
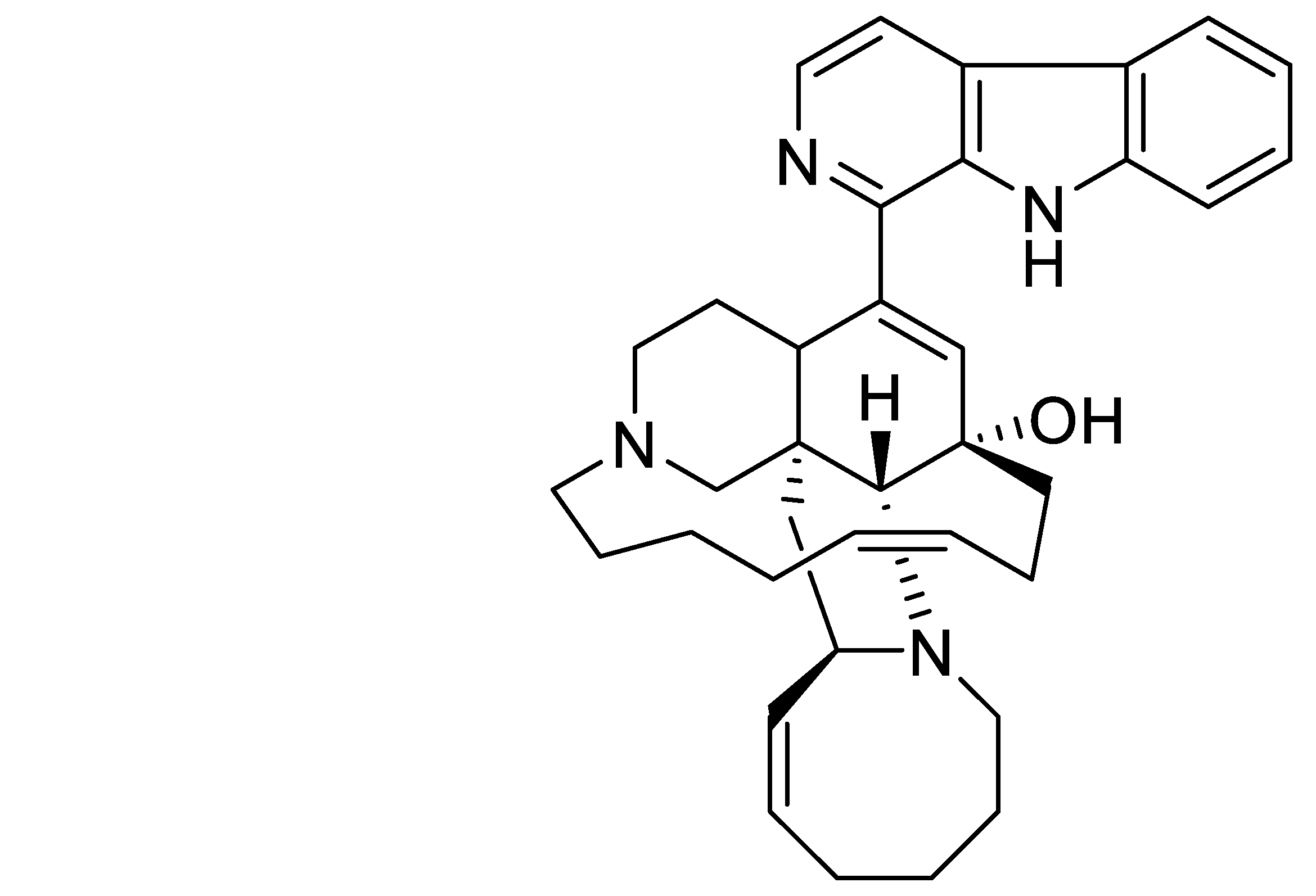
RSK1 vs. RSK2 Inhibitory Activity of the Marine β-Carboline Alkaloid Manzamine A: A Biochemical, Cervical Cancer Protein Expression, and Computational Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Effect of MZA on a Protein Kinases Screening Array and Docking Simulations
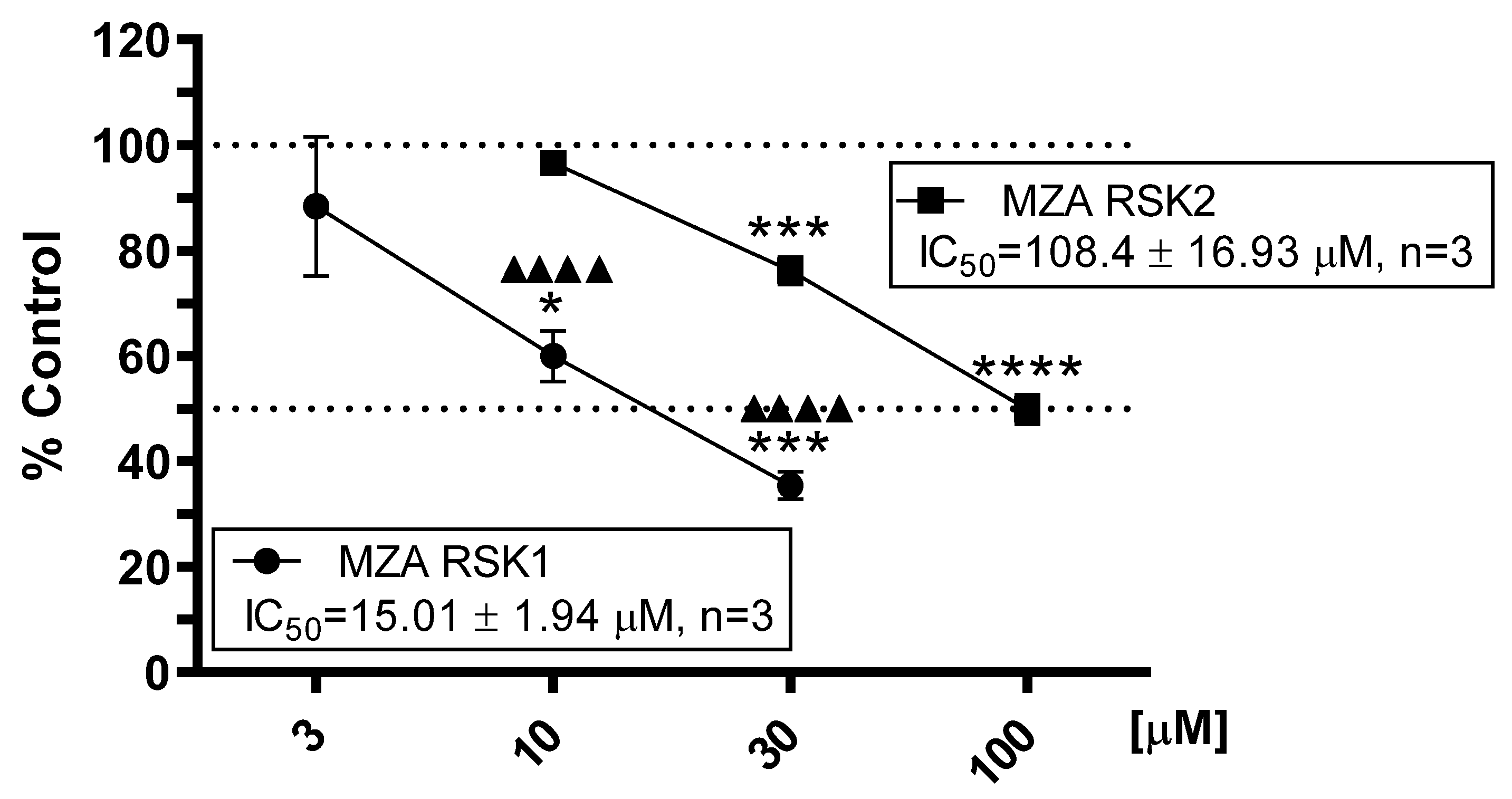
2.2. Effect of MZA on RSK1 and RSK2 Protein Kinase Assays
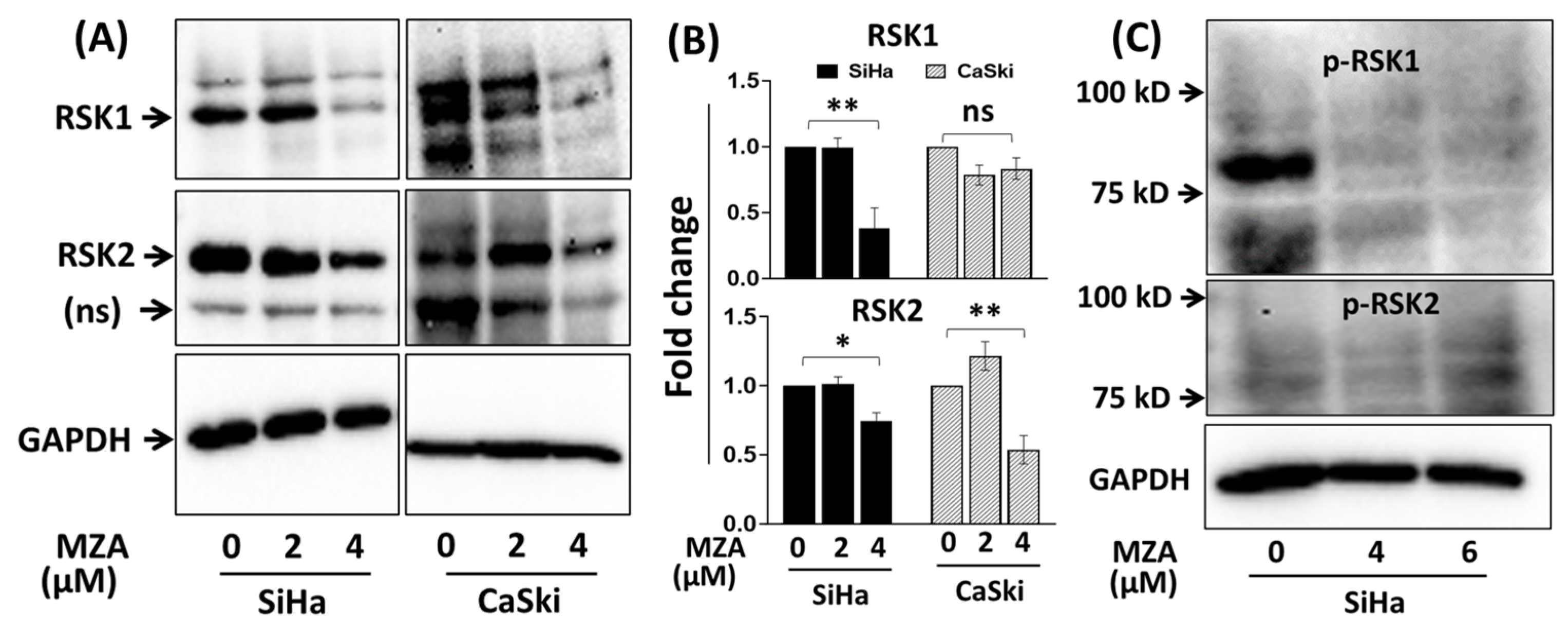
2.3. Effect of MZA on RSK1 and RSK2 in Human Cervical Carcinoma Cell Lines
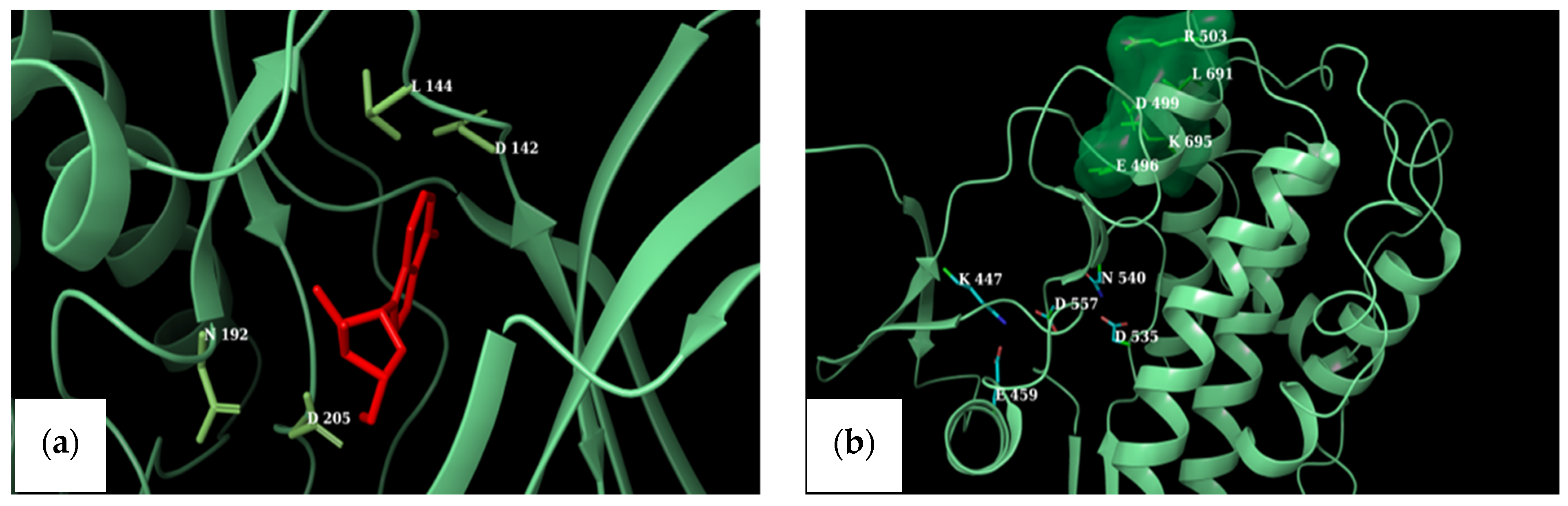
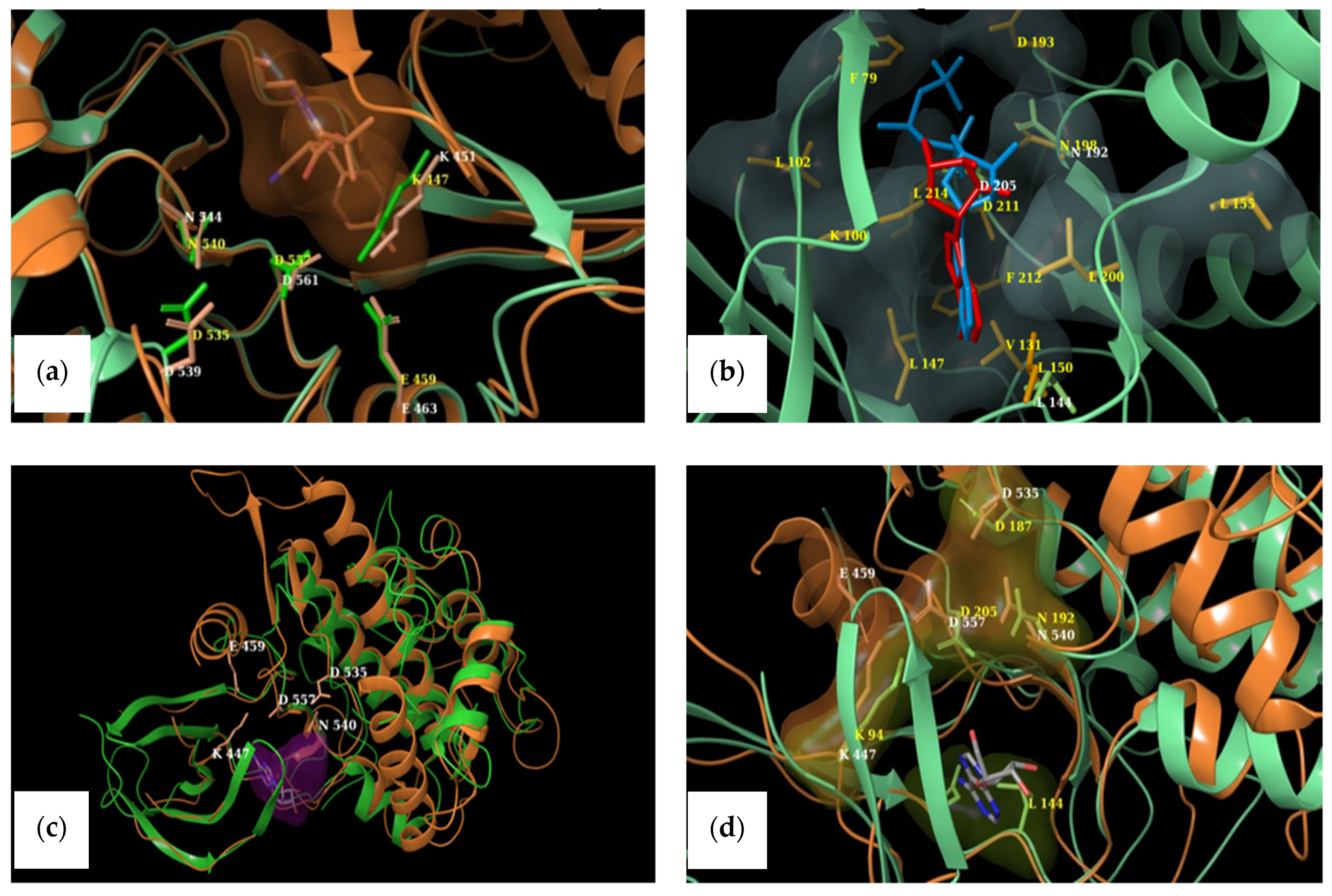
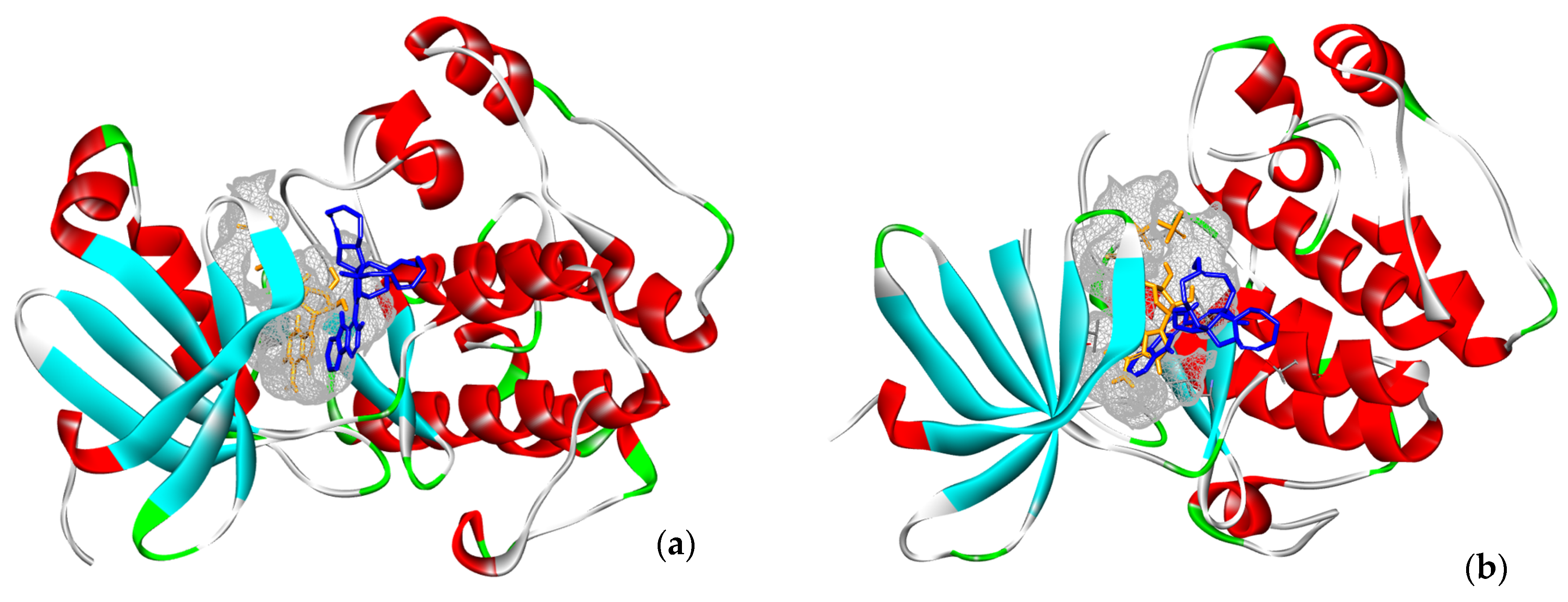
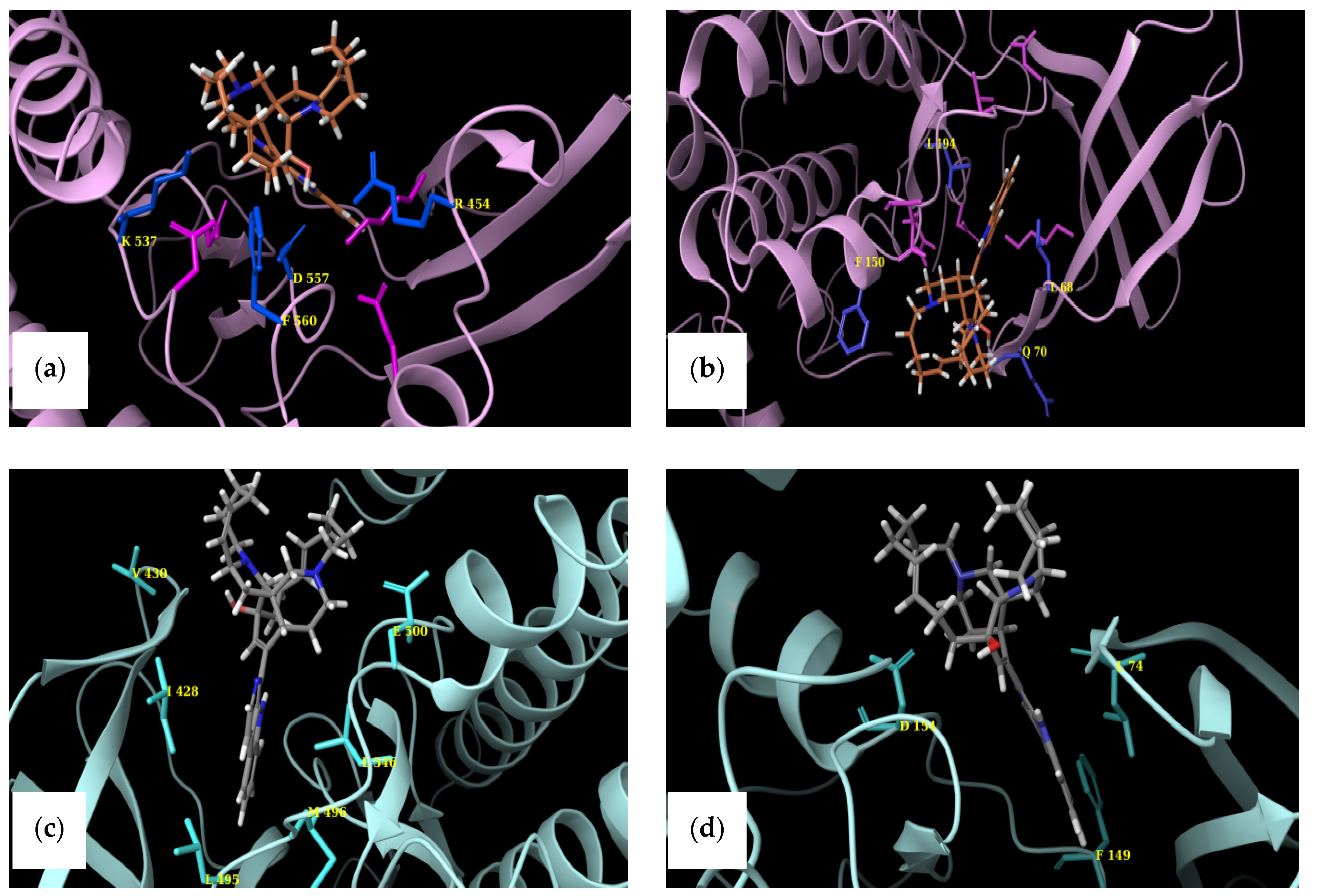
2.4. Structural Comparisons
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Protein Kinase Activity Assays
4.3. RSK1 and RSK2 Protein Kinase Assays
4.4. Computational Methods
4.4.1. Molecular Docking with AutoDock Vina
4.4.2. Selection of Targets and Identification of Critical Amino Acids
4.4.3. Protein Preparation
4.4.4. Docking Glide 5.7
4.5. Immunoblotting for RSK1 and RSK2 in SiHa and CaSki Human Cervical Carcinoma Cancer Cell Lines
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mayer, A.M.S.; Guerrero, A.J.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2021, 19, 49. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, N.; Zhang, T.; Zhang, B.; Sajeevan, T.P.; Joseph, V.; Armstrong, L.; He, S.; Yan, X.; Naman, C.B. A Systematic Review of Recently Reported Marine Derived Natural Product Kinase Inhibitors. Mar. Drugs 2019, 17, 493. [Google Scholar] [CrossRef] [Green Version]
- Sakai, R.; Higa, T. Manzamine A, a novel antitumor alkaloid from a sponge. J. Am. Chem. Soc. 1986, 108, 6404–6405. [Google Scholar] [CrossRef]
- Wahba, A.E.; Fromentin, Y.; Zou, Y.; Hamann, M.T. Acantholactone, a new manzamine related alkaloid with an unprecedented δ-lactone and ε-lactam ring system. Tetrahedron Lett. 2012, 53, 6329–6331. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.F.; Hamann, M.T.; Hill, R.; Kelly, M. The manzamine alkaloids. Alkaloids Chem. Biol. 2003, 60, 207–285. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, E.A.; Johnson, J.D.; Linley, P.A.; Gunasekera, S.E.; Wright, A.E. A novel activity from an old compound: Manzamine A reduces the metastatic potential of AsPC-1 pancreatic cancer cells and sensitizes them to TRAIL-induced apoptosis. Investig. New Drugs 2011, 29, 777–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ang, K.K.; Holmes, M.J.; Kara, U.A. Immune-mediated parasite clearance in mice infected with Plasmodium berghei following treatment with manzamine A. Parasitol. Res. 2001, 87, 715–721. [Google Scholar] [CrossRef]
- Mayer, A.M.; Hall, M.L.; Lynch, S.M.; Gunasekera, S.P.; Sennett, S.H.; Pomponi, S.A. Differential modulation of microglia superoxide anion and thromboxane B2 generation by the marine manzamines. BMC. Pharmacol. 2005, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Yousaf, M.; Hammond, N.L.; Peng, J.; Wahyuono, S.; McIntosh, K.A.; Charman, W.N.; Mayer, A.M.; Hamann, M.T. New manzamine alkaloids from an Indo-Pacific sponge. Pharmacokinetics, oral availability, and the significant activity of several manzamines against HIV-I, AIDS opportunistic infections, and inflammatory diseases. J. Med. Chem. 2004, 47, 3512–3517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palem, J.R.; Mudit, M.; Hsia, S.V.; Sayed, K.A. Discovery and preliminary structure-activity relationship of the marine natural product manzamines as herpes simplex virus type-1 inhibitors. Z. Naturforsch. C 2017, 72, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Kudrimoti, S.; Prasanna, S.; Odde, S.; Doerksen, R.J.; Pennaka, H.K.; Choo, Y.M.; Rao, K.V.; Tekwani, B.L.; Madgula, V.; et al. Structure-activity relationship and mechanism of action studies of manzamine analogues for the control of neuroinflammation and cerebral infections. J. Med. Chem. 2010, 53, 61–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Higuchi, R.; Miyamoto, T.; Van Soest, R.W. Neuritogenic activity-guided isolation of a free base form manzamine A from a marine sponge, Acanthostrongylophora aff. ingens (Thiele, 1899). Chem. Pharm. Bull. 2008, 56, 866–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguchi, K.; Fujiwara, Y.; Hayashida, A.; Horlad, H.; Kato, H.; Rotinsulu, H.; Losung, F.; Mangindaan, R.E.; de Voogd, N.J.; Takeya, M.; et al. Manzamine A, a marine-derived alkaloid, inhibits accumulation of cholesterol ester in macrophages and suppresses hyperlipidemia and atherosclerosis in vivo. Bioorg. Med. Chem. 2013, 21, 3831–3838. [Google Scholar] [CrossRef]
- Shilabin, A.G.; Kasanah, N.; Tekwani, B.L.; Hamann, M.T. Kinetic studies and bioactivity of potential manzamine prodrugs. J. Nat. Prod. 2008, 71, 1218–1221. [Google Scholar] [CrossRef] [Green Version]
- Hamann, M.; Alonso, D.; Martín-Aparicio, E.; Fuertes, A.; Pérez-Puerto, M.J.; Castro, A.; Morales, S.; Navarro, M.L.; Del Monte-Millán, M.; Medina, M.; et al. Glycogen synthase kinase-3 (GSK-3) inhibitory activity and structure-activity relationship (SAR) studies of the manzamine alkaloids. Potential for Alzheimer’s disease. J. Nat. Prod. 2007, 70, 1397–1405. [Google Scholar] [CrossRef]
- Kallifatidis, G.; Hoepfner, D.; Jaeg, T.; Guzmán, E.A.; Wright, A.E. The marine natural product manzamine A targets vacuolar ATPases and inhibits autophagy in pancreatic cancer cells. Mar. Drugs 2013, 11, 3500–3516. [Google Scholar] [CrossRef] [Green Version]
- Karan, D.; Dubey, S.; Pirisi, L.; Nagel, A.; Pina, I.; Choo, Y.M.; Hamann, M.T. The marine natural product Manzamine A inhibits cervical cancer by targeting the SIX1 protein. J. Nat. Prod. 2020, 83, 286–295. [Google Scholar] [CrossRef]
- Li, W.; Qin, Y.; Zhou, R.; Liu, Y.; Zhang, G. High expression of SIX1 is an independent predictor of poor prognosis in endometrial cancer. Am. J. Transl. Res. 2021, 13, 2840–2848. [Google Scholar]
- Romeo, Y.; Zhang, X.; Roux, P.P. Regulation and function of the RSK family of protein kinases. Biochem. J. 2012, 441, 553–569. [Google Scholar] [CrossRef] [Green Version]
- Heffron, D.; Mandell, J.W. Differential localization of MAPK-activated protein kinases RSK1 and MSK1 in mouse brain. Mol. Brain Res. 2005, 136, 134–141. [Google Scholar] [CrossRef]
- Chandrasena, K.; Hall, M.L.; Mayer, A.M.S. The mechanism of the marine B-carboline thromboxane B2 inhibitor Manzamine A: Possible involvement of rat brain microglia p90 ribosoml S6 kinase 1. FASEB J 2013, 27, 888.1. [Google Scholar] [CrossRef]
- Pereira, P.M.; Schneider, A.; Pannetier, S.; Heron, D.; Hanauer, A. Coffin-Lowry syndrome. Eur. J. Hum. Genet. 2010, 18, 627–633. [Google Scholar] [CrossRef]
- Cho, Y.Y.; Yao, K.; Kim, H.G.; Kang, B.S.; Zheng, D.; Bode, A.M.; Dong, Z. Ribosomal S6 kinase 2 is a key regulator in tumor promoter induced cell transformation. Cancer Res. 2007, 67, 8104–8112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redman, E.K.; Brookes, P.S.; Karcz, M.K. Role of p90RSK in regulating the Crabtree effect: Implications for cancer. Biochem. Soc. Trans. 2013, 41, 124–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapkota, G.P.; Cummings, L.; Newell, F.S.; Armstrong, C.; Bain, J.; Frodin, M.; Grauert, M.; Hoffmann, M.; Schnapp, G.; Steegmaier, M.; et al. BI-D1870 is a specific inhibitor of the p90 RSK (ribosomal S6 kinase) isoforms in vitro and in vivo. Biochem. J. 2007, 401, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.A.; Poteet-Smith, C.E.; Xu, Y.; Errington, T.M.; Hecht, S.M.; Lannigan, D.A. Identification of the first specific inhibitor of p90 ribosomal S6 kinase (RSK) reveals an unexpected role for RSK in cancer cell proliferation. Cancer Res. 2005, 65, 1027–1034. [Google Scholar]
- Cohen, M.S.; Zhang, C.; Shokat, K.M.; Taunton, J. Structural bioinformatics-based design of selective, irreversible kinase inhibitors. Science 2005, 308, 1318–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, S.P.; Reddy, H.; Caivano, M.; Cohen, P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem. J. 2000, 351, 95–105. [Google Scholar] [CrossRef]
- Bain, J.; McLauchlan, H.; Elliott, M.; Cohen, P. The specificities of protein kinase inhibitors: An update. Biochem. J. 2003, 371, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malakhova, M.; Kurinov, I.; Liu, K.; Zheng, D.; D’Angelo, I.; Shim, J.H.; Steinman, V.; Bode, A.M.; Dong, Z. Structural diversity of the active N-terminal kinase domain of p90 ribosomal S6 kinase 2. PLoS ONE 2009, 4, e8044. [Google Scholar] [CrossRef]
- Ikuta, M.; Kornienko, M.; Byrne, N.; Reid, J.C.; Mizuarai, S.; Kotani, H.; Munshi, S.K. Crystal structures of the N-terminal kinase domain of human RSK1 bound to three different ligands: Implications for the design of RSK1 specific inhibitors. Protein Sci. 2007, 16, 2626–2635. [Google Scholar] [CrossRef] [Green Version]
- Fisher, T.L.; Blenis, J. Evidence for two catalytically active kinase domains in pp90rsk. Mol. Cell. Biol. 1996, 16, 1212–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.K.; Berman, H.M.; Bhikadiya, C.; Bi, C.; Chen, L.; Di Costanzo, L.; Christie, C.; Dalenberg, K.; Duarte, J.M.; Dutta, S.; et al. RCSB Protein Data Bank: Biological macromolecular structures enabling research and education in fundamental biology, biomedicine, biotechnology and energy. Nucleic Acids Res. 2019, 47, D464–D474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utepbergenov, D.; Derewenda, Z.S. The unusual mechanism of inhibition of the p90 ribosomal S6 kinase (RSK) by flavonol rhamnosides. Biochim. Biophys. Acta 2013, 1834, 1285–1291. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Fu, T.M.; Nan, J.; Liu, C.; Li, L.F.; Su, X.D. Structural basis for the autoinhibition of the C-terminal kinase domain of human RSK1. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 680–685. [Google Scholar] [CrossRef]
- Serafimova, I.M.; Pufall, M.A.; Krishnan, S.; Duda, K.; Cohen, M.S.; Maglathlin, R.L.; McFarland, J.M.; Miller, R.M.; Frodin, M.; Taunton, J. Reversible targeting of noncatalytic cysteines with chemically tuned electrophiles. Nat. Chem. Biol. 2012, 8, 471–476. [Google Scholar] [CrossRef]
- Utepbergenov, D.; Derewenda, U.; Olekhnovich, N.; Szukalska, G.; Banerjee, B.; Hilinski, M.K.; Lannigan, D.A.; Stukenberg, P.T.; Derewenda, Z.S. Insights into the inhibition of the p90 ribosomal S6 kinase (RSK) by the flavonol glycoside SL0101 from the 1.5 A crystal structure of the N-terminal domain of RSK2 with bound inhibitor. Biochemistry 2012, 51, 6499–6510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derewenda, U.; Artamonov, M.; Szukalska, G.; Utepbergenov, D.; Olekhnovich, N.; Parikh, H.I.; Kellogg, G.E.; Somlyo, A.V.; Derewenda, Z.S. Identification of quercitrin as an inhibitor of the p90 S6 ribosomal kinase (RSK): Structure of its complex with the N-terminal domain of RSK2 at 1.8 A resolution. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malakhova, M.; Tereshko, V.; Lee, S.Y.; Yao, K.; Cho, Y.Y.; Bode, A.; Dong, Z. Structural basis for activation of the autoinhibitory C-terminal kinase domain of p90 RSK2. Nat. Struct. Mol. Biol. 2008, 15, 112–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein Kinase | Full Name | MZA 1 μM * | PDB ID | Predicted Binding Affinity (Kcal/mol) | |
|---|---|---|---|---|---|---|
| ATP | MZA | |||||
| 1 | MAPKAP-K1α (RSK-1) | MAPK-Activated Protein Kinase-1α | 68 | 2Z7Q | −7.6 | −9.5 |
| 2 | SGK | serum and glucocorticoid-induced kinase | 39 | 2R5T | −9.3 | −8.6 |
| 3 | LCK | lymphocyte kinase | 29 | 3MPM | −8.6 | −10.9 |
| 4 | MSK1 | mitogen and stress-activated protein kinase-1 | 27 | 3KN5 | −8.3 | −8.5 |
| 5 | GSK3β | glycogen synthase kinase-3β | 27 | 1Q5K | −7.4 | −8.5 |
| 6 | NEK 6 | NIMA-related protein kinase 6 | 26 | n/a | ||
| 7 | SAPK2α/P38 | stress-activated protein kinase-2α | 25 | 5OMH | −7.1 | −9.1 |
| 8 | PKBα | protein kinase B | 25 | 3CQW | -8.9 | −6.9 |
| 9 | JNK1a1 | c-Jun N-terminal kinase | 20 | 1UKI | −7.6 | −7.9 |
| 10 | S6K1 | p70 ribosomal protein S6 kinase | 18 | 3A60 | −7.9 | −8.6 |
| 11 | DYRK1A | dual tyrosine phosphorylated and regulated kinase 1A | 18 | 2VX3 | −8.2 | −7.9 |
| 12 | CK2 | casein kinase-2 | 17 | 1J91 | −7 | −8 |
| 13 | SAPK2β/p38β2 | stress-activated protein kinase-2β | 16 | n/a | ||
| 14 | CHK1 | checkpoint kinase-1 | 14 | 3U9N | −7.9 | −8.2 |
| 15 | CSK | C-terminal Src Kinase | 11 | 1BYG | −7.7 | −9.2 |
| 16 | MKK1 | MAPK kinase (mitogen-activated protein kinase) | 10 | 1S9J | −8.9 | −9.4 |
| 17 | PKA | cyclic AMP-dependent protein kinase | 10 | 2JDS | −9.1 | −7.9 |
| 18 | PKα | protein kinase C | 10 | 3IW4 | −8.5 | −8.7 |
| 19 | CK1 | casein kinase-1 | 10> | 1CSN | −8.3 | −8.9 |
| 20 | MAPKAP-K2 (RSK-2) | MAPK-activated protein kinase-2 | 10> | 3G51 | −9.5 | −7.6 |
| 21 | MAPK2/ERK2 | mitogen-activated protein kinase | 10> | 2OJJ | −8.6 | −7.9 |
| 22 | PRAK | p38-regulated/activated kinase | 10> | n/a | ||
| 23 | NEK2α | NIMA-related protein kinase 2α | 10> | 5M53 | −7.8 | −9.9 |
| 24 | AMPK | AMP-activated protein kinase | 10> | 2YA3 | −9 | −9.6 |
| 25 | SAPK4/P38d | stress-activated protein kinase-4 | 10> | 5EKN | −7.8 | −7.4 |
| 26 | PHK | phosphorylase kinase | 10> | 2Y7J | −7.1 | −9.6 |
| 27 | SAPK3/P38γ | stress-activated protein kinase-3 | 10> | 1CM8 | −7.5 | −11.8 |
| 28 | PDK1 | 3-phosphoinositide-dependent protein kinase-1 | 10> | 5LVL | −8.9 | −8.8 |
| 29 | ROCK-II | Rho-dependent protein kinase | 10> | 6ED6 | −8.5 | −6 |
| 30 | CDK2/cyclin A | cyclin-dependent kinase 2-cyclin A complex | 10> | 4EZ7 | −8.8 | −7.2 |
| RSK1 | RSK2 | ||
|---|---|---|---|
| CTKD | NTKD | CTKD | NTKD |
| 3RNY | 2Z7Q | 4D9T | 3G51 |
| RSK1 [32] | RSK2 [31,36] |
|---|---|
| N-Terminal | |
| Phe 73 e | Phe 79 |
| Asp 142 a | Asp 148 b |
| Leu 144 c | Leu 150 c |
| Lys 94 c | Lys 100 c |
| Asp 205 c | Asp 211 c |
| Asn 192 c | Asn 198 c |
| Val 125 e | Val 131 |
| Asp 187 d | Asp 193 |
| RSK1 [37] | RSK2 [38] |
| C-Terminal | |
| Residues around Glu496 | Equivalent residues |
| Leu 691 | Pro 696 |
| Glu 496 | Glu 500 |
| Arg 503 | Arg 507 |
| Asp 499 | Asp 503 |
| Cys 432 | Cys 436 |
| Cys 556 | Cys 560 |
| Thr 489 | Thr 493 |
| ATP-binding pocket | Equivalent residues |
| Lys 447 | Lys 451 |
| Glu 459 | Glu 463 |
| Asp 557 | Asp 561 |
| Asp 535 | Asp 539 |
| Asn 540 | Asn 544 |
| PDB ID | X-Center | Y-Center | Z-Center |
|---|---|---|---|
| 1BYG | 27.128 | 45.519 | 13.045 |
| 1CM8 | 41.771 | 75.375 | 4.443 |
| 1CSN | −24.576 | 49.359 | 51.847 |
| 1J91 | −11.675 | 3.876 | 3.645 |
| 1Q5K | 22.619 | 22.938 | 8.581 |
| 1S9J | 32.089 | 35.875 | 41.601 |
| 1UKI | 1.562 | 39.364 | 29.179 |
| 2JDS | 9.808 | 9.157 | −0.179 |
| 2OJJ | −13.28 | 13.521 | 38.972 |
| 2R5T | 33.954 | 32.472 | 64.595 |
| 2VX3 | 39.08 | 22.423 | −37.766 |
| 2Y7J | −14.917 | −6.949 | 40.443 |
| 2YA3 | 8.821 | 10.086 | 30.369 |
| 2Z7Q | 0.188 | −3.734 | 23.374 |
| 3A60 | −6.672 | 3.557 | 38.526 |
| 3CQW | 4.789 | 2.807 | 17.128 |
| 3G51 | 23.207 | 32.364 | 87.888 |
| 3IW4 | 4.829 | 30.195 | 53.056 |
| 3KN5 | 25.772 | 36.512 | 76.29 |
| 3MPM | 25.455 | 11.311 | 51.812 |
| 3U9N | 12.605 | −3.555 | 10.933 |
| 4EZ7 | −1.982 | 8.889 | −28.626 |
| 5EKN | −12.503 | 12.799 | −28.136 |
| 5LVL | −14.083 | −4.449 | −8.027 |
| 5M53 | 31.849 | −4.082 | 13.028 |
| 5OMH | −2.482 | −1.883 | −19.355 |
| 6ED6 | 26.752 | 45.831 | 53.583 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer, A.M.S.; Hall, M.L.; Lach, J.; Clifford, J.; Chandrasena, K.; Canton, C.; Kontoyianni, M.; Choo, Y.-M.; Karan, D.; Hamann, M.T. RSK1 vs. RSK2 Inhibitory Activity of the Marine β-Carboline Alkaloid Manzamine A: A Biochemical, Cervical Cancer Protein Expression, and Computational Study. Mar. Drugs 2021, 19, 506. https://doi.org/10.3390/md19090506
Mayer AMS, Hall ML, Lach J, Clifford J, Chandrasena K, Canton C, Kontoyianni M, Choo Y-M, Karan D, Hamann MT. RSK1 vs. RSK2 Inhibitory Activity of the Marine β-Carboline Alkaloid Manzamine A: A Biochemical, Cervical Cancer Protein Expression, and Computational Study. Marine Drugs. 2021; 19(9):506. https://doi.org/10.3390/md19090506
Chicago/Turabian StyleMayer, Alejandro M. S., Mary L. Hall, Joseph Lach, Jonathan Clifford, Kevin Chandrasena, Caitlin Canton, Maria Kontoyianni, Yeun-Mun Choo, Dev Karan, and Mark T. Hamann. 2021. "RSK1 vs. RSK2 Inhibitory Activity of the Marine β-Carboline Alkaloid Manzamine A: A Biochemical, Cervical Cancer Protein Expression, and Computational Study" Marine Drugs 19, no. 9: 506. https://doi.org/10.3390/md19090506
APA StyleMayer, A. M. S., Hall, M. L., Lach, J., Clifford, J., Chandrasena, K., Canton, C., Kontoyianni, M., Choo, Y. -M., Karan, D., & Hamann, M. T. (2021). RSK1 vs. RSK2 Inhibitory Activity of the Marine β-Carboline Alkaloid Manzamine A: A Biochemical, Cervical Cancer Protein Expression, and Computational Study. Marine Drugs, 19(9), 506. https://doi.org/10.3390/md19090506