
Anti-Inflammatory Polyketides from an Alga-Derived Fungus Aspergillus ochraceopetaliformis SCSIO 41020

 , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
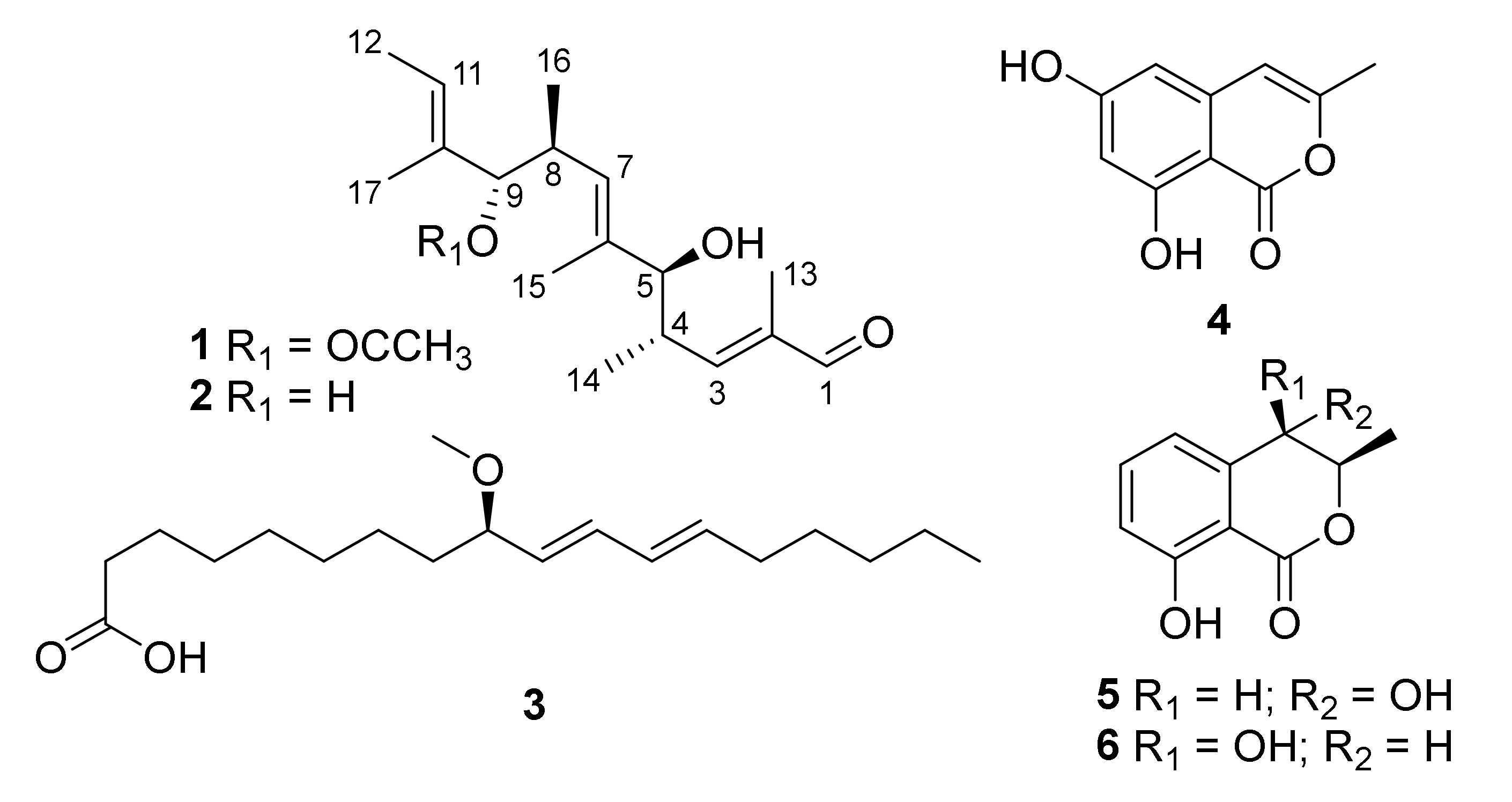
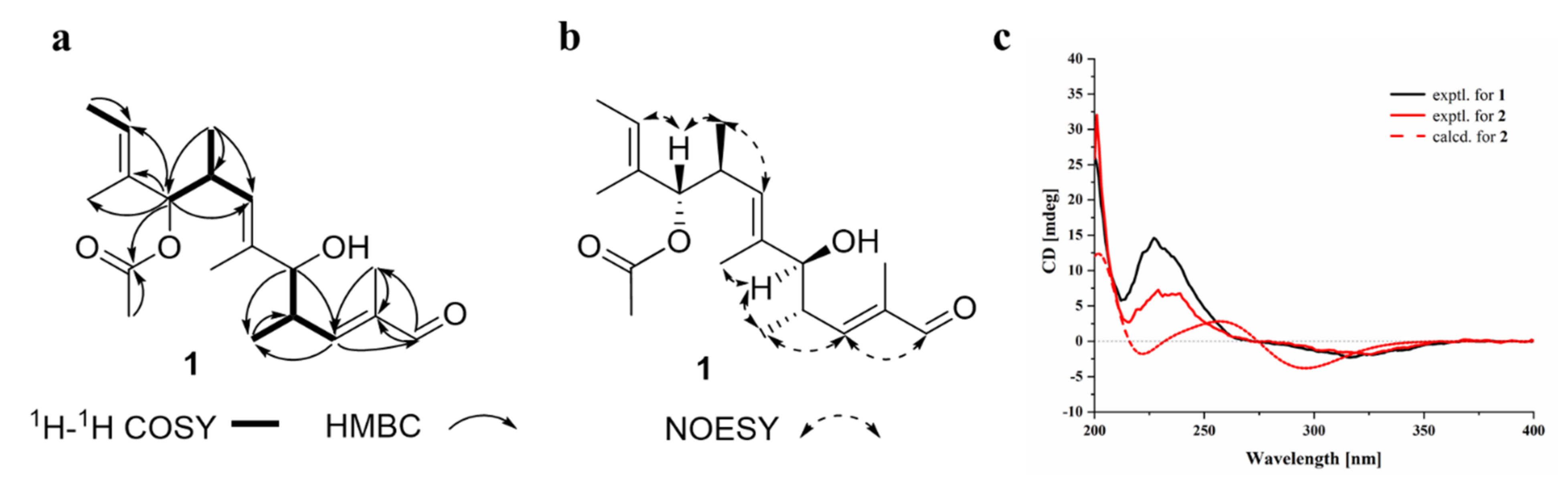
2.1. Structural Determination
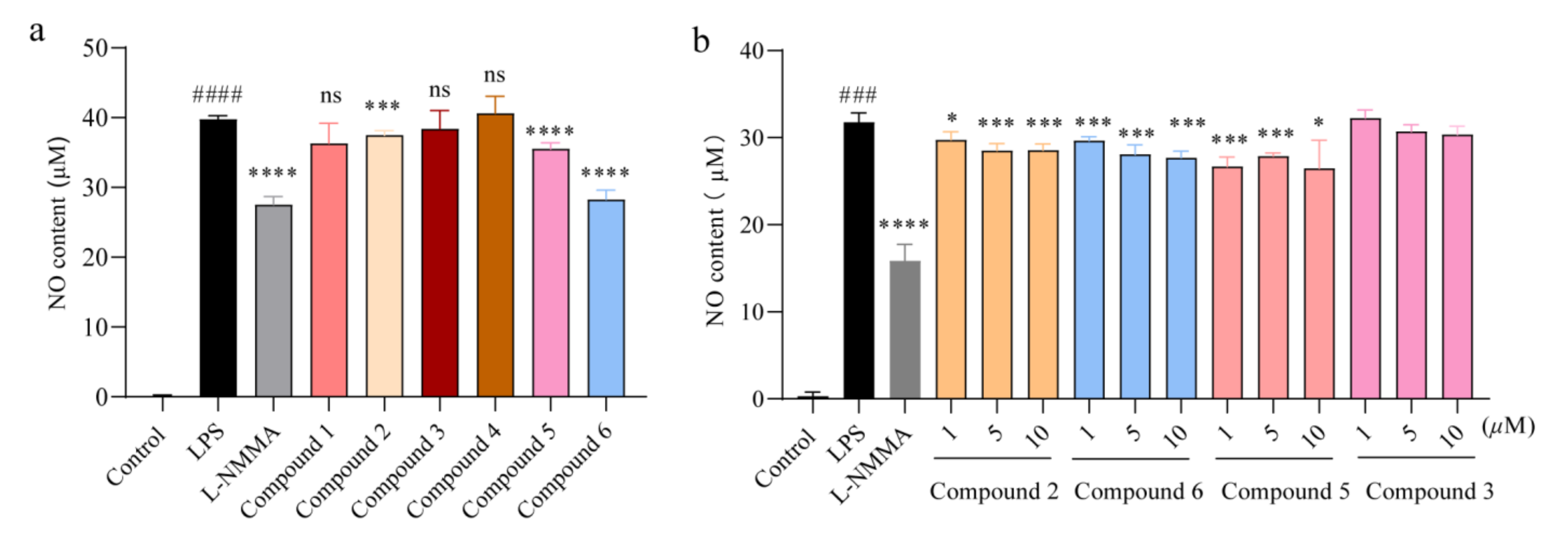
2.2. Compounds 2, 5, and 6 Inhibited the Overproduction of Nitric Oxide in a Dose-Dependent Manner
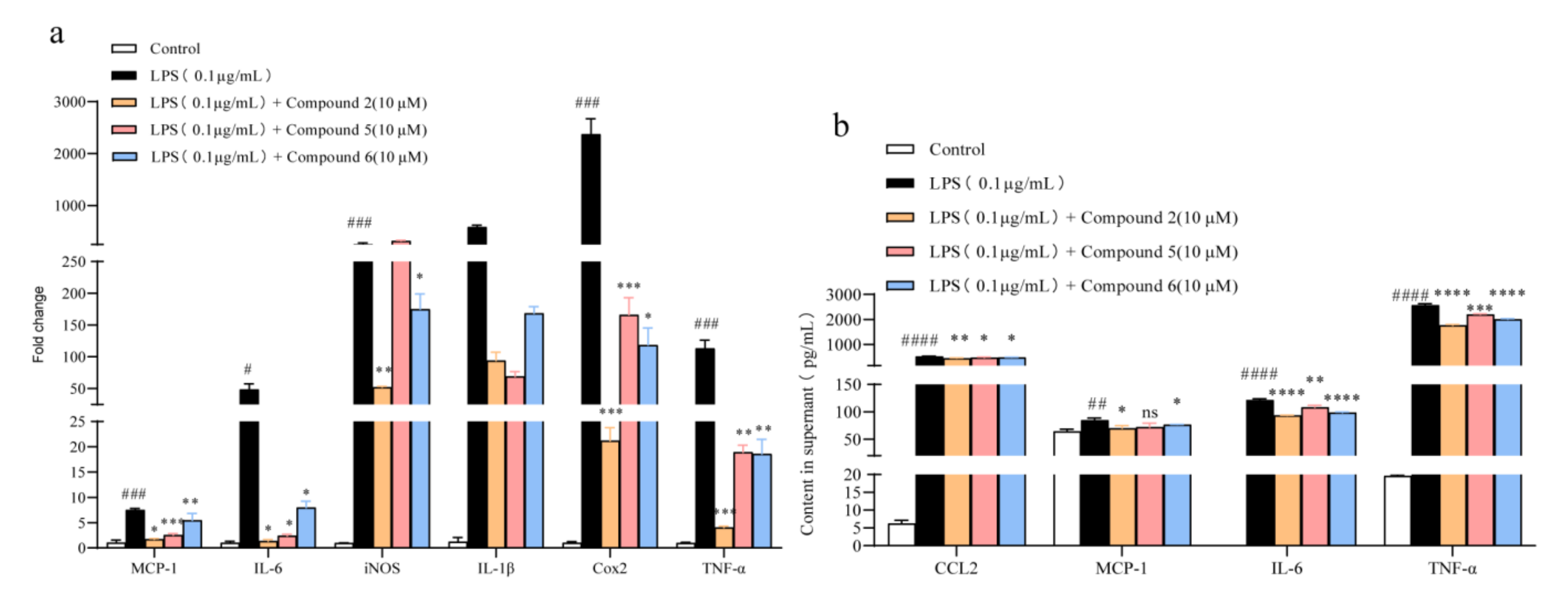
2.3. Compounds 2, 5, and 6 Repressed the Secretion of Pro-Inflammatory Cytokines in Raw 264.7 Macrophages
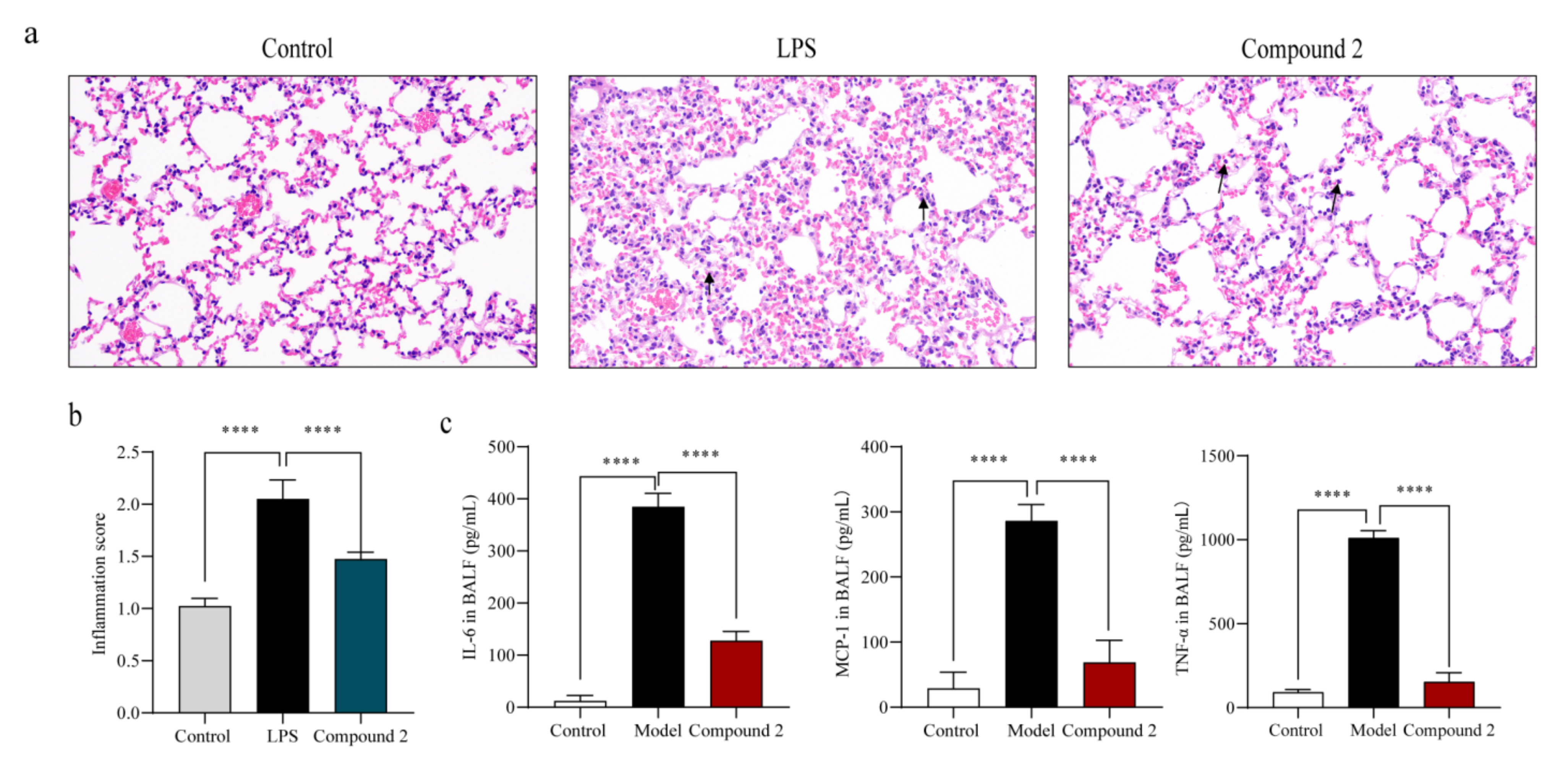
2.4. Compound 2 Attenuated the Lung Injury in LPS-Treated Mice
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fungal Material
3.3. Fermentation and Extraction
3.4. Isolation and Purification
3.5. ECD Calculation of Compound 2
3.6. Spectroscopic Data of Compound 1
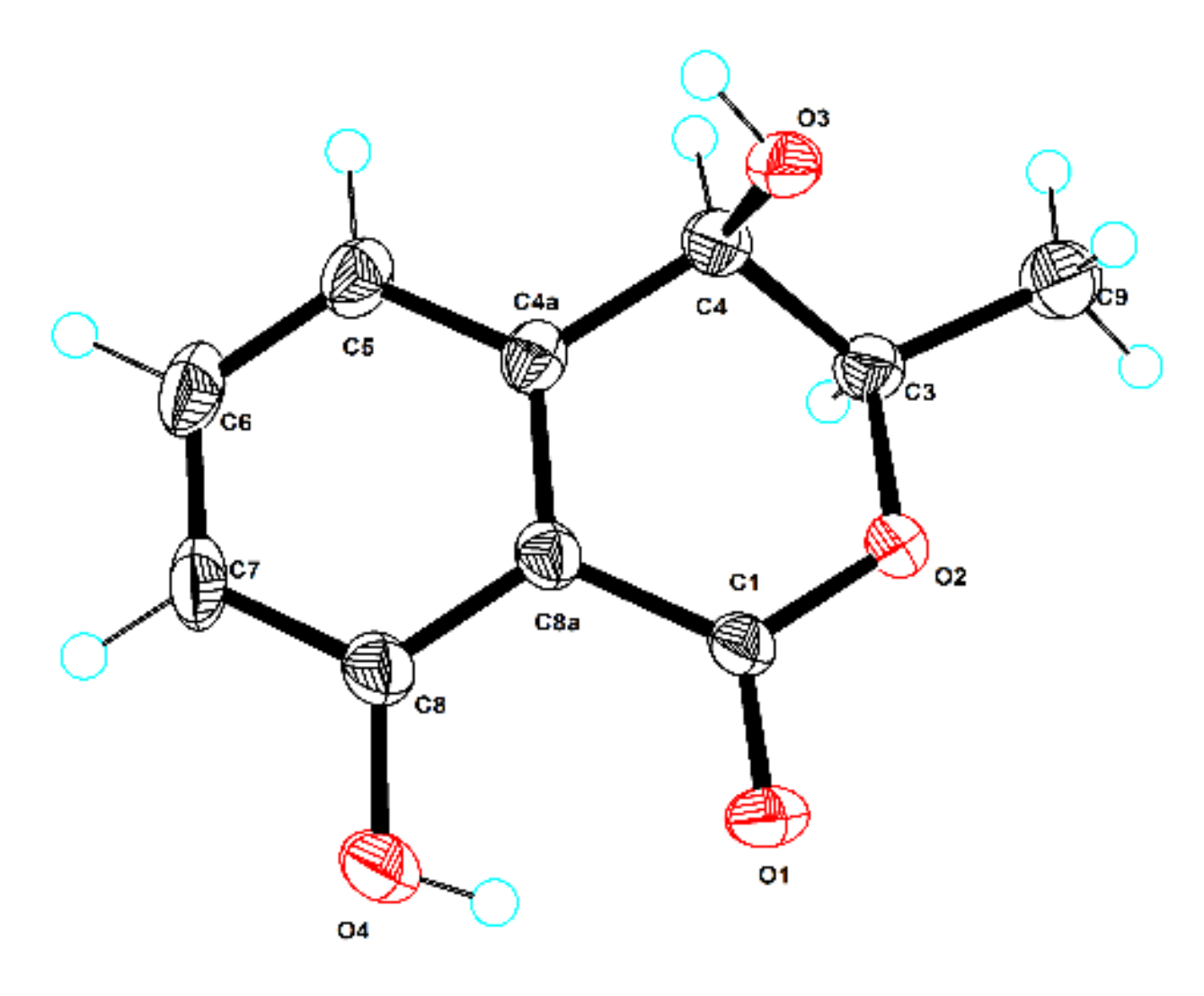
3.7. X-ray Crystallographic Analysis
3.8. LPS-Induced RAW264.7 Macrophages Inflammation Model
3.9. CCK-8 Assay
3.10. Nitric Oxide Production Assay
3.11. Cytokines Detection
3.12. Gene Expression Analysis
3.13. Animal Experimental Protocol
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Al-Samkari, H.; Berliner, N. Hemophagocytic lymphohistiocytosis. Annu. Rev. Pathol-Mech. 2018, 13, 27–49. [Google Scholar] [CrossRef]
- Barton, E.B. Molecule of the month IL-6: Insights into novel biological activities. Clin. Immunol. Immunopathol. 1997, 85, 16–20. [Google Scholar] [CrossRef]
- Barton, B.; Shortall, J.; Jackson, J. Interleukins 6 and 11 protect mice from mortality in a staphylococcal enterotoxin-induced toxic shock model. Infect. Immun. 1996, 64, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Opal, S.M.; DePalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. CSH Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Kang, S.; Tanaka, T.; Narazaki, M.; Kishimoto, T. Targeting interleukin-6 signaling in clinic. Immunity 2019, 50, 1007–1023. [Google Scholar] [CrossRef]
- Lin, S.; Zhou, Z.; Zhao, H.; Xu, C.; Guo, Y.; Gao, S.; Mei, X.; Tian, H. TNF promotes M1 polarization through mitochondrial metabolism in injured spinal cord. Free Radical. Biol. Med. 2021, 172, 622–632. [Google Scholar] [CrossRef]
- Wang, B.G.; Gloer, J.B.; Ji, N.Y.; Zhao, J.C. Halogenated organic molecules of Rhodomelaceae origin: Chemistry and biology. Chem. Rev. 2013, 113, 3632–3685. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2022, 38. [Google Scholar] [CrossRef]
- Orfali, R.; Aboseada, M.A.; Abdel-Wahab, N.M.; Hassan, H.M.; Perveen, S.; Ameen, F.; Alturki, E.; Abdelmohsen, U.R. Recent updates on the bioactive compounds of the marine-derived genus Aspergillus. RSC Adv. 2021, 11, 17116–17150. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Larsen, T.O. Chemodiversity in the genus Aspergillus. Appl. Microbiol. Biotechnol. 2015, 99, 7859–7877. [Google Scholar] [CrossRef]
- Cai, J.; Chen, C.; Tan, Y.; Chen, W.; Luo, X.; Luo, L.; Yang, B.; Liu, Y.; Zhou, X. Bioactive Polyketide and diketopiperazine derivatives from the mangrove-sediment-derived fungus Aspergillus sp. SCSIO41407. Molecules 2021, 26, 4851. [Google Scholar] [CrossRef]
- Chen, C.M.; Chen, W.H.; Pang, X.Y.; Liao, S.R.; Wang, J.F.; Lin, X.P.; Yang, B.; Zhou, X.F.; Luo, X.W.; Liu, Y.H. Pyrrolyl 4-quinolone alkaloids from the mangrove endophytic fungus Penicillium steckii SCSIO 41025: Chiral resolution, configurational assignment, and enzyme inhibitory activities. Phytochemistry 2021, 186, 112730. [Google Scholar] [CrossRef]
- Tawfike, A.F.; Romli, M.; Clements, C.; Abbott, G.; Young, L.; Schumacher, M.; Diederich, M.; Farag, M.; Edrada-Ebel, R. Isolation of anticancer and anti-trypanosome secondary metabolites from the endophytic fungus Aspergillus flocculus via bioactivity guided isolation and MS based metabolomics. J. Chromatogr. B 2019, 1106–1107, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Matsumori, N.; Kaneno, D.; Murata, M.; Nakamura, H.; Tachibana, K. Stereochemical determination of acyclic structures based on carbon-proton spin-coupling constants. A Method of Configuration Analysis for Natural Products. J. Org. Chem. 1999, 64, 866–876. [Google Scholar] [CrossRef]
- Wang, H.; Qi, H.; Zhang, S.Y.; Song, W.S.; Zhang, L.Q.; Xiang, W.S.; Wang, J.D. Sarubicinols A-C, cytotoxic benzoxazoles from a Streptomyces. J. Nat. Prod. 2022. [Google Scholar] [CrossRef]
- Kohno, J.; Nishio, M.; Sakurai, M.; Kawano, K.; Hiramatsu, H.; Kameda, N.; Kishi, N.; Yamashita, T.; Okuda, T.; Komatsubara, S. Isolation and structure determination of TMC-151s: Novel polyketide antibiotics from Gliocladium catenulatum Gilman & Abbott TC 1280. Tetrahedron 1999, 55, 7771–7786. [Google Scholar]
- Lin, S.; Zhang, Y.L.; Liu, M.T.; Zi, J.C.; Gan, M.L.; Song, W.X.; Fan, X.N.; Wang, S.J.; Yang, Y.C.; Shi, J.G. Methoxylated fatty acids from the bark of Fraxinus sieboldiana. J. Asian Nat. Prod. Res. 2012, 14, 235–243. [Google Scholar] [CrossRef]
- Singh, B.; Parshad, R.; Khajuria, R.K.; Guru, S.K.; Pathania, A.S.; Sharma, R.; Chib, R.; Aravinda, S.; Gupta, V.K.; Khan, I.A.; et al. Saccharonol B, a new cytotoxic methylated isocoumarin from Saccharomonospora azurea. Tetrahedron Lett. 2013, 54, 6695–6699. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Lin, X.P.; Liu, J.; Kaliyaperumal, K.; Ai, W.; Ju, Z.R.; Yang, B.; Wang, J.; Yang, X.W.; Liu, Y. Ascomycotin A, a new citromycetin analogue produced by Ascomycota sp. Ind19F07 isolated from deep sea sediment. Nat. Prod. Res. 2015, 29, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Chávez, J.; Figueroa, M.; González, M.d.C.; Glenn, A.E.; Mata, R. α-Glucosidase inhibitors from a Xylaria feejeensis associated with Hintonia latiflora. J. Nat. Prod. 2015, 78, 730–735. [Google Scholar] [CrossRef]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Luo, X.W.; Lin, X.P.; Tao, H.M.; Wang, J.F.; Li, J.Y.; Yang, B.; Zhou, X.F.; Liu, Y.H. Isochromophilones A–F, cytotoxic chloroazaphilones from the marine mangrove endophytic fungus Diaporthe sp. SCSIO 41011. J. Nat. Prod. 2018, 81, 934–941. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pos. | δC Type | δH (J in Hz) | HMBC | 1H-1H COSY | ||
|---|---|---|---|---|---|---|
| 1 | 195.4, CH | 9.33 (s) | C-2, C-13 | |||
| 2 | 138.0, C | |||||
| 3 | 158.6, CH | 6.56 (dd, 9.5, 1.0) | C-1, C-13 | H-4 | ||
| 4 | 37.3, CH | 2.77–2.82 (m) | C-3, C-14 | H-3, H-5, H3-14 | ||
| 5 | 78.9, CH | 3.74 (d, 7.0) | C-3, C-4, C-6, C-7, C-14, C-15 | H-4 | ||
| 6 | 137.2, C | |||||
| 7 | 127.9, CH | 5.15 (d, 9.5) | C-5, C-15 | H-8 | ||
| 8 | 33.7, CH | 2.69–2.74 (m) | 2.69–2.74 (m) | 2.69–2.74 (m) | C-7, C-9, C-16 | H-7, H-9, H3-16 |
| 9 | 82.3, CH | 4.87 (d, 8.5) | C-7, C-8, C-10, C-11, C-17, C-18 | H-8 | ||
| 10 | 132.8, C | |||||
| 11 | 123.3, CH | 5.43 (q, 5.5) | C-9, C-12 | H3-12 | ||
| 12 | 12.2, CH3 | 1.56 (s) | C-11 | H-11 | ||
| 13 | 9.1, CH3 | 1.65 (d, 1.0) | C-1, C-3, C-4 | |||
| 14 | 16.7, CH3 | 0.89 (d, 7.0) | C-3, C-4, C-5 | H-4 | ||
| 15 | 12.8, CH3 | 1.57 (d, 1.0) | C-5, C-6, C-7 | |||
| 16 | 17.3, CH3 | 0.78 (d, 7.0) | C-7, C-8, C-9 | H-8 | ||
| 17 | 11.4, CH3 | 1.55 (s) | C-9 | |||
| 18 | 169.4, C | |||||
| 19 | 20.9, CH3 | 1.88 (s) | C-18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Ren, X.; Tao, H.; Cai, W.; Chen, Y.; Luo, X.; Guo, P.; Liu, Y. Anti-Inflammatory Polyketides from an Alga-Derived Fungus Aspergillus ochraceopetaliformis SCSIO 41020. Mar. Drugs 2022, 20, 295. https://doi.org/10.3390/md20050295
Chen C, Ren X, Tao H, Cai W, Chen Y, Luo X, Guo P, Liu Y. Anti-Inflammatory Polyketides from an Alga-Derived Fungus Aspergillus ochraceopetaliformis SCSIO 41020. Marine Drugs. 2022; 20(5):295. https://doi.org/10.3390/md20050295
Chicago/Turabian StyleChen, Chunmei, Xue Ren, Huaming Tao, Wenteng Cai, Yuchi Chen, Xiaowei Luo, Peng Guo, and Yonghong Liu. 2022. "Anti-Inflammatory Polyketides from an Alga-Derived Fungus Aspergillus ochraceopetaliformis SCSIO 41020" Marine Drugs 20, no. 5: 295. https://doi.org/10.3390/md20050295
APA StyleChen, C., Ren, X., Tao, H., Cai, W., Chen, Y., Luo, X., Guo, P., & Liu, Y. (2022). Anti-Inflammatory Polyketides from an Alga-Derived Fungus Aspergillus ochraceopetaliformis SCSIO 41020. Marine Drugs, 20(5), 295. https://doi.org/10.3390/md20050295