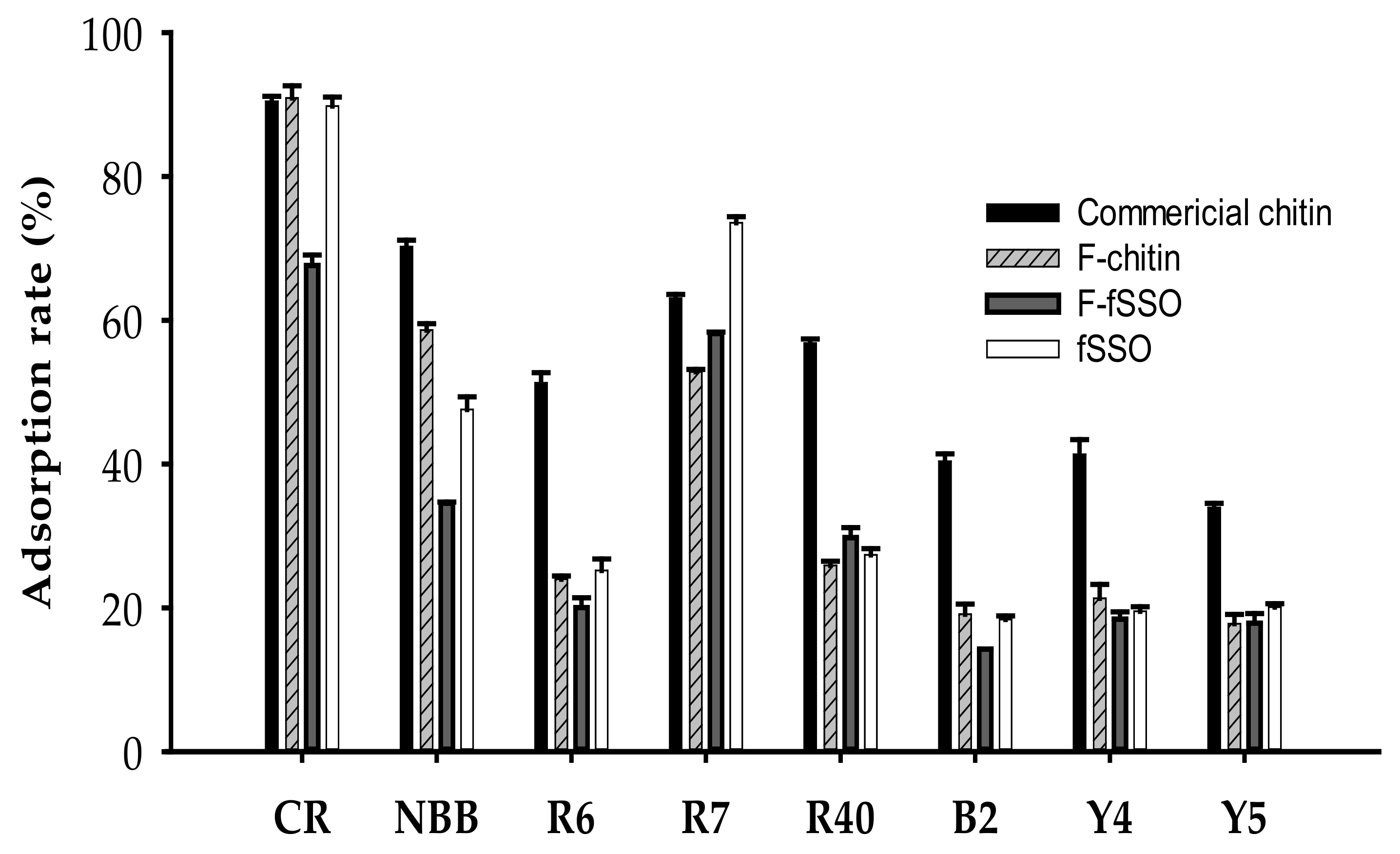
2.6. Dyes Adsorption Ability of Chitins
3. Materials and Methods
3.1. Materials
3.2. Isolation and Identification of Protease-Producing Bacteria
3.3. Measurement of Protease Activity
3.4. Gel Electrophoresis
3.5. Effect of Temperature and pH on Protease Activity and Stability
3.6. Effect of Various Chemicals on the Activity of Protease
3.7. Substrate Specificity
3.8. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity
3.9. α-Glucosidase Inhibitory Activity
3.10. Fermentation Conditions
3.11. Deproteinization Assay
3.12. Chitin Extraction
3.13. FTIR Analysis
3.14. Dye Adsorption
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kaya, M.; Mujtaba, M.; Ehrlich, H.; Salaberria, A.M.; Baran, T.; Amemiya, C.T.; Galli, R.; Akyuz, L.; Sargin, I.; Labidi, J. On chemistry of γ-chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Yan, N.; Chen, X. Sustainability: Don’t waste seafood waste. Nature 2015, 524, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chio, S. Deproteinization of shrimp and crab shell with the protease of Pseudomonas aeruginosa K-187. Enzym. Microb. Technol. 1998, 22, 629–633. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Vo, T.P.K.; Nguyen, A.D.; Wang, S.-L. Chitin extraction from shrimp waste by liquid fermentation using an alkaline protease-producing strain, Brevibacillus parabrevis. Int. J. Biol. Macromol. 2019, 131, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Mechri, S.; Sellem, I.; Bouacem, K.; Jabeur, F.; Laribi-Habchi, H.; Mellouli, L.; Hacène, H.; Bouanane-Darenfed, A.; Jaouadi, B. A biological clean processing approach for the valorization of speckled shrimp Metapenaeus monoceros by-product as a source of bioactive compounds. Environ. Sci. Pollut. Res. 2020, 27, 15842–15855. [Google Scholar] [CrossRef]
- Ningthoujam, D.S.; Kshetri, P.; Sanasam, S.; Nimaichand, S. Screening, identification of best producers and optimization of extracellular proteases from moderately halophilic alkalithermotolerant indigenous actinomycetes. World Appl. Sci. J. 2009, 7, 907–916. [Google Scholar]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Antonie van Leeuwenhoek 1994, 64, 253–260. [Google Scholar] [CrossRef]
- Shida, O.; Takagi, H.; Kadowaki, K.; Nakamura, L.K.; Komagata, K. Transfer of Bacillus alginolyticus, Bacillus chondroitinus, Bacillus curdlanolyticus, Bacillus glucanolyticus, Bacillus kobensis, and Bacillus thiaminolyticus to the genus Paenibacillus and emended description of the genus Paenibacillus. Int. J. Syst. Evol. Microbiol. 1997, 47, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Bae, C.; Jeon, J.; Chun, S.; Oh, H.W.; Hong, S.G.; Baek, K.; Moon, E.Y.; Bae, K.S. Paenibacillus elgii sp. nov. with broad antimicrobial activity. Int. J. Syst. Evol. Microbiol. 2004, 54, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.K.; Selitrennikoff, C.P. Plant and bacterial chitinases differ in antifungal activity. Microbiology 1988, 134, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Jung, H.; Kim, K.Y.; Park, S.K. An effective biocontrol bioformulation against Phytophthora blight of pepper using growth mixtures of combined chitinolytic bacteria under different field conditions. Eur. J. Plant Pathol. 2007, 120, 373–382. [Google Scholar] [CrossRef]
- Lee, D.; Song, H. Antimicrobial activity by Paenibacillus elgii DS381 and its antimicrobial substances against microbial residents on human skin and pathogenic bacteria. Microbiol. Soc. Korea 2018, 54, 244–253. [Google Scholar]
- Garg, V.; Gupta, R.; Yadav, A.B.; Kumar, R. Dye removal from aqueous solution by adsorption on treated sawdust. Bioresour. Technol. 2003, 89, 121–124. [Google Scholar] [CrossRef]
- Liu, J.; Yu, H.; Wang, L. Superior absorption capacity of tremella like ferrocene-based metal-organic framework in removal of organic dye from water. J. Hazard. Mater. 2020, 392, 122274. [Google Scholar] [CrossRef]
- Buntić, A.V.; Pavlović, M.D.; Antonović, D.G.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. A treatment of wastewater containing basic dyes by the use of new strain Streptomyces microflavus CKS6. J. Clean. Prod. 2017, 148, 347–354. [Google Scholar] [CrossRef]
- Piaskowski, K.; Świderska-Dąbrowska, R.; Zarzycki, P.K. Dye removal from water and wastewater using various physical, chemical, and biological processes. J. AOAC Int. 2018, 101, 1371–1384. [Google Scholar] [CrossRef]
- Mckay, G.; Blair, H.; Gardner, J. Adsorption of dyes on chitin. Equilibrium studies. J. Appl. Polym. Sci. 1982, 27, 3043–3057. [Google Scholar] [CrossRef]
- Akkaya, G.; Uzun, I.; Güzel, F. Kinetics of the adsorption of reactive dyes by chitin. Dye Pigment. 2007, 73, 168–177. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Wang, C.-L.; Wang, S.-L. Microbial conversion of shrimp heads to proteases and chitin as an effective dye adsorbent. Polymers 2020, 12, 2228. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.-W.; Wu, C.-C.; Cheng, W.-T.; Chen, Y.-C.; Wang, C.-L.; Wang, I.-L.; Wang, S.-L. Exopolysaccharides and antimicrobial biosurfactants produced by Paenibacillus macerans TKU029. Appl. Biochem. Biotechnol. 2013, 172, 933–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S. Utilization of seafood processing by-products for production of proteases by Paenibacillus sp. TKU052 and their application in biopeptides preparation. Mar. Drugs 2020, 18, 574. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Wen, I.-H.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Conversion of shrimp head waste for production of a thermotolerant, detergent-stable, alkaline protease by Paenibacillus sp. Catalysts 2019, 9, 798. [Google Scholar] [CrossRef] [Green Version]
- Aw, Y.-K.; Ong, K.-S.; Lee, L.-H.; Cheow, Y.-L.; Yule, C.; Lee, S.-M. Newly isolated Paenibacillus tyrfis sp. nov.; from Malaysian tropical peat swamp soil with broad spectrum antimicrobial activity. Front. Microbiol. 2016, 7, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Park, S.K.; Hur, J.Y.; Kim, Y.C. Purification and characterization of a major extracellular chitinase from a biocontrol bacterium, Paenibacillus elgii HOA73. Plant Pathol. J. 2017, 33, 318–328. [Google Scholar] [CrossRef] [Green Version]
- Hrabák, J.; Martínek, K. Screening of secreted proteases of Paenibacillus larvae by using substrate-SDS-polyacrylamide gel electrophoresis. J. Apic. Res. 2007, 46, 160–164. [Google Scholar] [CrossRef]
- Alvarez, V.; Von Der Weid, I.; Seldin, L.; Santos, A. Influence of growth conditions on the production of extracellular proteolytic enzymes in Paenibacillus peoriae NRRL BD-62 and Paenibacillus polymyxa SCE2. Lett. Appl. Microbiol. 2006, 43, 625–630. [Google Scholar] [CrossRef]
- Kim, Y.C.; Hur, J.Y.; Park, S.K. Biocontrol of Botrytis cinerea by chitin-based cultures of Paenibacillus elgii HOA73. Eur. J. Plant. Pathol. 2019, 155, 253–263. [Google Scholar] [CrossRef]
- Paul, T.; Halder, S.K.; Das, A.; Bera, S.; Maity, C.; Mandal, A.; Das, P.S.; Mohapatra, P.K.D.; Pati, B.R.; Mondal, K.C. Exploi-tation of chicken feather waste as a plant growth promoting agent using keratinase producing novel isolate Paenibacillus woosongensis TKB2. Biocatal. Agric. Biotechnol. 2013, 2, 50–57. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Conversion of squid pens to chitosanases and proteases via Paenibacillus sp. TKU042. Mar. Drugs 2018, 16, 83. [Google Scholar] [CrossRef] [Green Version]
- Hang, F.; Liu, P.; Wang, Q.; Han, J.; Wu, Z.; Gao, C.; Liu, Z.; Zhang, H.; Chen, W. High milk-clotting activity expressed by the newly isolated Paenibacillus spp. strain BD3526. Molecules 2016, 21, 73. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Lu, Z.; Bie, X.; Yao, Z.; Wang, Y.; Lu, Y.; Guo, Y. Purification and characterization of a novel anticoagulant and fibri-nolytic enzyme produced by endophytic bacterium Paenibacillus polymyxa EJS-3. Thromb. Res. 2010, 126, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pan, Y.; She, Q.; Chen, L. A novel carboxyl-terminal protease derived from Paenibacillus lautus CHN26 exhibiting high activities at multiple sites of substrates. BMC Biotechnol. 2013, 13, 89. [Google Scholar] [CrossRef] [Green Version]
- Rai, S.K.; Roy, J.K.; Mukherjee, A.K. Characterisation of a detergent-stable alkaline protease from a novel thermophilic strain Paenibacillus tezpurensis sp. nov. AS-S24-II. Appl. Microbiol. Biotechnol. 2009, 85, 1437–1450. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Dhandapani, B.; Mahadevan, S. Optimization studies on production of a salt-tolerant protease from Pseu-domonas aeruginosa strain BC1 and its application on tannery saline wastewater treatment. Braz. J. Microbiol. 2011, 42, 1506–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products—A review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- De Oliveira, C.F.; Correa, A.P.F.; Coletto, D.; Daroit, D.J.; Cladera-Olivera, F.; Brandelli, A. Soy protein hydrolysis with microbial protease to improve antioxidant and functional properties. J. Food Sci. Technol. 2014, 52, 2668–2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Anti-α-glucosidase activity by a protease from Bacillus licheniformis. Molecules 2019, 24, 691. [Google Scholar] [CrossRef] [Green Version]
- Henriques, A.; Vázquez, J.; Valcarcel, J.; Mendes, R.; Bandarra, N.; Pires, C. Characterization of protein hydrolysates from fish discards and by-products from the north-west Spain fishing fleet as potential sources of bioactive peptides. Mar. Drugs 2021, 19, 338. [Google Scholar] [CrossRef]
- Bhaskar, N.; Suresh, P.V.; Sakhare, P.Z.; Sachindra, N.M. Shrimp biowaste fermentation with Pediococcus acidolactici CFR2182: Optimization of fermentation conditions by response surface methodology and effect of optimized conditions on deprotein-ation/demineralization and carotenoid recovery. Enzym. Microb. Technol. 2007, 40, 1427–1434. [Google Scholar] [CrossRef]
- Sini, T.K.; Santhosh, S.; Mathew, P.T. Study on the production of chitin and chitosan from shrimp shell by using Bacillus subtilis fermentation. Carbohydr. Res. 2007, 342, 2423–2429. [Google Scholar] [CrossRef]
- Panêlová, M.; Machoviĉ, V.; Březina, M. Characterization and sorption properties of Aspergillus niger waste biomass. Cent. Eur. J. Chem. 2003, 1, 192–200. [Google Scholar] [CrossRef]
- Chen, X.; Chew, S.L.; Kerton, F.; Yan, N. Direct conversion of chitin into a N-containing furan derivative. Green Chem. 2014, 16, 2204–2212. [Google Scholar] [CrossRef] [Green Version]
- Hassainia, A.; Satha, H.; Boufi, S. Chitin from Agaricus bisporus: Extraction and characterization. Int. J. Biol. Macromol. 2018, 117, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Duran, C.; Ozdes, D.; Gundogdu, A.; Senturk, H.B. Kinetics and isotherm analysis of basic dyes adsorption onto almond shell (Prunus dulcis) as a low-cost Adsorbent. J. Chem. Eng. Data 2011, 56, 2136–2147. [Google Scholar] [CrossRef]
- Piccin, J.S.; Dotto, G.L.; Vieira, M.L.; Pinto, L.A. Kinetics and mechanism of the food dye FD&C Red 40 adsorption onto chi-tosan. J. Chem. Eng. Data 2011, 56, 3759–3765. [Google Scholar]
- Srinivasan, A.; Viraraghavan, T. Decolorization of dye wastewaters by biosorbents: A review. J. Environ. Manag. 2010, 91, 1915–1929. [Google Scholar] [CrossRef]
- Sánchez-Duarte, R.G.; Sánchez-Machado, D.I.; López-Cervantes, J.; Murrieta, M.A.C. Adsorption of allura red dye by cross-linked chitosan from shrimp waste. Water Sci. Technol. 2012, 65, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-L.; Chen, Y.-C.; Yen, Y.-H.; Liang, T.-W. Utilisation of chitinous materials in pigment adsorption. Food Chem. 2012, 135, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-L.; Yu, H.-T.; Tsai, M.-H.; Doan, C.T.; Nguyen, V.B.; Do, V.C.; Nguyen, A.D. Conversion of squid pens to chitosanases and dye adsorbents via Bacillus cereus. Res. Chem. Intermed. 2018, 44, 4903–4911. [Google Scholar] [CrossRef]
- Prado, A.G.; Torres, J.D.; Faria, E.A.; Dias, S.C. Comparative adsorption studies of indigo carmine dye on chitin and chitosan. J. Colloid Interface Sci. 2004, 277, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.-W.; Huang, C.-T.; Dzung, N.A.; Wang, S.-L. Squid pen chitin chitooligomers as food colorants absorbers. Mar. Drugs 2015, 13, 681–696. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.W.; Huffaker, R.C. Loss of ribulose 1,5-diphosphate carboxylase and increase in proteolytic activity during senescence of detached primary barley leaves. Plant Physiol. 1975, 55, 1009–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, D.F.; Saturnino, T.P.; Fernandes, F.F.; Mazzola, P.G.; Silveira, E.; Tambourgi, E.B. Azocasein substrate for determination of proteolytic activity: Reexamining a traditional method using bromelain samples. Biomed. Res. Int. 2016, 2016, 8409183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Ramnani, P. Microbial keratinases and their prospective applications: An overview. Appl. Microbiol. Biotechnol. 2006, 70, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Production of a thermostable chitosanase from shrimp heads via Paenibacillus mucilaginosus TKU032 conversion and its application in the preparation of bioactive chitosan oligosaccharides. Mar. Drugs 2019, 17, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.N.; Doan, C.T.; Nguyen, A.D.; Wang, S.-L. Antioxidant and anti-diabetes potential of water-soluble chitosan-glucose derivatives produced by Maillard reaction. Polymers 2019, 11, 1714. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Mizani, A.M.; Aminlari, B.M. A new process for deproteinization of chitin from shrimp head waste. In Proceedings of the European Congress of Chemical Engineering (ECCE-6), Copenhagen, Denmark, 16–20 September 2007; pp. 16–20. [Google Scholar]
- Rao, M.S.; Muñoz, J.; Stevens, W.F. Critical factors in chitin production by fermentation of shrimp biowaste. Appl. Microbiol. Biotechnol. 2000, 54, 808–813. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MW (kDa) | C/N Source | Ref. |
|---|---|---|---|
| P. elgii TKU051 | 100, 57, 43, 34 | SPP | This study |
| P. woosongenisis TKB2 | chicken feather | [28] | |
| P. mucilaginosus TKU032 | 32 | shrimp head powder | [19] |
| Paenibacillus sp. TKU047 | 32 | shrimp head powder | [22] |
| Paenibacillus sp. TKU042 | 35 | SPP | [29] |
| Paenibacillus spp. BD3526 | 35 | wheat bran | [30] |
| P. polymyxa EJS-3 | 63.3 | tryptone, beef extract | [31] |
| P. larvae | 87, 74, 42, 40 | MYPGP broth | [25] |
| Paenibacillus sp. TKU052 | 31 1 | demineralized crab shell powder | [21] |
| P. lautus CHN26 | 53.92 2 | [32] | |
| P. tezpurensis sp. now. AS-S24-II | 43 | casein and ammonium sulphate | [33] |
| P. polymyxa SCE2 and P. peoriae NRRL BD-62 | 210, 50, 35, 20 | thiamine/biotin/nitrogen broth | [26] |
| Chemical | Concentration | Relative Activity (%) |
|---|---|---|
| None | 100.00 ± 1.54 | |
| MgSO4 | 5 mM | 113.34 ± 1.15 |
| CuSO 4 | 5 mM | NA |
| FeSO4 | 5 mM | 87.24 ± 2.58 |
| CaCl2 | 5 mM | 108.12 ± 2.80 |
| ZnSO4 | 5 mM | NA |
| MnSO4 | 5 mM | 364.04 ± 7.50 |
| ZnCl2 | 5 mM | NA |
| BaCl2 | 5 mM | 99.07 ± 2.28 |
| PMSF | 5 mM | 52.81 ± 8.18 |
| EDTA | 5 mM | 20.16 ± 0.76 |
| SDS | 2 mM | 22.50 ± 2.65 |
| β-ME | 2 mM | 88.91 ± 3.93 |
| Tween 20 | 2% | 177.26 ± 0.44 |
| Tween 40 | 2% | 204.59 ± 5.14 |
| Triton X-100 | 2% | 127.96 ± 7.62 |
| Substrate | Relative Activity (%) |
|---|---|
| Casein | 100.00 ± 0.19 |
| Hemoglobin | 40.80 ± 0.16 |
| Albumin | 36.28 ± 0.41 |
| Myoglobin | 31.80 ± 0.20 |
| Elastin | 25.80 ± 0.34 |
| Fibrin | 29.07 ± 0.44 |
| Gelatin | 23.54 ± 0.62 |
| Keratin | 58.85 ± 0.15 |
| SSP | 28.49 ± 0.24 |
| SHP | 24.01 ± 0.46 |
| FMP | 34.96 ± 0.29 |
| SMP | 44.50 ± 0.93 |
| FHP | 31.99 ± 0.78 |
| SBP | 29.81 ± 0.22 |
| Kind of Shrimp Waste | Deproteinization Rate (%) | Residual Protein Content (%) | ||
|---|---|---|---|---|
| Fermentation | Nonfermentation | Fermentation | Nonfermentation | |
| cSHL | 95.90 ± 0.18 | 50.29 ± 1.71 | 1.99 ± 0.13 | 24.17 ± 0.47 |
| cSHO | 94.47 ± 0.66 | 42.38 ± 3.54 | 2.68 ± 0.31 | 27.91 ± 1.09 |
| cSSL | 94.61 ± 0.29 | 38.00 ± 0.45 | 2.25 ± 0.12 | 25.86 ± 0.10 |
| cSSO | 94.40 ± 0.24 | 43.09 ± 4.89 | 2.37 ± 0.07 | 24.03 ± 2.17 |
| fSHL | 96.86 ± 0.33 | 55.43 ± 1.11 | 1.51 ± 0.19 | 21.45 ± 0.57 |
| fSHO | 95.16 ± 0.26 | 53.25 ± 3.97 | 2.22 ± 0.15 | 21.43 ± 1.99 |
| fSSL | 95.23 ± 0.14 | 50.59 ± 3.41 | 2.10 ± 0.04 | 21.53 ± 2.04 |
| fSSO | 94.36 ± 0.19 | 46.74 ± 2.59 | 2.46 ± 0.01 | 23.67 ± 2.23 |
| Amount of fSSO | Deproteinization Rate (%) | Residual Protein Content (%) | ||
|---|---|---|---|---|
| Fermentation | Nonfermentation | Fermentation | Nonfermentation | |
| 1% | 97.45 ± 0.03 | 51.48 ± 1.45 | 1.11 ± 0.04 | 21.15 ± 0.70 |
| 2% | 46.93 ± 4.18 | 50.21 ± 2.16 | 23.12 ± 1.96 | 21.69 ± 0.35 |
| 3% | 50.60 ± 3.32 | 42.94 ± 0.57 | 21.51 ± 1.07 | 24.89 ± 0.73 |
| 4% | 48.81 ± 3.66 | 16.26 ± 0.30 | 22.31 ± 1.78 | 36.54 ± 1.47 |
| 5% | 33.34 ± 0.06 | 8.85 ± 3.66 | 29.08 ± 1.12 | 39.72 ± 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-H.; Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, C.-L.; Wang, S.-L. Proteases Production and Chitin Preparation from the Liquid Fermentation of Chitinous Fishery By-Products by Paenibacillus elgii. Mar. Drugs 2021, 19, 477. https://doi.org/10.3390/md19090477
Lee D-H, Doan CT, Tran TN, Nguyen VB, Nguyen AD, Wang C-L, Wang S-L. Proteases Production and Chitin Preparation from the Liquid Fermentation of Chitinous Fishery By-Products by Paenibacillus elgii. Marine Drugs. 2021; 19(9):477. https://doi.org/10.3390/md19090477
Chicago/Turabian StyleLee, Dan-Hsin, Chien Thang Doan, Thi Ngoc Tran, Van Bon Nguyen, Anh Dzung Nguyen, Chuan-Lu Wang, and San-Lang Wang. 2021. "Proteases Production and Chitin Preparation from the Liquid Fermentation of Chitinous Fishery By-Products by Paenibacillus elgii" Marine Drugs 19, no. 9: 477. https://doi.org/10.3390/md19090477
APA StyleLee, D.-H., Doan, C. T., Tran, T. N., Nguyen, V. B., Nguyen, A. D., Wang, C.-L., & Wang, S.-L. (2021). Proteases Production and Chitin Preparation from the Liquid Fermentation of Chitinous Fishery By-Products by Paenibacillus elgii. Marine Drugs, 19(9), 477. https://doi.org/10.3390/md19090477