
In Vivo Anticoagulant and Antithrombic Activity of Depolymerized Glycosaminoglycan from Apostichopus japonicus and Dynamic Effect–Exposure Relationship in Rat Plasma

 and
and
Abstract
:1. Introduction
2. Results
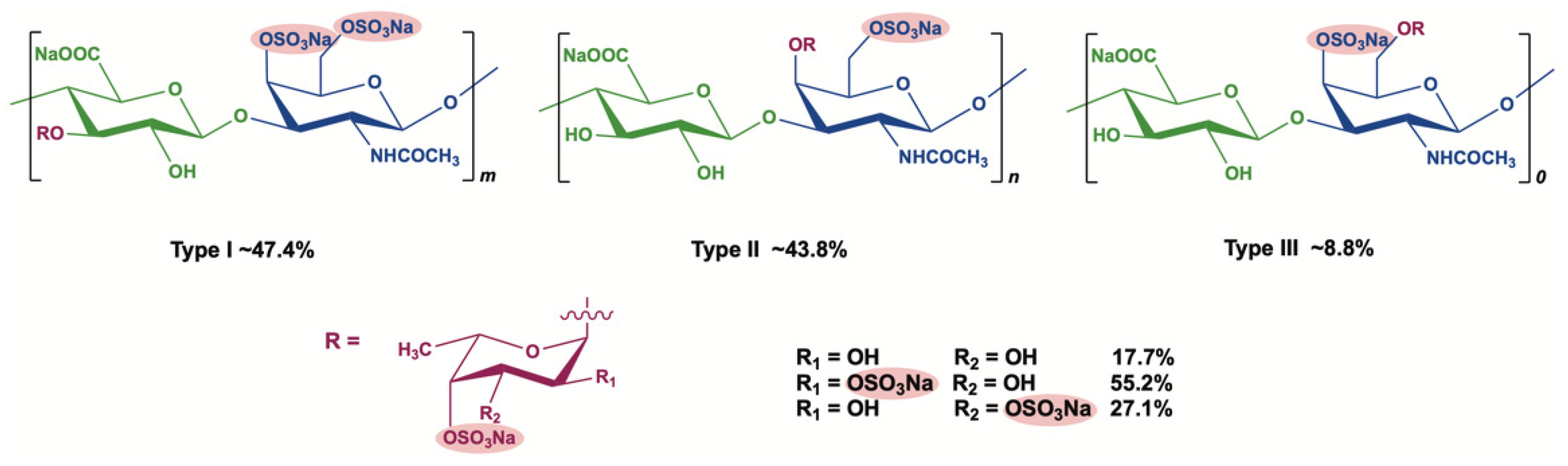
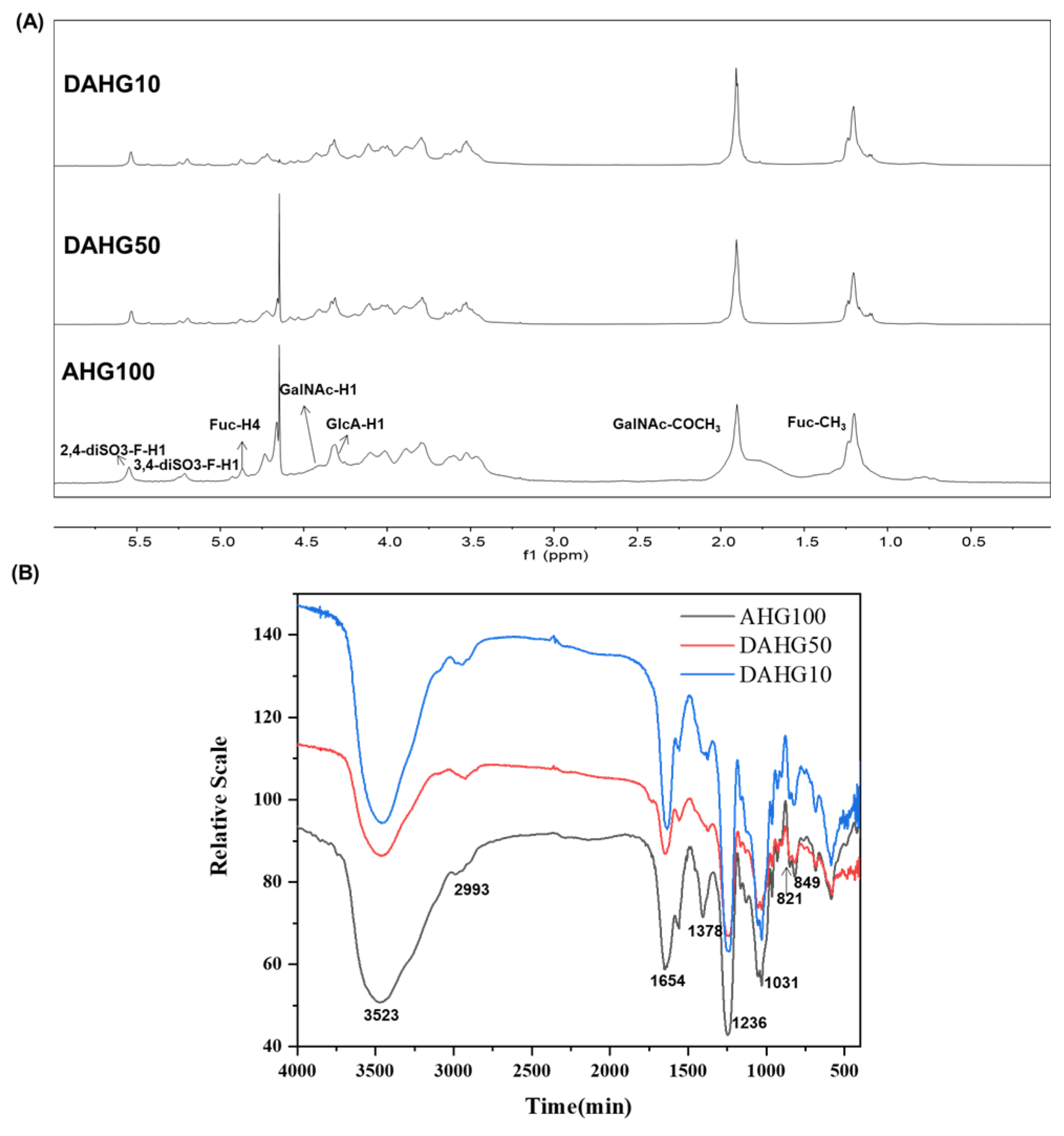
2.1. Characterization of AHG100 and DAHGs
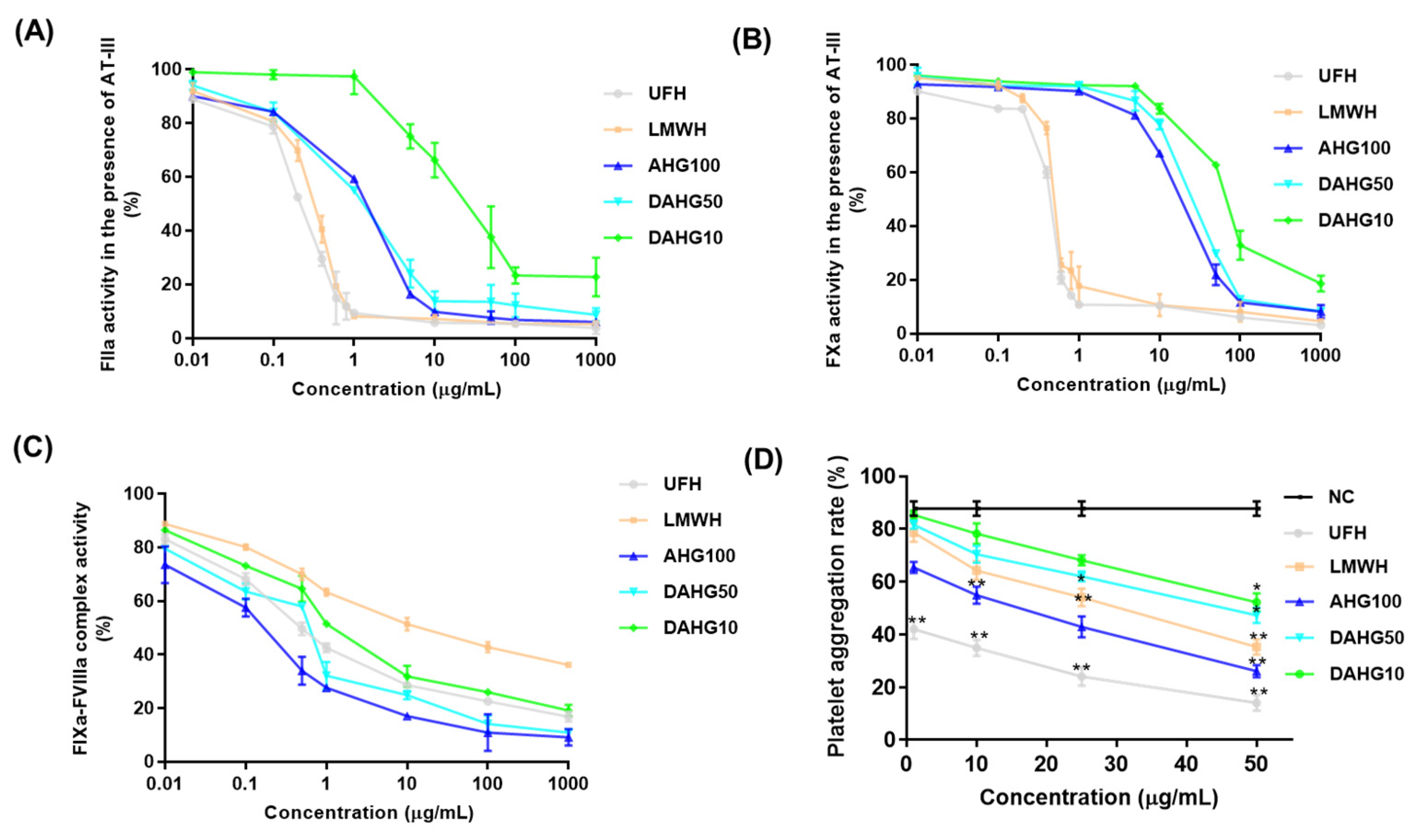
2.2. Inhibitory Effects of AHG100 and DAHGs on Coagulation (Co)Factors In Vitro
2.3. Effect of AHG100 and DAHGs on Thrombin-Induced Platelet Aggregation
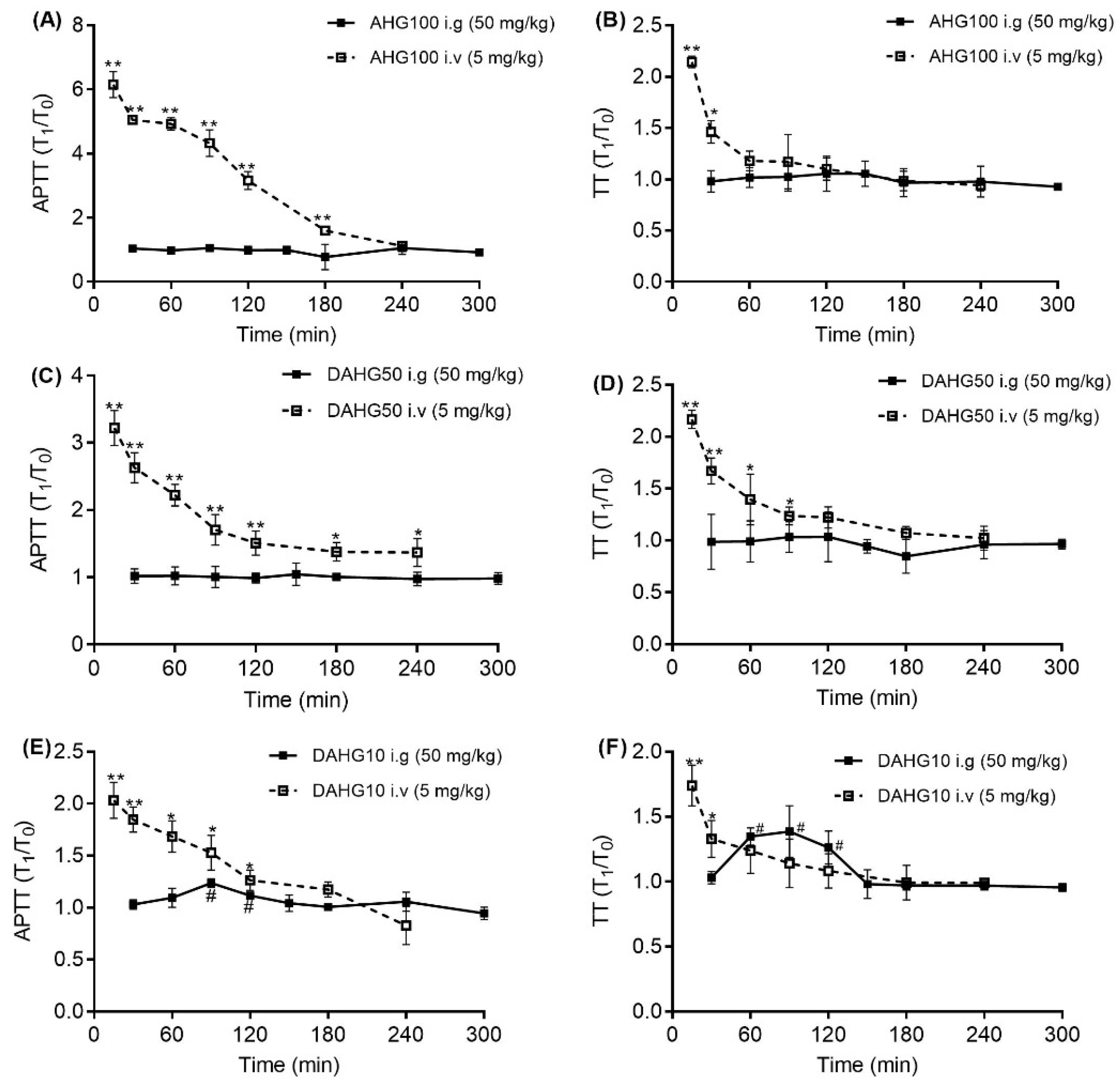
2.4. Effect of AHG100 and DAHGs on Clotting Time (CT) and Bleeding Time (BT)
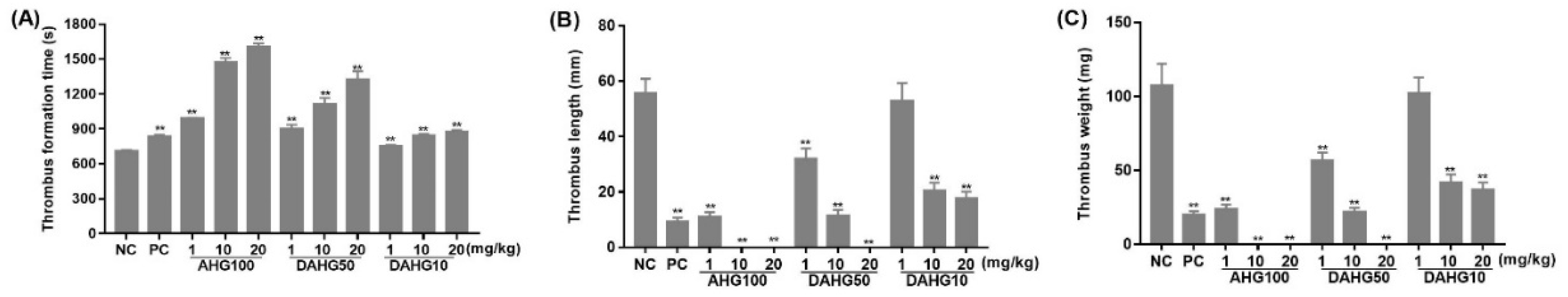
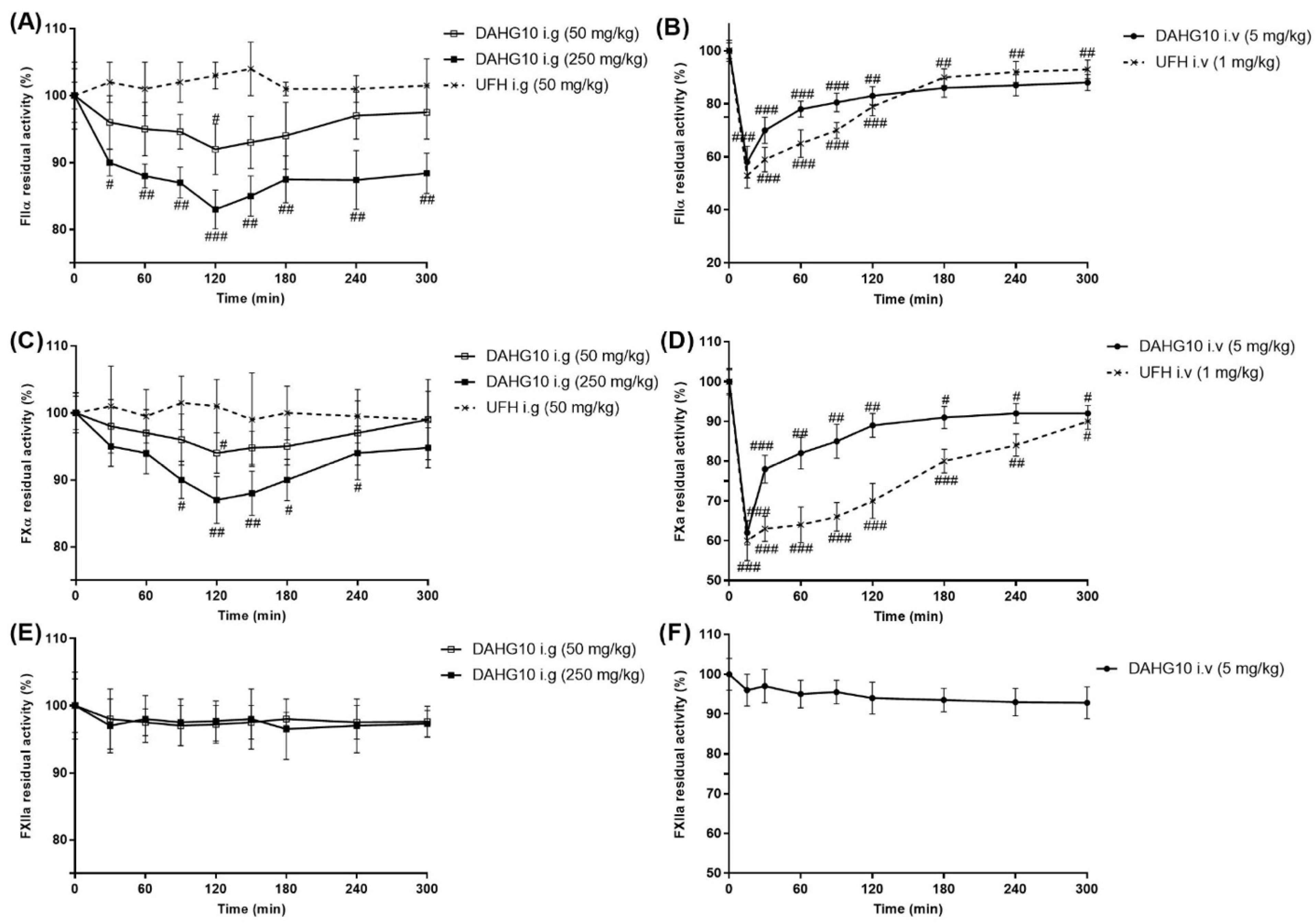
2.5. Anti-Thrombus Action of AHG100 and DAHGs
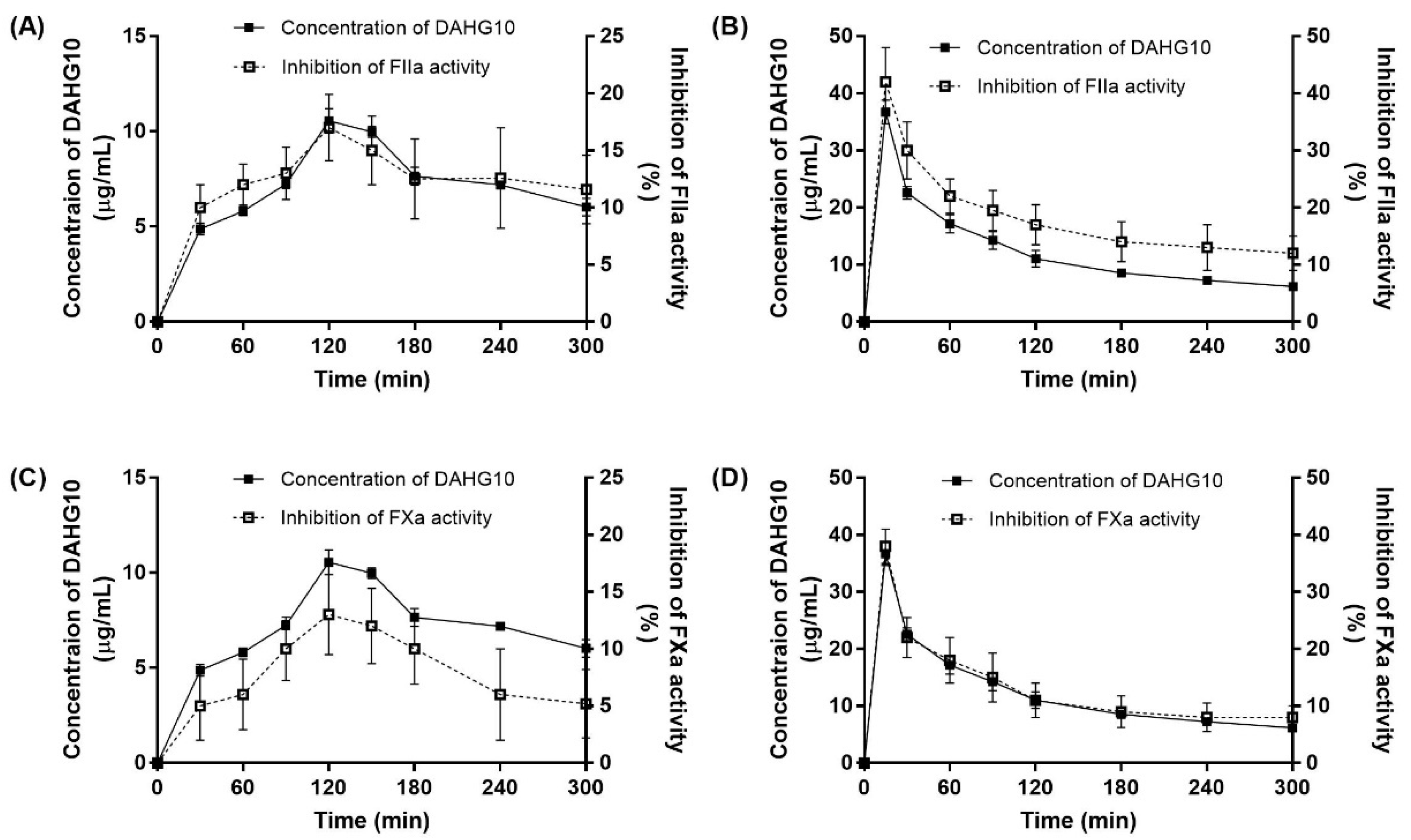
2.6. Correlation between Exposure and Anticoagulant Activity in Rats
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. The Preparation of AHG100 and DAHGs
4.3. Effects of AHG100 and DAHGs on Coagulation (Co)Factors In Vitro
4.4. Effect of AHG100 and DAHGs on Thrombin-Induced Platelet Aggregation
4.5. Effect of AHG100 and DAHGs on CT and BT
4.6. Anti-Thrombus Action of AHG100 and DAHGs in Rats
4.7. Correlation between Exposure and Anticoagulant Activity in Rats
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Egidy Assenza, G.; Dimopoulos, K.; Budts, W.; Donti, A.; Economy, K.E.; Gargiulo, G.D.; Gatzoulis, M.; Landzberg, M.J.; Valente, A.M.; Roos-Hesselink, J. Management of acute cardiovascular complications in pregnancy. Eur. Heart J. 2021, 42, 4224–4240. [Google Scholar] [CrossRef]
- Forouzandehmehr, M.; Ghoytasi, I.; Shamloo, A.; Ghosi, S. Particles in coronary circulation: A review on modelling for drug carrier design. Mater. Des. 2022, 216, 110511–110522. [Google Scholar] [CrossRef]
- Levy, J.H.; Douketis, J.; Weitz, J.I. Reversal agents for non-vitamin k antagonist oral anticoagulants. Nat. Rev. Cardiol. 2018, 15, 273–281. [Google Scholar] [CrossRef]
- Kelton, J.G.; Warkentin, T.E. Heparin-induced thrombocytopenia: A historical perspective. Blood 2008, 112, 2607–2616. [Google Scholar] [CrossRef] [Green Version]
- Onishi, A.; St Ange, K.; Dordick, J.S.; Linhardt, R.J. Heparin and anticoagulation. Front. Biosci. 2016, 21, 1372–1392. [Google Scholar] [CrossRef]
- Schulman, S.; Kakkar, A.K.; Goldhaber, S.Z.; Schellong, S.; Eriksson, H.; Mismetti, P.; Christiansen, A.V.; Friedman, J.; Maulf, F.L.; Peter, N.; et al. Treatment of acute venous thromboembolism with dabigatran or warfarin and pooled analysis. Circulation 2014, 129, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, H.; Lin, L.; Yao, W.; Zhao, J.; Wu, M.; Li, Z. Synthesis of fucosylated chondroitin sulfate nonasaccharide as a novel anticoagulant targeting intrinsic factor Xase complex. Angew. Chem. Int. Ed. Engl. 2018, 57, 12880–12885. [Google Scholar] [CrossRef]
- Eikelboom, J.W.; Zelenkofske, S.L.; Rusconi, C.P. Coagulation factor IXa as a target for treatment and prophylaxis of venous thromboembolism. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, R.J.; Santos, G.R.; Mourão, P.A. Effects of polysaccharides enriched in 2,4-disulfated fucose units on coagulation, thrombosis and bleeding. Practical and conceptual implications. Thromb. Haemost. 2009, 102, 829–836. [Google Scholar] [CrossRef] [Green Version]
- Nagase, H.; Enjyoji, K.; Minamiguchi, K.; Kitazato, K.; Kitazato, K.; Saito, H.; Kato, H. Depolymerized holothurian glycosaminoglycan with novel anticoagulant actions: Antithrombin III- and heparin cofactor II-independent inhibition of factor X activation by factor IXa-factor VIIIa complex and heparin cofactor II-dependent inhibition of thrombin. Blood 1995, 85, 1527–1534. [Google Scholar]
- Wu, M.; Wen, D.; Gao, N.; Xiao, C.; Yang, L.; Xu, L.; Lian, W.; Peng, W.; Jiang, J.; Zhao, J. Anticoagulant and antithrombotic evaluation of native fucosylated chondroitin sulfates and their derivatives as selective inhibitors of intrinsic factor Xase. Eur. J. Med. Chem. 2015, 92, 257–269. [Google Scholar] [CrossRef]
- Xiao, C.; Lian, W.; Zhou, L.; Gao, N.; Xu, L.; Chen, J.; Wu, M.; Peng, W.; Zhao, J. Interactions between depolymerized fucosylated glycosaminoglycan and coagulation proteases or inhibitors. Thromb. Res. 2016, 146, 59–68. [Google Scholar] [CrossRef]
- Fonseca, R.J.C.; Mouro, P.A.S. Fucosylated chondroitin sulfate as a new oral antithrombotic agent. Thromb. Haemost. 2007, 96, 822–829. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, M.; Wang, D.; Tao, W.; Liu, D.; Zhang, F.; Linhardt, R.J.; Ye, X.; Chen, S. Oral administration of fucosylated chondroitin sulfate oligomers in gastro-resistant microcapsules exhibits a safe antithrombotic activity. Thromb. Haemost. 2021, 121, 15–26. [Google Scholar] [CrossRef]
- Imanari, T.; Washio, Y.; Huang, Y.; Toyoda, H.; Suzuki, A.; Toida, T. Oral absorption and clearance of partially depolymerized fucosyl chondroitin sulfate from sea cucumber. Thromb. Res. 1999, 93, 129–135. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Jiang, T.; Lv, L.; Zhang, B.; Lv, Z. Depolymerized glycosaminoglycan and its anticoagulant activities from sea cucumber Apostichopus japonicus. Int. J. Biol. Macromol. 2015, 72, 699–705. [Google Scholar] [CrossRef]
- Ma, M.; Wang, H.; Yuan, Y.; Wang, Y.; Yang, S.; Lv, Z. The absorption of glycosaminoglycans of different molecular weight obtained from Apostichopus japonicus: An in vitro and in situ study. Food Funct. 2021, 12, 5551–5562. [Google Scholar] [CrossRef]
- Chen, R.; Yin, R.; Li, S.; Pan, Y.; Mao, H.; Cai, Y.; Lin, L.; Wang, W.; Zhang, T.; Zhou, L.; et al. Structural characterization of oligosaccharides from free radical depolymerized fucosylated glycosaminoglycan and suggested mechanism of depolymerization. Carbohydr. Polym. 2021, 270, 118368. [Google Scholar] [CrossRef]
- Wu, M.; Xu, S.; Zhao, J.; Kang, H.; Ding, H. Physicochemical characteristics and anticoagulant activities of low molecular weight fractions by free-radical depolymerization of a fucosylated chondroitin sulphate from sea cucumber Thelenata ananas. Food Chem. 2010, 122, 716–723. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, K.A. Antithrombotic activity of methanolic extract of Umbilicaria esculenta. J. Ethnopharmacol. 2006, 105, 342–345. [Google Scholar] [CrossRef]
- Adhana, R.; Chaurasiya, R.; Verma, A. Comparison of bleeding time and clotting time between males and females. J. Physiol. Pharmacol. 2018, 8, 1388–1390. [Google Scholar] [CrossRef]
- He, D.; Wang, Y.; Yang, S.; Lv, Z. LC-MS/MS method for the determination of the concentration of depolymerized Apostichopus japonicus holothurian glycosaminoglycans in rat plasma and its preliminary pharmacokinetics. Chin. J. Mar. Drugs 2020, 39, 52–58. [Google Scholar]
- Lin, L.; Zhao, L.; Gao, N.; Yin, R.; Li, S.; Sun, H.; Zhou, L.; Zhao, G.; Purcell, S.W.; Zhao, J. From multi-target anticoagulants to doacs, and intrinsic coagulation factor inhibitors. Blood Rev. 2020, 39, 100615–100623. [Google Scholar] [CrossRef]
- Li, H.; Yuan, Q.; Lv, K.; Ma, H.; Gao, C.; Liu, Y.; Zhang, S.; Zhao, L. Low-molecular-weight fucosylated glycosaminoglycan and its oligosaccharides from sea cucumber as novel anticoagulants: A review. Carbohydr. Polym. 2021, 251, 117034–117044. [Google Scholar] [CrossRef]
- Lin, L.; Xu, L.; Xiao, C.; Zhou, L.; Gao, N.; Wu, M.; Zhao, J. Plasma contact activation by a fucosylated chondroitin sulfate and its structure–activity relationship study. Glycobiology 2018, 28, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Sun, H.; Xiao, C.; Yang, L.; Lin, L.; Yin, R.; Li, Z.; Zhang, H.; Ji, X.; Zhao, J. Effects of native fucosylated glycosaminoglycan, its depolymerized derivatives on intrinsic factor Xase, coagulation, thrombosis, and hemorrhagic risk. Thromb. Haemost. 2020, 120, 607–619. [Google Scholar] [CrossRef]
- Zhao, L.; Qin, Y.; Guan, R.; Zheng, W.; Liu, J.; Zhao, J. Digestibility of fucosylated glycosaminoglycan from sea cucumber and its effects on digestive enzymes under simulated salivary and gastrointestinal conditions. Carbohydr. Polym. 2018, 186, 217–225. [Google Scholar] [CrossRef]
- Zhu, Z.; Dong, X.; Yan, C.; Ai, C.; Zhou, D.; Yang, J.; Zhang, H.; Liu, X.; Song, S.; Xiao, H.; et al. Structural features and digestive behavior of fucosylated chondroitin sulfate from sea cucumbers Stichopus japonicus. J. Agric. Food Chem. 2019, 67, 10534–10542. [Google Scholar] [CrossRef]
- Kamikubo, Y.; Mendolicchio, G.L.; Zampolli, A.; Marchese, P.; Gale, A.J.; Krishnaswamy, S.; Gruber, A.; Petersen, L.C.; Ruf, W.; Ruggeri, Z.M. Distinct mechanism of anti-hemophilic FVIII activation by nascent FXa in the tissue factor-FVIIa coagulation initiation complex resistant to FXa-directed inhibition. Blood 2014, 124, 2803. [Google Scholar] [CrossRef]
- Neves, A.R.; Correia-da-Silva, M.; Sousa, E.; Pinto, M. Strategies to overcome heparins’ low oral bioavailability. Pharmaceuticals 2016, 9, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Li, J.; Wang, D.; Ding, T.; Hu, Y.; Ye, X.; Linhardt, R.J.; Chen, S. Molecular size is important for the safety and selective inhibition of intrinsic factor Xase for fucosylated chondroitin sulfate. Carbohydr. Polym. 2017, 178, 180–189. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Jiang, T.; Lv, Z. Novel branch patterns and anticoagulant activity of glycosaminoglycan from sea cucumber Apostichopus japonicus. Int. J. Biol. Macromol. 2015, 72, 911–918. [Google Scholar] [CrossRef]
- Born, G.V.R. Aggregation of bood platelets by adenosine diphosphate and its reversal. Nature 1962, 194, 927–929. [Google Scholar] [CrossRef]
- Born, G.V.; Cross, M.J. The aggregation of blood platelets. J. Physiol. 1963, 168, 178–195. [Google Scholar] [CrossRef]
- Wang, Y.; Shao, J.; Yao, S.; Zhang, S.; Yan, J.; Wang, H.; Chen, Y. Study on the antithrombotic activity of Umbilicaria esculenta polysaccharide. Carbohydr. Polym. 2014, 105, 231–236. [Google Scholar] [CrossRef]
- Zhang, Y.; Huo, M.; Zhou, J.; Xie, S. Pksolver: An add-in program for pharmacokinetic and pharmacodynamic data analysis in microsoft excel. Comput. Methods Programs Biomed. 2010, 99, 306–314. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Mw (Da) | Chemical Composition (Molar Ratio) | ||
|---|---|---|---|---|
| GlcA | GalN | Fuc | ||
| AHG100 | 94,156 | 1.04 | 1.00 | 0.90 |
| DAHG50 | 51,318 | 1.02 | 1.00 | 0.91 |
| DAHG10 | 9972 | 1.02 | 1.00 | 0.94 |
| Parameter | DAHG10 |
|---|---|
| T1/2 (min) | 367.92 ± 169.29 |
| Tmax (min) | 120 |
| Cmax (mg/mL) | 10.56 ± 0.66 |
| AUC0–t (min·μg/mL) | 2111.49 ± 39.75 |
| AUC0–∞ (min·μg/mL) | 5373.71 ± 1578.63 |
| Vd (mL/kg) | 23,655.92 ± 4718.83 |
| MRT0–t (min) | 159.70 ± 1.07 |
| MRT0–∞ (min) | 571.78 ± 227.69 |
| Cl (mL/min/kg) | 50.08 ± 16.40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; He, D.; Duan, L.; Lv, L.; Gao, Q.; Wang, Y.; Yang, S.; Lv, Z. In Vivo Anticoagulant and Antithrombic Activity of Depolymerized Glycosaminoglycan from Apostichopus japonicus and Dynamic Effect–Exposure Relationship in Rat Plasma. Mar. Drugs 2022, 20, 631. https://doi.org/10.3390/md20100631
Wang H, He D, Duan L, Lv L, Gao Q, Wang Y, Yang S, Lv Z. In Vivo Anticoagulant and Antithrombic Activity of Depolymerized Glycosaminoglycan from Apostichopus japonicus and Dynamic Effect–Exposure Relationship in Rat Plasma. Marine Drugs. 2022; 20(10):631. https://doi.org/10.3390/md20100631
Chicago/Turabian StyleWang, Han, Dandan He, Linlin Duan, Lv Lv, Qun Gao, Yuanhong Wang, Shuang Yang, and Zhihua Lv. 2022. "In Vivo Anticoagulant and Antithrombic Activity of Depolymerized Glycosaminoglycan from Apostichopus japonicus and Dynamic Effect–Exposure Relationship in Rat Plasma" Marine Drugs 20, no. 10: 631. https://doi.org/10.3390/md20100631
APA StyleWang, H., He, D., Duan, L., Lv, L., Gao, Q., Wang, Y., Yang, S., & Lv, Z. (2022). In Vivo Anticoagulant and Antithrombic Activity of Depolymerized Glycosaminoglycan from Apostichopus japonicus and Dynamic Effect–Exposure Relationship in Rat Plasma. Marine Drugs, 20(10), 631. https://doi.org/10.3390/md20100631