
Network Pharmacological Analysis of the Red Sea Sponge Hyrtios erectus Extract to Reveal Anticancer Efficacy of Corresponding Loaded Niosomes

 ,
,  , , ,
, , ,  and
and 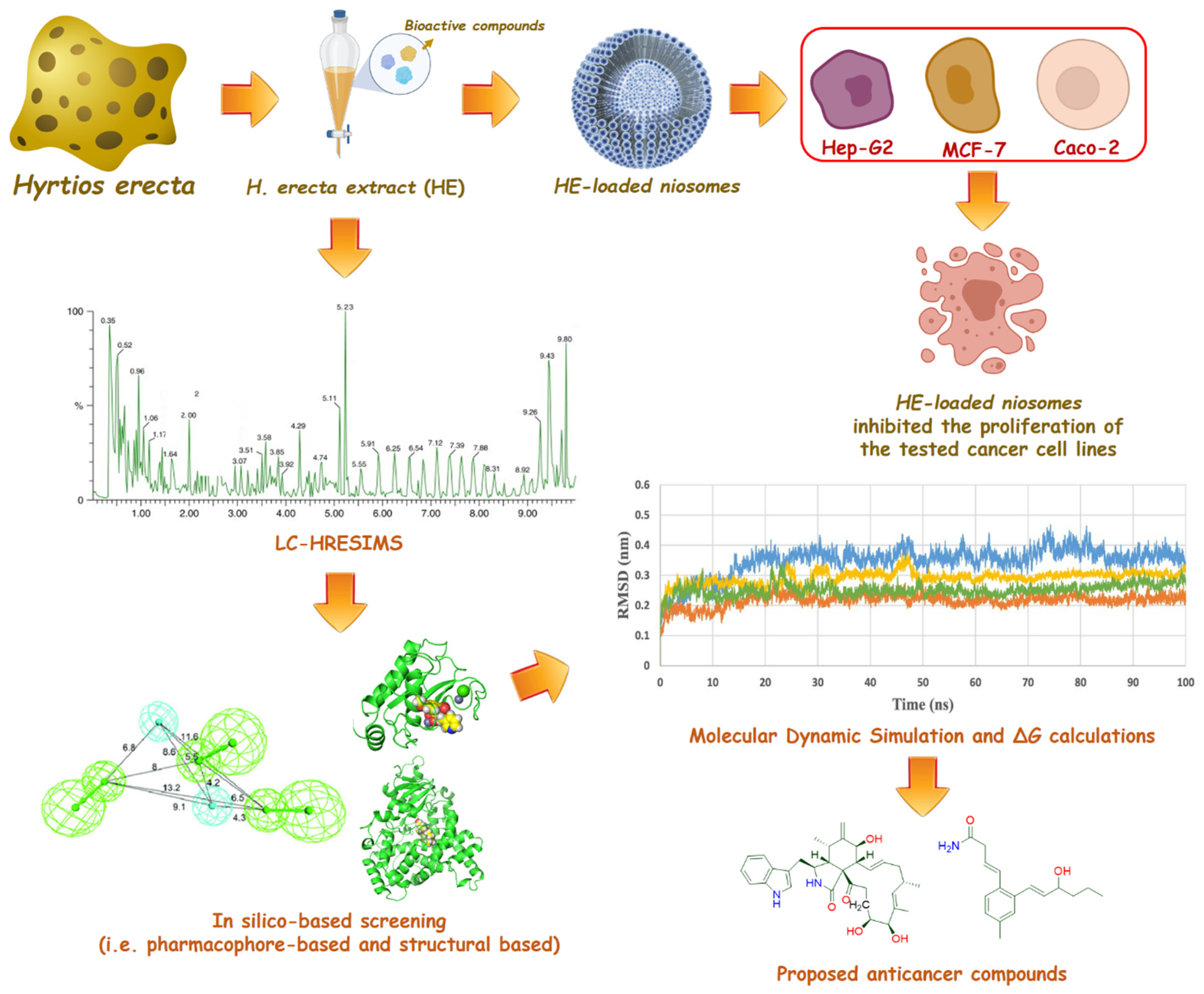{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. LC-HRMS Chemical Profiling
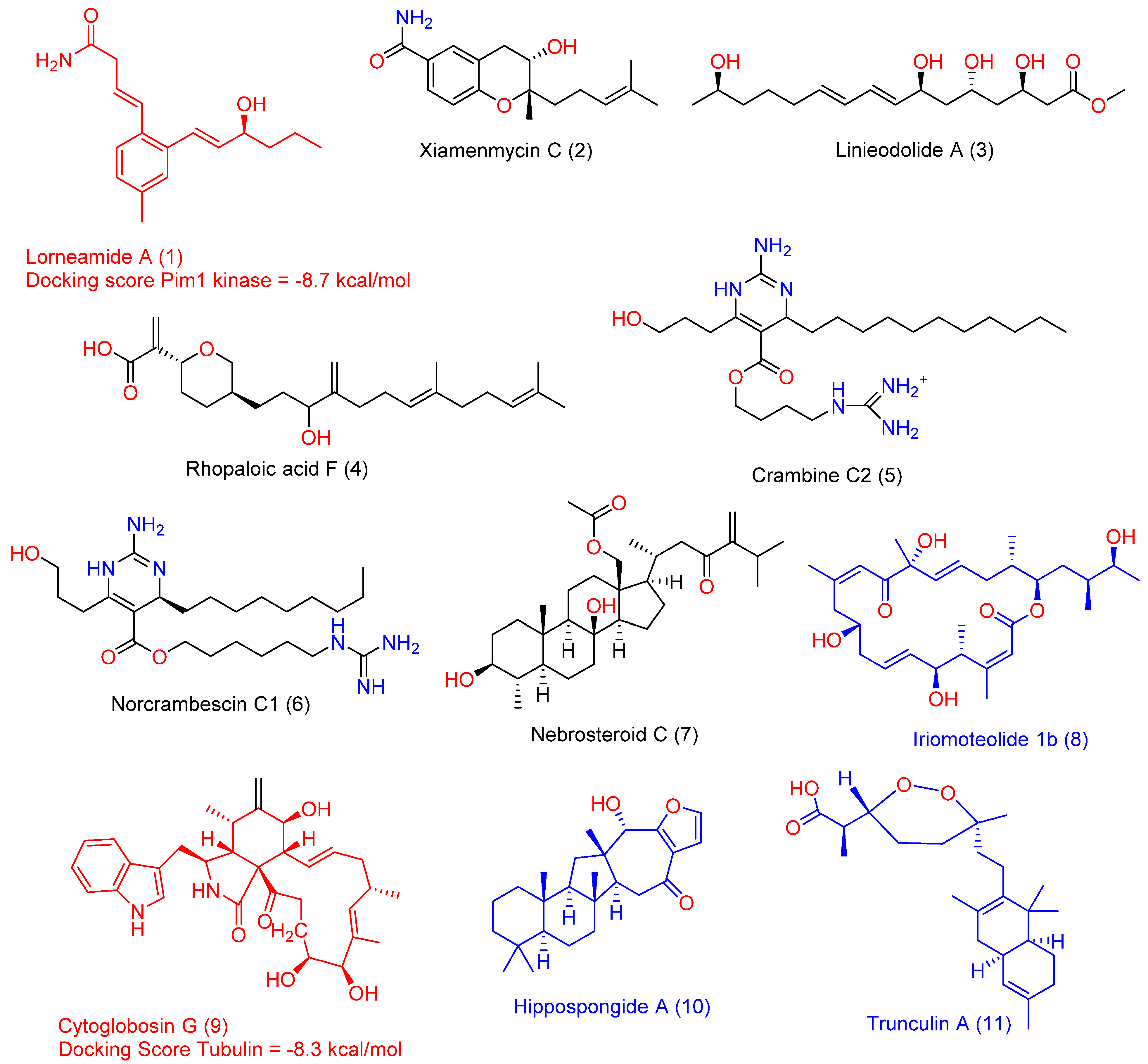
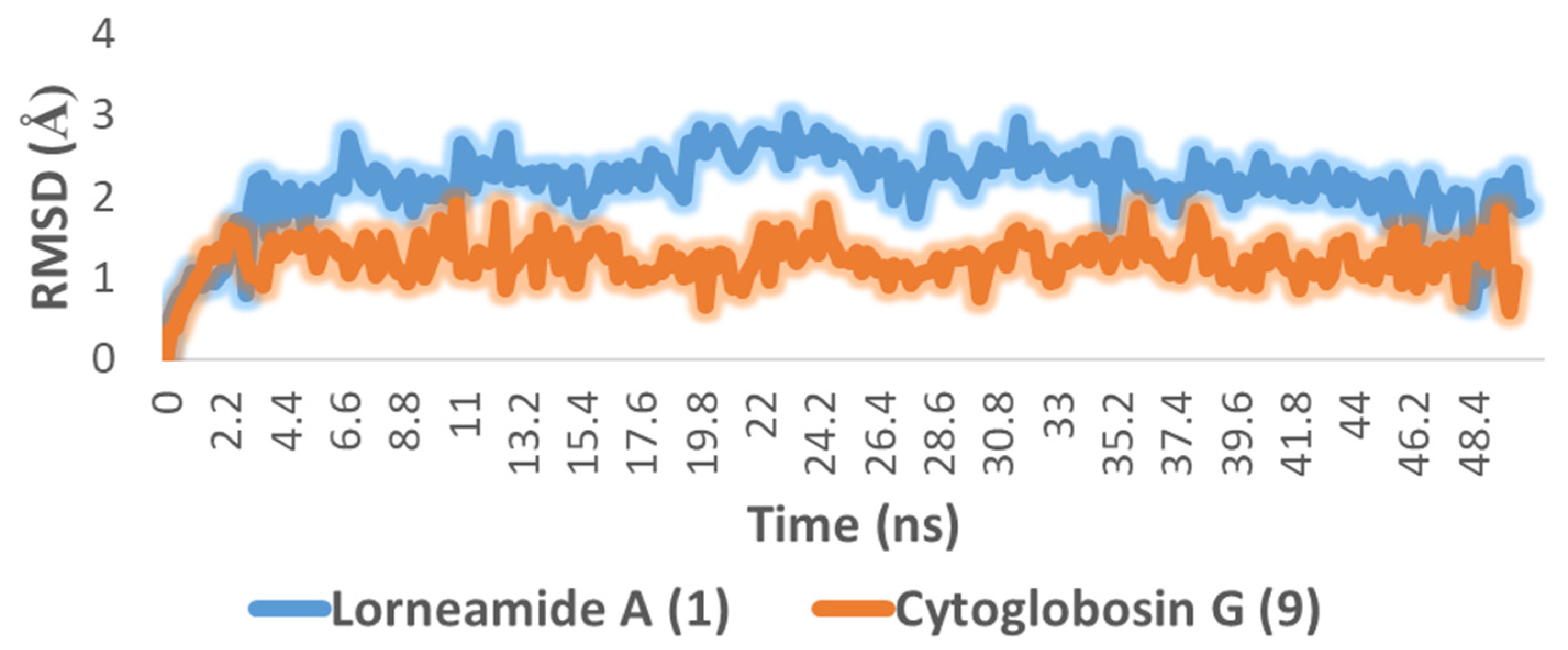
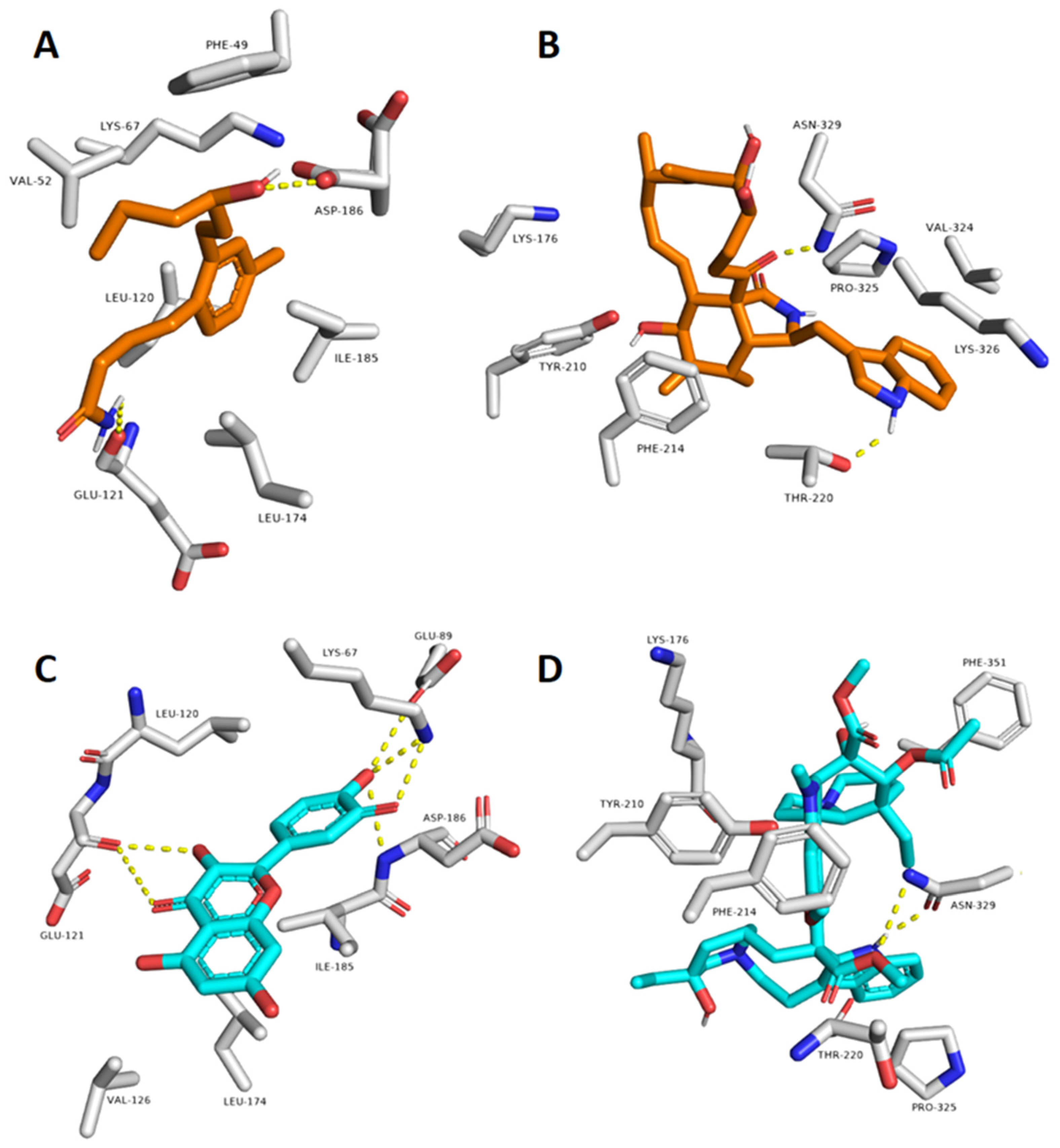
2.2. In Silico-Based Determination of the Active Metabolites
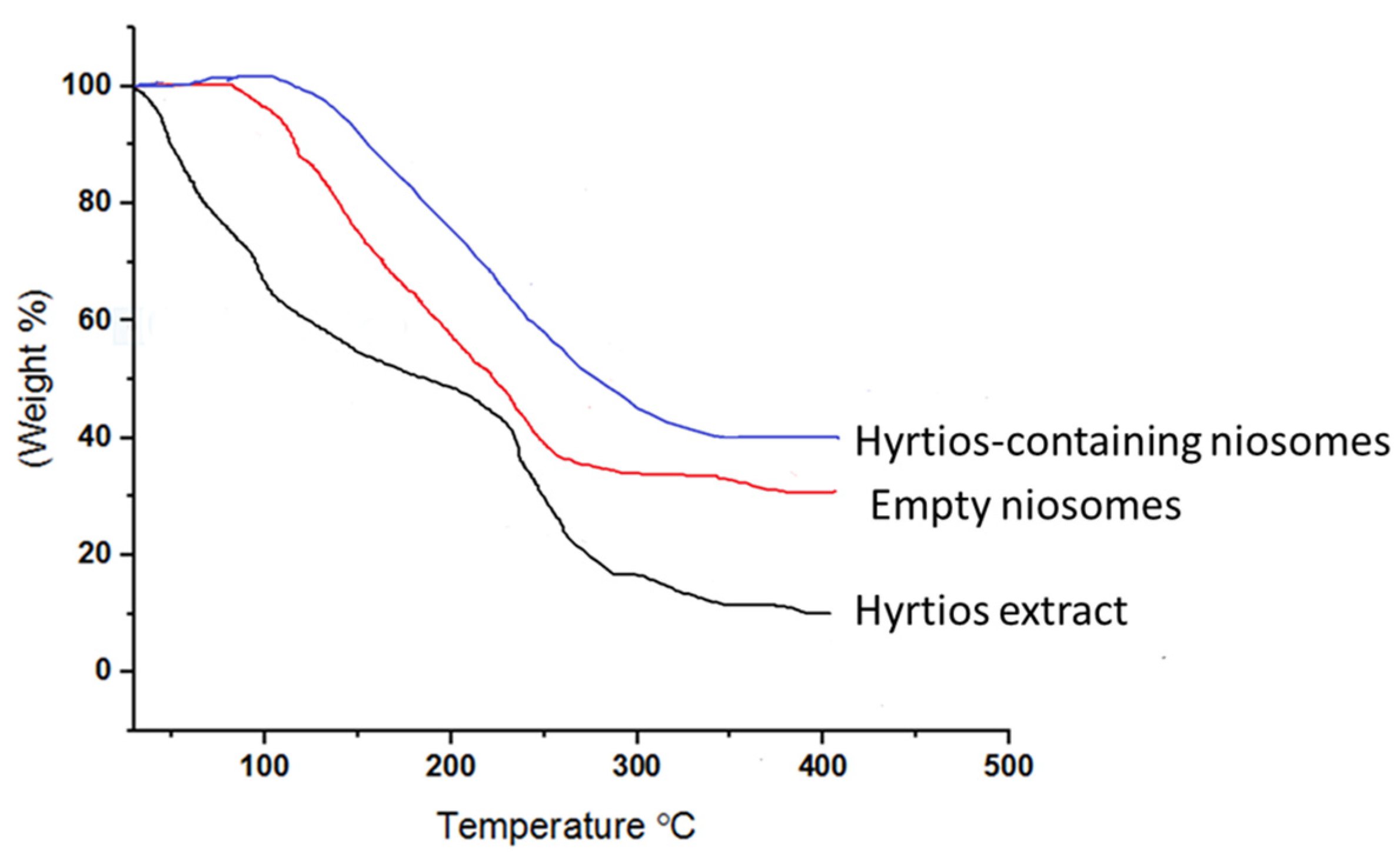
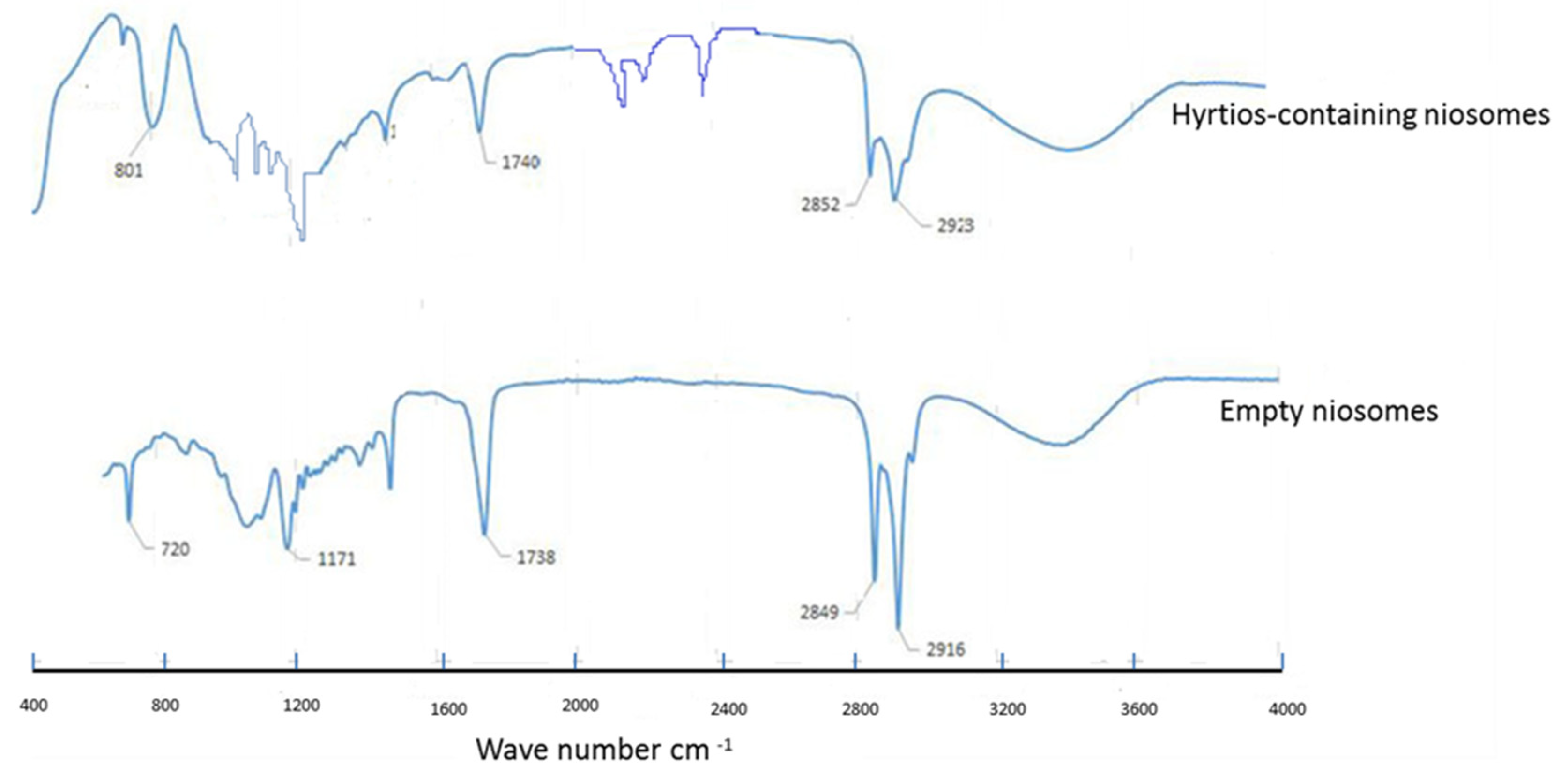
2.3. HE-Containing Niosomes
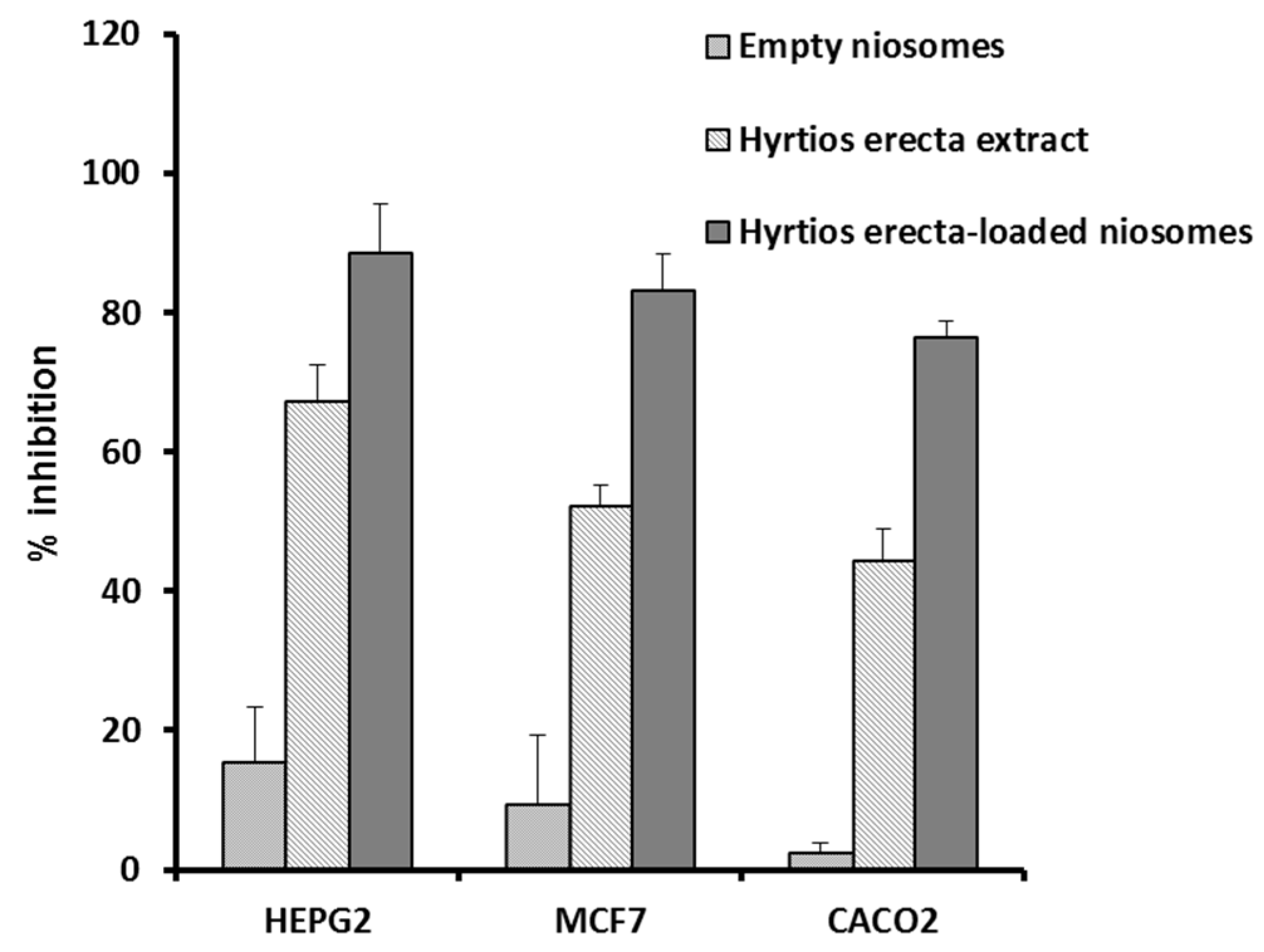
2.4. Antiproliferative Potential of the HE and Its Niosomal Formulation
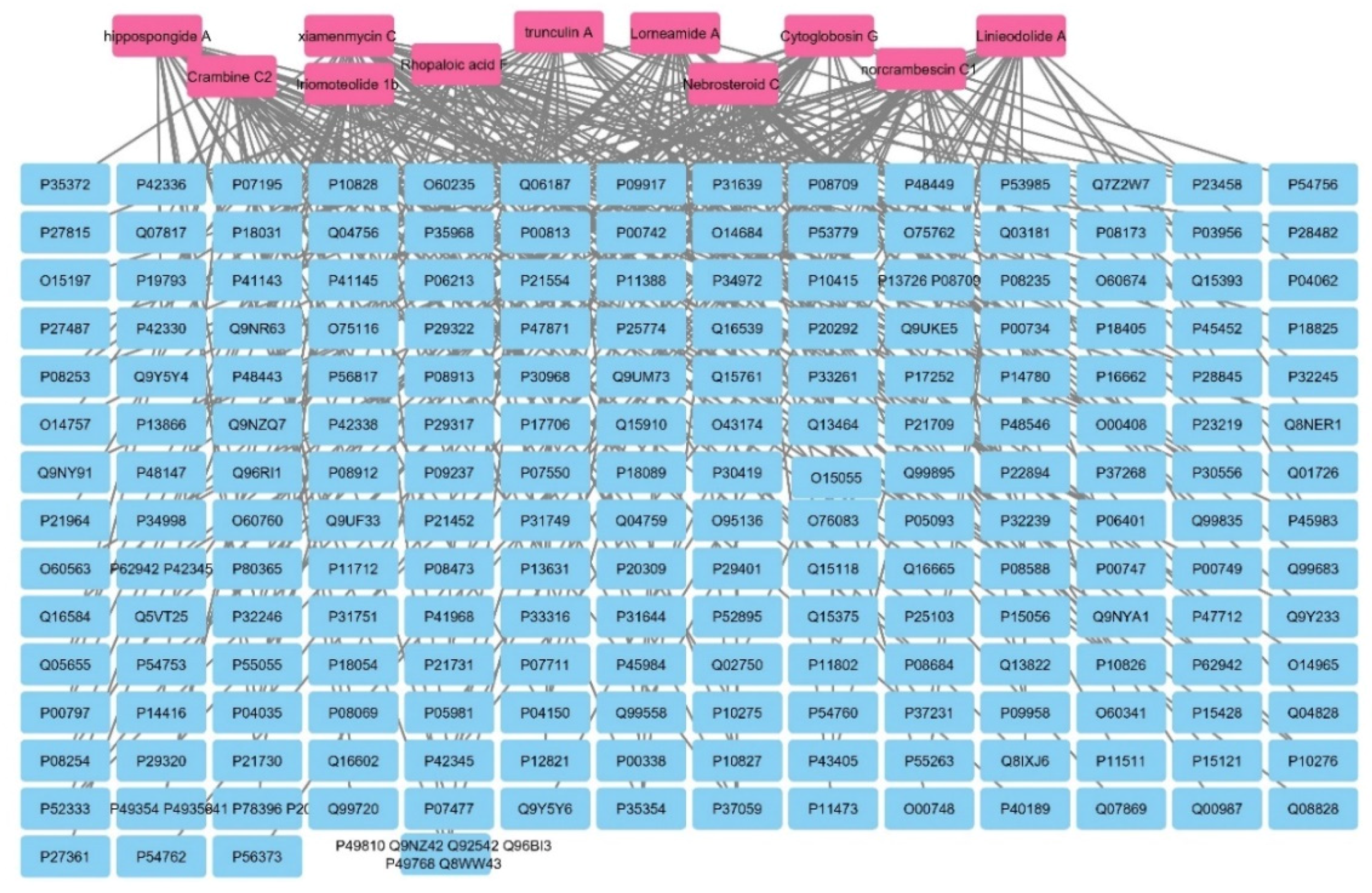
2.5. Network Pharmacology
2.5.1. Networks
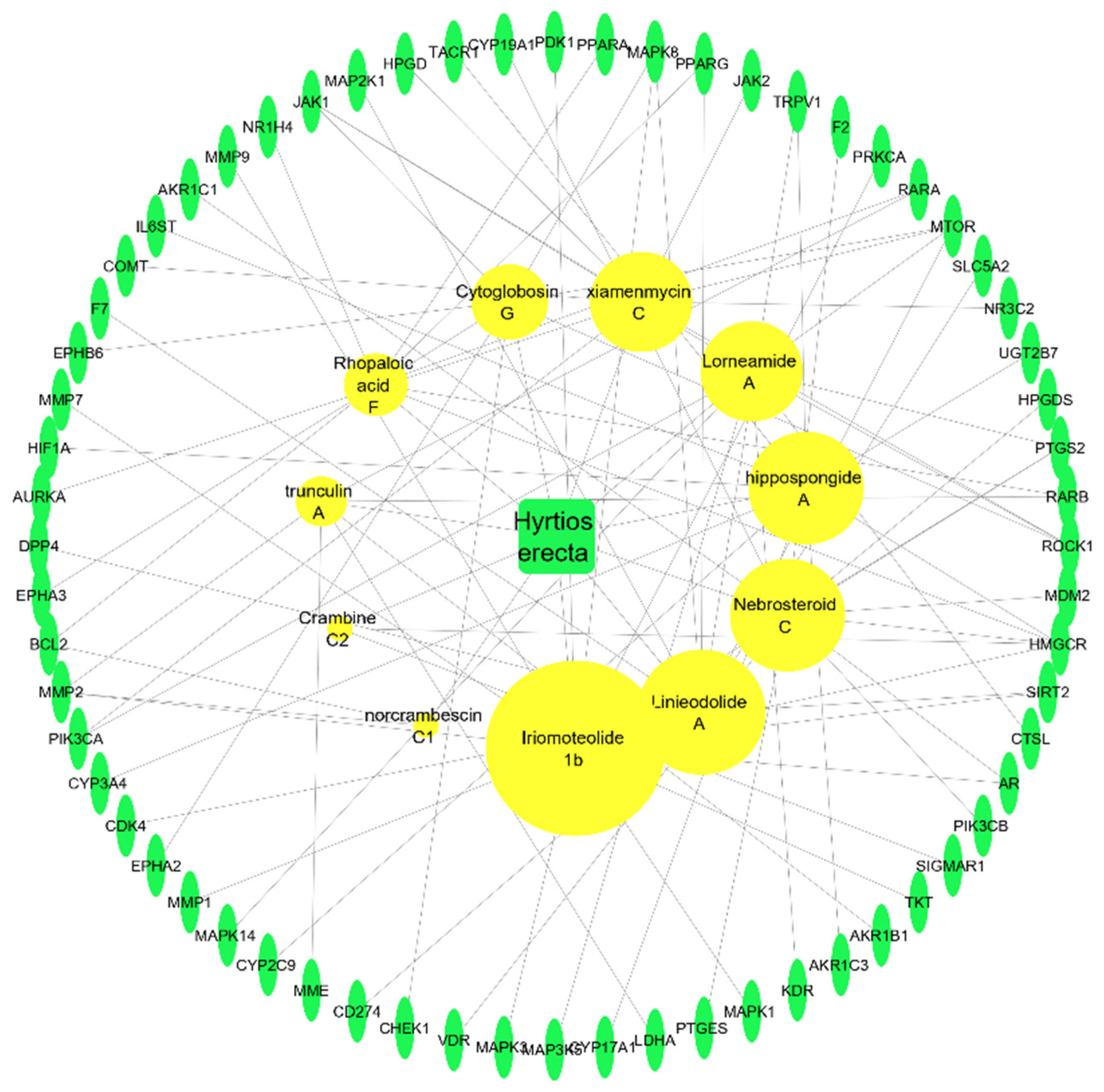
Sponge–Metabolite Network
The Metabolites–Target Genes Network
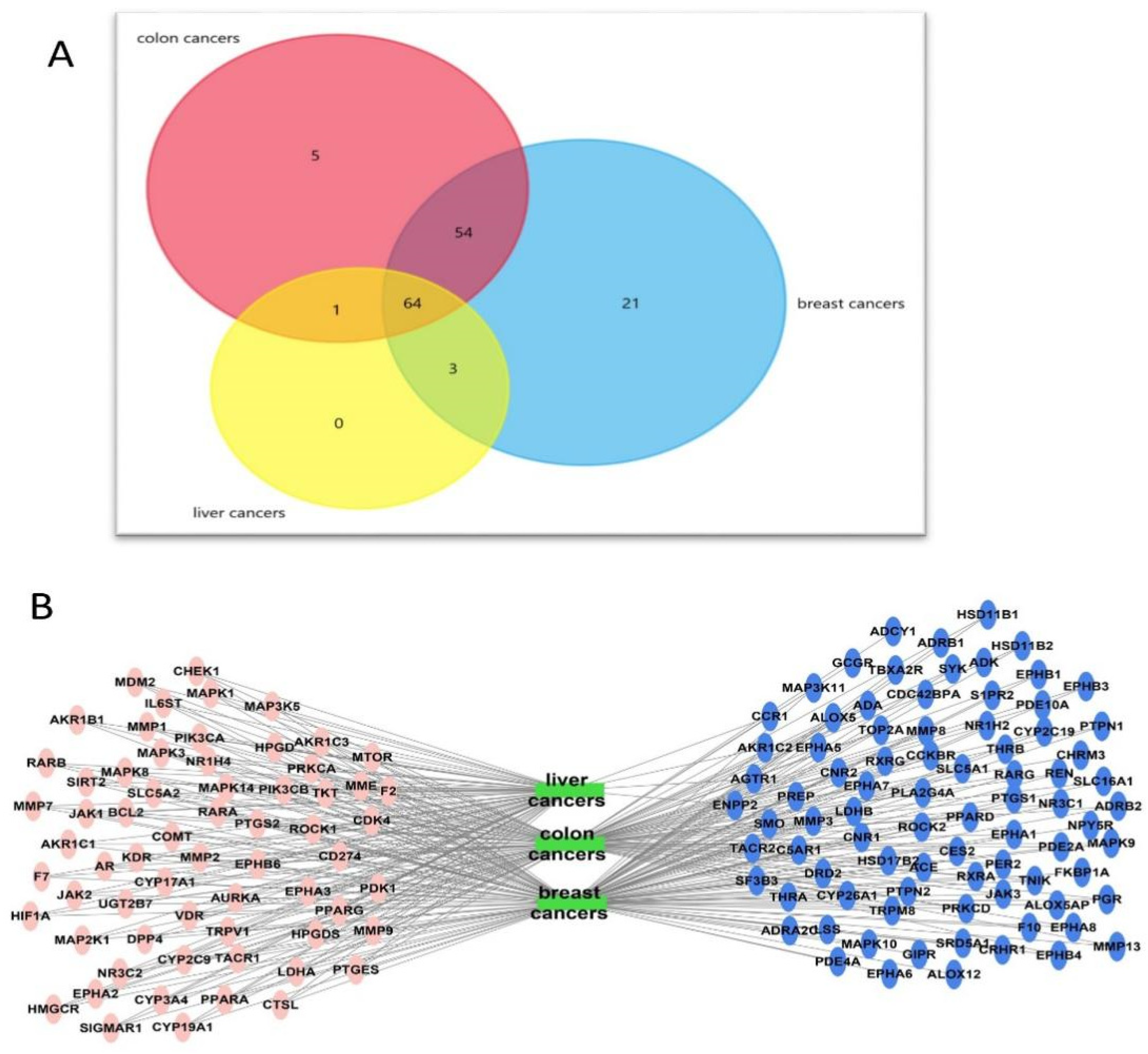
Targets–Cancer Types (Breast, Colon, and Hepatocellular) Network
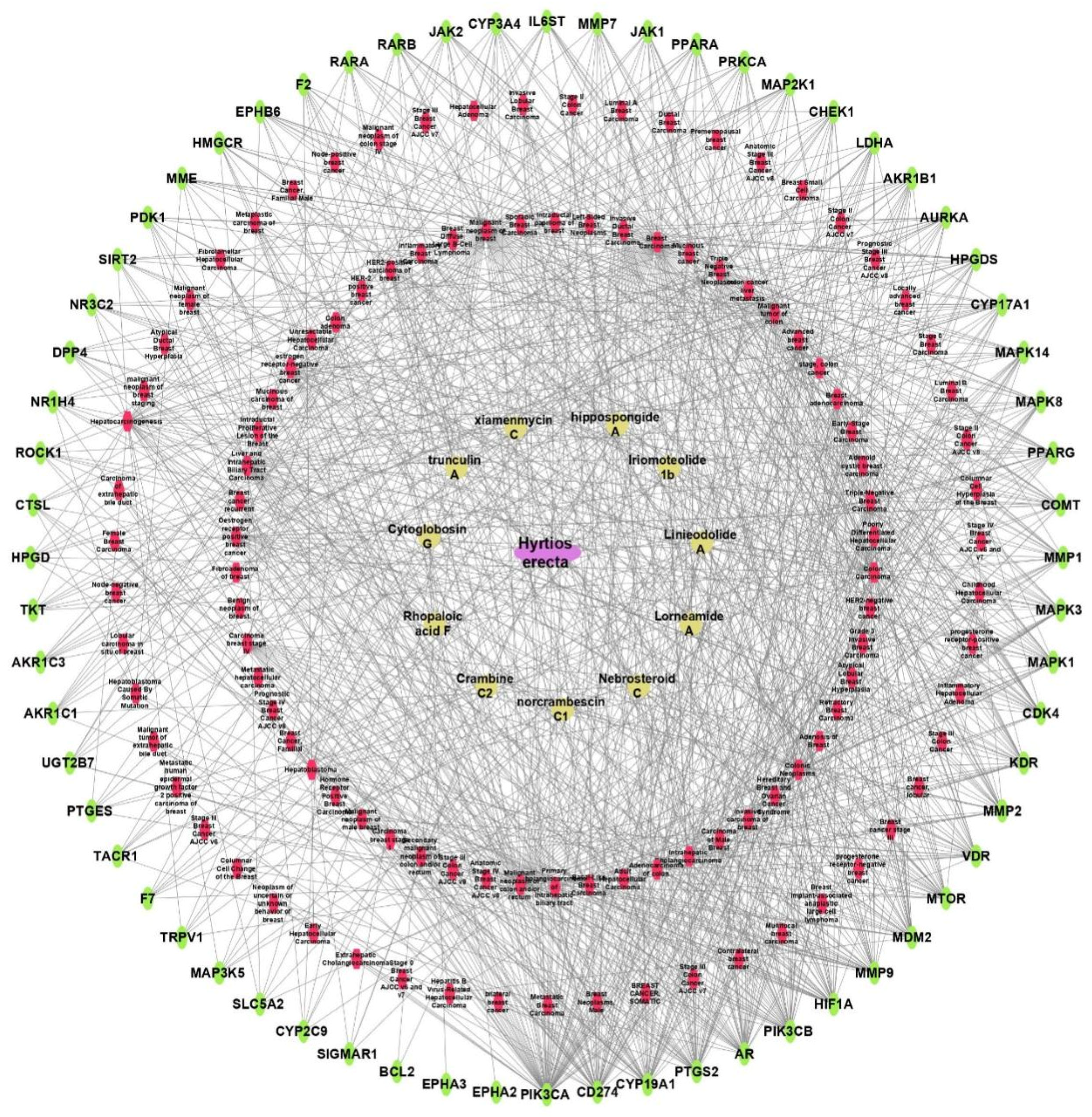
Sponge–Metabolite–Intersected Cancer Genes
Sponge–Metabolites–Intersected Genes–Classes of Types of Cancer
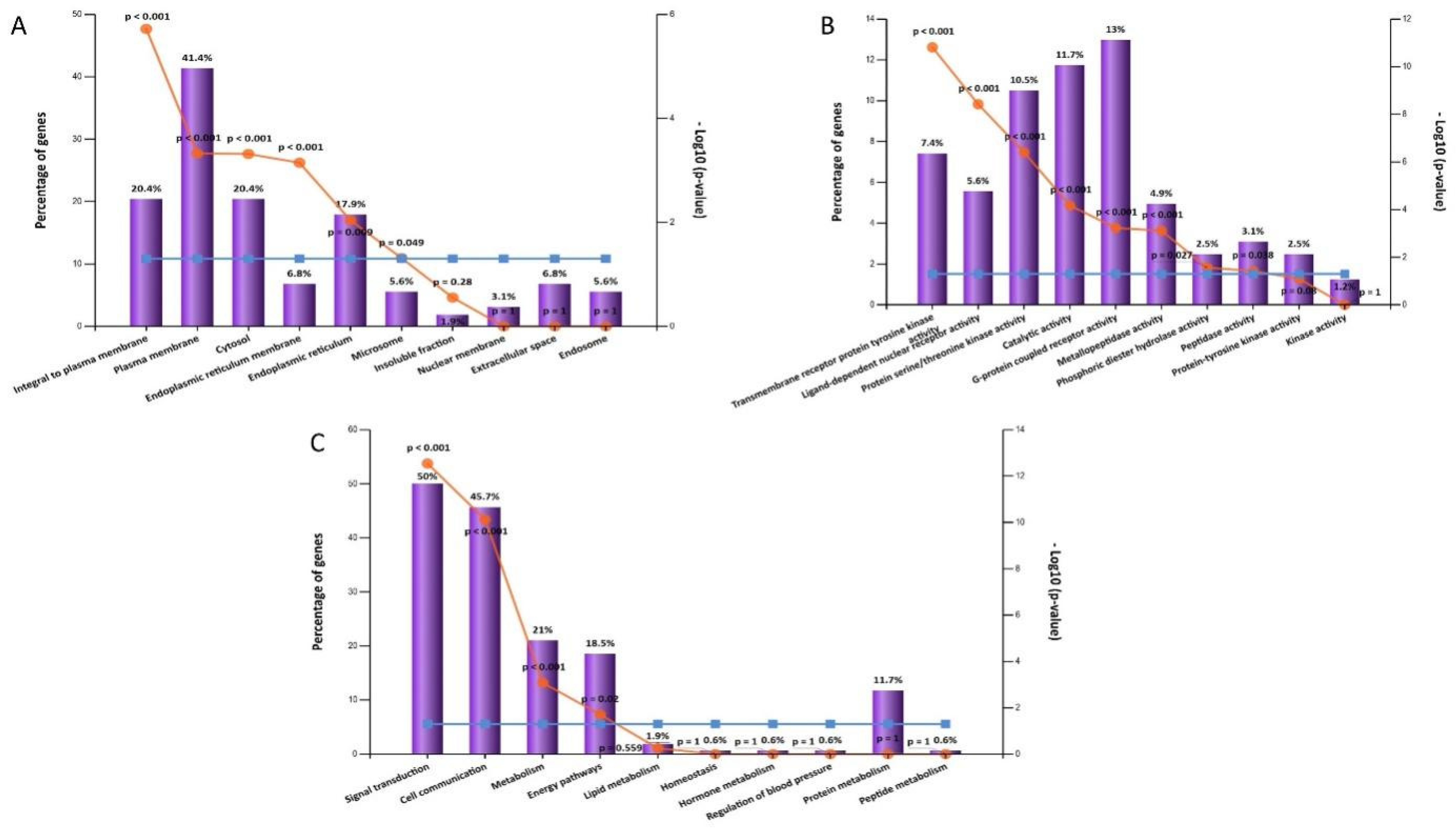
2.5.2. Gene Ontology and Enrichment Analysis
3. Material and Methods
3.1. Extraction of Sponge Material
3.2. LC-HRMS Analysis
3.3. Preparation of Niosomes of Hyrtios Erectus Extract
3.3.1. Transmission Electron Microscopy (TEM)
3.3.2. Size of HE-Containing Niosomes
3.3.3. FTIR and TGA of HE-Containing Niosomes
3.4. In Vitro MTT Assay
3.4.1. Conditions of Cell Culture
3.4.2. Antiproliferative Potential of HE and HE-Containing Niosomes
3.5. Molecular Modeling
3.5.1. In Silico Biological Activity Predictions
3.5.2. Prediction of the Potential Protein Targets
3.6. Network Pharmacology Study
3.6.1. Networks Construction
Sponge–Metabolite Network
The Metabolites–Target Genes Network
Targets–Cancer Types (Breast, Colon, and Hepatocellular)
Sponge–Metabolite–Intersected Cancer Genes
Sponge–Metabolites–Intersected Genes–Classes of Types of Cancer
3.6.2. Gene ontology and Enrichment Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hulvat, M.C. Cancer incidence and trends. Surg. Clin. 2020, 100, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Shady, N.H.; Abdelmohsen, U.R.; AboulMagd, A.M.; Amin, M.N.; Ahmed, S.; Fouad, M.A.; Kamel, M.S. Cytotoxic potential of the Red Sea sponge Amphimedon sp. supported by in silico modelling and dereplication analysis. Nat. Prod. Res. 2020, 35, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shady, N.H.; El-Hossary, E.M.; Fouad, M.A.; Gulder, T.A.; Kamel, M.S.; Abdelmohsen, U.R. Bioactive natural products of marine sponges from the genus Hyrtios. Molecules 2017, 22, 781. [Google Scholar] [CrossRef] [Green Version]
- Sen Gupta, B.K. Foraminifera in marginal marine environments. In Modern Foraminifera; Springer: Berlin/Heidelberg, Germany, 1999; pp. 141–159. [Google Scholar]
- Taufa, T.; Subramani, R.; Northcote, P.T.; Keyzers, R.A. Natural products from tongan marine organisms. Molecules 2021, 26, 4534. [Google Scholar] [CrossRef] [PubMed]
- Bourguet-Kondracki, M.-L.; Debitus, C.; Guyot, M. Dipuupehedione, a cytotoxic new red dimer from a new Caledonian marine sponge Hyrtios sp. Tetrahedron Lett. 1996, 37, 3861–3864. [Google Scholar] [CrossRef]
- Bourguet-Kondracki, M.-L.; Lacombe, F.; Guyot, M. Methanol adduct of puupehenone, a biologically active derivative from the marine sponge Hyrtios species. J. Nat. Prod. 1999, 62, 1304–1305. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Tsuda, M.; Fromont, J.; Kobayashi, J.I. Hyrtinadine A, a bis-indole alkaloid from a marine sponge. J. Nat. Prod. 2007, 70, 423–424. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Tan, R.; Melody, N.; Cichacz, Z.A.; Herald, D.L.; Hoard, M.S.; Pettit, R.K.; Chapuis, J.-C. Antineoplastic agents 397: Isolation and structure of sesterstatins 4 and 5 from hyrtios erecta (The Republic of Maldives). Bioorganic Med. Chem. Lett. 1998, 8, 2093–2098. [Google Scholar] [CrossRef]
- Gonzalez, M.A. Scalarane Sesterterpenoids; Bentham Science Publishers: Sharjah, United Arab Emirates, 2010; Volume 6, pp. 178–206. [Google Scholar]
- Chakraborty, K.; Francis, P. Apoptotic effect of chromanone derivative, hyrtiosone A from marine demosponge Hyrtios erectus in hepatocellular carcinoma HepG2 cells. Bioorganic Chem. 2021, 114, 105119. [Google Scholar] [CrossRef]
- Alishahi, A. Application of nanotechnology in marine-based products: A review. J. Aquat. Food Prod. Technol. 2015, 24, 150211123002003. [Google Scholar] [CrossRef]
- Durak, S.; Esmaeili Rad, M.; Alp Yetisgin, A.; Eda Sutova, H.; Kutlu, O.; Cetinel, S.; Zarrabi, A. Niosomal drug delivery systems for ocular disease—Recent advances and future prospects. Nanomaterials 2020, 10, 1191. [Google Scholar] [CrossRef] [PubMed]
- Lohumi, A. A novel drug delivery system: Niosomes review. J. Drug Deliv. Ther. 2012, 2, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Capon, R.J.; Skene, C.; Lacey, E.; Gill, J.H.; Wicker, J.; Heiland, K.; Friedel, T. Lorneamides A and B: Two new aromatic amides from a Southern Australian marine actinomycete. J. Nat. Prod. 2000, 63, 1682–1683. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.-Y.; Liu, X.-Y.; Li, X.-Y.; Yang, H.-G.; Li, L.; Jiao, X.-Z.; Xie, P. Total synthesis of xiamenmycin C and all of its stereoisomers: Stereochemical revision. J. Asian Nat. Prod. Res. 2016, 18, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Mondol, M.A.M.; Tareq, F.S.; Kim, J.H.; Lee, M.A.; Lee, H.S.; Lee, J.S.; Lee, Y.J.; Shin, H.J. New antimicrobial compounds from a marine-derived Bacillus sp. J. Antibiot. 2013, 66, 89–95. [Google Scholar] [CrossRef]
- Craig, K.S.; Williams, D.E.; Hollander, I.; Frommer, E.; Mallon, R.; Collins, K.; Wojciechowicz, D.; Tahir, A. Novel sesterterpenoid and norsesterterpenoid RCE-protease inhibitors isolated from the marine sponge Hippospongia sp. Tetrahedron Lett. 2002, 43, 4801–4804. [Google Scholar] [CrossRef]
- Berlinck, R.G.S.; Braekman, J.C.; Daloze, D.; Bruno, I.; Riccio, R.; Rogeau, D.; Amade, P. Crambines C1 and C2: Two further ichthyotoxic guanidine alkaloids from the sponge Crambe crambe. J. Nat. Prod. 1992, 55, 528–532. [Google Scholar] [CrossRef]
- Bondu, S.; Genta-Jouve, G.; Leiros, M.; Vale, C.; Guigonis, J.M.; Botana, L.M.; Thomas, O.P. Additional bioactive guanidine alkaloids from the Mediterranean sponge Crambe crambe. RSC Adv. 2012, 2, 2828–2835. [Google Scholar] [CrossRef]
- Huang, Y.C.; Wen, Z.H.; Wang, S.K.; Hsu, C.H.; Duh, C.Y. New anti-inflammatory 4-methylated steroids from the Formosan soft coral Nephthea chabroli. Steroids 2008, 73, 1181–1186. [Google Scholar] [CrossRef]
- Tsuda, M.; Oguchi, K.; Iwamoto, R.; Okamoto, Y.; Fukushi, E.; Kawabata, J.; Ozawa, T.; Masuda, A. Iriomoteolides-1b and-1c, 20-membered macrolides from a marine dinoflagellate Amphidinium species. J. Nat. Prod. 2007, 70, 1661–1663. [Google Scholar] [CrossRef]
- Cui, C.M.; Li, X.M.; Li, C.S.; Proksch, P.; Wang, B.G. Cytoglobosins A-G, Cytochalasans from a Marine-Derived Endophytic Fungus, Chaetomium globosum QEN-14. J. Nat. Prod. 2010, 73, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Tseng, S.W.; Liu, L.L.; Chou, Y.L.; Ho, Y.S.; Lu, M.C.; Su, J.H. Cytotoxic Sesterterpenoids from a Sponge Hippospongia sp. Mar. Drugs 2012, 10, 987–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capon, R.J.; MacLeod, J.K.; Willis, A.C. Trunculins A and B; norsesterterpene cyclic peroxides from a marine sponge; Latrunculia brevis. J. Org. Chem. 1987, 52, 339–342. [Google Scholar] [CrossRef]
- Sayed, A.M.; Alhadrami, H.A.; El-Hawary, S.S.; Mohammed, R.; Hassan, H.M.; Rateb, M.E.; Abdelmohsen, U.R.; Bakeer, W. Discovery of two brominated oxindole alkaloids as Staphylococcal DNA gyrase and pyruvate kinase inhibitors via inverse virtual screening. Microorganisms 2020, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Lagunin, A.; Stepanchikova, A.; Filimonov, D.; Poroikov, V. Internet server for on-line prediction of the biological activity spectrum for a substance. Bioinformatics 2000, 16, 747–748. [Google Scholar] [CrossRef] [Green Version]
- Coué, M.; Brenner, S.L.; Spector, I.; Korn, E.D. Inhibition of actin polymerization by latrunculin A. FEBS Lett. 1987, 213, 316–318. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.K.; Yuan, H. Enantioselective syntheses of the proposed structures of cytotoxic macrolides iriomoteolide-1a and-1b. Org. Lett. 2010, 12, 3120–3123. [Google Scholar] [CrossRef] [Green Version]
- Parker, L.J.; Taruya, S.; Tsuganezawa, K.; Ogawa, N.; Mikuni, J.; Honda, K.; Tomabechi, Y.; Handa, N.; Shirouzu, M.; Yokoyama, S. Kinase crystal identification and ATP-competitive inhibitor screening using the fluorescent ligand SKF86002. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 392–404. [Google Scholar] [CrossRef]
- Waight, A.B.; Bargsten, K.; Doronina, S.; Steinmetz, M.O.; Sussman, D.; Prota, A.E. Structural basis of microtubule destabilization by potent auristatin anti-mitotics. PLoS ONE 2016, 11, e0160890. [Google Scholar] [CrossRef] [Green Version]
- Wiranowska, M.; Singh, R.; Falahat, R.; Williams, E.; Johnson, J.O.; Alcantar, N. Preferential drug delivery to tumor cells than normal cells using a tunable niosome–chitosan double package nanodelivery system: A novel in vitro model. Cancer Nanotechnol. 2020, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Musa, A.; Elmaidomy, A.H.; Sayed, A.M.; Alzarea, S.I.; Al-Sanea, M.M.; Mostafa, E.M.; Hendawy, O.M.; Abdelgawad, M.A.; Youssif, K.A.; Refaat, H. Cytotoxic potential, metabolic profiling, and liposomes of Coscinoderma sp. crude extract supported by in silico analysis. Int. J. Nanomed. 2021, 16, 3861. [Google Scholar] [CrossRef] [PubMed]
- Refaat, H.; Naguib, Y.W.; Elsayed, M.; Sarhan, H.A.; Alaaeldin, E. Modified spraying technique and response surface methodology for the preparation and optimization of propolis liposomes of enhanced anti-proliferative activity against human melanoma cell line A375. Pharmaceutics 2019, 11, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, R.W.; Gonçalves, M.M.; Fachi, M.M.; Vilhena, R.D.O.; Pontarolo, R.; Maluf, D.F. UPLC–QToF-MS characterization of blackberry extracts of cultivars ‘Tupy’,‘Guarani’, and ‘Xavante’: Development of extract-loaded niosomes. Rev. Bras. De Farmacogn. 2020, 30, 519–527. [Google Scholar] [CrossRef]
- Abouelela, M.E.; Assaf, H.K.; Abdelhamid, R.A.; Elkhyat, E.S.; Sayed, A.M.; Oszako, T.; Belbahri, L.; Zowalaty, A.E.E.; Abdelkader, M.S.A. Identification of potential SARS-CoV-2 main protease and spike protein inhibitors from the genus Aloe: An in silico study for drug development. Molecules 2021, 26, 1767. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.; Rateb, M.E.; Hassan, H.M.; Alonazi, M.; Gomaa, M.R.; Mahrous, N.; GabAllah, M.; Kandeil, A.; Perveen, S.; Abdelmohsen, U.R. Sinapic acid suppresses SARS CoV-2 replication by targeting its envelope protein. Antibiotics 2021, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Thissera, B.; Sayed, A.M.; Hassan, M.H.; Abdelwahab, S.F.; Amaeze, N.; Semler, V.T.; Alenezi, F.N.; Yaseen, M.; Alhadrami, H.A.; Belbahri, L. Bioguided isolation of cyclopenin analogues as potential SARS-CoV-2 Mpro inhibitors from Penicillium citrinum TDPEF34. Biomolecules 2021, 11, 1366. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassermann, A.M.; Bajorath, J. BindingDB and ChEMBL: Online compound databases for drug discovery. Expert Opin. Drug Discov. 2011, 6, 683–687. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [Green Version]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Franz, M.; Lopes, C.T.; Huck, G.; Dong, Y.; Sumer, O.; Bader, G.D. Cytoscape.js: A graph theory library for visualisation and analysis. Bioinformatics 2016, 32, 309–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.Y.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36 (Suppl. S1), D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou-Taleb, H.A.; Sayed, A.M.; Refaat, H.; Alsenani, F.; Alaaeldin, E.; Mokhtar, F.A.; Abdelmohsen, U.R.; Shady, N.H. Network Pharmacological Analysis of the Red Sea Sponge Hyrtios erectus Extract to Reveal Anticancer Efficacy of Corresponding Loaded Niosomes. Mar. Drugs 2022, 20, 628. https://doi.org/10.3390/md20100628
Abou-Taleb HA, Sayed AM, Refaat H, Alsenani F, Alaaeldin E, Mokhtar FA, Abdelmohsen UR, Shady NH. Network Pharmacological Analysis of the Red Sea Sponge Hyrtios erectus Extract to Reveal Anticancer Efficacy of Corresponding Loaded Niosomes. Marine Drugs. 2022; 20(10):628. https://doi.org/10.3390/md20100628
Chicago/Turabian StyleAbou-Taleb, Heba A., Ahmed M. Sayed, Hesham Refaat, Faisal Alsenani, Eman Alaaeldin, Fatma A. Mokhtar, Usama Ramadan Abdelmohsen, and Nourhan Hisham Shady. 2022. "Network Pharmacological Analysis of the Red Sea Sponge Hyrtios erectus Extract to Reveal Anticancer Efficacy of Corresponding Loaded Niosomes" Marine Drugs 20, no. 10: 628. https://doi.org/10.3390/md20100628
APA StyleAbou-Taleb, H. A., Sayed, A. M., Refaat, H., Alsenani, F., Alaaeldin, E., Mokhtar, F. A., Abdelmohsen, U. R., & Shady, N. H. (2022). Network Pharmacological Analysis of the Red Sea Sponge Hyrtios erectus Extract to Reveal Anticancer Efficacy of Corresponding Loaded Niosomes. Marine Drugs, 20(10), 628. https://doi.org/10.3390/md20100628