
Comparative Metabolomics Reveals Fungal Conversion of Co-Existing Bacterial Metabolites within a Synthetic Aspergillus-Streptomyces Community

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
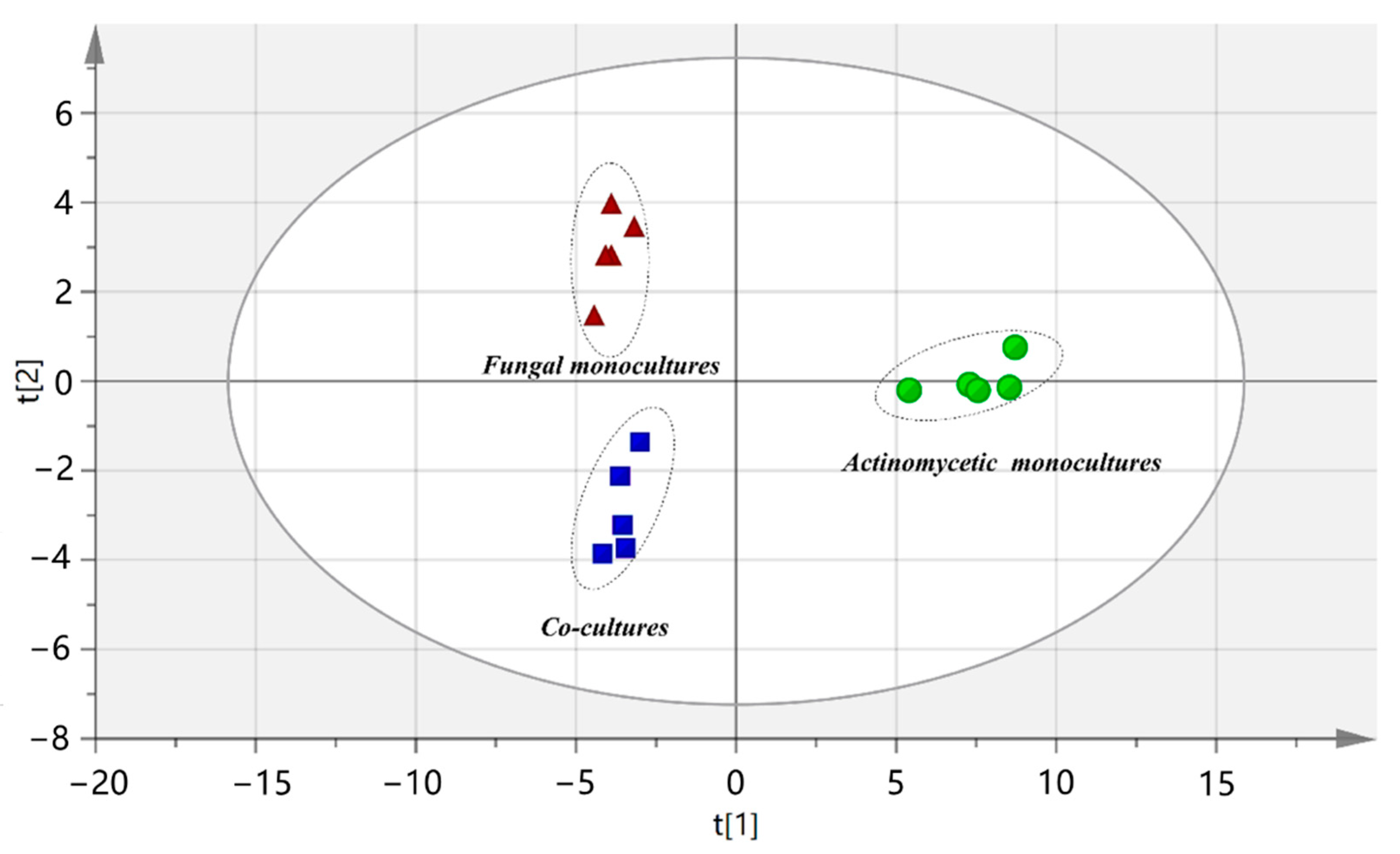
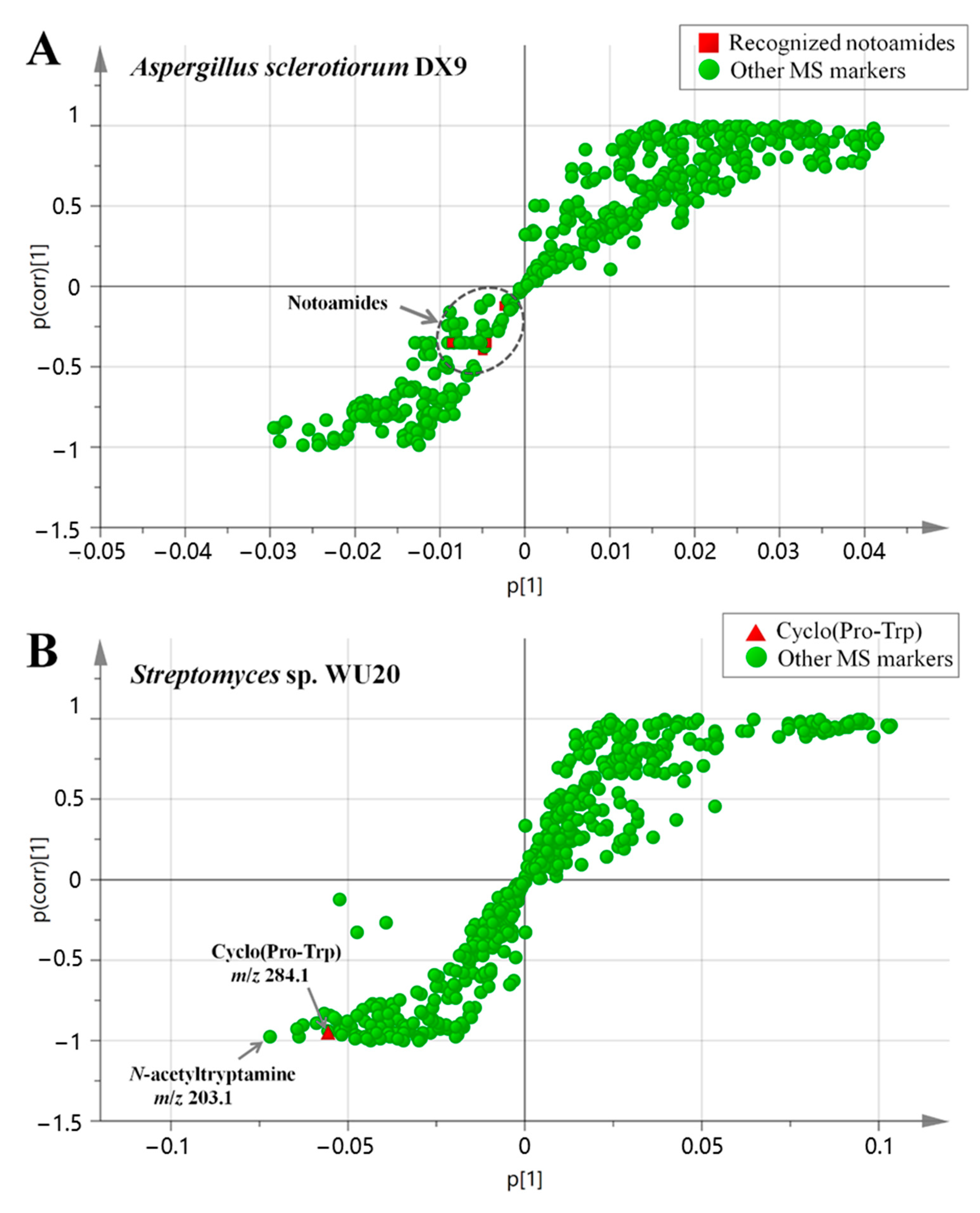
2.1. Mass Spectrometry-Based Comparative Metabolomics Profiling of Synthetic Multispecies Microbial Communities Locating Major Discriminating Compounds Induced by Metabolically-Mediated Interactions
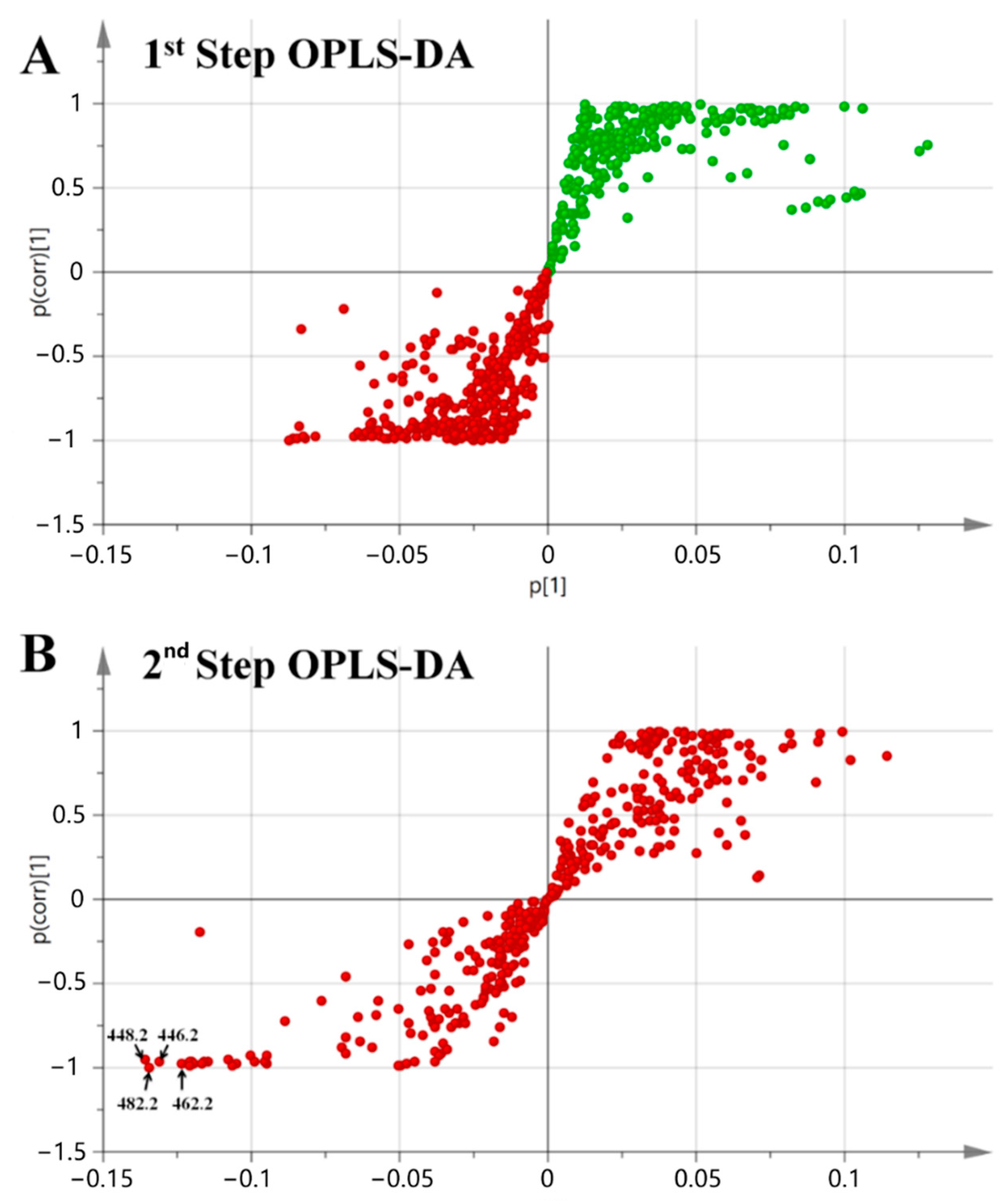
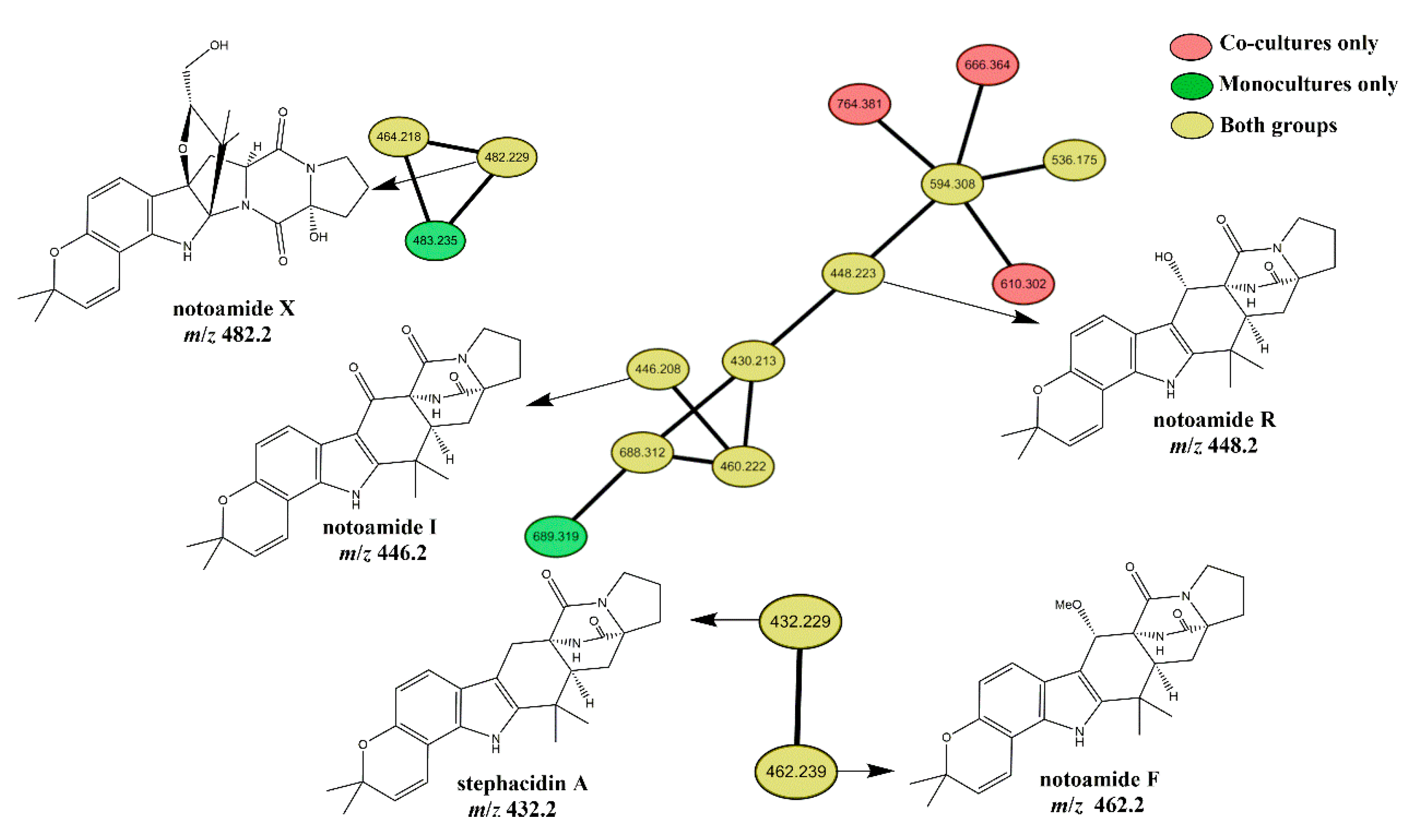
2.2. Mass Spectral Molecular Networking Facilitating Structural Elucidation of Target Co-Culture-Induced Metabolites
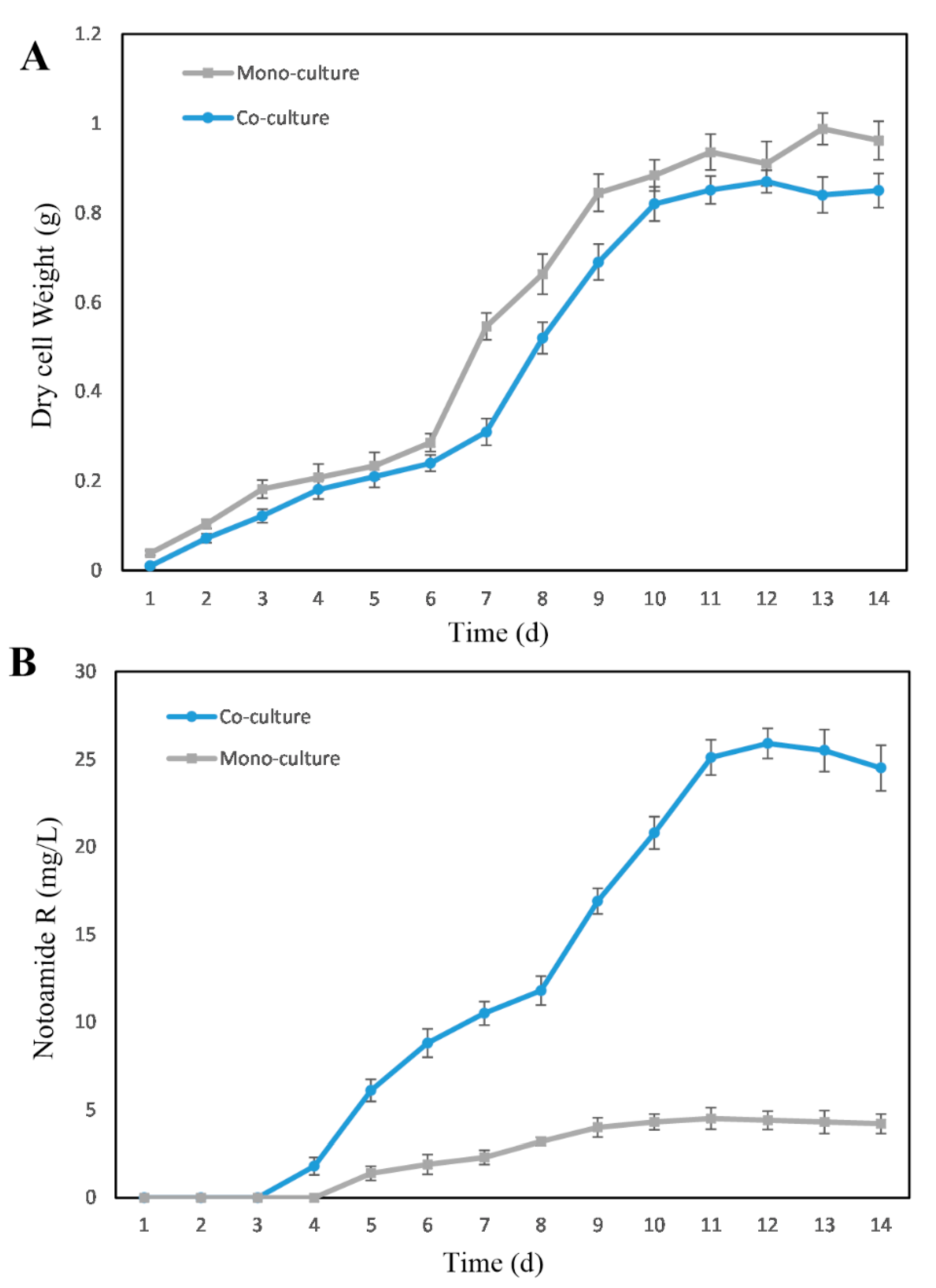
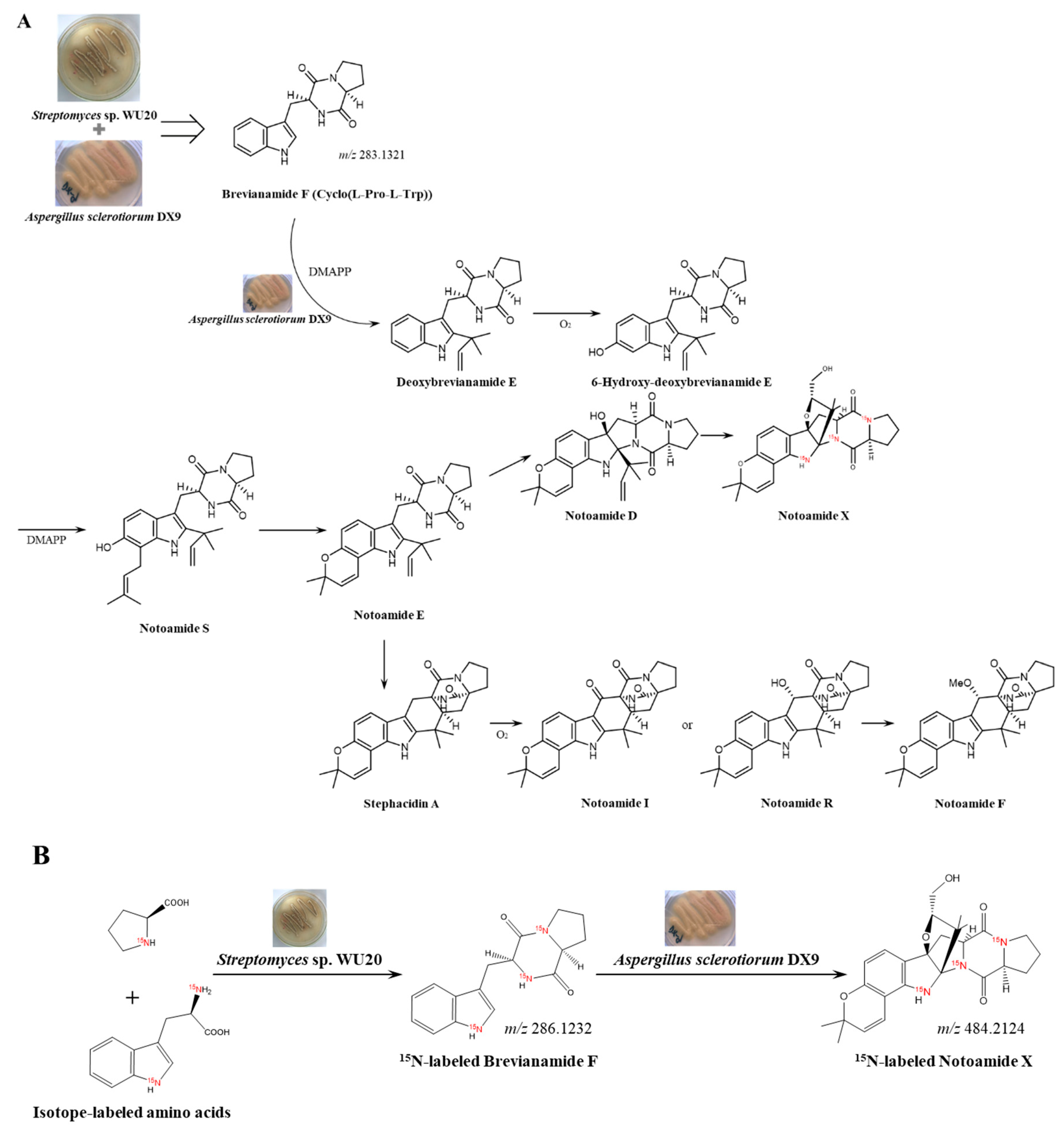
2.3. Induction-Effect-Guided Isolation Revealing Cyclo(Pro-Trp) Secreted by Streptomyces as Potential Molecular Inducers of Fungal Secondary Metabolites in the Co-Cultures
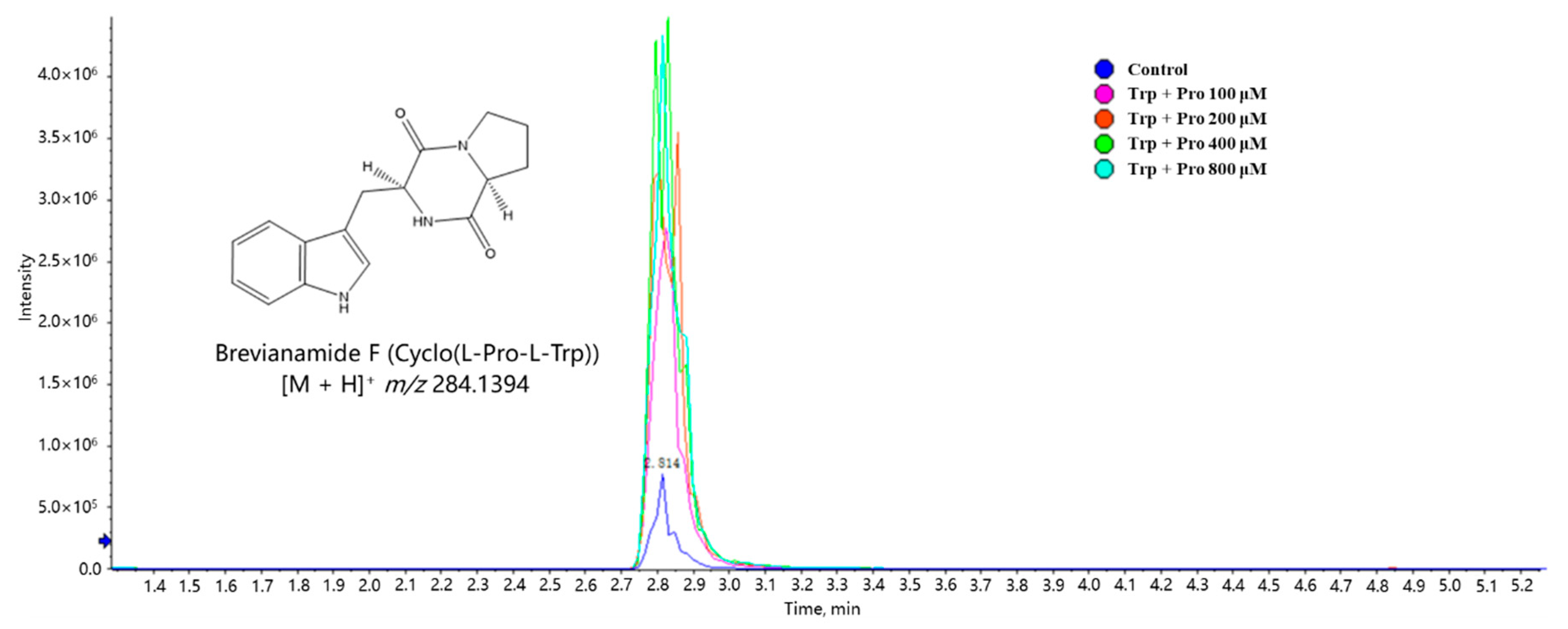
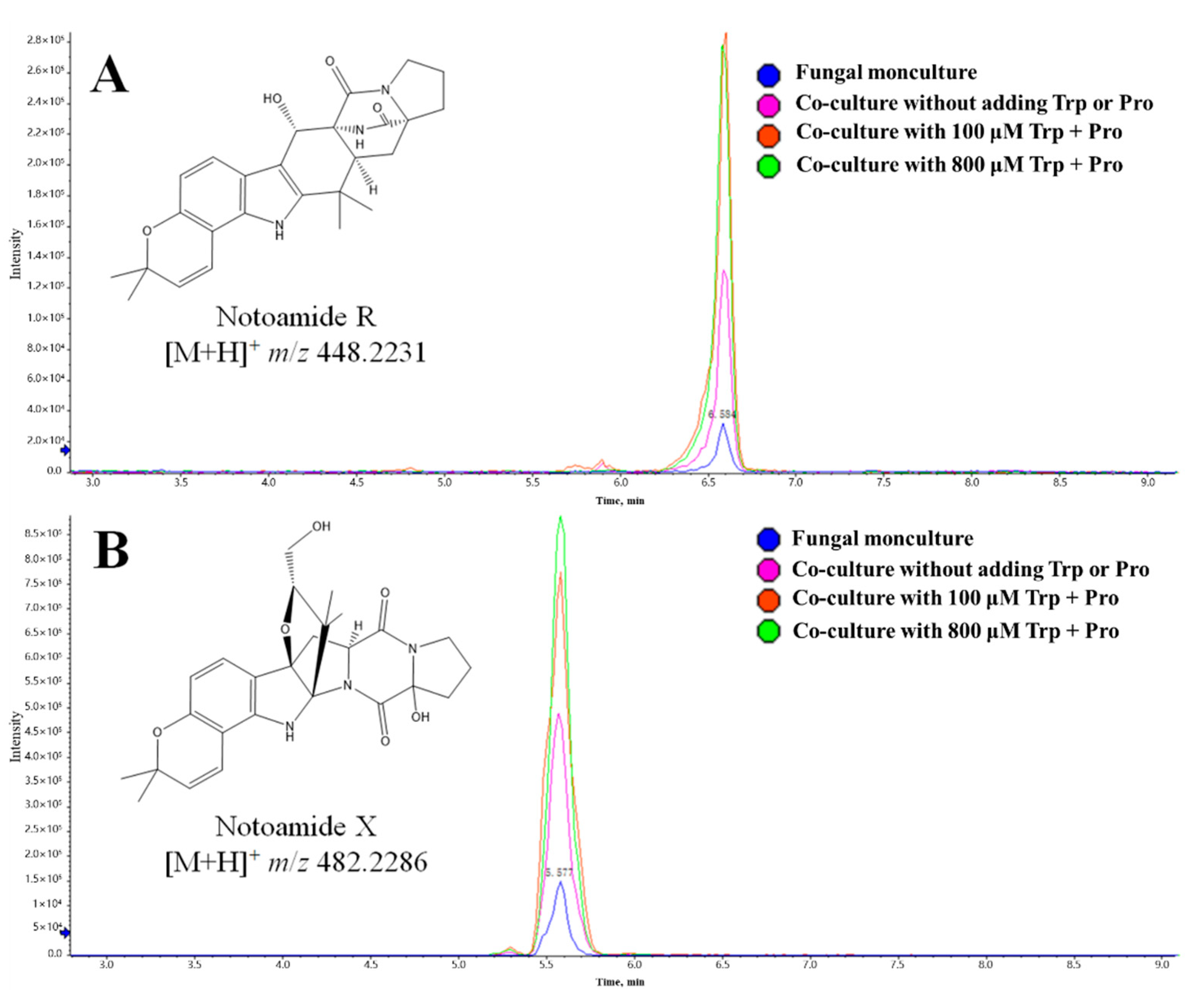
2.4. Further Investigation on the Effects of Exogenous Amino Acids on the Metabolism of Both Monocultures and Co-Cultures
3. Materials and Methods
3.1. Hydrothermal Vent Microbial Strains Isolation
3.2. Culturing Conditions for Co-Cultures
3.3. Metabolomics Sampling and UHPLC-ESI-HR-MS Analysis
3.4. Data Processing, Molecular Networking and Multivariate Data Analysis
3.5. Induction Effect-Guided Isolation of Cyclo(L-Pro-L-Trp)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Connor, S.E. Engineering of secondary metabolism. Annu. Rev. Genet. 2015, 49, 71–94. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Netzker, T.; Flak, M.; Krespach, M.K.; Stroe, M.C.; Weber, J.; Schroeckh, V.; Brakhage, A.A. Microbial interactions trigger the production of antibiotics. Curr. Opin. Microbiol. 2018, 45, 117–123. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodhi, A.F.; Zhang, Y.; Adil, M.; Deng, Y. Antibiotic discovery: Combining isolation chip (iChip) technology and co-culture technique. Appl. Microbiol. Biotechnol. 2018, 102, 7333–7341. [Google Scholar] [CrossRef] [PubMed]
- Toghueo, R.; Sahal, D.; Boyom, F.F. Recent advances in inducing endophytic fungal specialized metabolites using small molecule elicitors including epigenetic modifiers. Phytochemistry 2020, 174, 112338. [Google Scholar] [CrossRef]
- Jones, J.A.; Wang, X. Use of bacterial co-cultures for the efficient production of chemicals. Curr. Opin. Biotechnol. 2018, 53, 33–38. [Google Scholar] [CrossRef]
- Arora, D.; Gupta, P.; Jaglan, S.; Roullier, C.; Grovel, O.; Bertrand, S. Expanding the chemical diversity through microorganisms co-culture: Current status and outlook. Biotechnol. Adv. 2020, 40, 107521. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kakeya, H. Cryptic Chemical Communication: Secondary Metabolic Responses Revealed by Microbial Co-culture. Chem Asian J. 2020, 15, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Salim, A.A.; Capon, R.J. Chaunopyran A: Co-Cultivation of Marine dMollusk Derived Fungi Activates a Rare Class of 2-Alkenyl-Tetrahydropyran. J. Nat. Prod. 2017, 80, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Vallet, M.; Vanbellingen, Q.P.; Fu, T.; Le Caer, J.P.; Della-Negra, S.; Touboul, D.; Duncan, K.R. An Integrative Approach to Decipher the Chemical Antagonism between the Competing Endophytes Paraconiothyrium variabile and Bacillus subtilis. J. Nat. Prod. 2017, 80, 2863–2873. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Kim, H.K.; van Wezel, G.P.; Choi, Y.H. Metabolomics in the natural products field-a gateway to novel antibiotics. Drug Discov. Today 2015, 13, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Krug, D.; Hüttel, S.; Müller, R. Improving natural products identification through targeted LC-MS/MS in an untargeted secondary metabolomics workflow. Anal. Chem. 2014, 86, 10780–10788. [Google Scholar] [CrossRef] [PubMed]
- Covington, B.C.; Spraggins, J.M.; Ynigez-Gutierrez, A.E.; Hylton, Z.B.; Bachmann, B.O. Response of Secondary Metabolism of Hypogean Actinobacterial Genera to Chemical and Biological Stimuli. Appl. Environ. Microbiol. 2018, 84, e01125-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, C.; Mausz, M.A.; Pohnert, G. A co-culturing/metabolomics approach to investigate chemically mediated interactions of planktonic organisms reveals influence of bacteria on diatom metabolism. Metabolomics 2013, 9, 349–359. [Google Scholar] [CrossRef]
- Fischbach, M.A.; Segre, J.A. Signaling in host-associated microbial communities. Cell 2016, 164, 1288–1300. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Pan, C.; Wang, K.; Chen, X.; Wu, X.; Chen, C.A.; Wu, B. Synthetic multispecies microbial communities reveals shifts in secondary metabolism and facilitates cryptic natural product discovery. Environ. Microbiol. 2017, 19, 3606–3618. [Google Scholar] [CrossRef]
- Shi, Y.; Pan, C.; Cen, S.; Fu, L.; Cao, X.; Wang, H.; Wu, B. Comparative metabolomics reveals defence-related modification of citrinin by Penicillium citrinum within a synthetic Penicillium-Pseudomonas community. Environ. Microbiol. 2019, 21, 496–510. [Google Scholar] [CrossRef] [Green Version]
- Nordström, A.; Want, E.; Northen, T.; Lehtiö, J.; Siuzdak, G. Multiple ionization mass spectrometry strategy used to reveal the complexity of metabolomics. Anal. Chem. 2008, 80, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Choi, Y.H.; Verberne, M.; Lefeber, A.W.; Erkelens, C.; Verpoorte, R. Metabolic fingerprinting of wild type and transgenic tobacco plants by 1H NMR and multivariate analysis technique. Phytochemistry 2004, 65, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Bylesjö, M.; Rantalainen, M.; Cloarec, O.; Nicholson, J.K.; Holmes, E.; Trygg, J. OPLS discriminant analysis: Combining the strengths of PLS-DA and SIMCA classification. J. Chemometr. 2006, 20, 341–351. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Umaoka, H.; Yoshikawa, K.; Ikeda, T.; Hirota, H. Notoamide O, a structurally unprecedented prenylated indole alkaloid, and notoamides P-R from a marine-derived fungus, Aspergillus sp. J. Nat. Prod. 2010, 73, 1438–1440. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Kato, H.; Samizo, M.; Nojiri, Y.; Onuki, H.; Hirota, H.; Ohta, T. Notoamides F-K, prenylated indole alkaloids isolated from a marine-derived Aspergillus sp. J. Nat. Prod. 2008, 71, 2064–2067. [Google Scholar] [CrossRef]
- Maiya, S.; Grundmann, A.; Li, S.M.; Turner, G. The fumitremorgin gene cluster of Aspergillus fumigatus: Identification of a gene encoding brevianamide F synthetase. Chembiochem 2006, 7, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Sunderhaus, J.D.; Sherman, D.H.; Williams, R.M. Studies on the biosynthesis of the stephacidin and notoamide natural products: A stereochemical and genetic conundrum. Isr. J. Chem. 2011, 51, 442–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendisch, V.F. Microbial Production of Amino Acid-Related Compounds. Adv. Biochem. Eng. Biotechnol. 2017, 159, 255–269. [Google Scholar] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Ma, Y.; Wei, J.; Ge, Y.; Jiang, W.; He, S.; Wu, X.; Zhang, X.; Wu, B. Comparative Metabolomics Reveals Fungal Conversion of Co-Existing Bacterial Metabolites within a Synthetic Aspergillus-Streptomyces Community. Mar. Drugs 2021, 19, 526. https://doi.org/10.3390/md19090526
Shi Y, Ma Y, Wei J, Ge Y, Jiang W, He S, Wu X, Zhang X, Wu B. Comparative Metabolomics Reveals Fungal Conversion of Co-Existing Bacterial Metabolites within a Synthetic Aspergillus-Streptomyces Community. Marine Drugs. 2021; 19(9):526. https://doi.org/10.3390/md19090526
Chicago/Turabian StyleShi, Yutong, Yihan Ma, Jihua Wei, Yichao Ge, Wei Jiang, Shan He, Xiaodan Wu, Xiaoqin Zhang, and Bin Wu. 2021. "Comparative Metabolomics Reveals Fungal Conversion of Co-Existing Bacterial Metabolites within a Synthetic Aspergillus-Streptomyces Community" Marine Drugs 19, no. 9: 526. https://doi.org/10.3390/md19090526
APA StyleShi, Y., Ma, Y., Wei, J., Ge, Y., Jiang, W., He, S., Wu, X., Zhang, X., & Wu, B. (2021). Comparative Metabolomics Reveals Fungal Conversion of Co-Existing Bacterial Metabolites within a Synthetic Aspergillus-Streptomyces Community. Marine Drugs, 19(9), 526. https://doi.org/10.3390/md19090526