
Antifouling Activity of Halogenated Compounds Derived from the Red Alga Sphaerococcus coronopifolius: Potential for the Development of Environmentally Friendly Solutions

 , ,
, ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Identification of Metabolites from Sphaerococcus coronopifolius
2.2. Evaluation of Antifouling Activity
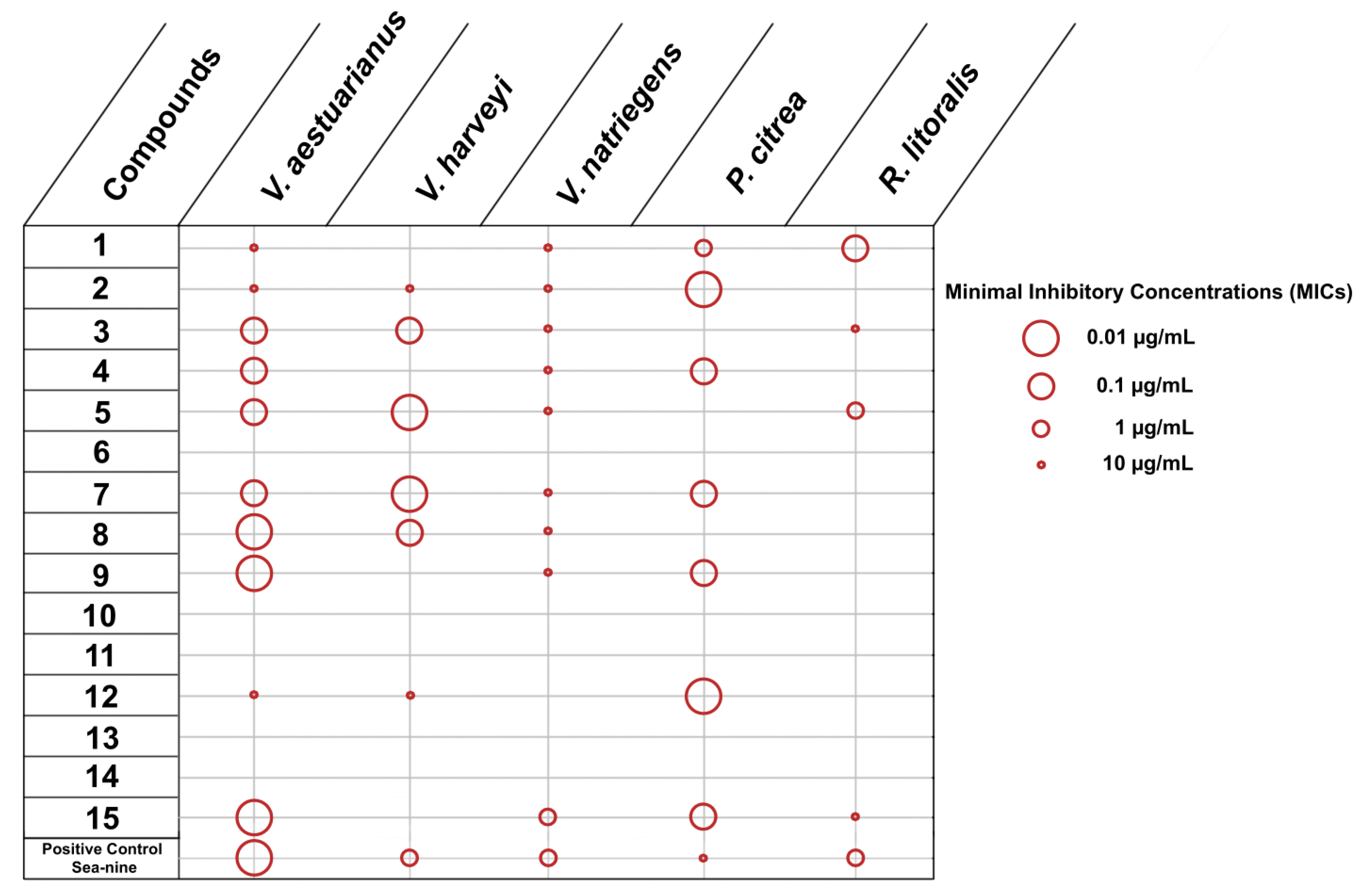
2.2.1. Inhibition of Adhesion and Growth of Marine Bacteria and Microalgae
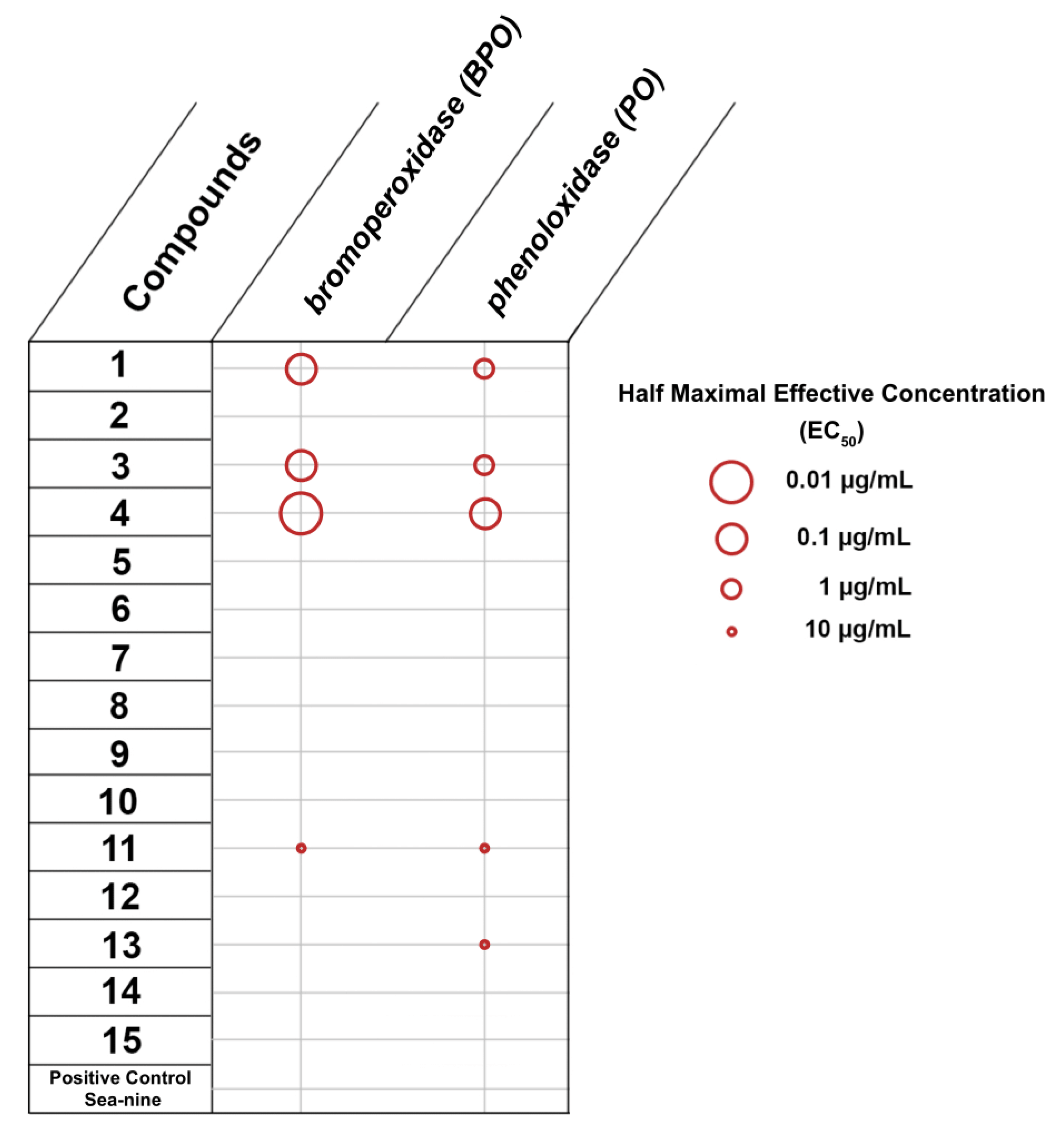
2.2.2. Inhibition of Bromoperoxidase and Phenoloxidase
2.2.3. Toxicity Evaluation of the Compounds
2.3. Overall Antifouling Potential
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Extraction and Isolation
3.4. Antifouling Evaluation Assays
3.5. Toxicity Assessment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Maréchal, J.-P.; Hellio, C. Challenges for the development of new non-toxic antifouling solutions. Int. J. Mol. Sci. 2009, 10, 4623–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovio, E.; Fauchon, M.; Toueix, Y.; Mehiri, M.; Varese, G.C.; Hellio, C. The Sponge-Associated Fungus Eurotium Chevalieri MUT 2316 and Its Bioactive Molecules: Potential Applications in the Field of Antifouling. Mar. Biotechnol. 2019, 21, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Saha, M.; Goecke, F.; Bhadury, P. Minireview: Algal Natural Compounds and Extracts as Antifoulants. J. Appl. Phycol. 2018, 30, 1859–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine Biofilms: Diversity of Communities and of Chemical Cues. Environ. Microbiol Rep. 2019, 11, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Fitridge, I.; Dempster, T.; Guenther, J.; de Nys, R. The Impact and Control of Biofouling in Marine Aquaculture: A Review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef]
- Bannister, J.; Sievers, M.; Bush, F.; Bloecher, N. Biofouling in Marine Aquaculture: A Review of Recent Research and Developments. Biofouling 2019, 35, 631–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Ning, C. Latest Research Progress of Marine Microbiological Corrosion and Bio-Fouling, and New Approaches of Marine Anti-Corrosion and Anti-Fouling. Bioact. Mater. 2019, 4, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic Impact of Biofouling on a Naval Surface Ship. Biofouling 2011, 27, 87–98. [Google Scholar] [CrossRef]
- Jadidi, P.; Zeinoddini, M. Influence of Hard Marine Fouling on Energy Harvesting from Vortex-Induced Vibrations of a Single-Cylinder. Renew. Energy 2020, 152, 516–528. [Google Scholar] [CrossRef]
- Amara, I.; Miled, W.; Slama, R.B.; Ladhari, N. Antifouling Processes and Toxicity Effects of Antifouling Paints on Marine Environment. A Review. Environ. Toxicol. Pharmacol. 2018, 57, 115–130. [Google Scholar] [CrossRef]
- Yebra, D.M.; Kiil, S.; Weinell, C.E.; Dam-Johansen, K. Presence and Effects of Marine Microbial Biofilms on Biocide-Based Antifouling Paints. Biofouling 2006, 22, 33–41. [Google Scholar] [CrossRef]
- Onduka, T.; Mizuno, K.; Shikata, T.; Mastubara, T.; Onitsuka, G.; Hamaguchi, M. Assessment of the Risk Posed by Three Antifouling Biocides to Pacific Oyster Embryos and Larvae in Hiroshima Bay, Japan. Environ. Sci Pollut Res. 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.R.; Ariffin, M.M.; Omar, T.F.T.; Ghazali, A.; Sheikh, M.A.; Shazili, N.A.M.; Bachok, Z. Antifouling Paint Biocides (Irgarol 1051 and Diuron) in the Selected Ports of Peninsular Malaysia: Occurrence, Seasonal Variation, and Ecological Risk Assessment. Environ. Sci. Pollut. Res. 2021, 28, 52247–52257. [Google Scholar] [CrossRef] [PubMed]
- Watermann, B.T.; Daehne, B.; Sievers, S.; Dannenberg, R.; Overbeke, J.C.; Klijnstra, J.W.; Heemken, O. Bioassays and Selected Chemical Analysis of Biocide-Free Antifouling Coatings. Chemosphere 2005, 60, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Kymenvaara, S.; Tegnér Anker, H.; Baaner, L.; Ekroos, A.; Gipperth, L.; Seppälä, J. Regulating Antifouling Paints for Leisure Boats—A Patchwork of Rules Across Three Baltic Sea Countries. Nord. Environ. Law J. 2017, 1, 7–32. Available online: http://nordiskmiljoratt.se/onewebmedia/Kymenvaara%20et%20al.pdf (accessed on 1 December 2021).
- Jones, B.; Bolam, T. Copper Speciation Survey from UK Marinas, Harbours and Estuaries. Mar. Pollut Bull. 2007, 54, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.-Y.; Li, Z.; Xu, Y.; Li, Y.; Fusetani, N. Mini-Review: Marine Natural Products and Their Synthetic Analogs as Antifouling Compounds: 2009-2014. Biofouling 2015, 31, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Alemán-Vega, M.; Sánchez-Lozano, I.; Hernández-Guerrero, C.J.; Hellio, C.; Quintana, E.T. Exploring Antifouling Activity of Biosurfactants Producing Marine Bacteria Isolated from Gulf of California. Int. J. Mol. Sci. 2020, 21, 6068. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lozano, I.; Hernández-Guerrero, C.J.; Muñoz-Ochoa, M.; Hellio, C. Biomimetic Approaches for the Development of New Antifouling Solutions: Study of Incorporation of Macroalgae and Sponge Extracts for the Development of New Environmentally-Friendly Coatings. Int. J. Mol. Sci. 2019, 20, 4863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Yu, L.; Mou, J.; Wu, D.; Xu, M.; Zhou, P.; Ren, Y. Research Strategies to Develop Environmentally Friendly Marine Antifouling Coatings. Mar. Drugs 2020, 18, 371. [Google Scholar] [CrossRef] [PubMed]
- Carson, R.T.; Damon, M.; Johnson, L.T.; Gonzalez, J.A. Conceptual Issues in Designing a Policy to Phase out Metal-Based Antifouling Paints on Recreational Boats in San Diego Bay. J. Environ. Manag. 2009, 90, 2460–2468. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Lewis, J.A.; Johnston, E.L. Antifouling Strategies: History and Regulation, Ecological Impacts and Mitigation. Mar. Pollut. Bull. 2011, 62, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef] [PubMed]
- Levert, A.; Foulon, V.; Fauchon, M.; Tapissier-Bontemps, N.; Banaigs, B.; Hellio, C. Antifouling Activity of Meroterpenes Isolated from the Ascidian Aplidium Aff. Densum. Mar. Biotechnol. 2021, 23, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Shinde, P.; Banerjee, P.; Mandhare, A. Marine Natural Products as Source of New Drugs: A Patent Review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef] [PubMed]
- Matulja, D.; Wittine, K.; Malatesti, N.; Laclef, S.; Turks, M.; Markovic, M.K.; Ambrožić, G.; Marković, D. Marine Natural Products with High Anticancer Activities. Curr. Med. Chem. 2020, 27, 1243–1307. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, I.P.S.; Nah, J.-W.; Jeon, Y.-J. Potential Anti-Inflammatory Natural Products from Marine Algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Martínez Andrade, K.A.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, E. Mining Natural Product Biosynthesis in Eukaryotic Algae. Mar. Drugs 2020, 18, 90. [Google Scholar] [CrossRef] [Green Version]
- Piazza, V.; Roussis, V.; Garaventa, F.; Greco, G.; Smyrniotopoulos, V.; Vagias, C.; Faimali, M. Terpenes from the red alga Sphaerococcus coronopifolius inhibit the settlement of barnacles. Mar. Biotechnol. 2011, 13, 764–772. [Google Scholar] [CrossRef]
- Fattorusso, E.; Magno, S.; Santacroce, C.; Sica, D.; Di Blasio, B.; Pedone, C.; Impellizzeri, G.; Mangiafico, S.; Oriente, G.; Piattelli, M.; et al. Bromosphaerol, a new bromine containing diterpenoid from the red alga Sphaerococcus coronopifolius. Gazz. Chim. Ital. 1976, 106, 779–783. [Google Scholar]
- Cafieri, F.; Ciminiello, P.; Fattorusso, E.; Mangoni, A. Two novel bromoditerpenes from the red alga Sphaerococcus coronopifolius. Gazz. Chim. Ital. 1990, 120, 139–142. [Google Scholar]
- Cafieri, F.; Ciminiello, P.; Fattorusso, E.; Santacroce, C. 12S-Hydroxybromosphaerol, a new bromoditerpene from the red alga Sphaerococcus coronopifolius. Experientia 1982, 38, 298–299. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Quesada, A.; Vagias, C.; Moreau, D.; Roussakis, C.; Roussis, V. Cytotoxic bromoditerpenes from the red alga Sphaerococcus coronopifolius. Tetrahedron 2008, 64, 5184–5190. [Google Scholar] [CrossRef]
- Cafieri, F.; De Napoli, L.; Fattorusso, E.; Santacroce, C. Diterpenes from the red alga Sphaerococcus coronopifolius. Phytochemistry 1987, 26, 471–473. [Google Scholar] [CrossRef]
- Fenical, W.; Finer, J.; Clardy, J. Sphaerococcenol A, a new rearranged bromo-diterpene from the red alga Sphaerococcus coronopifolius. Tetrahedron Lett. 1976, 17, 731–734. [Google Scholar] [CrossRef]
- De Rosa, S.; De Stefano, S.; Scarpelli, P.; Zavodnik, N. Terpenes from the red alga Sphaerococcus coronopifolius of the North Adriatic Sea. Phytochemistry 1988, 27, 1875–1878. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; de Andrade Tomaz, A.C.; Vanderlei de Souza, M.F.; da Cunha, E.V.L.; Kiss, R.; Mathieu, V.; Ioannou, E.; Roussis, V. Halogenated diterpenes with in vitro antitumor activity from the red alga Sphaerococcus coronopifolius. Mar. Drugs 2020, 18, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyrniotopoulos, V.; Vagias, C.; Rahman, M.M.; Gibbons, S.; Roussis, V. Brominated diterpenes with antibacterial activity from the red alga Sphaerococcus coronopifolius. J. Nat. Prod. 2008, 71, 1386–1392. [Google Scholar] [CrossRef]
- Cafieri, F.; De Napoli, L.; Fattorusso, E.; Impellizzeri, G.; Piattelli, M.; Sciuto, S. Bromosphaerodiol, a minor bromo compound from the red alga Sphaerococcus coronopifolius. Experientia 1977, 33, 1549–1550. [Google Scholar] [CrossRef]
- Cafieri, F.; Ciminiello, P.; Santacroce, C.; Fattorusso, E. (1S)-1,2-Dihydro-1-hydroxybromosphaerol, a minor bromoditerpene from the red alga Sphaerococcus coronopifolius. Phytochemistry 1982, 21, 2412–2413. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Vagias, C.; Bruyere, C.; Lamoral-Theys, D.; Kiss, R.; Roussis, V. Structure and in vitro antitumor activity evaluation of brominated diterpenes from the red alga Sphaerococcus coronopifolius. Bioorg. Med. Chem. 2010, 18, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Faure, R.; Ramanoelina, A.R.P.; Rokotonirainy, O.; Bianchini, J.P.; Gaydou, E. Two-dimensional nuclear magnetic resonance of sesquiterpenes. Application to complete assignment of 1H and 13C NMR spectra of some aromadendrane derivatives. Mag. Reson. Chem. 1991, 29, 969–971. [Google Scholar] [CrossRef]
- Almeida, J.R.; Correia-da-Silva, M.; Sousa, E.; Antunes, J.; Pinto, M.; Vasconcelos, V.; Cunha, I. Antifouling Potential of Nature-Inspired Sulfated Compounds. Sci. Rep. 2017, 7, 42424. [Google Scholar] [CrossRef] [Green Version]
- Ralston, E.; Swain, G. Bioinspiration--the Solution for Biofouling Control? Bioinspir. Biomim. 2009, 4, 015007. [Google Scholar] [CrossRef] [PubMed]
- Marechal, J.-P.; Hellio, C. Antifouling Activity against Barnacle Cypris Larvae: Do Target Species Matter (Amphibalanus Amphitrite versus Semibalanus Balanoides)? Int. Biodeterior. Biodegrad. 2010, 65, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Niemann, H.; Hagenow, J.; Chung, M.-Y.; Hellio, C.; Weber, H.; Proksch, P. SAR of Sponge-Inspired Hemibastadin Congeners Inhibiting Blue Mussel PhenolOxidase. Mar. Drugs 2015, 13, 3061–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebret, K.; Thabard, M.; Hellio, C. Algae as Marine Fouling Organisms: Adhesion Damage and Prevention. Adv. Mar. Antifouling Coat. Tech. 2009, 1, 80–112. [Google Scholar] [CrossRef]
- Dobretsov, S.; Coutinho, R.; Rittschof, D.; Salta, M.; Ragazzola, F.; Hellio, C. The Oceans Are Changing: Impact of Ocean Warming and Acidification on Biofouling Communities. Biofouling 2019, 35, 585–595. [Google Scholar] [CrossRef]
- Bitton, R.; Ben-Yehuda, M.; Davidovich, M.; Balazs, Y.; Potin, P.; Delage, L.; Colin, C.; Bianco-Peled, H. Structure of Algal-Born Phenolic Polymeric Adhesives. Macromol. Biosci. 2006, 6, 737–746. [Google Scholar] [CrossRef]
- Chiovitti, A.; Heraud, P.; Dugdale, T.M.; Hodson, O.M.; Curtain, R.C.A.; Dagastine, R.R.; Wood, B.R.; Wetherbee, R. Divalent Cations Stabilize the Aggregation of Sulfated Glycoproteins in the Adhesive Nanofibers of the Biofouling Diatom Toxarium Undulatum. Soft. Matter. 2008, 4, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Dimartino, S.; Savory, D.M.; Fraser-Miller, S.J.; Gordon, K.C.; McQuillan, A.J. Microscopic and Infrared Spectroscopic Comparison of the Underwater Adhesives Produced by Germlings of the Brown Seaweed Species Durvillaea Antarctica and Hormosira Banksii. J. Royal Soc. Interface 2016, 13, 20151083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vreeland, V.; Waite, J.; Epstein, L. Minireview—Polyphenols and Oxidases in Substratum Adhesion by Marine Algae and Mussels. J. Phycol. 1998, 34, 1–8. [Google Scholar] [CrossRef]
- La Barre, S.; Potin, P.; Leblanc, C.; Delage, L. The Halogenated Metabolism of Brown Algae (Phaeophyta), Its Biological Importance and Its Environmental Significance. Mar. Drugs 2010, 8, 988–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellio, C.; Bourgougnon, N.; Gal, Y.L. Phenoloxidase (E.C. 1.14.18.1) from the Byssus Gland of Mytilus Edulis: Purification, Partial Characterization and Application for Screening Products with Potential Antifouling Activities. Biofouling 2000, 16, 235–244. [Google Scholar] [CrossRef]
- Alves, C.; Serrano, E.; Silva, J.; Rodrigues, C.; Pinteus, S.; Gaspar, H.; Botana, L.M.; Alpoim, M.C.; Pedrosa, R. Sphaerococcus Coronopifolius Bromoterpenes as Potential Cancer Stem Cell-Targeting Agents. Biomed. Pharm. 2020, 128, 110275. [Google Scholar] [CrossRef]
- Moodie, L.W.K.; Cervin, G.; Trepos, R.; Labriere, C.; Hellio, C.; Pavia, H.; Svenson, J. Design and Biological Evaluation of Antifouling Dihydrostilbene Oxime Hybrids. Mar. Biotechnol. 2018, 20, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Hellio, C.; Trepos, R.; Aguila-Ramírez, R.N.; Hernández-Guerrero, C.J. Protocol for Assessing Antifouling Activities of Macroalgal Extracts. Methods Mol. Biol. 2015, 1308, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Chambers, L.D.; Hellio, C.; Stokes, K.R.; Dennington, S.P.; Goodes, L.R.; Wood, R.J.K.; Walsh, F.C. Investigation of Chondrus Crispus as a Potential Source of New Antifouling Agents. Int. Biodeterior. Biodegrad. 2011, 7, 939–946. [Google Scholar] [CrossRef]
- Hellio, C.; De La Broise, D.; Dufossé, L.; Le Gal, Y.; Bourgougnon, N. Inhibition of Marine Bacteria by Extracts of Macroalgae: Potential Use for Environmentally Friendly Antifouling Paints. Mar. Environ. Res. 2001, 52, 231–247. [Google Scholar] [CrossRef]
- Hellio, C.; Bremer, G.; Pons, A.M.; Le Gal, Y.; Bourgougnon, N. Inhibition of the Development of Microorganisms (Bacteria and Fungi) by Extracts of Marine Algae from Brittany, France. Appl. Microbiol. Biotechnol. 2000, 54, 543–549. [Google Scholar] [CrossRef]
- Chen, J.; Li, H.; Zhao, Z.; Xia, X.; Li, B.; Zhang, J.; Yan, X. Diterpenes from the Marine Algae of the Genus Dictyota. Mar. Drugs 2018, 16, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-Y.; Hao, H.-L.; Lau, S.C.K.; Wang, H.-Y.; Han, Y.; Dong, L.-M.; Huang, R.-M. Biodiversity and Antifouling Activity of Fungi Associated with Two Soft Corals from the South China Sea. Arch. Microbiol. 2019, 201, 757–767. [Google Scholar] [CrossRef]
- Tobal, I.E.; Roncero, A.M.; Moro, R.F.; Díez, D.; Marcos, I.S. Antibacterial Natural Halimanes: Potential Source of Novel Antibiofilm Agents. Molecules 2020, 25, 1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring Structural Diversity of Microbe Secondary Metabolites Using OSMAC Strategy: A Literature Review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Navarri, M.; Jégou, C.; Meslet-Cladière, L.; Brillet, B.; Barbier, G.; Burgaud, G.; Fleury, Y. Deep Subseafloor Fungi as an Untapped Reservoir of Amphipathic Antimicrobial Compounds. Mar. Drugs 2016, 14, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quémener, M.; Kikionis, S.; Fauchon, M.; Toueix, Y.; Aulanier, F.; Makris, A.M.; Roussis, V.; Ioannou, E.; Hellio, C. Antifouling Activity of Halogenated Compounds Derived from the Red Alga Sphaerococcus coronopifolius: Potential for the Development of Environmentally Friendly Solutions. Mar. Drugs 2022, 20, 32. https://doi.org/10.3390/md20010032
Quémener M, Kikionis S, Fauchon M, Toueix Y, Aulanier F, Makris AM, Roussis V, Ioannou E, Hellio C. Antifouling Activity of Halogenated Compounds Derived from the Red Alga Sphaerococcus coronopifolius: Potential for the Development of Environmentally Friendly Solutions. Marine Drugs. 2022; 20(1):32. https://doi.org/10.3390/md20010032
Chicago/Turabian StyleQuémener, Maxence, Stefanos Kikionis, Marilyne Fauchon, Yannick Toueix, Fanny Aulanier, Antonios M. Makris, Vassilios Roussis, Efstathia Ioannou, and Claire Hellio. 2022. "Antifouling Activity of Halogenated Compounds Derived from the Red Alga Sphaerococcus coronopifolius: Potential for the Development of Environmentally Friendly Solutions" Marine Drugs 20, no. 1: 32. https://doi.org/10.3390/md20010032
APA StyleQuémener, M., Kikionis, S., Fauchon, M., Toueix, Y., Aulanier, F., Makris, A. M., Roussis, V., Ioannou, E., & Hellio, C. (2022). Antifouling Activity of Halogenated Compounds Derived from the Red Alga Sphaerococcus coronopifolius: Potential for the Development of Environmentally Friendly Solutions. Marine Drugs, 20(1), 32. https://doi.org/10.3390/md20010032