
An Overview of Potential Seaweed-Derived Bioactive Compounds for Pharmaceutical Applications



Abstract
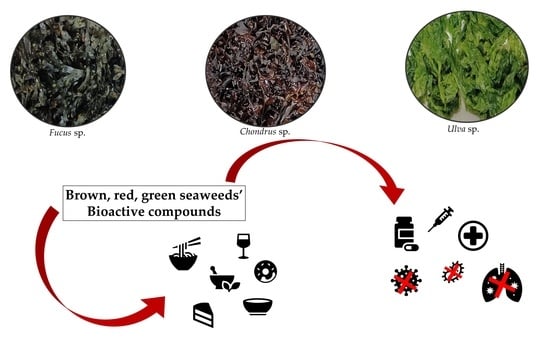
:
1. Introduction
2. Socio-Ecological Relevance of Seaweeds and Classification
Bioactive Compounds of Seaweeds and Their Potential to Ameliorate Human Health and Welfare

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Seaweed Bioactive Compounds | Application and Properties | Principal Source | Reference | |
|---|---|---|---|---|
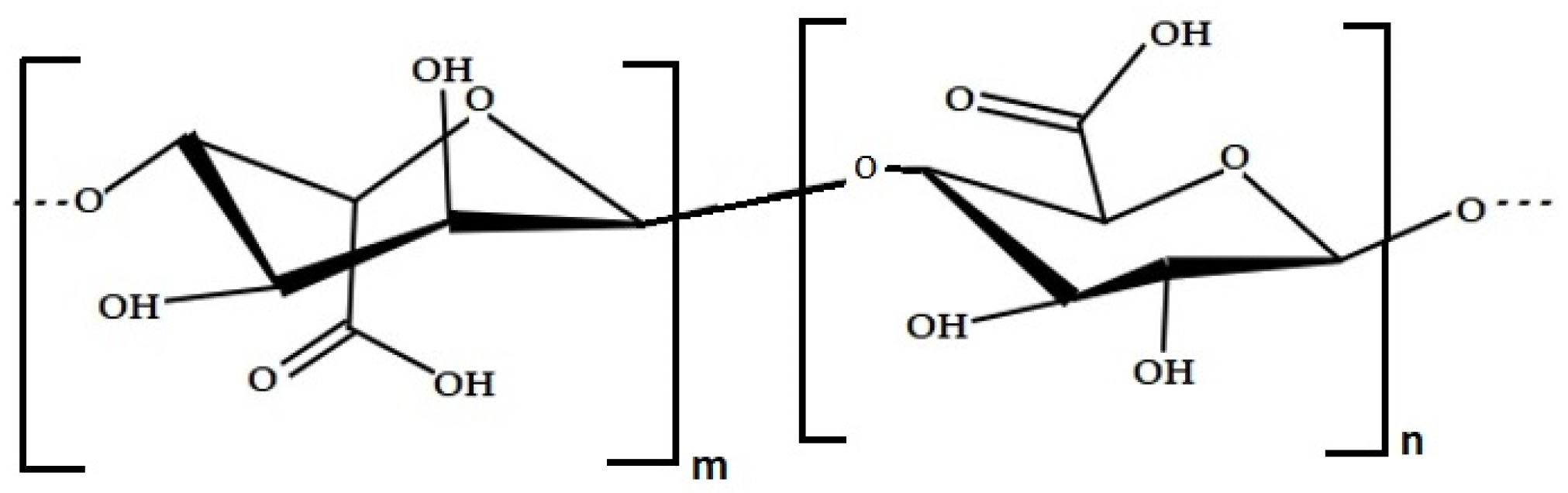
| Polysaccharides | Alginate | Used as stabilizer and thickening agent in food products and medicine | Brown seaweed (Laminaria sp., Ascophyllum nodosum) | [71,73,74] |
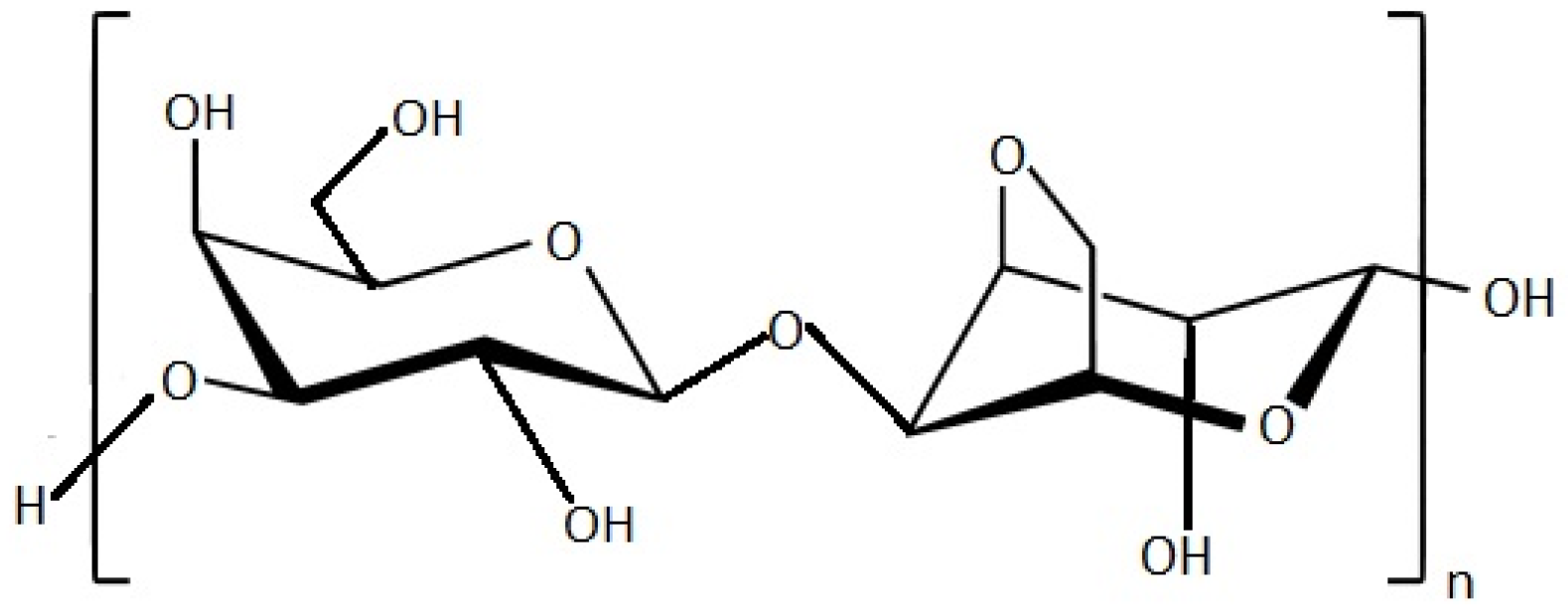
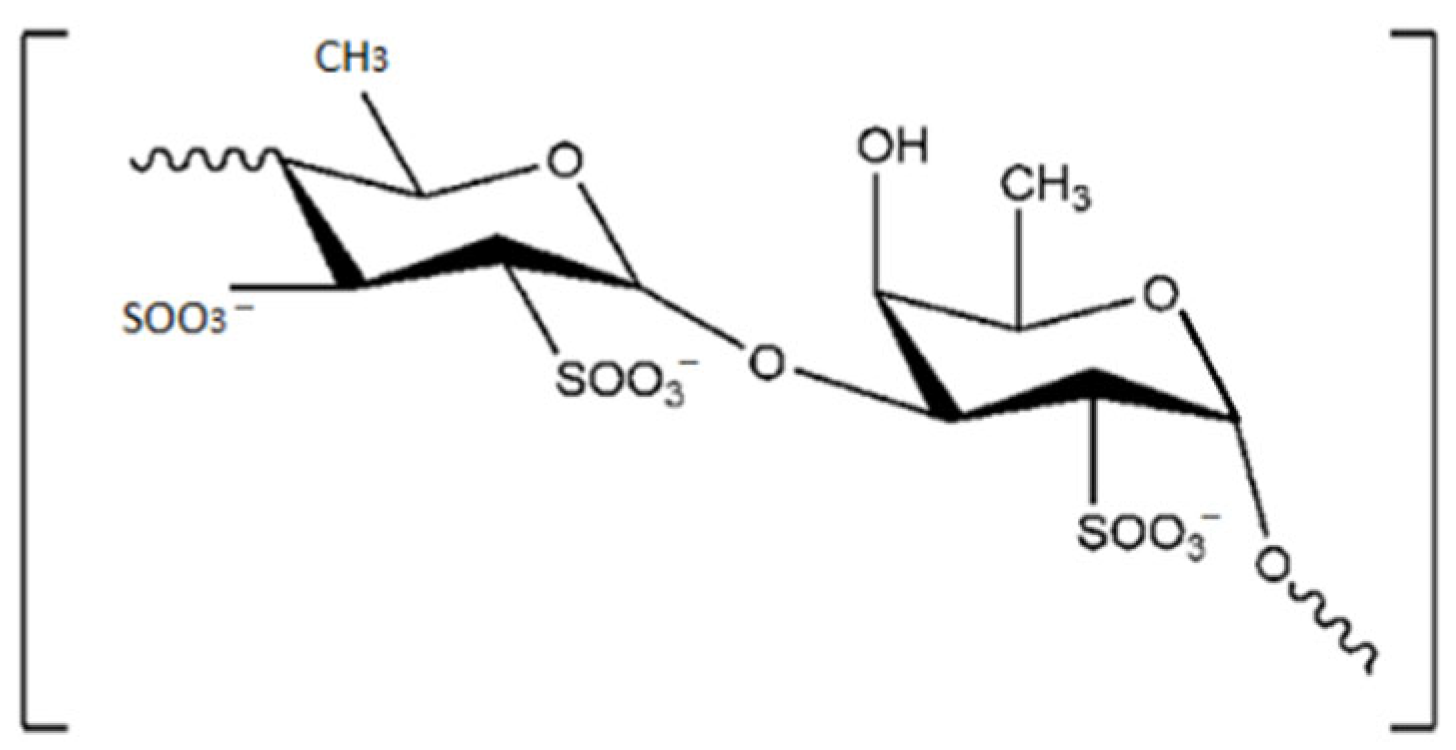
| Fucoidan | Antiproliferative, Antimicrobial and Antiviral activity Anticoagulant activity Antidiabetic activity | Brown seaweed (Undaria pinnatifida, Fucus sp.) | [105,106,107,108,109,110,111,112,113,114] | |
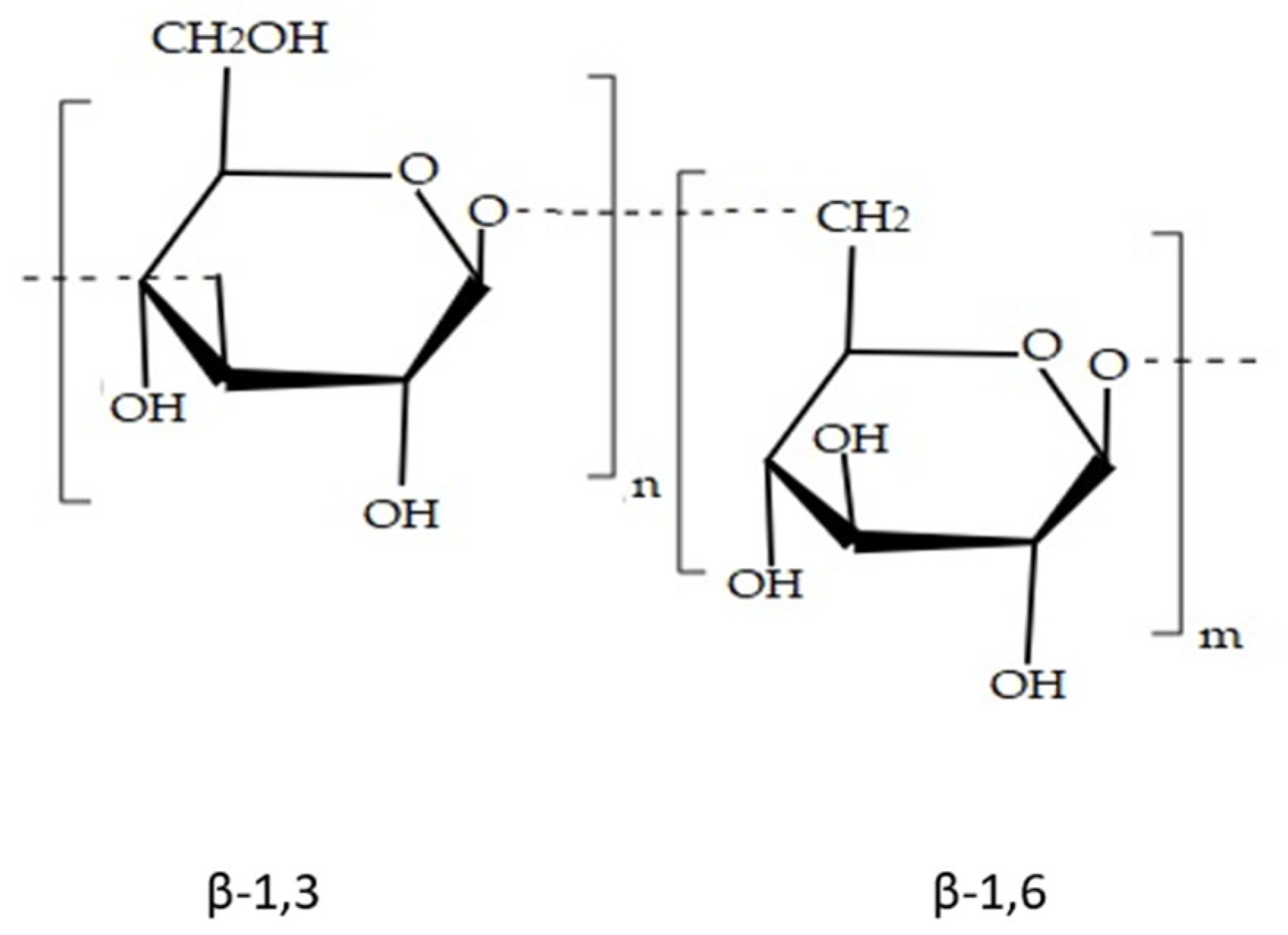
| Laminarin | Used in food industry and biomedicine because of its nutraceutical properties; immunostimulatory, antitumour and antioxidant activity | Brown seaweed (Laminaria sp.) | [75,76,77,115] | |
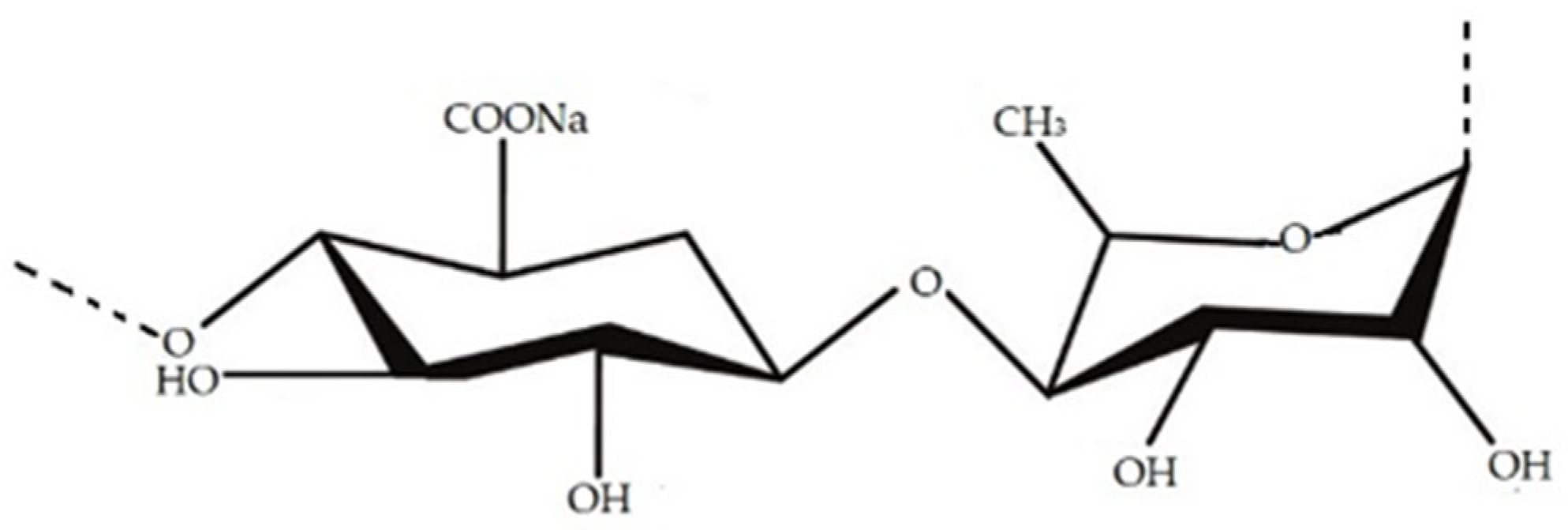
| Agar | Used in food products and pharmaceutical field as jellifiers, stabilisers, thickeners and emulsifiers | Red seaweed (Gracilaria sp.) | [116,117,118,119,120] | |
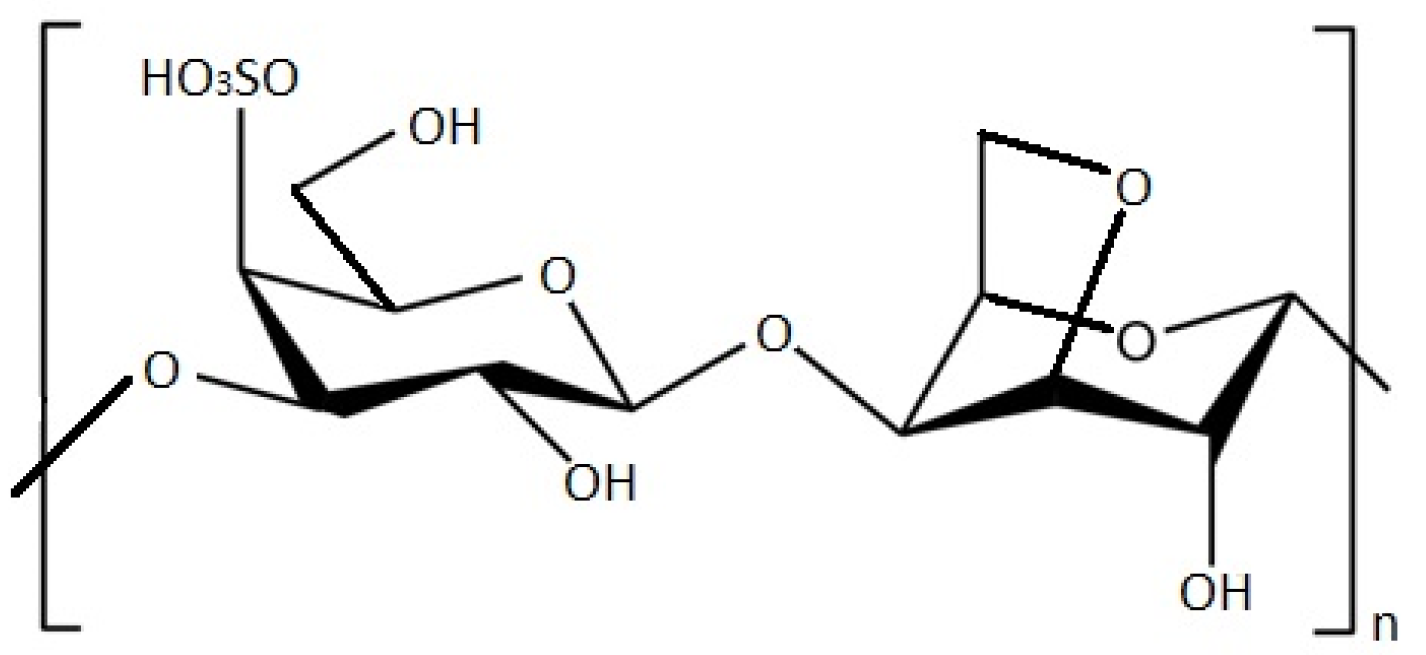
| Carrageenan | Red seaweed (Gigartina sp., Chondrus sp.) | [116,117,121,122,123] | ||
| Porphyran | Anti-inflammatory, antioxidant, antihyperlipidemic and anticancer activities | Red seaweed (Porphyra sp.) | [59,60,61] | |
| Ulvan | Immunostimulatory, antitumoural, antiviral activities | Green seaweed (Ulva sp.) | [62,79,80,81,124] | |
| Polyphenols | Phlorotannin | antimicrobial, antioxidant, antiviral, anticancer, anti-inflammatory, antidiabetic properties | Brown seaweed (Ecklonia sp., Eisenia sp., Laminaria sp., Undaria pinnafitida) | [86,87,88,90,91,125] |
3. Seaweeds in Pharmaceutical Studies and Applications
3.1. Phylum Ochrophyta, Class Phaeophyceae
3.2. Phylum Rhodophyta
3.3. Phylum Chlorophyta
4. Use of Seaweeds in Traditional and Modern Pharmacology
5. Main Conclusions of the Pharmaceutical Potential of Marine Macroalgae and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lyu, M.; Wang, Y.F.; Fan, G.W.; Wang, X.Y.; Xu, S.Y.; Zhu, Y. Balancing herbal medicine and functional food for prevention and treatment of cardiometabolic diseases through modulating gut microbiota. Front. Microbiol. 2017, 8, 2146. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L. Biological and therapeutic properties of the seaweed polysaccharides. Int. Biol. Rev. 2018, 2, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2011, 23, 321–335. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Vaskovsky, V.E.; Titlyanova, T.V. Fatty acids of marine algae from the pacific coast of North California. Bot. Mar. 2002, 45, 17–22. [Google Scholar] [CrossRef]
- Agregán, R.; Munekata, P.E.; Domínguez, R.; Carballo, J.; Franco, D.; Lorenzo, J.M. Proximate composition, phenolic content and in vitro antioxidant activity of aqueous extracts of the seaweeds Ascophyllum nodosum, Bifurcaria bifurcata and Fucus vesiculosus. Effect of addition of the extracts on the oxidative stability of canola oil unde. Food Res. Int. 2017, 99, 986–994. [Google Scholar] [CrossRef]
- Rajapakse, N.; Kim, S.K. Nutritional and Digestive Health Benefits of Seaweed. In Advances in Food and Nutrition Research, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 64, ISBN 9780123876690. [Google Scholar]
- Charoensiddhi, S.; Conlon, M.A.; Franco, C.M.M.; Zhang, W. The development of seaweed-derived bioactive compounds for use as prebiotics and nutraceuticals using enzyme technologies. Trends Food Sci. Technol. 2017, 70, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, J.; Wang, S.; Xu, X. Porphyra species: A mini-review of its pharmacological and nutritional properties. J. Med. Food 2016, 19, 111–119. [Google Scholar] [CrossRef]
- Arvinda Swamy, M.L. Chapter 6: Marine Algal Sources for Treating Bacterial Diseases. In Advances in Food and Nutrition Research, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 64, ISBN 9780123876690. [Google Scholar]
- Gheda, S.F.; El-Adawi, H.I.; El-Deeb, N.M. Antiviral Profile of Brown and Red Seaweed Polysaccharides Against Hepatitis C Virus. Iran. J. Pharm. Res. IJPR 2016, 15, 483–491. [Google Scholar]
- Duarte, C.M.; Wu, J.; Xiao, X.; Bruhn, A.; Krause-Jensen, D. Can seaweed farming play a role in climate change mitigation and adaptation? Front. Mar. Sci. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Kim, J.K.; Yarish, C.; Hwang, E.K.; Park, M.; Kim, Y. Seaweed aquaculture: Cultivation technologies, challenges and its ecosystem services. Algae 2017, 32, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cabral, P.; Levrel, H.; Viard, F.; Frangoudes, K.; Girard, S.; Scemama, P. Ecosystem services assessment and compensation costs for installing seaweed farms. Mar. Policy 2016, 71, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Hasselström, L.; Visch, W.; Gröndahl, F.; Nylund, G.M.; Pavia, H. The impact of seaweed cultivation on ecosystem services—A case study from the west coast of Sweden. Mar. Pollut. Bull. 2018, 133, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Food Agriculture Organization. De La Pesca Y La Acuicultura; Food Agriculture Organization: Rome, Italy, 2020; Volume 3, ISBN 0000000340472. [Google Scholar]
- Dixit, D.C.; Reddy, C.R.K.; Balar, N.; Suthar, P.; Gajaria, T.; Gadhavi, D.K. Assessment of the Nutritive, Biochemical, Antioxidant and Antibacterial Potential of Eight Tropical Macro algae Along Kachchh Coast, India as Human Food Supplements. J. Aquat. Food Prod. Technol. 2018, 27, 61–79. [Google Scholar] [CrossRef]
- McLachlan, J. Macroalgae (seaweeds): Industrial resources and their utilization. Biosalin. Action Bioprod. Saline Water 1985, 157, 137–157. [Google Scholar] [CrossRef]
- Barba, F.J. Microalgae and seaweeds for food applications: Challenges and perspectives. Food Res. Int. 2017, 99, 969–970. [Google Scholar] [CrossRef]
- Pereira, L.; Amado, A.M.; Critchley, A.T.; van de Velde, F.; Ribeiro-Claro, P.J.A. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FT-Raman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef] [Green Version]
- Mabeau, S.; Fleurence, J. Seaweed in food products: Biochemical and nutritional aspects. Trends Food Sci. Technol. 1993, 4, 103–107. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F.J. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef] [Green Version]
- De Melo, N.S.M.; Cardoso, L.G.; de Castro Nunes, J.M.; Brito, G.B.; Caires, T.A.; de Souza, C.O.; Portz, L.; Druzian, J.I. Effects of dry and rainy seasons on the chemical composition of Ulva fasciata, Crassiphycus corneus, and Sargassum vulgare seaweeds in tropical environment. Rev. Bras. Bot. 2021, 44, 331–344. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, M.; Alalawy, A.I.; Almutairi, F.M.; Al-Duais, M.A.; Wang, J.; Salama, E.S. Identification and characterization of marine seaweeds for biocompounds production. Environ. Technol. Innov. 2021, 24, 101848. [Google Scholar] [CrossRef]
- Hamid, S.S.; Wakayama, M.; Ichihara, K.; Sakurai, K.; Ashino, Y.; Kadowaki, R.; Soga, T.; Tomita, M. Metabolome profiling of various seaweed species discriminates between brown, red, and green algae. Planta 2019, 249, 1921–1947. [Google Scholar] [CrossRef] [PubMed]
- Cermeño, M.; Kleekayai, T.; Amigo-Benavent, M.; Harnedy-Rothwell, P.; FitzGerald, R.J. Current knowledge on the extraction, purification, identification, and validation of bioactive peptides from seaweed. Electrophoresis 2020, 41, 1694–1717. [Google Scholar] [CrossRef] [PubMed]
- Seely, G.R.; Duncan, M.J.; Vidaver, W.E. Preparative and analytical extraction of pigments from brown algae with dimethyl sulfoxide. Mar. Biol. 1972, 12, 184–188. [Google Scholar] [CrossRef]
- Denis, C.; Ledorze, C.; Jaouen, P.; Fleurence, J. Comparison of different procedures for the extraction and partial purification of R-phycoerythrin from the red macroalga Grateloupia turuturu. Bot. Mar. 2009, 52, 278–281. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Algal carotenoids 54. Carotenoids of brown algae (Phaeophyceae). Biochem. Syst. Ecol. 1994, 22, 31–41. [Google Scholar] [CrossRef]
- Aryee, A.N.; Agyei, D.; Akanbi, T.O. Recovery and utilization of seaweed pigments in food processing. Curr. Opin. Food Sci. 2018, 19, 113–119. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, G.A.; Hearst, J.E. Genetics and molecular biology of carotenoid pigment biosynthesis. FASEB J. 1996, 10, 228–237. [Google Scholar] [CrossRef]
- Dumay, J.; Morançais, M.; Munier, M.; Le Guillard, C.; Fleurence, J. Phycoerythrins: Valuable Proteinic Pigments in Red Seaweeds; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71, ISBN 9780124080621. [Google Scholar]
- Pangestuti, R.; Kim, S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Suleria, H.; Osborne, S.; Masci, P.; Gobe, G. Marine-Based Nutraceuticals: An Innovative Trend in the Food and Supplement Industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [Green Version]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Kuhnle, G.G.; et al. Re-evaluation of carrageenan (E 407) and processed Eucheuma seaweed (E 407a) as food additives. EFSA J. 2018, 16, e05238. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Kim, S.K. Present and Future Prospects of Seaweeds in Developing Functional Foods. In Advances in Food and Nutrition Research, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 64, ISBN 9780123876690. [Google Scholar]
- Tanna, B.; Mishra, A. Nutraceutical Potential of Seaweed Polysaccharides: Structure, Bioactivity, Safety, and Toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biancarosa, I.; Belghit, I.; Bruckner, C.G.; Liland, N.S.; Waagbø, R.; Amlund, H.; Heesch, S.; Lock, E.-J.; Irene, B.; Belghit, I.; et al. Chemical characterization of 21 species of marine macroalgae common in Norwegian waters: Benefits of and limitations to their potential use in food and feed. J. Sci. Food Agric. 2018, 98, 2035–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Said, G.F.; El-Sikaily, A. Chemical composition of some seaweed from Mediterranean Sea coast, Egypt. Environ. Monit. Assess. 2013, 185, 6089–6099. [Google Scholar] [CrossRef] [Green Version]
- Roleda, M.Y.; Skjermo, J.; Marfaing, H.; Jónsdóttir, R.; Rebours, C.; Gietl, A.; Stengel, D.B.; Nitschke, U. Iodine content in bulk biomass of wild-harvested and cultivated edible seaweeds: Inherent variations determine species-specific daily allowable consumption. Food Chem. 2018, 254, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Teas, J.; Pino, S.; Critchley, A.; Braverman, L.E. Variability of Iodine Content in Common Commercially Available Edible Seaweeds. Thyroid 2004, 14, 836–841. [Google Scholar] [CrossRef]
- Romarís-Hortas, V.; García-Sartal, C.; Barciela-Alonso, M.d.C.; Domínguez-González, R.; Moreda-Piñeiro, A.; Bermejo-Barrera, P. Bioavailability study using an in vitro method of iodine and bromine in edible seaweed. Food Chem. 2011, 124, 1747–1752. [Google Scholar] [CrossRef]
- Romarís-Hortas, V.; Moreda-Piñeiro, A.; Bermejo-Barrera, P. Microwave assisted extraction of iodine and bromine from edible seaweed for inductively coupled plasma-mass spectrometry determination. Talanta 2009, 79, 947–952. [Google Scholar] [CrossRef]
- Sebastiani, G.; Barbero, A.H.; Borrás-Novel, C.; Casanova, M.A.; Aldecoa-Bilbao, V.; Andreu-Fernández, V.; Tutusaus, M.P.; Martínez, S.F.; Roig, M.D.G.; García-Algar, O. The effects of vegetarian and vegan diet during pregnancy on the health of mothers and offspring. Nutrients 2019, 11, 557. [Google Scholar] [CrossRef] [Green Version]
- Lyon, T.D.B.; Ann Robin, P.; Watson, W.S.; Littlejohn, D. Determination of elevated concentrations of bromine in serum by ICP-MS and ICP-OES. J. Anal. At. Spectrom. 2005, 20, 757–759. [Google Scholar] [CrossRef]
- Covaci, A.; Voorspoels, S.; Ramos, L.; Neels, H.; Blust, R. Recent developments in the analysis of brominated flame retardants and brominated natural compounds. J. Chromatogr. A 2007, 1153, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, K.R.R.; Kulkarni, M.G.; Stirk, W.A.; Van Staden, J. Bioactive Metabolites and Value-Added Products from Marine Macroalgae. In Seafood Processing By-Products: Trends and Applications; Springer: Berlin/Heidelberg, Germany, 2013; Chapter 21; pp. 423–440. ISBN 9781461495901. [Google Scholar]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Martin, A.; Joseph, J.A. Essential fatty acids and the brain: Possible health implications. Int. J. Dev. Neurosci. 2000, 18, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Nobre, M.E.P.; Correia, A.O.; Borges, M.d.B.; Sampaio, T.M.A.; Chakraborty, S.A.; Gonçalves, D.d.O.; Brito, G.A.d.C.; Leal, L.K.A.M.; Felipe, C.F.B.; Lucetti, D.L.; et al. Eicosapentaenoic acid and docosahexaenoic acid exert anti-inflammatory and antinociceptive effects in rodents at low doses. Nutr. Res. 2013, 33, 422–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosch, B.J.; Magnusson, M.; Paul, N.A.; de Nys, R. Total lipid and fatty acid composition of seaweeds for the selection of species for oil-based biofuel and bioproducts. GCB Bioenergy 2012, 4, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Rohani-Ghadikolaei, K.; Abdulalian, E.; Ng, W.K. Evaluation of the proximate, fatty acid and mineral composition of representative green, brown and red seaweeds from the Persian Gulf of Iran as potential food and feed resources. J. Food Sci. Technol. 2012, 49, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Al-Adilah, H.; Al-Sharrah, T.K.; Al-Bader, D.; Ebel, R.; Küpper, F.C.; Kumari, P. Assessment of arabian gulf seaweeds from kuwait as sources of nutritionally important polyunsaturated fatty acids (Pufas). Foods 2021, 10, 2442. [Google Scholar] [CrossRef]
- Kumar, M.; Kumari, P.; Trivedi, N.; Shukla, M.K.; Gupta, V.; Reddy, C.R.K.; Jha, B. Minerals, PUFAs and antioxidant properties of some tropical seaweeds from Saurashtra coast of India. J. Appl. Phycol. 2011, 23, 797–810. [Google Scholar] [CrossRef]
- Khan, M.N.A.; Cho, J.Y.; Lee, M.C.; Kang, J.Y.; Nam, G.P.; Fujii, H.; Hong, Y.K. Isolation of two anti-inflammatory and one pro-inflammatory polyunsaturated fatty acids from the brown seaweed Undaria pinnatifida. J. Agric. Food Chem. 2007, 55, 6984–6988. [Google Scholar] [CrossRef]
- Rinaudo, M. Seaweed Polysaccharides. Compr. Glycosci. Chem. Syst. Biol. 2007, 2–4, 691–735. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ammar, H.H.; Lajili, S.; Ben Said, R.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Physico-Chemical characterization and pharmacological evaluation of sulfated polysaccharides from three species of Mediterranean brown algae of the genus Cystoseira. DARU J. Pharm. Sci. 2015, 23, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Isaka, S.; Cho, K.; Nakazono, S.; Abu, R.; Ueno, M.; Kim, D.; Oda, T. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int. J. Biol. Macromol. 2015, 74, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, W.; Xiao, L.; Liu, C.; Qi, H.; Zhang, Z. In vivo antihyperlipidemic and antioxidant activity of porphyran in hyperlipidemic mice. Carbohydr. Polym. 2017, 174, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Deng, Z.; Geng, L.; Wang, J.; Zhang, Q. In vitro evaluation of the neuroprotective effect of oligo-porphyran from Porphyra yezoensis in PC12 cells. J. Appl. Phycol. 2019, 31, 2559–2571. [Google Scholar] [CrossRef]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/Akt signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Araki, C. Structure of the Agarose Constituent of Agar-agar. Bull. Chem. Soc. Jpn. 1956, 29, 543–544. [Google Scholar] [CrossRef]
- ARAKI, C. Some Recent Studies on the Polysaccharides of Agarophytes; Pergamon Press Ltd.: Oxford, UK, 1966. [Google Scholar]
- Matsuhashi, T. Agar. Chapter 1. In Food Gels; Harris, P., Ed.; Elsevier Science Publishers Ltd.: Amsterdam, The Netherlands, 1990; pp. 1–51. [Google Scholar]
- Zhong, H.; Gao, X.; Cheng, C.; Liu, C.; Wang, Q.; Han, X. The Structural Characteristics of Seaweed Polysaccharides and Their Application in Gel Drug Delivery Systems. Mar. Drugs 2020, 18, 658. [Google Scholar] [CrossRef]
- Therkelsen, G.H. Carrageenan, 3rd ed.; Academic Press Inc.: Cambridge, MA, USA, 2012; ISBN 9780127462530. [Google Scholar]
- Zia, K.M.; Tabasum, S.; Nasif, M.; Sultan, N.; Aslam, N.; Noreen, A.; Zuber, M. A review on synthesis, properties and applications of natural polymer based carrageenan blends and composites. Int. J. Biol. Macromol. 2017, 96, 282–301. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Setyawidati, N.A.R.; Puspita, M.; Kaimuddin, A.H.; Widowati, I.; Deslandes, E.; Bourgougnon, N.; Stiger-Pouvreau, V. Seasonal biomass and alginate stock assessment of three abundant genera of brown macroalgae using multispectral high resolution satellite remote sensing: A case study at Ekas Bay (Lombok, Indonesia). Mar. Pollut. Bull. 2018, 131, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Abdul Khalil, H.P.S.; Lai, T.K.; Tye, Y.Y.; Rizal, S.; Chong, E.W.N.; Yap, S.W.; Hamzah, A.A.; Nurul Fazita, M.R.; Paridah, M.T. A review of extractions of seaweed hydrocolloids: Properties and applications. Express Polym. Lett. 2018, 12, 296–317. [Google Scholar] [CrossRef]
- Alba, K.; Kontogiorgos, V. Seaweed Polysaccharides (Agar, Alginate Carrageenan); Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128140451. [Google Scholar]
- Szekalska, M.; Puciłowska, A.; Szymańska, E.; Ciosek, P.; Winnicka, K. Alginate: Current Use and Future Perspectives in Pharmaceutical and Biomedical Applications. Int. J. Polym. Sci. 2016, 2016, 7697031. [Google Scholar] [CrossRef] [Green Version]
- Rinaudo, M. Biomaterials based on a natural polysaccharide: Alginate. Tip 2014, 17, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, Y.J.; Kim, H.J.; Kim, Y.S.; Park, W. Immunostimulatory effect of laminarin on RAW 264.7 mouse macrophages. Molecules 2012, 17, 5404–5411. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.I.; Kim, H.J.; Lee, J.W. Structural feature and antioxidant activity of low molecular weight laminarin degraded by gamma irradiation. Food Chem. 2011, 129, 520–523. [Google Scholar] [CrossRef]
- Paradossi, G.; Cavalieri, F.; Pizzoferrato, L.; Liquori, A.M. A physico-chemical study on the polysaccharide ulvan from hot water extraction of the macroalga Ulva. Int. J. Biol. Macromol. 1999, 25, 309–315. [Google Scholar] [CrossRef]
- Lakshmi, D.S.; Sankaranarayanan, S.; Gajaria, T.K.; Li, G.; Kujawski, W.; Kujawa, J.; Navia, R. A short review on the valorization of green seaweeds and ulvan: Feedstock for chemicals and biomaterials. Biomolecules 2020, 10, 991. [Google Scholar] [CrossRef]
- Madub, K.; Goonoo, N.; Gimié, F.; Ait Arsa, I.; Schönherr, H.; Bhaw-Luximon, A. Green seaweeds ulvan-cellulose scaffolds enhance in vitro cell growth and in vivo angiogenesis for skin tissue engineering. Carbohydr. Polym. 2021, 251, 117025. [Google Scholar] [CrossRef]
- Alves, A.; Sousa, R.A.; Reis, R.L. In vitro cytotoxicity assessment of ulvan, a polysaccharide extracted from green algae. Phyther. Res. 2013, 27, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Kelman, D.; Posner, E.K.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, K.; Iwamura, Y.; Shibata, T.; Hirayama, I.; Nakamura, T. Bactericidal activity of phlorotannins from the brown alga Ecklonia kurome. J. Antimicrob. Chemother. 2002, 50, 889–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, A.R.; Shin, T.S.; Lee, M.S.; Park, J.Y.; Park, K.E.; Yoon, N.Y.; Kim, J.S.; Choi, J.S.; Jang, B.C.; Byun, D.S.; et al. Isolation and Identification of Phlorotannins From Ecklonia stolonifera With Antioxidant and Anti-Inflammatory Properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef]
- Artan, M.; Li, Y.; Karadeniz, F.; Lee, S.H.; Kim, M.M.; Kim, S.K. Anti-HIV-1 activity of phloroglucinol derivative, 6,6′-bieckol, from Ecklonia cava. Bioorganic Med. Chem. 2008, 16, 7921–7926. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of apoptosis by phloroglucinol derivative from Ecklonia cava in MCF-7 human breast cancer cells. Food Chem. Toxicol. 2009, 47, 1653–1658. [Google Scholar] [CrossRef]
- Parys, S.; Kehraus, S.; Krick, A.; Glombitza, K.W.; Carmeli, S.; Klimo, K.; Gerhäuser, C.; König, G.M. In vitro chemopreventive potential of fucophlorethols from the brown alga Fucus vesiculosus L. by anti-oxidant activity and inhibition of selected cytochrome P450 enzymes. Phytochemistry 2010, 71, 221–229. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Aderogba, M.A.; Ndhlala, A.R.; Stirk, W.A.; Van Staden, J. Acetylcholinesterase inhibitory activity of phlorotannins isolated from the brown alga, Ecklonia maxima (Osbeck) Papenfuss. Food Res. Int. 2013, 54, 1250–1254. [Google Scholar] [CrossRef]
- Jung, H.A.; Hyun, S.K.; Kim, H.R.; Choi, J.S. Angiotensin-converting enzyme I inhibitory activity of phlorotannins from Ecklonia stolonifera. Fish. Sci. 2006, 72, 1292–1299. [Google Scholar] [CrossRef]
- Moon, C.J.; Sung-Ho, K.; Jong-Choon, K.; Hyun, J.W.; Lee, N.H.; Park, J.W.; Shin, T. Protective Effect of Phlorotannin Components Phloroglucinol and Eckol on Radiation-induced Intestinal Injury in Mice. Phyther. Res. 2007, 22, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS ONE 2012, 7, e31145. [Google Scholar] [CrossRef]
- Rabi, T.; Bishayee, A. Terpenoids and breast cancer chemoprevention. Breast Cancer Res. Treat. 2009, 115, 223–239. [Google Scholar] [CrossRef]
- Liby, K.T.; Yore, M.M.; Sporn, M.B. Triterpenoids and rexinoids as multifunctional agents for the prevention and treatment of cancer. Nat. Rev. Cancer 2007, 7, 357–369. [Google Scholar] [CrossRef]
- Hannan, M.A.; Sohag, A.A.M.; Dash, R.; Haque, M.N.; Mohibbullah, M.; Oktaviani, D.F.; Hossain, M.T.; Choi, H.J.; Moon, I.S. Phytosterols of marine algae: Insights into the potential health benefits and molecular pharmacology. Phytomedicine 2020, 69, 153201. [Google Scholar] [CrossRef]
- Hoang, M.H.; Jia, Y.; Jun, H.J.; Lee, J.H.; Lee, B.Y.; Lee, S.J. Fucosterol is a selective liver X receptor modulator that regulates the expression of key genes in cholesterol homeostasis in macrophages, hepatocytes, and intestinal cells. J. Agric. Food Chem. 2012, 60, 11567–11575. [Google Scholar] [CrossRef]
- Sivaperumal, P.; Sankar, T.V.; Viswanathan Nair, P.G. Heavy metal concentrations in fish, shellfish and fish products from internal markets of India vis-a-vis international standards. Food Chem. 2007, 102, 612–620. [Google Scholar] [CrossRef]
- Çelik, U.; Oehlenschläger, J. High contents of cadmium, lead, zinc and copper in popular fishery products sold in Turkish supermarkets. Food Control 2007, 18, 258–261. [Google Scholar] [CrossRef]
- Paz, S.; Rubio, C.; Frías, I.; Gutiérrez, Á.J.; González-Weller, D.; Martín, V.; Revert, C.; Hardisson, A. Toxic metals (Al, Cd, Pb and Hg) in the most consumed edible seaweeds in Europe. Chemosphere 2019, 218, 879–884. [Google Scholar] [CrossRef] [PubMed]
- EU Commission Regulation (EU) No 488/2014 of 12 May 2014. Off. J. Eur. Union. 2014. Available online: https://eur-lex.europa.eu/legal-content/FR/TXT/PDF/?uri=CELEX:32014R0488&from=EN (accessed on 22 January 2022).
- Filippini, M.; Baldisserotto, A.; Menotta, S.; Fedrizzi, G.; Rubini, S.; Gigliotti, D.; Valpiani, G.; Buzzi, R.; Manfredini, S.; Vertuani, S. Heavy metals and potential risks in edible seaweed on the market in Italy. Chemosphere 2021, 263, 127983. [Google Scholar] [CrossRef] [PubMed]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Kalimuthu, S.; Kim, S.K. Chapter 7. Fucoidan, A Sulfated Polysaccharides from Brown Algae as Therapeutic Target for Cancer. In Handbook of Anticancer Drugs from Marine Origin; Springer: Cham, Switzerland, 2015; pp. 145–164. ISBN 9783319071459. [Google Scholar]
- Shibata, H.; Iimuro, M.; Uchiya, N.; Kawamori, T.; Nagaoka, M.; Ueyama, S.; Hashimoto, S.; Yokokura, T.; Sugimura, T.; Wakabayashi, K. Preventive effects of Cladosiphon fucoidan against Helicobacter pylori infection in Mongolian gerbils. Helicobacter 2003, 8, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. In vitro antioxidant and antibacterial activity of sulfated polysaccharides isolated from Spatoglossum asperum. Carbohydr. Polym. 2017, 170, 296–304. [Google Scholar] [CrossRef]
- Ayrapetyan, O.N.; Obluchinskaya, E.D.; Zhurishkina, E.V.; Skorik, Y.A.; Lebedev, D.V.; Kulminskaya, A.A.; Lapina, I.M. Antibacterial properties of fucoidans from the brown algae Fucus vesiculosus L. of the barents sea. Biology 2021, 10, 67. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Y.; Cao, M.J.; Liu, G.M.; Chen, Q.; Sun, L.; Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohydr. Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The comparative analysis of antiviral activity of native and modified fucoidans from brown algae Fucus evanescens in Vitro and in Vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- De Azevedo, T.C.G.; Bezerra, M.E.B.; Santos, M.d.G.d.L.; Souza, L.A.; Marques, C.T.; Benevides, N.M.B.; Leite, E.L. Heparinoids algal and their anticoagulant, hemorrhagic activities and platelet aggregation. Biomed. Pharmacother. 2009, 63, 477–483. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M. Pilot clinical study to evaluate the anticoagulant activity of fucoidan. Blood Coagul. Fibrinolysis 2009, 20, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structural Dependence of Sulfated Polysaccharide for Diabetes Management: Fucoidan From Undaria pinnatifida Inhibiting α-Glucosidase More Strongly Than α-Amylase and Amyloglucosidase. Front. Pharmacol. 2020, 11, 831. [Google Scholar] [CrossRef] [PubMed]
- Gahan, D.A.; Lynch, M.B.; Callan, J.J.; O’Sullivan, J.T.; O’Doherty, J.V. Performance of weanling piglets offered low-, medium- or high-lactose diets supplemented with a seaweed extract from Laminaria spp. Animal 2009, 3, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.K. Algal biotechnology industries and research activities in China. J. Appl. Phycol. 2001, 13, 375–380. [Google Scholar] [CrossRef]
- Cardozo, K.H.M.M.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.E.; Valiente, O.G.; Mainardi, V.; Bello, J.L.; Vélez, H.; Rosado, A. Isolation and characterization of an antitumor active agar-type polysaccharide of Gracilaria dominguensis. Carbohydr. Res. 1989, 190, 77–83. [Google Scholar] [CrossRef]
- Marinho-Soriano, E.; Bourret, E. Polysaccharides from the red seaweed Gracilaria dura (Gracilariales, Rhodophyta). Bioresour. Technol. 2005, 96, 379–382. [Google Scholar] [CrossRef]
- Sousa, A.M.M.; Sereno, A.M.; Hilliou, L.; Gonçalves, M.P. Biodegradable agar extracted from Gracilaria vermiculophylla: Film properties and application to edible coating. Mater. Sci. Forum 2010, 636–637, 739–744. [Google Scholar] [CrossRef]
- Pujol, C.A.; Scolaro, L.A.; Ciancia, M.; Matulewicz, M.C.; Cerezo, A.S.; Damonte, E.B. Antiviral activity of a carrageenan from Gigartina skottsbergii against intraperitoneal murine Herpes simplex virus infection. Planta Med. 2006, 72, 121–125. [Google Scholar] [CrossRef]
- Pereira, L.; Van De Velde, F. Portuguese carrageenophytes: Carrageenan composition and geographic distribution of eight species (Gigartinales, Rhodophyta). Carbohydr. Polym. 2011, 84, 614–623. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Sheng, W.; Yao, W.; Wang, C. Effect of low molecular λ-carrageenan from Chondrus ocellatus on antitumor H-22 activity of 5-Fu. Pharmacol. Res. 2006, 53, 129–134. [Google Scholar] [CrossRef]
- Ivanova, V.; Rouseva, R.; Kolarova, M.; Serkedjieva, J.; Rachev, R.; Manolova, N. Isolation of a polysaccharide with antiviral effect from Ulva lactuca. Prep. Biochem. 1994, 24, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-X.; Wijesekara, I.; Li, Y.; Kim, S.-K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, K.; Zhao, Y.; Wang, J.; Wang, B.; Qu, Y. The Embodiment of Characteristic Teaching of Marine Pharmacology-Marine Seaweed Polysaccharides. In Proceedings of the 2nd International Seminar on Education Research and Social Science (ISERSS 2019), Kuala Lumpur, Malaysia, 25–27 May 2019; Volume 322, pp. 181–183. [Google Scholar] [CrossRef]
- Murali, K.; Rajendran, V.; Ramalingam, R. Indispensability of herbal drug standardization. J. Pharmacogn. Phytochem. 2017, 6, 47–49. [Google Scholar]
- Abhishek, D.; Jyoti, P.; Savan, D.; Sumitra, C. Pharmacognostic standardization of Chaetomorpha antennina and Ulva lactuca, green seaweeds from Gujarat coast. J. Pharmacogn. Phytochem. 2018, 7, 3863–3870. [Google Scholar]
- Devi, J.A.I.; Balan, G.S.; Periyanayagam, K. Pharmacognostical study and phytochemical evaluation of brown seaweed Sargassum wightii. J. Coast. Life Med. 2013, 1, 199–204. [Google Scholar] [CrossRef]
- Devi, R.M.; Loganathan, P.; Kalaiarasi, J. Pharmacognostical and phytochemical analysis of Sargassum cinereum (Turner) C. Agardh. J. Pharmacogn. Phytochem. 2018, 7, 2233–2238. [Google Scholar]
- Shikov, A.N.; Flisyuk, E.V.; Obluchinskaya, E.D.; Pozharitskaya, O.N. Pharmacokinetics of Marine-Derived Drugs. Mar. Drugs 2020, 18, 557. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and tissue distribution of fucoidan from Fucus vesiculosus after oral administration to rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [Green Version]
- Arunkumar, M.; Mahalakshmi, M.; Ashokkumar, V.; Aravind, M.K.; Gunaseelan, S.; Mohankumar, V.; Ashokkumar, B.; Varalakshmi, P. Evaluation of seaweed sulfated polysaccharides as natural antagonists targeting Salmonella typhi OmpF: Molecular docking and pharmacokinetic profiling. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 8. [Google Scholar] [CrossRef]
- Shannon, E.; Conlon, M.; Hayes, M. Seaweed components as potential modulators of the gut microbiota. Mar. Drugs 2021, 19, 358. [Google Scholar] [CrossRef] [PubMed]
- Ventura, S.; Rodrigues, M.; Falcão, A.; Alves, G. Safety evidence on the administration of Fucus vesiculosus L. (bladderwrack) extract and lamotrigine: Data from pharmacokinetic studies in the rat. Drug Chem. Toxicol. 2020, 43, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Lionberger Topical nonsteroidal anti-inflammatory drugs for the treatment of pain due to soft tissue injury: Diclofenac epolamine topical patch. J. Pain Res. 2010, 2010, 223–233. [CrossRef] [Green Version]
- Fireman, S.; Toledano, O.; Neimann, K.; Loboda, N.; Dayan, N. A look at emerging delivery systems for topical drug products. Dermatol. Ther. 2011, 24, 477–488. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Obluchinskaya, E.D.; Vuorela, H. The pharmacokinetics of fucoidan after topical application to rats. Mar. Drugs 2019, 17, 687. [Google Scholar] [CrossRef] [Green Version]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Flisyuk, E.V.; Shikov, A.N. Formulation, optimization and in vivo evaluation of fucoidan-based cream with anti-inflammatory properties. Mar. Drugs 2021, 19, 643. [Google Scholar] [CrossRef]
- Mei, C.H.; Zhou, S.C.; Zhu, L.; Ming, J.X.; Zeng, F.D.; Xu, R. Antitumor effects of laminaria extract fucoxanthin on lung cancer. Mar. Drugs 2017, 15, 39. [Google Scholar] [CrossRef]
- Chung, T.W.; Choi, H.J.; Lee, J.Y.; Jeong, H.S.; Kim, C.H.; Joo, M.; Choi, J.Y.; Han, C.W.; Kim, S.Y.; Choi, J.S.; et al. Marine algal fucoxanthin inhibits the metastatic potential of cancer cells. Biochem. Biophys. Res. Commun. 2013, 439, 580–585. [Google Scholar] [CrossRef]
- Atya, M.E.; El-Hawiet, A.; Alyeldeen, M.A.; Ghareeb, D.A.; Abdel-Daim, M.M.; El-Sadek, M.M. In vitro biological activities and in vivo hepatoprotective role of brown algae-isolated fucoidans. Environ. Sci. Pollut. Res. 2021, 28, 19664–19676. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; White, W.; Lu, J. Extracts from New Zealand Undaria pinnatifida Containing Fucoxanthin as Potential Functional Biomaterials against Cancer in vitro. J. Funct. Biomater. 2014, 5, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Boo, H.J.; Hyun, J.H.; Kim, S.C.; Kang, J.I.; Kim, M.K.; Kim, S.Y.; Cho, H.; Yoo, E.S.; Kang, H.K. Fucoidan from Undaria pinnatifida induces apoptosis in A549 human lung carcinoma cells. Phyther. Res. 2011, 25, 1082–1086. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Ooi, V.E.C.; Chan, P.K.S.; Ang, P.O. Isolation and characterization of a sulfated polysaccharide from the brown alga Sargassum patens and determination of its anti-herpes activity. Biochem. Cell Biol. 2003, 81, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Chiu, L.C.M.; Ooi, V.E.C.; Chan, P.K.S.; Ang, P.O. Antiviral property and mode of action of a sulphated polysaccharide from Sargassum patens against Herpes simplex virus type 2. Int. J. Antimicrob. Agents 2004, 24, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Chkhikvishvili, I.D.; Ramazanov, Z.M. Phenolic substances of brown algae and their antioxidant activity. Appl. Biochem. Microbiol. 2000, 36, 289–291. [Google Scholar] [CrossRef]
- Chellappan, D.K.; Chellian, J.; Leong, J.Q.; Liaw, Y.Y. Biological and therapeutic potential of the edible brown marine seaweed Padina australis and their pharmacological mechanisms. J. Trop. Biol. Conserv. 2020, 17, 251–271. [Google Scholar]
- Custódio, L.; Silvestre, L.; Rocha, M.I.; Rodrigues, M.J.; Vizetto-Duarte, C.; Pereira, H.; Barreira, L.; Varela, J. Methanol extracts from Cystoseira tamariscifolia and Cystoseira nodicaulis are able to inhibit cholinesterases and protect a human dopaminergic cell line from hydrogen peroxide-induced cytotoxicity. Pharm. Biol. 2016, 54, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Yoon, N.Y.; Chung, H.Y.; Kim, H.R.; Choi, J.S. Acetyl- and butyrylcholinesterase inhibitory activities of sterols and phlorotannins from Ecklonia stolonifera. Fish. Sci. 2008, 74, 200–207. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-Proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef]
- Yang, H.; Zeng, M.; Dong, S.; Liu, Z.; Li, R. Anti-proliferative activity of phlorotannin extracts from brown algae Laminaria japonica Aresch. Chin. J. Oceanol. Limnol. 2010, 28, 122–130. [Google Scholar] [CrossRef]
- Gonçalves-Fernández, C.; Sineiro, J.; Moreira, R.; Gualillo, O. Extraction and characterization of phlorotannin-enriched fractions from the Atlantic seaweed Bifurcaria bifurcata and evaluation of their cytotoxic activity in murine cell line. J. Appl. Phycol. 2019, 31, 2573–2583. [Google Scholar] [CrossRef]
- Corona, G.; Coman, M.M.; Spencer, J.P.E.; Rowland, I. Digested and fermented seaweed phlorotannins reduce DNA damage and inhibit growth of HT-29 colon cancer cells. Proc. Nutr. Soc. 2014, 73, 262519. [Google Scholar] [CrossRef] [Green Version]
- Catarino, M.D.; Fernandes, I.; Oliveira, H.; Carrascal, M.; Ferreira, R.; Silva, A.M.S.; Cruz, M.T.; Mateus, N.; Cardoso, S.M. Antitumor activity of Fucus vesiculosus-derived phlorotannins through activation of apoptotic signals in gastric and colorectal tumor cell lines. Int. J. Mol. Sci. 2021, 22, 7604. [Google Scholar] [CrossRef]
- Kim, M.M.; Kim, S.K. Effect of phloroglucinol on oxidative stress and inflammation. Food Chem. Toxicol. 2010, 48, 2925–2933. [Google Scholar] [CrossRef]
- Na, H.J.; Moon, P.D.; Ko, S.G.; Lee, H.J.; Jung, H.A.; Hong, S.H.; Seo, Y.; Oh, J.M.; Lee, B.H.; Choi, B.W.; et al. Sargassum hemiphyllum inhibits atopic allergic reaction via the regulation of inflammatory mediators. J. Pharmacol. Sci. 2005, 97, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirk, W.A.; Reinecke, D.L.; Van Staden, J. Seasonal variation in antifungal, antibacterial and acetylcholinesterase activity in seven South African seaweeds. J. Appl. Phycol. 2007, 19, 271–276. [Google Scholar] [CrossRef]
- Heo, S.J.; Ko, S.C.; Kang, S.M.; Cha, S.H.; Lee, S.H.; Kang, D.H.; Jung, W.K.; Affan, A.; Oh, C.; Jeon, Y.J. Inhibitory effect of diphlorethohydroxycarmalol on melanogenesis and its protective effect against UV-B radiation-induced cell damage. Food Chem. Toxicol. 2010, 48, 1355–1361. [Google Scholar] [CrossRef]
- Heo, S.J.; Ko, S.C.; Cha, S.H.; Kang, D.H.; Park, H.S.; Choi, Y.U.; Kim, D.; Jung, W.K.; Jeon, Y.J. Effect of phlorotannins isolated from Ecklonia cava on melanogenesis and their protective effect against photo-oxidative stress induced by UV-B radiation. Toxicol. Vitr. 2009, 23, 1123–1130. [Google Scholar] [CrossRef]
- Lee, D.S.; Eom, S.H.; Jeong, S.Y.; Shin, H.J.; Je, J.Y.; Lee, E.W.; Chung, Y.H.; Kim, Y.M.; Kang, C.K.; Lee, M.S. Anti-methicillin-resistant Staphylococcus aureus (MRSA) substance from the marine bacterium Pseudomonas sp. UJ-6. Environ. Toxicol. Pharmacol. 2013, 35, 171–177. [Google Scholar] [CrossRef]
- Lee, D.S.; Kang, M.S.; Hwang, H.J.; Eom, S.H.; Yang, J.Y.; Lee, M.S.; Lee, W.J.; Jeon, Y.J.; Choi, J.S.; Kim, Y.M. Synergistic effect between dieckol from Ecklonia stolonifera and β-lactams against methicillin-resistant Staphylococcus aureus. Biotechnol. Bioprocess Eng. 2008, 13, 758–764. [Google Scholar] [CrossRef]
- Shim, S.Y.; Choi, J.S.; Byun, D.S. Inhibitory effects of phloroglucinol derivatives isolated from Ecklonia stolonifera on FcεRI expression. Bioorganic Med. Chem. 2009, 17, 4734–4739. [Google Scholar] [CrossRef]
- Myung, C.S.; Shin, H.C.; Hai, Y.B.; Soo, J.Y.; Bong, H.L.; Jong, S.K. Improvement of memory by dieckol and phlorofucofuroeckol in ethanol-treated mice: Possible involvement of the inhibition of acetylcholinesterase. Arch. Pharm. Res. 2005, 28, 691–698. [Google Scholar] [CrossRef]
- Shibata, T.; Fujimoto, K.; Nagayama, K.; Yamaguchi, K.; Nakamura, T. Inhibitory activity of brown algal phlorotannins against hyaluronidase. Int. J. Food Sci. Technol. 2002, 37, 703–709. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Eom, T.K.; Kim, M.M.; Kim, S.K. Inhibitory effect of phlorotannins isolated from Ecklonia cava on mushroom tyrosinase activity and melanin formation in mouse B16F10 melanoma cells. J. Agric. Food Chem. 2009, 57, 4124–4129. [Google Scholar] [CrossRef]
- Kim, K.N.; Ahn, G.; Heo, S.J.; Kang, S.M.; Kang, M.C.; Yang, H.M.; Kim, D.; Roh, S.W.; Kim, S.K.; Jeon, B.T.; et al. Inhibition of tumor growth in vitro and in vivo by fucoxanthin against melanoma B16F10 cells. Environ. Toxicol. Pharmacol. 2013, 35, 39–46. [Google Scholar] [CrossRef]
- Jia, R.B.; Wu, J.; Li, Z.R.; Ou, Z.R.; Lin, L.; Sun, B.; Zhao, M. Structural characterization of polysaccharides from three seaweed species and their hypoglycemic and hypolipidemic activities in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 155, 1040–1049. [Google Scholar] [CrossRef]
- Sugiura, Y.; Matsuda, K.; Okamoto, T.; Kakinuma, M.; Amano, H. Anti-allergic effects of the brown alga Eisenia arborea on Brown Norway rats. Fish. Sci. 2008, 74, 180–186. [Google Scholar] [CrossRef]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentão, P.P. Phlorotannin extracts from fucales characterized by HPLC-DAD-ESI-MS n: Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef] [Green Version]
- Matsuhiro, B.; Conte, A.F.; Damonte, E.B.; Kolender, A.A.; Matulewicz, M.C.; Mejías, E.G.; Pujol, C.A.; Zúñiga, E.A. Structural analysis and antiviral activity of a sulfated galactan from the red seaweed Schizymenia binderi (Gigartinales, Rhodophyta). Carbohydr. Res. 2005, 340, 2392–2402. [Google Scholar] [CrossRef]
- Andrade, R.M.; Maria, N.; Benevides, B.; Maria, A.; Tovar, F. In vitro inhibition of thrombin generation by sulfated polysaccharides from the marine alga Solieria filiformis (Kützing) Gabrielson (Solieriaceae, Rhodophyta). Inibição in vitro de geração de trombina por polissacarídeos sulfatados da alga marinha. Acta Fish. Aquat. Resour. 2017, 5, 2. [Google Scholar] [CrossRef]
- Ali, A.J.; Abdulla, H.I.; Al-Nimer, M.S. The Pharmacological Effects of Kappa Carrageenan on Different Human Cell Lines and Genomic DNA: An in vitro study. Iraqi J. Pharm. Sci. 2021, 30, 189–195. [Google Scholar] [CrossRef]
- Cotas, J.; Marques, V.; Afonso, M.B.; Rodrigues, C.M.P.; Pereira, L. Antitumour Potential of Gigartina pistillata Carrageenans against Colorectal Cancer Stem Cell-Enriched Tumourspheres. Mar. Drugs 2020, 18, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, M.J.; Pujol, C.A.; Ciancia, M.; Noseda, M.D.; Matulewicz, M.C.; Damonte, E.B.; Cerezo, A.S. Antiherpetic and anticoagulant properties of carrageenans from the red seaweed Gigartina skottsbergii and their cyclized derivatives: Correlation between structure and biological activity. Int. J. Biol. Macromol. 1997, 20, 97–105. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Rathgeber, B.; MacIsaac, J.; Boulianne, M.; Brigitte, L.; Stratton, G.; Thomas, N.A.; Critchley, A.T.; Hafting, J.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, reduce Salmonella enteritidis in laying hens. Front. Microbiol. 2017, 8, 567. [Google Scholar] [CrossRef] [Green Version]
- Kulshreshtha, G.; Rathgeber, B.; Stratton, G.; Thomas, N.; Evans, F.; Critchley, A.; Hafting, J.; Prithiviraj, B. Immunology, health, and disease: Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, affects performance, egg quality, and gut microbiota of layer hens. Poult. Sci. 2014, 93, 2991–3001. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Critchley, A.; Rathgeber, B.; Stratton, G.; Banskota, A.H.; Hafting, J.; Prithiviraj, B. Antimicrobial effects of selected, cultivated red seaweeds and their components in combination with tetracycline, against poultry pathogen Salmonella enteritidis. J. Mar. Sci. Eng. 2020, 8, 511. [Google Scholar] [CrossRef]
- Brabakaran, A.; Thangaraju, N. In vitro evaluation of methanolic extract of red seaweeds against α-amylase and α-glucosidase enzyme inhibitory activity. Asian J. Pharm. Pharmacol. 2018, 4, 339–342. [Google Scholar] [CrossRef]
- du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the Red Seaweed Sarconema filiforme Attenuate Symptoms of Diet-Induced Metabolic Syndrome in Rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.X.; Mi, Y.; Cao, W.X.; Lim, P.E.; Xue, C.H.; Tang, Q.J. A pilot study on anti-obesity mechanisms of Kappaphycus alvarezii: The role of native κ-carrageenan and the leftover sans-carrageenan fraction. Nutrients 2019, 11, 1133. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Zhang, Y.; Sun-Waterhouse, D.; You, L.; Fu, X. Advantages of the polysaccharides from Gracilaria lemaneiformis over metformin in antidiabetic effects on streptozotocin-induced diabetic mice. RSC Adv. 2017, 7, 9141–9151. [Google Scholar] [CrossRef] [Green Version]
- Lins, K.O.A.L.; Bezerra, D.P.; Alves, A.P.N.N.; Alencar, N.M.N.; Lima, M.W.; Torres, V.M.; Farias, W.R.L.; Pessoa, C.; De Moraes, M.O.; Costa-Lotufo, L.V. Antitumor properties of a sulfated polysaccharide from the red seaweed Champia feldmannii (Diaz-Pifferer). J. Appl. Toxicol. 2009, 29, 20–26. [Google Scholar] [CrossRef]
- Nguyen, T.; Nguyen, T.; Nguyen, V.; Nguyen, T.P.; Tran, T.A.; Do, A.; Kim, S. Antidiabetic and antioxidant activities of red seaweed Laurencia dendroidea. Asian Pac. J. Trop. Biomed. 2019, 9, 501–509. [Google Scholar] [CrossRef]
- Koehn, F.E.; Gunasekera, S.P.; Niel, D.N.; Cross, S.S. Halituna, an usual diterpene aldehyde from the marine alga Halimeda tuna. Tetrahedron Lett. 1991, 2, 169–172. [Google Scholar] [CrossRef]
- Pinto, A.M.V.; Leite, J.P.G.; Ferreira, W.J.; Cavalcanti, D.N.; Villaça, R.C.; Giongo, V.; Teixeira, V.L.; Paixão, I.C.N.d.P. Marine natural seaweed products as potential antiviral drugs against bovine viral diarrhea virus. Rev. Bras. Farmacogn. 2012, 22, 813–817. [Google Scholar] [CrossRef] [Green Version]
- Canché Chay, C.I.; Cansino, R.G.; Espitia Pinzón, C.I.; Torres-Ochoa, R.O.; Martínez, R. Synthesis and anti-tuberculosis activity of the marine natural product caulerpin and its analogues. Mar. Drugs 2014, 12, 1757–1772. [Google Scholar] [CrossRef]
- Macedo, N.R.P.V.; Ribeiro, M.S.; Villaça, R.C.; Ferreira, W.; Pinto, A.M.; Teixeira, V.L.; Cirne-Santos, C.; Paixão, I.C.N.P.; Giongo, V. Caulerpin as a potential antiviral drug against Herpes simplex virus type 1. Rev. Bras. Farmacogn. 2012, 22, 861–867. [Google Scholar] [CrossRef]
- De Souza, É.T.; De Lira, D.P.; De Queiroz, A.C.; Da Silva, D.J.C.; De Aquino, A.B.; Campessato Mella, E.A.; Lorenzo, V.P.; De Miranda, G.E.C.; De Araújo-Júnior, J.X.; De Oliveira Chaves, M.C.; et al. The antinociceptive and anti-inflammatory activities of caulerpin, a bisindole alkaloid isolated from seaweeds of the genus Caulerpa. Mar. Drugs 2009, 7, 689–704. [Google Scholar] [CrossRef] [Green Version]
- Lucena, A.M.M.; Souza, C.R.M.; Jales, J.T.; Guedes, P.M.M.; De Miranda, G.E.C.; de Moura, A.M.A.; Araújo-Júnior, J.X.; Nascimento, G.J.; Scortecci, K.C.; Santos, B.V.O.; et al. The bisindole alkaloid caulerpin, from seaweeds of the genus Caulerpa, attenuated colon damage in murine colitis model. Mar. Drugs 2018, 16, 318. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.B.; Hayashi, K.; Maeda, M.; Hayashi, T. Antiherpetic activities of sulfated polysaccharides from green algae. Planta Med. 2004, 70, 813–817. [Google Scholar] [CrossRef]
- Tanna, B.; Yadav, S.; Mishra, A. Anti-Proliferative and ROS-inhibitory activities reveal the anticancer potential of Caulerpa species. Mol. Biol. Rep. 2020, 47, 7403–7411. [Google Scholar] [CrossRef]
- Saeed, A.M.; Abotaleb, S.I.; Alam, N.G.; Elmehalawy, A.A.; Gheda, S.F. In vitro assessment of antimicrobial, antioxidant and anticancer activities of some marine macroalgae. Egypt. J. Bot. 2020, 60, 81–96. [Google Scholar] [CrossRef]
- Zbakh, H.; Salhi, G.; Bochkov, V.; Ciudad, C.J.; Noé, V.; Hassoun, M.; Riadi, H. Insights on the anti-inflammatory and antitumor activities of extracts from the marine green alga Codium decorticatum. Eur. J. Integr. Med. 2020, 37, 101170. [Google Scholar] [CrossRef]
- Ripol, A.; Cardoso, C.; Afonso, C.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Bandarra, N.M. Composition, anti-inflammatory activity, and bioaccessibility of green seaweeds from fish pond aquaculture. Nat. Prod. Commun. 2018, 13, 603–608. [Google Scholar] [CrossRef]
- Indira, K.; Balakrishnan, S.; Srinivasan, M.; Bragadeeswaran, S.; Balasubramanian, T. Evaluation of in vitro antimicrobial property of seaweed (Halimeda tuna) from Tuticorin coast, Tamil Nadu, Southeast coast of India. Afr. J. Biotechnol. 2013, 12, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.Q.; Lim, C.S.; Sung, J.Y.; Choi, H.G.; Ha, I.; Han, J.S. Ulva conglobata, a marine algae, has neuroprotective and anti-inflammatory effects in murine hippocampal and microglial cells. Neurosci. Lett. 2006, 402, 154–158. [Google Scholar] [CrossRef]
- Reis, S.E.; Andrade, R.G.C.; Accardo, C.M.; Maia, L.F.; Oliveira, L.F.C.; Nader, H.B.; Aguiar, J.A.K.; Medeiros, V.P. Influence of sulfated polysaccharides from Ulva lactuca L. upon Xa and IIa coagulation factors and on venous blood clot formation. Algal Res. 2020, 45, 101750. [Google Scholar] [CrossRef]
- Celikler, S.; Tas, S.; Vatan, O.; Ziyanok-Ayvalik, S.; Yildiz, G.; Bilaloglu, R. Anti-Hyperglycemic and antigenotoxic potential of Ulva rigida ethanolic extract in the experimental diabetes mellitus. Food Chem. Toxicol. 2009, 47, 1837–1840. [Google Scholar] [CrossRef]
- Bitencourt, M.A.O.; Silva, H.M.D.; Abílio, G.M.F.; Miranda, G.E.C.; Moura, A.M.A.; de Araújo-Júnior, J.X.; Silveira, E.J.D.; Santos, B.V.O.; Souto, J.T. Anti-Inflammatory effects of methanolic extract of green algae Caulerpa mexicana in a murine model of ulcerative colitis. Rev. Bras. Farmacogn. 2015, 25, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Bonnett, R.; Mallams, A.K.; Tee, J.L.; Weedon, B.C.L.; McCormick, A. Fucoxanthin and related pigments. Chem. Commun. 1966, 15, 515–516. [Google Scholar] [CrossRef]
- Bonnett, R.; Mallams, A.K.; Spark, A.A.; Tee, J.L.; Weedon, B.C.L. Carotenoids and Related Compounds. Part XX.l Structure and Reactions of Fucoxanthin. J. Chem. Soc. C Org. 1969, 3, 429–454. [Google Scholar] [CrossRef]
- Hwang, P.A.; Phan, N.N.; Lu, W.J.; Hieu, B.T.N.; Lin, Y.C. Low-Molecular-Weight fucoidan and high-stability fucoxanthin from brown seaweed exert prebiotics and anti-inflammatory activities in Caco-2 cells. Food Nutr. Res. 2016, 60, 32033. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Luna, A.; Ávila-Román, J.; Oliveira, H.; Motilva, V.; Talero, E. Fucoxanthin and Rosmarinic Acid Combination Has Anti-Inflammatory Effects through Regulation of NLRP3 inflammasome in UVB-exposed HaCaT keratinocytes. Mar. Drugs 2019, 17, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-A y mice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef]
- Ganesan, P.; Matsubara, K.; Sugawara, T.; Hirata, T. Marine algal carotenoids inhibit angiogenesis by down-regulating FGF-2-mediated intracellular signals in vascular endothelial cells. Mol. Cell. Biochem. 2013, 380, 1–9. [Google Scholar] [CrossRef]
- Yan, X.; Chuda, Y.; Suzuki, M.; Nagata, T. Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci. Biotechnol. Biochem. 1999, 63, 605–607. [Google Scholar] [CrossRef]
- Airanthi, M.K.W.A.; Hosokawa, M.; Miyashita, K. Comparative Antioxidant Activity of Edible Japanese Brown Seaweeds. J. Food Sci. 2011, 76, C104–C111. [Google Scholar] [CrossRef]
- Sakthivel, R.; Devi, K.P. Antioxidant, Anti-Inflammatory and Anticancer Potential of Natural Bioactive Compounds from Seaweeds, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 63, ISBN 9780128179017. [Google Scholar]
- Sinha, S.; Astani, A.; Ghosh, T.; Schnitzler, P.; Ray, B. Polysaccharides from Sargassum tenerrimum: Structural features, chemical modification and antiviral activity. Phytochemistry 2010, 71, 235–242. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.K. Neuroprotective effects of marine algae. Mar. Drugs 2011, 9, 803–818. [Google Scholar] [CrossRef]
- Andrade, P.B.; Barbosa, M.; Matos, R.P.; Lopes, G.; Vinholes, J.; Mouga, T.; Valentão, P. Valuable compounds in macroalgae extracts. Food Chem. 2013, 138, 1819–1828. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Jouini, M.; Bel Haj Amor, H.; Mzoughi, Z.; Dridi, M.; Ben Said, R.; Bouraoui, A. Phytochemical Analysis and Evaluation of the Antioxidant, Anti-Inflammatory, and Antinociceptive Potential of Phlorotannin-Rich Fractions from Three Mediterranean Brown Seaweeds. Mar. Biotechnol. 2018, 20, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Shiino, M.; Watanabe, Y.; Umezawa, K. Synthesis of N-substituted N-nitrosohydroxylamines as inhibitors of mushroom tyrosinase. Bioorg. Med. Chem. 2001, 9, 1233–1240. [Google Scholar] [CrossRef]
- Yu, S.; Blennow, A.; Bojko, M.; Madsen, F.; Olsen, C.E.; Engelsen, S.B. Physico-Chemical characterization of floridean starch of red algae. Starch/Staerke 2002, 54, 66–74. [Google Scholar] [CrossRef]
- Cian, R.E.; Drago, S.R.; De Medina, F.S.; Martínez-Augustin, O. Proteins and carbohydrates from red seaweeds: Evidence for beneficial effects on gut function and microbiota. Mar. Drugs 2015, 13, 5358–5383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudtanatip, T.; Lynch, S.A.; Wongprasert, K.; Culloty, S.C. Assessment of the effects of sulfated polysaccharides extracted from the red seaweed Irish moss Chondrus crispus on the immune-stimulant activity in mussels Mytilus spp. Fish Shellfish Immunol. 2018, 75, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.H.; Alves, A.; Popa, E.G.; Reys, L.L.; Gomes, M.E.; Sousa, R.A.; Silva, S.S.; Mano, J.F.; Reis, R.L. Marine algae sulfated polysaccharides for tissue engineering and drug delivery approaches. Biomatter 2012, 2, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Gallagher, E.; Tasdemir, D.; Hayes, M. Heart health peptides from Macroalgae and their potential use in functional foods. J. Agric. Food Chem. 2011, 59, 6829–6836. [Google Scholar] [CrossRef]
- Lascombes, C.; Agoda-Tandjawa, G.; Boulenguer, P.; Le Garnec, C.; Gilles, M.; Mauduit, S.; Barey, P.; Langendorff, V. Starch-carrageenan interactions in aqueous media: Role of each polysaccharide chemical and macromolecular characteristics. Food Hydrocoll. 2017, 66, 176–189. [Google Scholar] [CrossRef]
- Talarico, L.B.; Damonte, E.B. Interference in dengue virus adsorption and uncoating by carrageenans. Virology 2007, 363, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Khotimchenko, M.; Tiasto, V.; Kalitnik, A.; Begun, M.; Khotimchenko, R.; Leonteva, E.; Bryukhovetskiy, I.; Khotimchenko, Y. Antitumor potential of carrageenans from marine red algae. Carbohydr. Polym. 2020, 246, 116568. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Sun, Y.P.; Xin, H.; Zhang, Y.; Li, Z.; Xu, Z. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Song, J. Preparation, structural characterization and in vitro antitumor activity of kappa-carrageenan oligosaccharide fraction from Kappaphycus striatum. J. Appl. Phycol. 2005, 17, 7–13. [Google Scholar] [CrossRef]
- Silva, F.R.F.; Dore, C.M.P.G.; Marques, C.T.; Nascimento, M.S.; Benevides, N.M.B.; Rocha, H.A.O.; Chavante, S.F.; Leite, E.L. Anticoagulant activity, paw edema and pleurisy induced carrageenan: Action of major types of commercial carrageenans. Carbohydr. Polym. 2010, 79, 26–33. [Google Scholar] [CrossRef]
- Pereira, M.G.; Benevides, N.M.B.; Melo, M.R.S.; Valente, A.P.; Melo, F.R.; Mourão, P.A.S. Structure and anticoagulant activity of a sulfated galactan from the red alga, Gelidium crinale. Is there a specific structural requirement for the anticoagulant action? Carbohydr. Res. 2005, 340, 2015–2023. [Google Scholar] [CrossRef]
- Rodrigues, J.A.G.; Torres, V.M.; de Alencar, D.B.; Sampaio, A.H.; Farias, W.R.L. Extraction and anticoagulant activity of sulfated polysaccharides from the red marine alga Halymenia pseudofloresia. Acta Sci. Biol. Sci. 2009, 40, 224–231. [Google Scholar]
- Farias, W.R.L.; Valente, A.P.; Pereira, M.S.; Mourão, P.A.S. Structure and anticoagulant activity of sulfated galactans. Isolation of a unique sulfated galactan from the red algae Botryocladia occidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. J. Biol. Chem. 2000, 275, 29299–29307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.X.; Chen, X.Q.; Cheong, K.L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 151, 344–354. [Google Scholar] [CrossRef]
- Aliste, A.J.; Vieira, F.F.; Del Mastro, N.L. Radiation effects on agar, alginates and carrageenan to be used as food additives. Radiat. Phys. Chem. 2000, 57, 305–308. [Google Scholar] [CrossRef]
- Phan, T.D.; Debeaufort, F.; Luu, D.; Voilley, A. Functional properties of edible agar-based and starch-based films for food quality preservation. J. Agric. Food Chem. 2005, 53, 973–981. [Google Scholar] [CrossRef]
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambré, C.; Leblanc, J.; et al. Re-Evaluation of agar (E 406) as a food additive. EFSA J. 2016, 14, e04645. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Fu, X.; Huang, L.; Xu, J.; Gao, X. Agar oligosaccharides: A review of preparation, structures, bioactivities and application. Carbohydr. Polym. 2021, 265, 118076. [Google Scholar] [CrossRef] [PubMed]
- Agbaje-Daniels, F.; Adeleye, A.; Nwankwo, D.; Adeniyi, B.; Seku, F.; Beukes, D. Antibacterial Activities of Selected Green Seaweeds from West African Coast. Ec Pharmacol. Toxicol. 2020, 4, 84–92. [Google Scholar]
- Pramitha, V.S.; Lipton, A.P. Antimicrobial effect of Ulva fasciata Delile, 1813 solvent extracts against multidrug resistant human pathogenic bacteria and fish pathogens. Indian J. Geo-Mar. Sci. 2014, 43, 2244–2253. [Google Scholar]
- Shanmughapriya, S.; Manilal, A.; Sujith, S.; Selvin, J.; Kiran, G.S.; Natarajaseenivasan, K. Antimicrobial activity of seaweeds extracts against multiresistant pathogens. Ann. Microbiol. 2008, 58, 535–541. [Google Scholar] [CrossRef]
- Suganya, S.; Ishwarya, R.; Jayakumar, R.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-anbr, M.N.; Vaseeharan, B. New insecticides and antimicrobials derived from Sargassum wightii and Halimeda gracillis seaweeds: Toxicity against mosquito vectors and antibiofilm activity against microbial pathogens. S. Afr. J. Bot. 2019, 125, 466–480. [Google Scholar] [CrossRef]
- Pitozzi, V.; Giovannelli, L.; Bardini, G.; Rotella, C.M.; Dolara, P. Oxidative DNA damage in peripheral blood cells in type 2 diabetes mellitus: Higher vulnerability of polymorphonuclear leukocytes. Mutat. Res.—Fundam. Mol. Mech. Mutagen. 2003, 529, 129–133. [Google Scholar] [CrossRef]
- Evans, M.D.; Dizdaroglu, M.; Cooke, M.S. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res. 2004, 567, 1–61. [Google Scholar] [CrossRef]
- Gómez-Meda, B.C.; Zúñiga-González, G.M.; Zamora-Perez, A.; Luisa Ramos-Ibarra, M.; Batista-González, C.M.; Torres-Mendoza, B.M. Folate supplementation of cyclophosphamide-treated mothers diminishes micronucleated erythrocytes in peripheral blood of newborn rats. Environ. Mol. Mutagen. 2004, 44, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Bonassi, S.; Znaor, A.; Ceppi, M.; Lando, C.; Chang, W.P.; Holland, N.; Kirsch-Volders, M.; Zeiger, E.; Ban, S.; Barale, R.; et al. An increased micronucleus frequency in peripheral blood lymphocytes predicts the risk of cancer in humans. Carcinogenesis 2007, 28, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Tanna, B.; Brahmbhatt, H.R.; Mishra, A. Phenolic, flavonoid, and amino acid compositions reveal that selected tropical seaweeds have the potential to be functional food ingredients. J. Food Process. Preserv. 2019, 43, e14266. [Google Scholar] [CrossRef]
- Jeong, S.C.; Jeong, Y.T.; Lee, S.M.; Kim, J.H. Immune-Modulating activities of polysaccharides extracted from brown algae Hizikia fusiforme. Biosci. Biotechnol. Biochem. 2015, 79, 1362–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yermak, I.M.; Sokolova, E.V.; Davydova, V.N.; Solov’eva, T.F.; Aminin, D.L.; Reunov, A.V.; Lapshina, L.A. Influence of red algal polysaccharides on biological activities and supramolecular structure of bacterial lipopolysaccharide. J. Appl. Phycol. 2016, 28, 619–627. [Google Scholar] [CrossRef]
- Wang, B.; Huang, H.; Xiong, H.P.; Xie, E.Y.; Li, Z.M. Analysis on nutrition constituents of Sargassum naozhouense sp. Food Res. Dev. 2010, 31, 195–197. [Google Scholar]
- Hong, D.D.; Hien, H.M.; Son, P.N. Seaweeds from Vietnam used for functional food, medicine and biofertilizer. J. Appl. Phycol. 2007, 19, 817–826. [Google Scholar] [CrossRef]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef]
- Uzuner, H.; Bauer, R.; Fan, T.P.; Guo, D.A.; Dias, A.; El-Nezami, H.; Efferth, T.; Williamson, E.M.; Heinrich, M.; Robinson, N.; et al. Traditional Chinese medicine research in the post-genomic era: Good practice, priorities, challenges and opportunities. J. Ethnopharmacol. 2012, 140, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Editorial Board of Zhong Hua Ben Cao (China Herbal). Zhonghua Ben Cao; Shanghai Science and Technology Publishing House: Shanghai, China, 1999. [Google Scholar]
- Song, X.H.; Zan, R.Z.; Yu, C.H.; Wang, F. Effects of modified Haizao Yuhu Decoction in experimental autoimmune thyroiditis rats. J. Ethnopharmacol. 2011, 135, 321–324. [Google Scholar] [CrossRef]
- Smith, E.L.; Abbott, A.P.; Ryder, K.S. Deep Eutectic Solvents (DESs) and Their Applications. Chem. Rev. 2014, 114, 11060–11082. [Google Scholar] [CrossRef] [Green Version]
- Zdanowicz, M.; Wilpiszewska, K.; Spychaj, T. Deep eutectic solvents for polysaccharides processing. A review. Carbohydr. Polym. 2018, 200, 361–380. [Google Scholar] [CrossRef]
- Saravana, P.S.; Cho, Y.N.; Woo, H.C.; Chun, B.S. Green and efficient extraction of polysaccharides from brown seaweed by adding deep eutectic solvent in subcritical water hydrolysis. J. Clean. Prod. 2018, 198, 1474–1484. [Google Scholar] [CrossRef]
- Nie, J.; Chen, D.; Lu, Y. Deep eutectic solvents based ultrasonic extraction of polysaccharides from edible brown Seaweed Sargassum horneri. J. Mar. Sci. Eng. 2020, 8, 440. [Google Scholar] [CrossRef]
- Li, G.; Row, K.H. Air Assisted Dispersive Liquid–Liquid Microextraction (AA-DLLME) Using Hydrophilic–Hydrophobic Deep Eutectic Solvents for the Isolation of Monosaccharides and Amino Acids from Kelp. Anal. Lett. 2020, 53, 188–202. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Zakharova, L.V.; Daurtseva, A.V.; Flisyuk, E.V.; Shikov, A.N. Efficacy of natural deep eutectic solvents for extraction of hydrophilic and lipophilic compounds from Fucus vesiculosus. Molecules 2021, 26, 4198. [Google Scholar] [CrossRef]
- Das, A.K.; Sharma, M.; Mondal, D.; Prasad, K. Deep eutectic solvents as efficient solvent system for the extraction of κ-carrageenan from Kappaphycus alvarezii. Carbohydr. Polym. 2016, 136, 930–935. [Google Scholar] [CrossRef]
- World Health Organization. World Health Organization Countries at the Core. WHO Medicines Strategy 2004–2007. World Health Organisation Geneva. 2007. Available online: http//www.who.int/management/background_4a.pdf (accessed on 14 January 2022).
- World Health Organization. WHO Traditional Medicine Strategy 2002–2005. World Health Organization, Geneva. 2005. Available online: http//www.wpro.who.int/health_technology/book_who_traditional_medicine_strategy_2002_2005.pdf (accessed on 1 January 2022).
- Nwobike, J.C. Pharmaceutical corporations and access to drugs indeveloping countries: The way forward. Int. J. Hum. Rights 2006, 4, 126–143. [Google Scholar]







| Species | Seaweed Compound | Pre-Clinical Study | Pharmaceutical Property | Reference |
|---|---|---|---|---|
| Phylum Ochrophyta, Class Phaeophyceae | ||||
| Laminaria japonica | Fucoxanthins | In vitro | Antitumoral activity on lung cancer cells | [140,141] |
| Colpomenia sinuosa, Sargassum prismaticum | Antitumoral activity on HCT-116, MCF-7, HepG-2 cells | [142] | ||
| Undaria pinnatifida | Antitumoral activity on Malme-3M, SiHa cells | [143] | ||
| Undaria pinnatifida | Fucoidans | Antitumoral activity on A549 cells | [144] | |
| Antidiabetic activity | [114] | |||
| Cladosiphon okamuranus | Antimicrobial activity against Helicobacter pylori | [106] | ||
| Spatoglossum asperum | Antimicrobial activity against Aeromonas hydrophila | [107] | ||
| Fucus vesiculosus | Antimicrobial activity against Escherichia coli, Staphylococcus epidermidis, S. aureus, Bacillus licheniformis | [108] | ||
| Anticoagulant activity | [111,112] | |||
| Laminaria japonica | Antimicrobial activity against Escherichia coli, Staphylococcus aureus | [109] | ||
| Fucus evanescens | Antiviral activity against HSV-1, HSV-2, ECHO-1, HIV-1 | [110] | ||
| Sargassum patens | Sulphate PSs | Antiviral activity against HSV-1, HVS-2 | [145,146] | |
| Padina australis | Phenols | Antimicrobial activity against Bacillus cereus, Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus | [147,148] | |
| Cystoseira tamariscifolia, C. nodicaulis | AChE-inhibitory activity | [149] | ||
| Ecklonia maxima, E. stolonifera, Ishige okamurae | AChE-inhibitory activity | [91,150] | ||
| Fucus vesiculosus, Alaria esculenta, Ascophyllum nodosum, Laminaria, japonica, Sargassum muticum, Bifurcaria bifurcata | Phlorotannins | Antitumoral activity on HFF-1, MKN-28, HT-29, Caco-2, BEL-7402, P388, ATDC5 cells | [151,152,153,154,155,156] | |
| Ecklonia cava | Anti-inflammatory activity | [157] | ||
| Sargassum hemiphylum | Treatment of atopic dermatitis | [158] | ||
| Dictyota humifusa | AChE-inhibitory activity | [159] | ||
| Sargassum hemiphylum | Treatment of atopic dermatitis | [158] | ||
| Ecklonia cava, E. stolonifera | UV photoprotection | [160,161] | ||
| Ecklonia kurome | Phlorotannins | Antimicrobial activity against Staphylococcus aureus, S. pyogenes, Bacillus cereus, Campylobacter fetus, C. jejuni, Escherichia coli, Salmonella enteritidis, S. typhimurium, Vibrio parahaemolyticus | [86] | |
| Ecklonia stolonifera | Phlorotannins (dieckol) | Antimicrobial activity against Staphylococcus aureus | [162,163] | |
| Phlorotannins (DHE, PFF-A) | Antiallergic effects | [164] | ||
| Eisenia sp., Ecklonia sp. | Phlorotannins (dieckol, PFFA) | AChE-inhibitory activity | [165] | |
| Eisenia bicyclis, Ecklonia kurome | Phlorotannins (PFFA, dieckol, eckol, bieckol) | Hyaluronidase-inhibition activity | [166] | |
| Ecklonia cava | Phlorotannins (7-phloroeckol, dieckol) | UV photoprotection in B16F10 melanoma cells | [167] | |
| Laminaria japonica | Fucoxanthins | In vivo | Antitumoral activity on lung cancer cells | [140,141] |
| Ishige okamurae | Antitumoral activity on B16-F10 cells | [168] | ||
| Colpomenia sinuosa, Sargassum prismaticum | Antioxidant activity and hepatoprotective ability | [142] | ||
| Fucus vesiculosus | Fucoidans | Anticoagulant activity | [113] | |
| Fucus evanescens | Antiviral activity against HSV-1, HSV-2, ECHO-1, HIV-1 | [110] | ||
| Sargassum fusiforme, Macrocystis pyrifera | Sulphate PSs | Antidiabetic activity | [169] | |
| Ecklonia cava | Phlorotannins | Antiproliferative activity on MCF-7 cells | [89] | |
| Eisenia arborea | Antiallergic effects | [170] | ||
| Cystoseira nodicaulis | Hyaluronidase-inhibition activity | [171] | ||
| Phylum Rhodophyta | ||||
| Schizymenia binderi | Carrageenans | In vitro | Antiviral activity against HSV-1, HSV-2 | [172] |
| Solieria filiformis | Anticoagulant activity | [173] | ||
| Gigartina sp., Chondrus sp., Eucheuma sp., Iridaea sp. | κ-carrageenans | Antiproliferative activity on HeLa cells, mammary cells fibroblasts | [174] | |
| Gigartina pistillata | ɩ/ε-carrageenans | Antitumoral activity on colorectal cancer stem cells | [175] | |
| Gigartina skottsbergii | ɩ- and k-carrageenans | Antiviral activity against HSV-1, HSV-2 | [121,176] | |
| Chondrus crispus, Sarcodiotheca gaudichaudii | Polysaccharides | Antimicrobial activity against Salmonella Enteritidis | [177,178,179] | |
| Pterocladia capillacea, Laurencia obtusa | Sulphate PSs | Antiviral activity against HCV | [10] | |
| Spyridia filamentosa | Methanolic extract | Antidiabetic activity | [180] | |
| Chondrus ocellatus | ʎ-carrageenans | In vivo | Antitumoral activity on H-22 cells | [123] |
| Sarconema filiforme | ι-carrageenans | Anti-obesity activity | [181] | |
| Kappaphycus alvarezii | k-carrageenans | Anti-obesity activity | [182] | |
| Gracilaria dominguensis | Agar-type PSs | Anticancer activity on EAC cells | [118] | |
| Chondrus crispus, Sarcodiotheca gaudichaudii | Polysaccharides | Antimicrobial activity against Salmonella Enteritidis | [177,178,179] | |
| Gracilaria lemaneiformis | Sulphate PSs | Anti-obesity, antidiabetic activities | [183] | |
| Champia parvula | Antitumoral activity on sarcoma 180 ascites cells | [184] | ||
| Laurencia dendroidea | Ethyl-acetate extract | Antidiabetic activity | [185] | |
| Phylum Chlorophyta | ||||
| Ulva lactuca | Ulvan | In vitro | Antitumoral activity on L929 cells | [81] |
| Antiviral activity against IAV | [124] | |||
| Halimeda tuna | Dipertene | Antiviral activity against coronavirus strain A5Y | [186] | |
| Caulerpa sp. | Caulerpin | Antiviral activity against BVDV | [187] | |
| Antimicrobial activity against Mycobacterium tuberculosis | [188] | |||
| Caulerpa racemosa | Antiviral activity against HSV-1 | [189] | ||
| Anti-inflammatory activity | [190,191] | |||
| Caulerpa brachypus | Antiviral activity against HSV-1 | [192] | ||
| Caulerpa racemosa, C. scalpelliformis | Phenols | Antiproliferative activity on Huh-7, HeLa cells | [193] | |
| Ulvs fasciata | Antiproliferative activity on PC3, HepG2 cells | [194] | ||
| Ulva lactuca | Antiproliferative activity on MCF-7, HeLa cells | [194] | ||
| Ulva lactuca, U. fasciata | Antimicrobial activity against Klebsiella pneumoniae, Proteus mirabilis; antifungal activity against Aspergillus flavus, A. s fumingatus, A. niger | [194] | ||
| Codium decorticatum | Extracts | Anti-inflammatory, anticancer activities | [195] | |
| Chaetomorpha linum, Rhizoclonium riparium, Ulva intestinalis, U. lactuca, U. prolifera | Anti-inflammatory activity | [196] | ||
| Halimeda tuna | Methanolic extracts | Antimicrobial activity against Staphylococcus aureus, Salmonella typhimurium, S. paratyphi, Klebsiella oxytoca, Escherichia coli; antifungal activity against Aspergillus niger, A. flavus, Alternaria alternaria, Candida albicans, Epidermophyton floccossum, | [197] | |
| Ulva conglobata | Antioxidant activity | [198] | ||
| Caulerpa racemosa | Caulerpin | In vivo | Anti-inflammatory activity | [190,191] |
| Ulva lactuca | Sulphated PSs | Anticoagulant activity | [199] | |
| Ulva rigida | Ethanolic extracts | Anti-hyperglycaemic activity | [200] | |
| Caulerpa mexicana | Methanolic extracts | Anti-inflammatory activity | [201] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lomartire, S.; Gonçalves, A.M.M. An Overview of Potential Seaweed-Derived Bioactive Compounds for Pharmaceutical Applications. Mar. Drugs 2022, 20, 141. https://doi.org/10.3390/md20020141
Lomartire S, Gonçalves AMM. An Overview of Potential Seaweed-Derived Bioactive Compounds for Pharmaceutical Applications. Marine Drugs. 2022; 20(2):141. https://doi.org/10.3390/md20020141
Chicago/Turabian StyleLomartire, Silvia, and Ana M. M. Gonçalves. 2022. "An Overview of Potential Seaweed-Derived Bioactive Compounds for Pharmaceutical Applications" Marine Drugs 20, no. 2: 141. https://doi.org/10.3390/md20020141
APA StyleLomartire, S., & Gonçalves, A. M. M. (2022). An Overview of Potential Seaweed-Derived Bioactive Compounds for Pharmaceutical Applications. Marine Drugs, 20(2), 141. https://doi.org/10.3390/md20020141