
Method Development Progress in Genetic Engineering of Thraustochytrids


Abstract
:1. Introduction
2. Transforming DNA into the Cells: Methods and Considerations
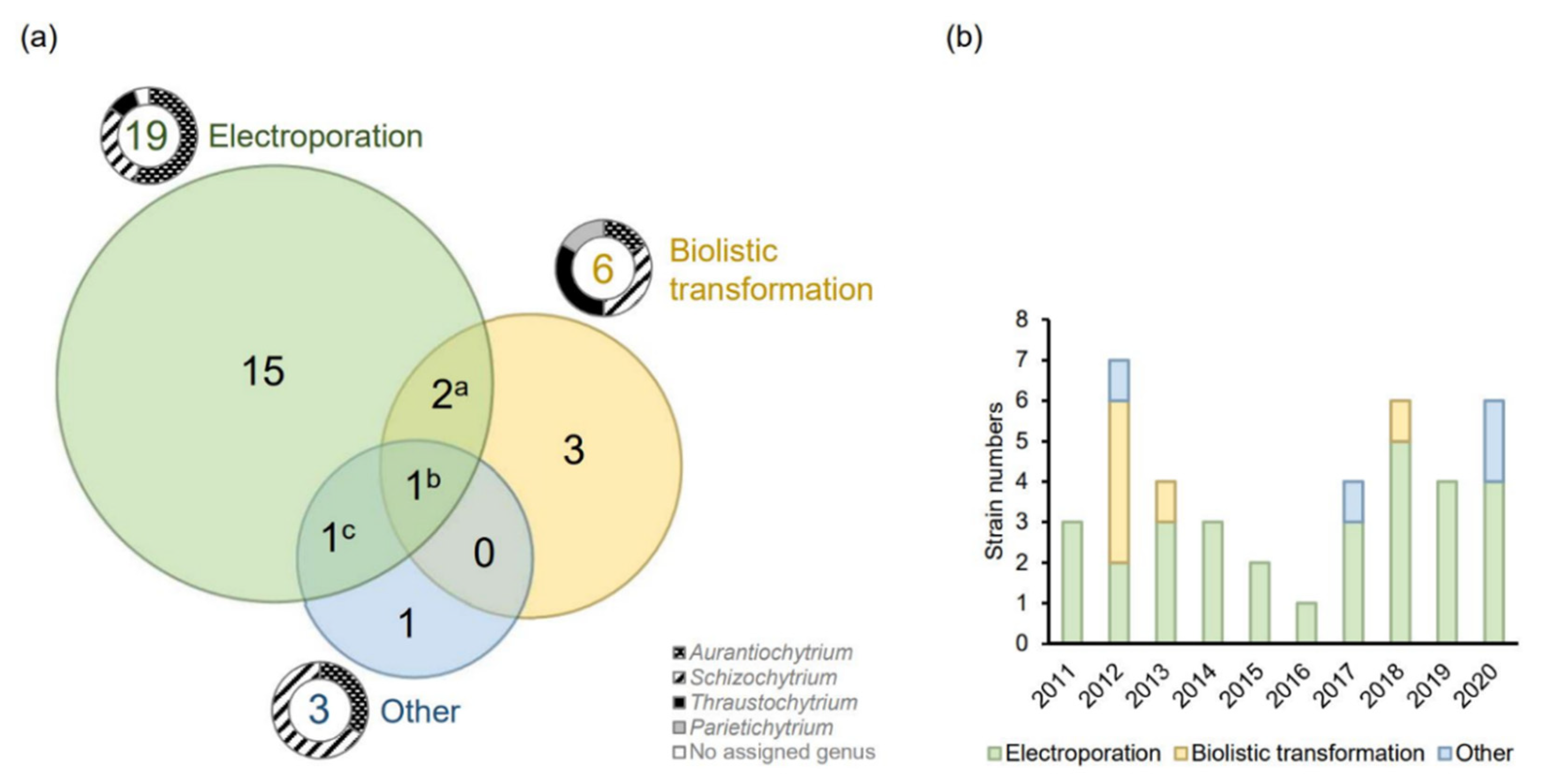
2.1. Transformation by Electroporation
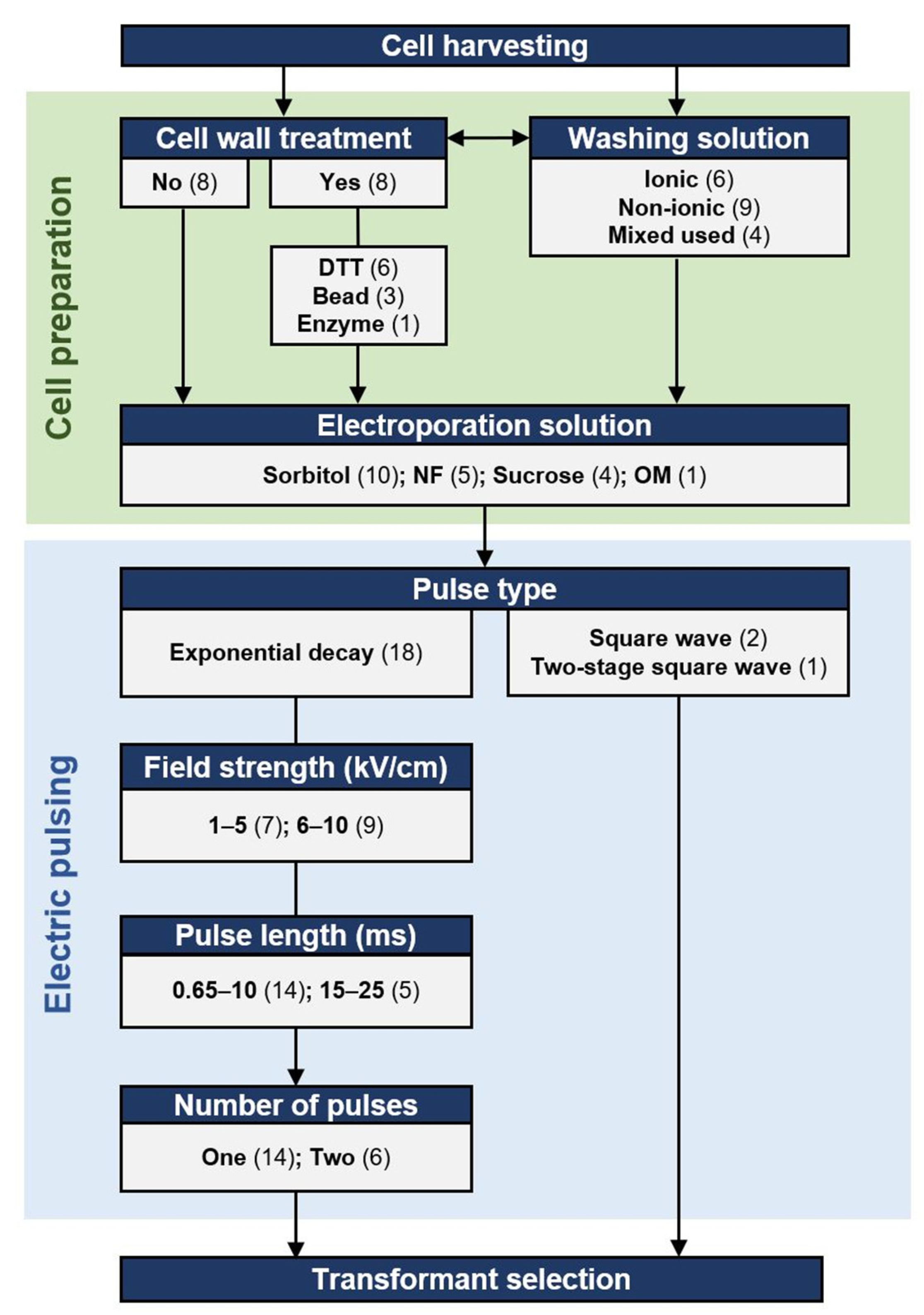
2.1.1. Pulse Types, Numbers, and the Parameters
2.1.2. Cell Wall Disruption or Removal
2.1.3. Effect of the Solutions Used to Prepare the Electrocompetent Cells
2.2. Transforming DNA into Thraustochytrid Cells by Non-Electroporation Methods
2.3. Other Strategies for Transferring DNA into Cells
3. The Properties of the DNA Affect the Outcome of the Transformation
3.1. The Presence and the Design of Homology Arms Affect Genome Integration
3.2. The Structure and Quantity of DNA Affect Transformation Frequencies
3.3. Strategies That Facilite Homologous Recombination
3.4. Application of Extrachromosomal DNAs
4. The Properties of the DNA Related to Gene Expression in Thraustochytrids
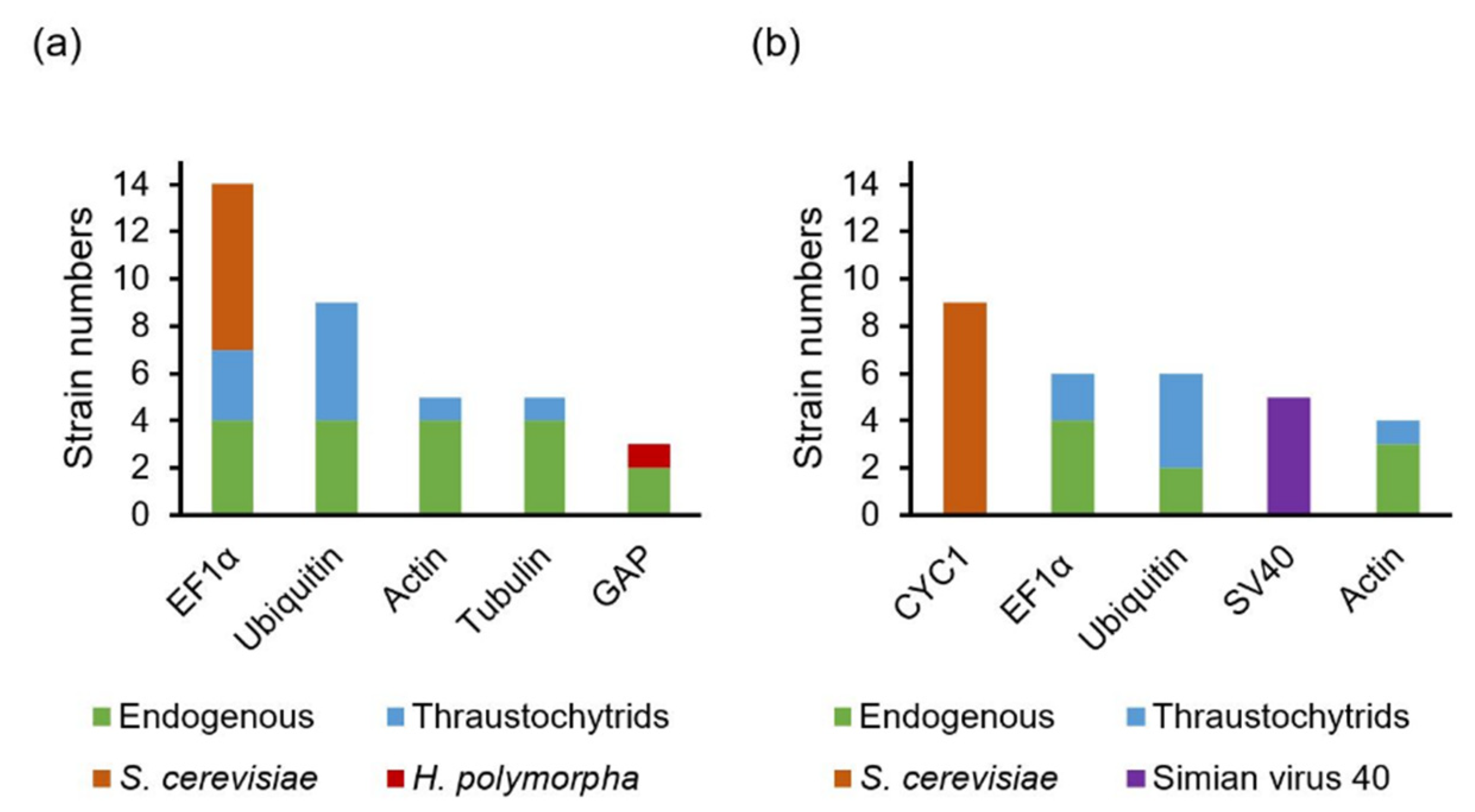
4.1. Promoters and Terminators Used for Controlling Cassette Expression Level
4.2. Antibiotics Resistance Genes for Transformant Selection in Thraustochytrids
4.3. Expression of Multiple Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Morabito, C.; Bournaud, C.; Maës, C.; Schuler, M.; Aiese Cigliano, R.; Dellero, Y.; Maréchal, E.; Amato, A.; Rébeillé, F. The lipid metabolism in thraustochytrids. Prog. Lipid Res. 2019, 76, 101007. [Google Scholar] [CrossRef]
- Yoon, H.S.; Andersen, R.A.; Boo, S.M.; Bhattacharya, D. Stramenopiles. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 721–731. [Google Scholar] [CrossRef]
- Marchan, L.F.; Chang, K.J.L.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Hauvermale, A.; Kuner, J.; Rosenzweig, B.; Guerra, D.; Diltz, S.; Metz, J.G. Fatty acid production in Schizochytrium sp.: Involvement of a polyunsaturated fatty acid synthase and a type I fatty acid synthase. Lipids 2006, 41, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Lippmeier, J.C.; Crawford, K.S.; Owen, C.B.; Rivas, A.A.; Metz, J.G.; Apt, K.E. Characterization of both polyunsaturated fatty acid biosynthetic pathways in Schizochytrium sp. Lipids 2009, 44, 621–630. [Google Scholar] [CrossRef]
- Du, F.; Wang, Y.-Z.; Xu, Y.-S.; Shi, T.-Q.; Liu, W.-Z.; Sun, X.-M.; Huang, H. Biotechnological production of lipid and terpenoid from thraustochytrids. Biotechnol. Adv. 2021, 48, 107725. [Google Scholar] [CrossRef]
- Bentsen, H. Dietary polyunsaturated fatty acids, brain function and mental health. Microb. Ecol. Health Dis. 2017, 28, 1281916. [Google Scholar] [CrossRef]
- Elagizi, A.; Lavie, C.J.; O’Keefe, E.; Marshall, K.; O’Keefe, J.H.; Milani, R.V. An update on omega-3 polyunsaturated fatty acids and cardiovascular health. Nutrients 2021, 13, 204. [Google Scholar] [CrossRef]
- Yamagata, K. Dietary docosahexaenoic acid inhibits neurodegeneration and prevents stroke. J. Neurosci. Res. 2021, 99, 561–572. [Google Scholar] [CrossRef]
- Aasen, I.M.; Ertesvåg, H.; Heggeset, T.M.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, S.; Dietrich, D.; Becker, J.; Kohlstedt, M.; Wittmann, C. Microbial production of polyunsaturated fatty acids—High-value ingredients for aquafeed, superfoods, and pharmaceuticals. Curr. Opin. Biotechnol. 2021, 69, 199–211. [Google Scholar] [CrossRef]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Huang, C.; Xu, Z.; Xu, H.; Wang, Z.; Yu, X. The strategies to reduce cost and improve productivity in DHA production by Aurantiochytrium sp.: From biochemical to genetic respects. Appl. Microbiol. Biotechnol. 2020, 104, 9433–9447. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Mining of squalene as a value-added byproduct from DHA producing marine thraustochytrid cultivated on food waste hydrolysate. Sci. Total Environ. 2020, 736, 139691. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kwak, M.; Seo, J.; Ju, J.; Heo, S.; Park, S.; Hong, W. Enhanced production of carotenoids using a Thraustochytrid microalgal strain containing high levels of docosahexaenoic acid-rich oil. Bioprocess Biosyst. Eng. 2018, 41, 1355–1370. [Google Scholar] [CrossRef]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Extracellular enzymes produced by marine eukaryotes, thraustochytrids. Biosci. Biotechnol. Biochem. 2009, 73, 180–182. [Google Scholar] [CrossRef] [Green Version]
- Nham Tran, T.L.; Miranda, A.F.; Gupta, A.; Puri, M.; Ball, A.S.; Adhikari, B.; Mouradov, A. The nutritional and pharmacological potential of new Australian thraustochytrids isolated from mangrove sediments. Mar. Drugs 2020, 18, 151. [Google Scholar] [CrossRef] [Green Version]
- Nivetha, K.; Rao, A.S.; Nair, A. Microbial production of omega-3 fatty acids: An overview. J. Appl. Microbiol. 2021. [Google Scholar] [CrossRef]
- Riley, L.A.; Guss, A.M. Approaches to genetic tool development for rapid domestication of non-model microorganisms. Biotechnol. Biofuels 2021, 14, 30. [Google Scholar] [CrossRef]
- Faktorová, D.; Nisbet, R.E.R.; Fernández Robledo, J.A.; Casacuberta, E.; Sudek, L.; Allen, A.E.; Ares, M., Jr.; Aresté, C.; Balestreri, C.; Barbrook, A.C.; et al. Genetic tool development in marine protists: Emerging model organisms for experimental cell biology. Nat. Methods 2020, 17, 481–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Sakiyama, R.; Iimi, Y.; Sekine, S.; Abe, E.; Nomura, K.H.; Nomura, K.; Ishibashi, Y.; Okino, N.; Hayashi, M.; et al. Regulation of TG accumulation and lipid droplet morphology by the novel TLDP1 in Aurantiochytrium limacinum F26-b. J. Lipid Res. 2017, 58, 2334–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nutahara, E.; Abe, E.; Uno, S.; Ishibashi, Y.; Watanabe, T.; Hayashi, M.; Okino, N.; Ito, M. The glycerol-3-phosphate acyltransferase PLAT2 functions in the generation of DHA-rich glycerolipids in Aurantiochytrium limacinum F26-b. PLoS ONE 2019, 14, e0211164. [Google Scholar] [CrossRef]
- Abe, E.; Ikeda, K.; Nutahara, E.; Hayashi, M.; Yamashita, A.; Taguchi, R.; Doi, K.; Honda, D.; Okino, N.; Ito, M. Novel lysophospholipid acyltransferase PLAT1 of Aurantiochytrium limacinum F26-b responsible for generation of palmitate-docosahexaenoate-phosphatidylcholine and phosphatidylethanolamine. PLoS ONE 2014, 9, e102377. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Sakaguchi, K.; Matsuda, T.; Abe, E.; Hama, Y.; Hayashi, M.; Honda, D.; Okita, Y.; Sugimoto, S.; Okino, N.; et al. Increase of eicosapentaenoic acid in thraustochytrids through thraustochytrid ubiquitin promoter-driven expression of a fatty acid Δ5 desaturase gene. Appl. Environ. Microbiol. 2011, 77, 3870–3876. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Aoki, K.; Okino, N.; Hayashi, M.; Ito, M. A thraustochytrid-specific lipase/phospholipase with unique positional specificity contributes to microbial competition and fatty acid acquisition from the environment. Sci. Rep. 2019, 9, 16357. [Google Scholar] [CrossRef]
- Ohara, J.; Sakaguchi, K.; Okita, Y.; Okino, N.; Ito, M. Two fatty acid elongases possessing C18-Δ6/C18-Δ9/C20-Δ5 or C16-Δ9 elongase activity in Thraustochytrium sp. ATCC 26185. Mar. Biotechnol. 2013, 15, 476–486. [Google Scholar] [CrossRef]
- Sakaguchi, K.; Matsuda, T.; Kobayashi, T.; Ohara, J.; Hamaguchi, R.; Abe, E.; Nagano, N.; Hayashi, M.; Ueda, M.; Honda, D.; et al. Versatile transformation system that is applicable to both multiple transgene expression and gene targeting for thraustochytrids. Appl. Environ. Microbiol. 2012, 78, 3193–3202. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Sakaguchi, K.; Kobayashi, T.; Abe, E.; Kurano, N.; Sato, A.; Okita, Y.; Sugimoto, S.; Hama, Y.; Hayashi, M.; et al. Molecular cloning of a Pinguiochrysis pyriformis oleate-specific microsomal Δ12-fatty acid desaturase and functional analysis in yeasts and thraustochytrids. J. Biochem. 2011, 150, 375–383. [Google Scholar] [CrossRef]
- Liu, Z.; Zang, X.; Cao, X.; Wang, Z.; Liu, C.; Sun, D.; Guo, Y.; Zhang, F.; Yang, Q.; Hou, P.; et al. Cloning of the pks3 gene of Aurantiochytrium limacinum and functional study of the 3-ketoacyl-ACP reductase and dehydratase enzyme domains. PLoS ONE 2018, 13, e0208853. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chen, H.; Zang, X.; Hou, P.; Zhou, B.; Liu, Y.; Wu, F.; Cao, X.; Zhang, X. Application of the Cre/loxP site-specific recombination system for gene transformation in Aurantiochytrium limacinum. Molecules 2015, 20, 10110–10121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okino, N.; Wakisaka, H.; Ishibashi, Y.; Ito, M. Visualization of endoplasmic reticulum and mitochondria in Aurantiochytrium limacinum by the expression of EGFP with cell organelle-specific targeting/retaining signals. Mar. Biotechnol. 2018, 20, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Sahara, T.; Okuyama, H.; Morita, N. Glass bead-based genetic transformation: An efficient method for transformation of thraustochytrid microorganisms. J. Oleo. Sci. 2017, 66, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Meng, T.; Ling, X.; Li, J.; Zheng, C.; Shi, Y.; Chen, Z.; Li, Z.; Li, Q.; Lu, Y.; et al. Overexpression of malonyl-CoA: ACP transacylase in Schizochytrium sp. to improve polyunsaturated fatty acid production. J. Agric. Food Chem. 2018, 66, 5382–5391. [Google Scholar] [CrossRef]
- Li, Z.; Chen, X.; Li, J.; Meng, T.; Wang, L.; Chen, Z.; Shi, Y.; Ling, X.; Luo, W.; Liang, D.; et al. Functions of PKS genes in lipid synthesis of Schizochytrium sp. by gene disruption and metabolomics analysis. Mar. Biotechnol. 2018, 20, 792–802. [Google Scholar] [CrossRef]
- Ling, X.; Zhou, H.; Yang, Q.; Yu, S.; Li, J.; Li, Z.; He, N.; Chen, C.; Lu, Y. Functions of enyolreductase (ER) domains of PKS cluster in lipid synthesis and enhancement of PUFAs accumulation in Schizochytrium limacinum SR21 using triclosan as a regulator of ER. Microorganisms 2020, 8, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Perez, C.M.T.; Kitahori, T.; Hata, K.; Aoi, M.; Takahashi, H.; Sakuma, T.; Okamura, Y.; Nakashimada, Y.; Yamamoto, T.; et al. Improvement of fatty acid productivity of thraustochytrid, Aurantiochytrium sp. by genome editing. J. Biosci. Bioeng. 2020, 131, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.-K.; Heo, S.-Y.; Oh, B.-R.; Kim, C.H.; Sohn, J.-H.; Yang, J.-W.; Kondo, A.; Seo, J.-W. A transgene expression system for the marine microalgae Aurantiochytrium sp. KRS101 using a mutant allele of the gene encoding ribosomal protein L44 as a selectable transformation marker for cycloheximide resistance. Bioprocess Biosyst. Eng. 2013, 36, 1191–1197. [Google Scholar] [CrossRef]
- Suen, Y.L.; Tang, H.; Huang, J.; Chen, F. Enhanced production of fatty acids and astaxanthin in Aurantiochytrium sp. by the expression of Vitreoscilla hemoglobin. J. Agric. Food Chem. 2014, 62, 12392–12398. [Google Scholar] [CrossRef]
- Liang, Y.; Liu, Y.; Tang, J.; Ma, J.; Cheng, J.J.; Daroch, M. Transcriptomic profiling and gene disruption revealed that two genes related to PUFAs/DHA biosynthesis may be essential for cell growth of Aurantiochytrium sp. Mar. Drugs 2018, 16, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.-Z.; Ma, Z.; Liu, Y.-J.; Feng, Y.; Sun, Z.; Cheng, Y.; Song, X.; Cui, Q. Overexpression of glucose-6-phosphate dehydrogenase enhanced the polyunsaturated fatty acid composition of Aurantiochytrium sp. SD116. Algal Res. 2016, 19, 138–145. [Google Scholar] [CrossRef]
- Wang, S.; Lan, C.; Wang, Z.; Wan, W.; Zhang, H.; Cui, Q.; Song, X. Optimizing eicosapentaenoic acid production by grafting a heterologous polyketide synthase pathway in the thraustochytrid Aurantiochytrium. J. Agric. Food Chem. 2020, 68, 11253–11260. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lan, C.; Wang, Z.; Wan, W.; Cui, Q.; Song, X. PUFA-synthase-specific PPTase enhanced the polyunsaturated fatty acid biosynthesis via the polyketide synthase pathway in Aurantiochytrium. Biotechnol. Biofuels 2020, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Liu, M.; He, M.; Ye, Y.; Huang, J. Illustrating and enhancing the biosynthesis of astaxanthin and docosahexaenoic acid in Aurantiochytrium sp. SK4. Mar. Drugs 2019, 17, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.-j.; Chen, S.-l.; Geng, L.-j.; Ji, X.-j.; Xu, X.; Song, P.; Gao, S.; Huang, H. Exploring the function of acyltransferase and domain replacement in order to change the polyunsaturated fatty acid profile of Schizochytrium sp. Algal Res. 2018, 29, 193–201. [Google Scholar] [CrossRef]
- Ren, L.J.; Zhuang, X.Y.; Chen, S.L.; Ji, X.J.; Huang, H. Introduction of omega-3 desaturase obviously changed the fatty acid profile and sterol content of Schizochytrium sp. J. Agric. Food Chem. 2015, 63, 9770–9776. [Google Scholar] [CrossRef]
- Zhang, S.; He, Y.; Sen, B.; Chen, X.; Xie, Y.; Keasling, J.D.; Wang, G. Alleviation of reactive oxygen species enhances PUFA accumulation in Schizochytrium sp. through regulating genes involved in lipid metabolism. Metab. Eng. Commun. 2018, 6, 39–48. [Google Scholar] [CrossRef]
- Wang, F.; Bi, Y.; Diao, J.; Lv, M.; Cui, J.; Chen, L.; Zhang, W. Metabolic engineering to enhance biosynthesis of both docosahexaenoic acid and odd-chain fatty acids in Schizochytrium sp. S31. Biotechnol. Biofuels 2019, 12, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Zhao, Z.; Wen, Y.; Chen, Z. Enhancement of docosahexaenoic acid production by overexpression of ATP-citrate lyase and acetyl-CoA carboxylase in Schizochytrium sp. Biotechnol. Biofuels 2020, 13, 131. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.-B.; Lin, X.-Z.; Wang, Z.-K.; Yang, S.-J.; Rong, H.; Ma, Y. Establishment of a transgene expression system for the marine microalga Schizochytrium by 18S rDNA-targeted homologous recombination. World J. Microbiol. Biotechnol. 2011, 27, 737–741. [Google Scholar] [CrossRef]
- Yan, J.; Cheng, R.; Lin, X.; You, S.; Li, K.; Rong, H.; Ma, Y. Overexpression of acetyl-CoA synthetase increased the biomass and fatty acid proportion in microalga Schizochytrium. Appl. Environ. Microbiol. 2013, 97, 1933–1939. [Google Scholar] [CrossRef]
- Ono, K.; Aki, T.; Kawamoto, S. Method for Introducing a Gene into Labyrinthulomycota. U.S. Patent 7888123B2, 15 February 2011. [Google Scholar]
- Rau, E.M.; Aasen, I.M.; Ertesvåg, H. A non-canonical Δ9-desaturase synthesizing palmitoleic acid identified in the thraustochytrid Aurantiochytrium sp. T66. Appl. Microbiol. Biotechnol. 2021, 105, 5931–5941. [Google Scholar] [CrossRef]
- Bayne, A.C.; Boltz, D.; Owen, C.; Betz, Y.; Maia, G.; Azadi, P.; Archer-Hartmann, S.; Zirkle, R.; Lippmeier, J.C. Vaccination against influenza with recombinant hemagglutinin expressed by Schizochytrium sp. confers protective immunity. PLoS ONE 2013, 8, e61790. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Sakaguchi, K.; Hamaguchi, R.; Kobayashi, T.; Abe, E.; Hama, Y.; Hayashi, M.; Honda, D.; Okita, Y.; Sugimoto, S.; et al. Analysis of Δ12-fatty acid desaturase function revealed that two distinct pathways are active for the synthesis of PUFAs in T. aureum ATCC 34304. J. Lipid Res. 2012, 53, 1210–1222. [Google Scholar] [CrossRef] [Green Version]
- Merkx-Jacques, A.; Rasmussen, H.; Muise, D.M.; Benjamin, J.J.R.; Kottwitz, H.; Tanner, K.; Milway, M.T.; Purdue, L.M.; Scaife, M.A.; Armenta, R.E.; et al. Engineering xylose metabolism in thraustochytrid T18. Biotechnol. Biofuels 2018, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.J.; Wang, Z.P.; Liang, M.J.; Wang, Z.; Liu, X.Y.; Hu, L.; Xia, J. One-step utilization of inulin for docosahexaenoic acid (DHA) production by recombinant Aurantiochytrium sp. carrying Kluyveromyces marxianus inulinase. Bioprocess Biosyst. Eng. 2020, 43, 1801–1811. [Google Scholar] [CrossRef]
- Hernández-Ramírez, J.; Wong-Arce, A.; González-Ortega, O.; Rosales-Mendoza, S. Expression in algae of a chimeric protein carrying several epitopes from tumor associated antigens. Int. J. Biol. Macromol. 2020, 147, 46–52. [Google Scholar] [CrossRef]
- Bañuelos-Hernández, B.; Monreal-Escalante, E.; González-Ortega, O.; Angulo, C.; Rosales-Mendoza, S. Algevir: An expression system for microalgae based on viral vectors. Front. Microbiol. 2017, 8, 1100. [Google Scholar] [CrossRef] [Green Version]
- Cheng, R.; Ma, R.; Li, K.; Rong, H.; Lin, X.; Wang, Z.; Yang, S.; Ma, Y. Agrobacterium tumefaciens mediated transformation of marine microalgae Schizochytrium. Microbiol. Res. 2012, 167, 179–186. [Google Scholar] [CrossRef]
- Kotnik, T.; Frey, W.; Sack, M.; Haberl Meglič, S.; Peterka, M.; Miklavčič, D. Electroporation-based applications in biotechnology. Trends Biotechnol. 2015, 33, 480–488. [Google Scholar] [CrossRef]
- Finer, J.J.; Finer, K.R.; Ponappa, T. Particle bombardment mediated transformation. In Plant Biotechnology: New Products and Applications; Hammond, J., McGarvey, P., Yusibov, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 59–80. [Google Scholar] [CrossRef]
- Huang, W.; Daboussi, F. Genetic and metabolic engineering in diatoms. Phil. Trans. R. Soc. B 2017, 372, 20160411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikkert, J.; Vidal, J.; Reisch, B. Stable transformation of plant cells by particle bombardment/biolistics. In Transgenic Plants: Methods and Protocols. Methods in Molecular Biology™; Peña, L., Ed.; Humana Press: Totowa, NJ, USA, 2005; Volume 286, pp. 61–78. [Google Scholar] [CrossRef]
- Michielse, C.B.; Hooykaas, P.J.; van den Hondel, C.A.; Ram, A.F. Agrobacterium-mediated transformation as a tool for functional genomics in fungi. Curr. Genet. 2005, 48, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Matamoros, M.F.; Villanueva, M.A.; Islas-Flores, T. Genetic transformation of cell-walled plant and algae cells: Delivering DNA through the cell wall. Brief. Funct. Genom. 2017, 17, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Reberšek, M.; Miklavčič, D. Advantages and disadvantages of different concepts of electroporation pulse generation. Automatika 2011, 52, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Dower, W.J.; Miller, J.F.; Ragsdale, C.W. High efficiency transformation of E. coli by high voltage electroporation. Nucleic Acids Res. 1988, 16, 6127–6145. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Electroporation. Cold Spring Harb. Protoc. 2019, 096271. [Google Scholar] [CrossRef] [Green Version]
- Hattermann, D.R.; Stacey, G. Efficient DNA transformation of Bradyrhizobium japonicum by electroporation. Appl. Environ. Microbiol. 1990, 56, 833–836. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.E.; Sprecher, S.L.; Keller, L.R. Introduction of exogenous DNA into Chlamydomonas reinhardtii by electroporation. Mol. Cell Biol. 1991, 11, 2328–2332. [Google Scholar] [CrossRef] [Green Version]
- Demiryurek, Y.; Nickaeen, M.; Zheng, M.; Yu, M.; Zahn, J.D.; Shreiber, D.I.; Lin, H.; Shan, J.W. Transport, resealing, and re-poration dynamics of two-pulse electroporation-mediated molecular delivery. Biochim. Biophys. Acta Biomembr. 2015, 1848, 1706–1714. [Google Scholar] [CrossRef] [Green Version]
- Satkauskas, S.; Bureau, M.F.; Puc, M.; Mahfoudi, A.; Scherman, D.; Miklavcic, D.; Mir, L.M. Mechanisms of in vivo DNA electrotransfer: Respective contributions of cell electropermeabilization and DNA electrophoresis. Mol. Ther. 2002, 5, 133–140. [Google Scholar] [CrossRef]
- Kandušer, M.; Miklavčič, D.; Pavlin, M. Mechanisms involved in gene electrotransfer using high- and low-voltage pulses—An in vitro study. Bioelectrochemistry 2009, 74, 265–271. [Google Scholar] [CrossRef]
- Zheng, H.-Z.; Liu, H.-H.; Chen, S.-X.; Lu, Z.-X.; Zhang, Z.-L.; Pang, D.-W.; Xie, Z.-X.; Shen, P. Yeast transformation process studied by fluorescence labeling technique. Bioconjug. Chem. 2005, 16, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Jeon, K.; Suresh, A.; Kim, Y.-C. Highly efficient molecular delivery into Chlamydomonas reinhardtii by electroporation. Korean J. Chem. Eng. 2013, 30, 1626–1630. [Google Scholar] [CrossRef]
- Muñoz, C.F.; de Jaeger, L.; Sturme, M.H.J.; Lip, K.Y.F.; Olijslager, J.W.J.; Springer, J.; Wolbert, E.J.H.; Martens, D.E.; Eggink, G.; Weusthuis, R.A.; et al. Improved DNA/protein delivery in microalgae—A simple and reliable method for the prediction of optimal electroporation settings. Algal Res. 2018, 33, 448–455. [Google Scholar] [CrossRef]
- Azencott, H.R.; Peter, G.F.; Prausnitz, M.R. Influence of the cell wall on intracellular delivery to algal cells by electroporation and sonication. Ultrasound Med. Biol. 2007, 33, 1805–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, B.; Eu, Y.-J.; Jeong, W.-J.; Kim, D.-P. Droplet electroporation in microfluidics for efficient cell transformation with or without cell wall removal. Lab Chip 2012, 12, 4483–4488. [Google Scholar] [CrossRef] [Green Version]
- De Nobel, J.G.; Dijkers, C.; Hooijberg, E.; Klis, F.M. Increased cell wall porosity in Saccharomyces cerevisiae after treatment with dithiothreitol or EDTA. J. Gen. Microbiol. 1989, 135, 2077–2084. [Google Scholar] [CrossRef] [Green Version]
- Ponton, J.; Jones, J.M. Analysis of cell wall extracts of Candida albicans by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and Western blot techniques. Infect. Immun. 1986, 53, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, S.; Hashimoto, W.; Murata, K. Transformation of Saccharomyces cerevisiae and other fungi. Bioeng. Bugs 2010, 1, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Kong, R.; Hu, L. Preparation of protoplasts from Chlorella protothecoides. World J. Microbiol. Biotechnol. 2012, 28, 1827–1830. [Google Scholar] [CrossRef]
- Braun, E.; Aach, H.G. Enzymatic degradation of the cell wall of Chlorella. Planta 1975, 126, 181–185. [Google Scholar] [CrossRef]
- Costanzo, M.C.; Fox, T.D. Transformation of yeast by agitation with glass beads. Genetics 1988, 120, 667–670. [Google Scholar] [CrossRef]
- Kindle, K.L. High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1990, 87, 1228–1232. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, G.L.; Panda, T. Electroporation: Basic principles, practical considerations and applications in molecular biology. Bioprocess Eng. 1997, 16, 261–264. [Google Scholar] [CrossRef]
- Benatuil, L.; Perez, J.M.; Belk, J.; Hsieh, C.-M. An improved yeast transformation method for the generation of very large human antibody libraries. Protein Eng. Des. Sel. 2010, 23, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.R.; Lundgren, B.R.; Grzeskowiak, B.R.; Mizuno, K.; Nomura, C.T. A rapid and efficient electroporation method for transformation of Halomonas sp. O-1. J. Microbiol. Methods 2016, 129, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, C.F.; Sturme, M.H.J.; D’Adamo, S.; Weusthuis, R.A.; Wijffels, R.H. Stable transformation of the green algae Acutodesmus obliquus and Neochloris oleoabundans based on E. coli conjugation. Algal Res. 2019, 39, 101453. [Google Scholar] [CrossRef]
- Sharma, A.K.; Nymark, M.; Sparstad, T.; Bones, A.M.; Winge, P. Transgene-free genome editing in marine algae by bacterial conjugation—Comparison with biolistic CRISPR/Cas9 transformation. Sci. Rep. 2018, 8, 14401. [Google Scholar] [CrossRef] [PubMed]
- Karas, B.J.; Diner, R.E.; Lefebvre, S.C.; McQuaid, J.; Phillips, A.P.R.; Noddings, C.M.; Brunson, J.K.; Valas, R.E.; Deerinck, T.J.; Jablanovic, J.; et al. Designer diatom episomes delivered by bacterial conjugation. Nat. Commun. 2015, 6, 6925. [Google Scholar] [CrossRef]
- Wang, S.; Lee, L.J. Micro-/nanofluidics based cell electroporation. Biomicrofluidics 2013, 7, 011301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, S.C.C.; Goyal, G.; Kim, P.W.; Koutsoubelis, N.; Keasling, J.D.; Adams, P.D.; Hillson, N.J.; Singh, A.K. A versatile microfluidic device for automating synthetic biology. ACS Synth. Biol. 2015, 4, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, M.B.; Esveld, D.C.; Valero, A.; Luttge, R.; Mastwijk, H.C.; Bartels, P.V.; van den Berg, A.; Boom, R.M. Electroporation of cells in microfluidic devices: A review. Anal. Bioanal. Chem. 2006, 385, 474. [Google Scholar] [CrossRef] [Green Version]
- Im, D.J.; Jeong, S.-N.; Yoo, B.S.; Kim, B.; Kim, D.-P.; Jeong, W.-J.; Kang, I.S. Digital microfluidic approach for efficient electroporation with high productivity: Transgene expression of microalgae without cell wall removal. Anal. Chem. 2015, 87, 6592–6599. [Google Scholar] [CrossRef]
- Kim, G.C.; Cheon, D.H.; Lee, Y. Challenge to overcome current limitations of cell-penetrating peptides. Biochim. Biophys. Acta Proteins Proteom. 2021, 1869, 140604. [Google Scholar] [CrossRef] [PubMed]
- Gadamchetty, P.; Mullapudi, P.L.V.; Sanagala, R.; Markandan, M.; Polumetla, A.K. Genetic transformation of Chlorella vulgaris mediated by HIV-TAT peptide. 3 Biotech 2019, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Jeon, S.; Kim, S.; Chang, Y.K.; Kim, Y.-C. Development of a pVEC peptide-based ribonucleoprotein (RNP) delivery system for genome editing using CRISPR/Cas9 in Chlamydomonas reinhardtii. Sci. Rep. 2020, 10, 22158. [Google Scholar] [CrossRef]
- Zorin, B.; Lu, Y.; Sizova, I.; Hegemann, P. Nuclear gene targeting in Chlamydomonas as exemplified by disruption of the PHOT gene. Gene 2009, 432, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-T.; Jiang, J.-Y.; Shi, T.-Q.; Sun, X.-M.; Zhao, Q.-Y.; Huang, H.; Ren, L.-J. Application of the CRISPR/Cas system for genome editing in microalgae. Appl. Microbiol. Biotechnol. 2019, 103, 3239–3248. [Google Scholar] [CrossRef] [PubMed]
- Diner, R.E.; Bielinski, V.A.; Dupont, C.L.; Allen, A.E.; Weyman, P.D. Refinement of the diatom episome maintenance sequence and improvement of conjugation-based DNA delivery methods. Front. Bioeng. Biotechnol. 2016, 4, 65. [Google Scholar] [CrossRef] [Green Version]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef]
- Angstenberger, M.; Krischer, J.; Aktaş, O.; Büchel, C. Knock-down of a ligIV homologue enables DNA integration via homologous recombination in the marine diatom Phaeodactylum tricornutum. ACS Synth. Biol. 2019, 8, 57–69. [Google Scholar] [CrossRef]
- Plecenikova, A.; Mages, W.; Andrésson, Ó.S.; Hrossova, D.; Valuchova, S.; Vlcek, D.; Slaninova, M. Studies on recombination processes in two Chlamydomonas reinhardtii endogenous genes, NIT1 and ARG7. Protist 2013, 164, 570–582. [Google Scholar] [CrossRef]
- Jiang, W.Z.; Weeks, D.P. A gene-within-a-gene Cas9/sgRNA hybrid construct enables gene editing and gene replacement strategies in Chlamydomonas reinhardtii. Algal Res. 2017, 26, 474–480. [Google Scholar] [CrossRef]
- Ferenczi, A.; Pyott, D.E.; Xipnitou, A.; Molnar, A. Efficient targeted DNA editing and replacement in Chlamydomonas reinhardtii using Cpf1 ribonucleoproteins and single-stranded DNA. Proc. Natl. Acad. Sci. USA 2017, 114, 13567–13572. [Google Scholar] [CrossRef] [Green Version]
- Belshaw, N.; Grouneva, I.; Aram, L.; Gal, A.; Hopes, A.; Mock, T. Efficient CRISPR/Cas-mediated homologous recombination in the model diatom Thalassiosira pseudonana. bioRxiv 2017, 215582. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Gao, M.; Lopez-Garcia, C.L.; Wu, Y.; Seetharam, A.S.; Severin, A.J.; Shao, Z. Centromeric DNA facilitates nonconventional yeast genetic engineering. ACS Synth. Biol. 2017, 6, 1545–1553. [Google Scholar] [CrossRef]
- Nymark, M.; Sharma, A.K.; Sparstad, T.; Bones, A.M.; Winge, P. A CRISPR/Cas9 system adapted for gene editing in marine algae. Sci. Rep. 2016, 6, 24951. [Google Scholar] [CrossRef] [PubMed]
- Hopes, A.; Nekrasov, V.; Kamoun, S.; Mock, T. Editing of the urease gene by CRISPR-Cas in the diatom Thalassiosira pseudonana. Plant Methods 2016, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Brueggeman, A.J.; Horken, K.M.; Plucinak, T.M.; Weeks, D.P. Successful transient expression of Cas9 and single guide RNA genes in Chlamydomonas reinhardtii. Eukaryot. Cell 2014, 13, 1465–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partow, S.; Siewers, V.; Bjørn, S.; Nielsen, J.; Maury, J. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae. Yeast 2010, 27, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Shao, Z.; Zhao, H.; Nair, N.; Wen, F.; Xu, J.H.; Zhao, H. Cloning and characterization of a panel of constitutive promoters for applications in pathway engineering in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2012, 109, 2082–2092. [Google Scholar] [CrossRef] [PubMed]
- Redden, H.; Morse, N.; Alper, H.S. The synthetic biology toolbox for tuning gene expression in yeast. FEMS Yeast Res. 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.-G.; Chen, J.-W.; Zheng, D.-L.; Balamurugan, S.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. High-efficiency promoter-driven coordinated regulation of multiple metabolic nodes elevates lipid accumulation in the model microalga Phaeodactylum tricornutum. Microb. Cell Fact. 2018, 17, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, M.; Díaz-Santos, E.; De la Vega, M.; Rodríguez, H.; Vargas, Á.; León, R. Promoter trapping in microalgae using the antibiotic paromomycin as selective agent. Mar. Drugs 2012, 10, 2749–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanishi, M.; Ito, Y.; Kintaka, R.; Imamura, C.; Katahira, S.; Ikeuchi, A.; Moriya, H.; Matsuyama, T. A genome-wide activity assessment of terminator regions in Saccharomyces cerevisiae provides a “terminatome“ toolbox. ACS Synth. Biol. 2013, 2, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.R.; Busser, K.; Comiskey, J.; Whittier, P.S.; Purcell, T.J.; Hoeffler, J.P. Small vectors for expression based on dominant drug resistance with direct multicopy selection. In Pichia Protocols. Methods in Molecular Biology™; Higgins, D.R., Cregg, J.M., Eds.; Humana Press: Totowa, NJ, USA, 1998; Volume 103, pp. 41–53. [Google Scholar] [CrossRef]
- Napierala, M.; Parniewski, P.; Pluciennik, A.; Wells, R.D. Long CTG.CAG repeat sequences markedly stimulate intramolecular recombination. J. Biol. Chem. 2002, 277, 34087–34100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Chen, O.; Wall, J.B.J.; Zheng, M.; Zhou, Y.; Wang, L.; Ruth Vaseghi, H.; Qian, L.; Liu, J. Systematic comparison of 2A peptides for cloning multi-genes in a polycistronic vector. Sci. Rep. 2017, 7, 2193. [Google Scholar] [CrossRef]
- Falciatore, A.; Casotti, R.; Leblanc, C.; Abrescia, C.; Bowler, C. Transformation of nonselectable reporter genes in marine diatoms. Mar. Biotechnol. 1999, 1, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.H.; Loke, S.; Croft, L.J.; Gleason, F.H.; Lange, L.; Pilgaard, B.; Trevathan-Tackett, S.M. First genome of Labyrinthula sp., an opportunistic seagrass pathogen, reveals novel Insight into marine protist phylogeny, ecology and CAZyme cell-wall degradation. Microb. Ecol. 2021, 82, 498–511. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Zeocin | Hygromycin | G418 | Blasticidin | Other | Reference |
|---|---|---|---|---|---|---|
| A. limacinum F26-b | 2000 | 500 | [21,22,23] | |||
| A. limacinum mh0186 | 500 * | 1000; 2000 * | 500 | 1200 | 500 (neomycin) | [24,25,26,27,28] |
| A. limacinum OUC168 | 5 | 100 (chloramphenicol) | [29] | |||
| A. limacinum OUC88 | 5 | 100 (chloramphenicol) | [30] | |||
| A. limacinum SR21 | 30; 50; 100 | 200 | 500 | [20,31,32,33,34,35,36,52] | ||
| Aurantiochytrium sp. KRS101 | 30 (cycloheximide) | [37] | ||||
| Aurantiochytrium sp. MP4 | 50 | [38] | ||||
| Aurantiochytrium sp. PKU#SW7 | 500 | 500 | [39] | |||
| Aurantiochytrium sp. RH-7A | 100 | [36] | ||||
| Aurantiochytrium sp. SD116 | 30; 50; 100 | 500 * | 50 *; 50 | 100 *(anhydrotetracycline) | [40,41,42] | |
| Aurantiochytrium sp. SK4 | 50 | [38,43] | ||||
| Aurantiochytrium sp. YLH70 | 15 | [56] | ||||
| Parietichytrium sp. TA04Bb | 2000 | 500 | 800 | [27] | ||
| Schizochytrium sp. S31 | 40; 50 | 100 | 50 (bleomycin); 250 (cefotaxime); 50 (paromomycin) | [5,47,48,53,57,58] | ||
| Schizochytrium sp. HX-308 | 1.5; 20 | [44,45] | ||||
| Schizochytrium sp. PKU#Mn4 | 800 | [46] | ||||
| Schizochytrium sp. TIO01 | 100 | [49] | ||||
| Schizochytrium sp. TIO1101 | 300 | [50,59] | ||||
| Schizochytrium sp. SEK 579 | 2000 | 500 | [27] | |||
| Schizochytrium sp. CB15-5 | 20 | [51] | ||||
| Thraustochytrid strain 12B | 500 | [32] | ||||
| T. aureum ATCC 34304 | 2000 | 1000; 2000 | 200–400 | [27,54] | ||
| Thraustochytrium sp. ONC-T18 | 250 | 400 | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rau, E.-M.; Ertesvåg, H. Method Development Progress in Genetic Engineering of Thraustochytrids. Mar. Drugs 2021, 19, 515. https://doi.org/10.3390/md19090515
Rau E-M, Ertesvåg H. Method Development Progress in Genetic Engineering of Thraustochytrids. Marine Drugs. 2021; 19(9):515. https://doi.org/10.3390/md19090515
Chicago/Turabian StyleRau, E-Ming, and Helga Ertesvåg. 2021. "Method Development Progress in Genetic Engineering of Thraustochytrids" Marine Drugs 19, no. 9: 515. https://doi.org/10.3390/md19090515
APA StyleRau, E.-M., & Ertesvåg, H. (2021). Method Development Progress in Genetic Engineering of Thraustochytrids. Marine Drugs, 19(9), 515. https://doi.org/10.3390/md19090515