
Scale-Up to Pilot of a Non-Axenic Culture of Thraustochytrids Using Digestate from Methanization as Nitrogen Source

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
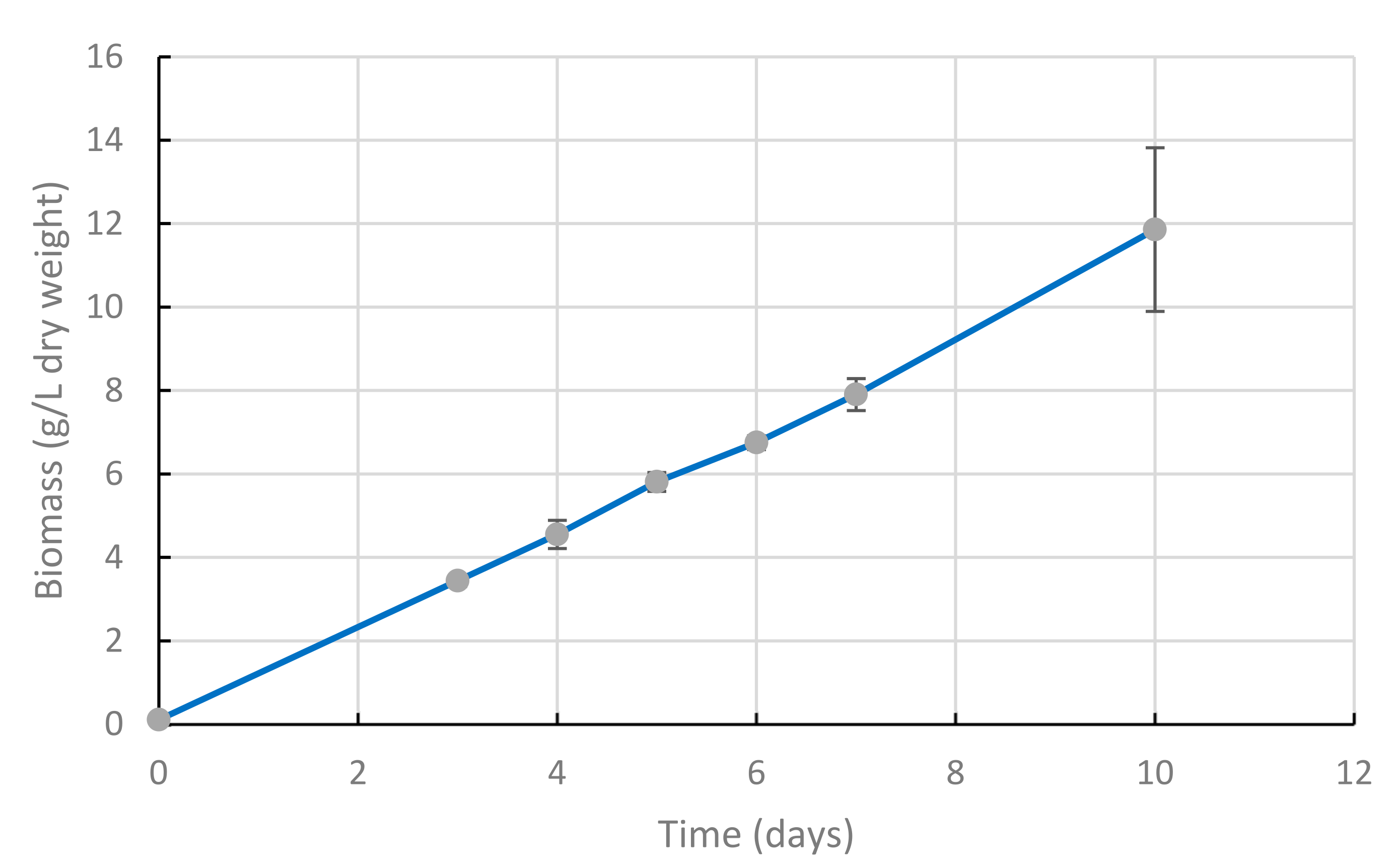
2.1. Cultures Submitted to Air Bubbling and Shaking Exhibit Linear Growth
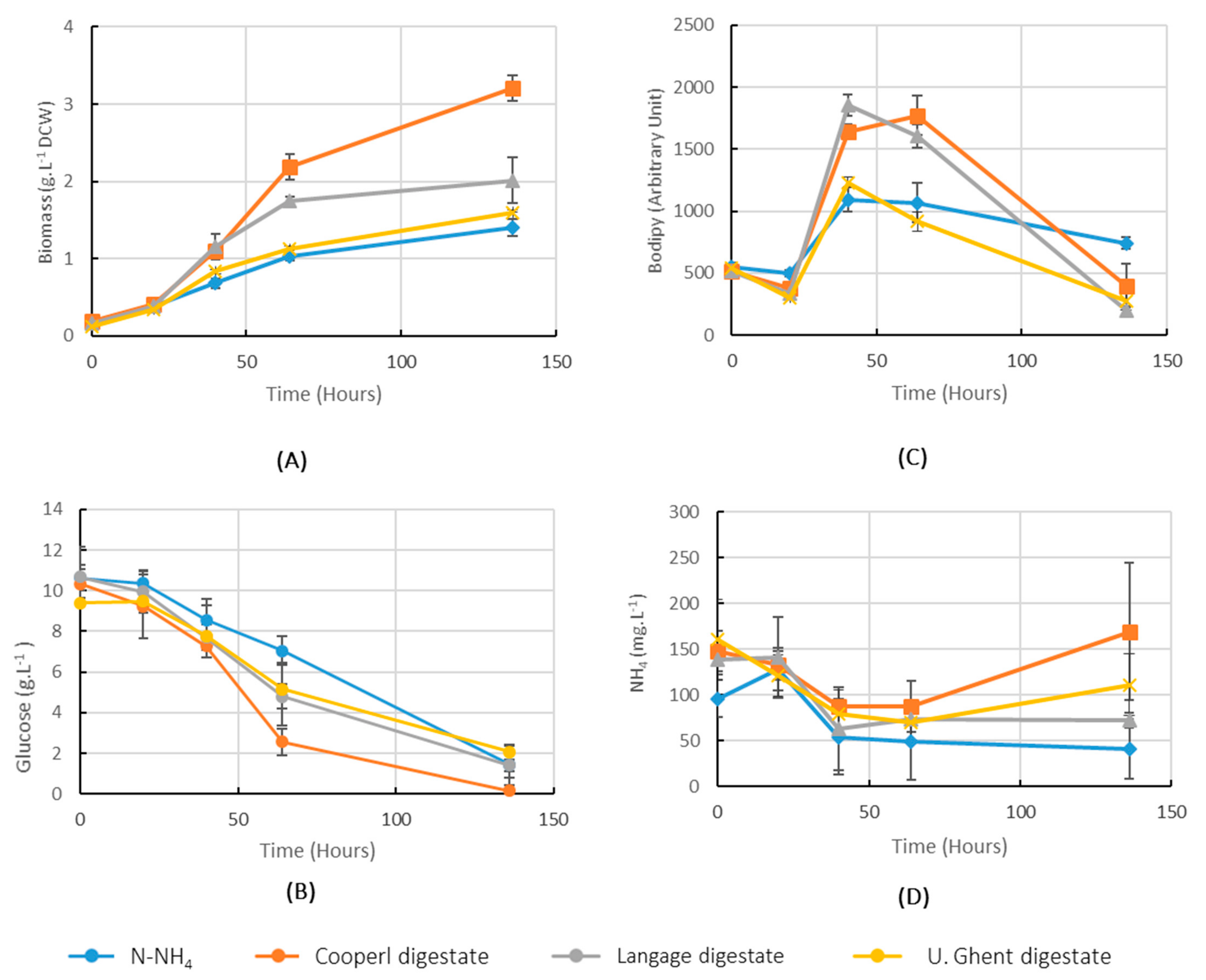
2.2. Comparison of Performance of Cultures in Digestates from Three Different Sources
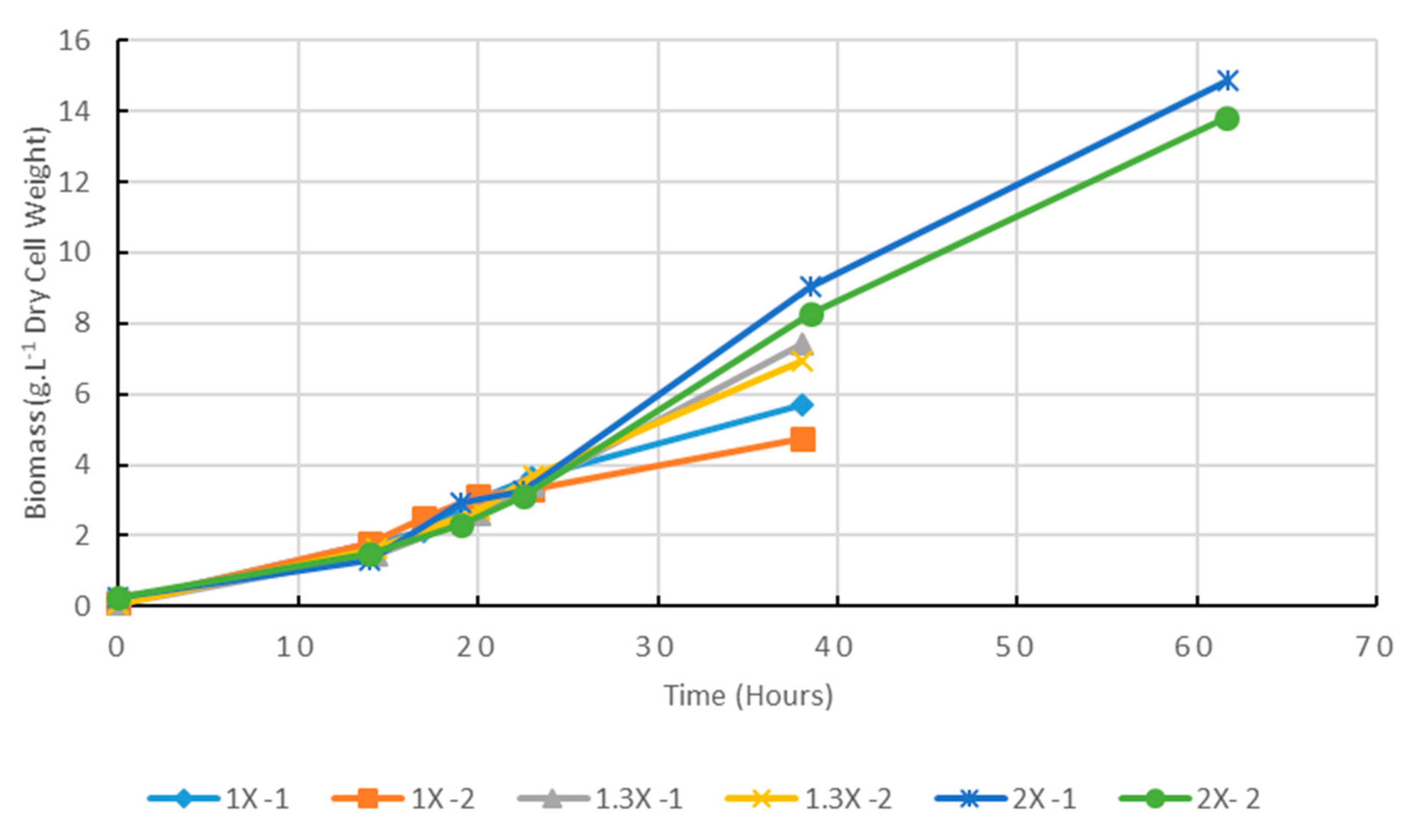
2.3. The Digestate Concentration Influences the Growth Kinetics in Flask Cultures
2.4. Batch Cultures Can Be Run in Non-Axenic Airlift Cylinders
2.5. Non-Axenic Cultivation in Pilot-Scale (500 L) Culture in Batch and Fed-Batch Modes Appears Feasible
2.6. Bacterial Contamination Assessment in Non-Axenic Cultures
2.7. Glucose Conversion and Nitrogen Trapping Efficiency
2.8. Lipids and Proteins in Pilot-Scale Production
2.9. Productivity and Conversion Yields of Substrate to Biomass
3. Discussion
3.1. Non-Axenic Cultivation
3.2. Substrates
3.3. Limiting Factor for Growth and Productivity
3.4. Lipids
3.5. AD Digestate Regulation
4. Materials and Methods
4.1. Culture Media
4.1.1. Modified YEP Medium for Lab Experiments and Pilot Precultures
4.1.2. Modified Guillard and Ryther F/2 Medium for Digestate Comparison Experiment
- Digestates and NH4Cl (297 g.L−1) solution were filter-sterilized through 0.2 µm filter.
- The vitamin solution was prepared as follows: cyanocobalamin: 5 mg; biotin: 5 mg; thiamine HCl: 1 g; distilled water: 500 mL, autoclaved 20 min at 120 °C.
- The trace element solution was prepared as follows: stock solution: CuSO4.5H2O: 100 mg, ZnSO4·7H2O: 220 mg, CoCl2·6H2O: 100 mg, MnCl2·4H2O: 1800 mg and Na2MoO4·2H2O: 60 mg. Distilled water: 200 mL. The final trace element solution contained stock solution: 20 mL; Na2EDTA: 4.36 g; FeCl3·6H2O: 3.15 g; distilled water: 980 mL. Solution was filter-sterilized through 0.2 µm.
- Each 800 mL culture was prepared independently as follows:
4.1.3. Culture Media for Digestate Concentration Tests
4.1.4. Medium for Non-Axenic Culture in 10 L Cylinder Experiments at Lab
4.1.5. Medium for Non-Axenic 400 or 500 L Culture in 800 L Cylinders at Pilot Plant
4.2. Thraustochytrid Strain
4.3. Pre-Cultures
4.3.1. Pre-Cultures for Lab Experiments
4.3.2. Pre-Cultures for Pilot-Scale Experiments
4.4. Cultures at Laboratory Scale
4.4.1. Scale-Up of Axenic 8 L Cultures in 20 L Carboys
4.4.2. Comparison of Three Sources of Digestate
4.4.3. Effect of Digestate Concentration on Growth Kinetics in Flask Cultures
4.4.4. Scale-up to Non-Axenic 6 L Cultures in 10 L Cylinders
4.5. Cultures at Pilot Scale
4.6. Analysis
4.6.1. Dry Cell Weight (DCW)
4.6.2. Monitoring of Culture Concentration and Cellular Parameters
4.6.3. Nutrient Content Analysis
4.6.4. Lipid Content Analysis
- Lipid Extraction:
- Separation of polar lipids (PL) and neutral lipids (NL):
- Fatty acid analysis in total lipids (TL), polar lipids (PL) and neutral lipids (NL):
4.6.5. Protein Analysis
4.6.6. Conversion Yield
4.6.7. On Line Measurements
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; Food and Agriculture Organization of the United Nations FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- Tocher, D.R. Omega-3 Long-Chain Polyunsaturated Fatty Acids and Aquaculture in Perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Arts, M.T.; Ackman, R.G.; Holub, B.J. “Essential Fatty Acids” in Aquatic Ecosystems: A Crucial Link between Diet and Human Health and Evolution. Can. J. Fish. Aquat. Sci. 2001, 58, 122–137. [Google Scholar] [CrossRef]
- Amminger, G.P.; Schäfer, M.R.; Schlögelhofer, M.; Klier, C.M.; McGorry, P.D. Longer-Term Outcome in the Prevention of Psychotic Disorders by the Vienna Omega-3 Study. Nat. Commun. 2015, 6, 7934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, M.A.; Broadhurst, C.L. The Role of Docosahexaenoic and the Marine Food Web as Determinants of Evolution and Hominid Brain Development: The Challenge for Human Sustainability. Nutr. Health 2012, 21, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture; Food and Agriculture Organization of the United Nations FAO: Rome, Italy, 2014. [Google Scholar]
- Lotze, H.K.; Tittensor, D.P.; Bryndum-Buchholz, A.; Eddy, T.D.; Cheung, W.W.L.; Galbraith, E.D.; Barange, M.; Barrier, N.; Bianchi, D.; Blanchard, J.L.; et al. Global Ensemble Projections Reveal Trophic Amplification of Ocean Biomass Declines with Climate Change. Proc. Natl. Acad. Sci. USA 2019, 116, 12907–12912. [Google Scholar] [CrossRef] [Green Version]
- Fossier Marchan, L.; Lee Chang, K.J.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, Ecology and Biotechnological Applications of Thraustochytrids: A Review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Karageorgou, D.; Katapodis, P.; Sharma, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Bioprospecting of Thraustochytrids for Omega-3 Fatty Acids: A Sustainable Approach to Reduce Dependency on Animal Sources. Trends Food Sci. Technol. 2021, 115, 433–444. [Google Scholar] [CrossRef]
- Orozco Colonia, B.S.; Vinícius de Melo Pereira, G.; Soccol, C.R. Omega-3 Microbial Oils from Marine Thraustochytrids as a Sustainable and Technological Solution: A Review and Patent Landscape. Trends Food Sci. Technol. 2020, 99, 244–256. [Google Scholar] [CrossRef]
- Du, F.; Wang, Y.-Z.; Xu, Y.-S.; Shi, T.-Q.; Liu, W.-Z.; Sun, X.-M.; Huang, H. Biotechnological Production of Lipid and Terpenoid from Thraustochytrids. Biotechnol. Adv. 2021, 48, 107725. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Mao, W.; Zhang, C.; Liu, L.; Zhang, Z.; Guo, C.; Lin, J. Effective and Economic Microbial Lipid Biosynthesis for Biodiesel Production by Two-Phase Whole-Cell Biocatalytic Process. J. Clean. Prod. 2021, 298, 126798. [Google Scholar] [CrossRef]
- Loïez, A. Production de La Levure de Panification Par Biotechnologie; Techniques de L’ingénieur Bioprocédés et Bioproductions 2003, Base Documentaire: TIP140WEB; Techniques de L’ingénieur: Paris, France, 2003. [Google Scholar]
- Gahlawat, G.; Kumari, P.; Bhagat, N.R. Technological Advances in the Production of Polyhydroxyalkanoate Biopolymers. Curr. Sustain. Renew. Energy Rep. 2020, 7, 73–83. [Google Scholar] [CrossRef]
- Lei, M.; Peng, X.; Sun, W.; Zhang, D.; Wang, Z.; Yang, Z.; Zhang, C.; Yu, B.; Niu, H.; Ying, H.; et al. Nonsterile L-Lysine Fermentation Using Engineered Phosphite-Grown Corynebacterium Glutamicum. ACS Omega 2021, 6, 10160–10167. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ren, L.-J.; Sun, G.-N.; Ji, X.-J.; Nie, Z.-K.; Huang, H. Batch, Fed-Batch and Repeated Fed-Batch Fermentation Processes of the Marine Thraustochytrid Schizochytrium sp. For Producing Docosahexaenoic Acid. Bioprocess Biosyst. Eng. 2013, 36, 1905–1912. [Google Scholar] [CrossRef]
- Ye, H.; He, Y.; Xie, Y.; Sen, B.; Wang, G. Fed-Batch Fermentation of Mixed Carbon Source Significantly Enhances the Production of Docosahexaenoic Acid in Thraustochytriidae sp. PKU#Mn16 by Differentially Regulating Fatty Acids Biosynthetic Pathways. Bioresour. Technol. 2020, 297, 122402. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Villarroel, M.P.; Roa, A.L.; Quilodrán, B.H. Use of Waste from Agroindustrial Sources as Substrate for Polyunsaturated Fatty Acids Production by Thraustochytrium Kinney VAL-B1. Int. J. Eng. Res. Afr. 2017, 33, 50–55. [Google Scholar] [CrossRef]
- Yamasaki, T.; Aki, T.; Shinozaki, M.; Taguchi, M.; Kawamoto, S.; Ono, K. Utilization of Shochu Distillery Wastewater for Production of Polyunsaturated Fatty Acids and Xanthophylls Using Thraustochytrid. J. Biosci. Bioeng. 2006, 102, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Smerilli, M.; Neureiter, M.; Wurz, S.; Haas, C.; Frühauf, S.; Fuchs, W. Direct Fermentation of Potato Starch and Potato Residues to Lactic Acid by Geobacillus Stearothermophilus under Non-Sterile Conditions. J. Chem. Technol. Biotechnol. 2015, 90, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monod, J. The Growth of Bacterial Cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ochoa, F.; Gomez, E. Bioreactor Scale-up and Oxygen Transfer Rate in Microbial Processes: An Overview. Biotechnol. Adv. 2009, 27, 153–176. [Google Scholar] [CrossRef]
- Collos, Y.; Harrison, P.J. Acclimation and Toxicity of High Ammonium Concentrations to Unicellular Algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef]
- Raghukumar, S. Bacterivory: A Novel Dual Role for Thraustochytrids in the Sea. Mar. Biol. 1992, 113, 165–169. [Google Scholar] [CrossRef]
- Dalsgaard, J.; St. John, M.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty Acid Trophic Markers in the Pelagic Marine Environment. In Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 2003; Volume 46, pp. 225–340. [Google Scholar]
- Najdek, M.; Debobbis, D.; Mioković, D.; Ivančić, I. Fatty Acid and Phytoplankton Compositions of Different Types of Mucilaginous Aggregates in the Northern Adriatic. J. Plankton Res. 2002, 24, 429–441. [Google Scholar] [CrossRef]
- Bonmatıí, A.; Flotats, X. Air Stripping of Ammonia from Pig Slurry: Characterisation and Feasibility as a Pre- or Post-Treatment to Mesophilic Anaerobic Digestion. Waste Manag. 2003, 23, 261–272. [Google Scholar] [CrossRef]
- Soudant, P.; Ventura, M.; Chauchat, L.; Guerreiro, M.; Le Grand, F.; Simon, V.; Collet, S.; Zambonino-Infante, J.-L.; Le Goïc, N.; Lambert, C.; et al. Evaluation of Aurantiochytrium mangrovei Biomass Grown on Digestate as a Feed Ingredient of Sea Bass, Dicentrarchus labrax, Juveniles and Larvae. 2022; submitted. [Google Scholar]
- Lisičar, J.; Scheper, T.; Barbe, S. Turning Industrial Baker’s Yeast Manufacture into a Powerful Zero Discharge Multipurpose Bioprocess. Ind. Biotechnol. 2017, 13, 184–191. [Google Scholar] [CrossRef]
- Jakobsen, A.N.; Aasen, I.M.; Josefsen, K.D.; Strøm, A.R. Accumulation of Docosahexaenoic Acid-Rich Lipid in Thraustochytrid Aurantiochytrium sp. Strain T66: Effects of N and P Starvation and O2 Limitation. Appl. Microbiol. Biotechnol. 2008, 80, 297–306. [Google Scholar] [CrossRef]
- Chi, Z.; Liu, Y.; Frear, C.; Chen, S. Study of a Two-Stage Growth of DHA-Producing Marine Algae Schizochytrium Limacinum SR21 with Shifting Dissolved Oxygen Level. Appl. Microbiol. Biotechnol. 2009, 81, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.; Guo, J.; Liu, X.; Zhang, X.; Wang, N.; Lu, Y.; Ng, I.-S. Impact of Carbon and Nitrogen Feeding Strategy on High Production of Biomass and Docosahexaenoic Acid (DHA) by Schizochytrium sp. LU310. Bioresour. Technol. 2015, 184, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.; Ryther, J.H. Studies of Marine Planktonic Diatoms. I. Cyclotella Nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Lelong, A.; Jolley, D.F.; Soudant, P.; Hégaret, H. Impact of Copper Exposure on Pseudo-Nitzschia spp. Physiology and Domoic Acid Production. Aquat. Toxicol. 2012, 118–119, 37–47. [Google Scholar] [CrossRef]
- Hansen, H.P.; Koroleff, F. Determination of Nutrients. In Methods of Seawater Analysis: Third, Completely Revised and Extended Edition; John and Wiley and Sons: Hoboken, NJ, USA, 2007; pp. 159–228. ISBN 978-3-527-61398-4. [Google Scholar]
- Eppley, R.W.; Coatsworth, J.L.; Solórzano, L. Studies of Nitrate Reductase in Marine Phytoplankton. Limnol. Oceanogr. 1969, 14, 194–205. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A Modified Single Solution Method for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Sardenne, F.; Bodin, N.; Metral, L.; Crottier, A.; Le Grand, F.; Bideau, A.; Brisset, B.; Bourjea, J.; Saraux, C.; Bonhommeau, S.; et al. Effects of Extraction Method and Storage of Dry Tissue on Marine Lipids and Fatty Acids. Anal. Chim. Acta 2019, 1051, 82–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marty, Y.; Soudant, P.; Perrotte, S.; Moal, J.; Dussauze, J.; Samain, J.F. Identification and Occurrence of a Novel Cis-4,7,10,Trans-13-Docosatetraenoic Fatty Acid in the Scallop Pecten maximus (L.). J. Chromatogr. A 1999, 839, 119–127. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Condition | Initial-Culture Medium | Feed | Culture Volume (L) | Culture Time (Days) | Biomass Concentration (g.L−1 ± S.D.) | Productivity (gDCW.L−1.day−1) | Yield | |
|---|---|---|---|---|---|---|---|---|
| Flask-500 mL axenic | 100 rpm | YEP modified | / | 0.25 | 14 | 4.3 | 0.31 | 0.18 |
| Flask-1 L axenic | 100 rpm + 0.4 VVM | Glucose + Salts | / | 0.8 | 5.67 | 3.21 ± 0.16 | 0.57 | 0.32 |
| Carboy-20 L axenic | 100 rpm + 0.4 VVM | YEP modified + 2.5% digestate | / | 6 | 2.75 | 2.49 | 0.91 | 0.10 |
| Cylinder-10 L non-axenic | Air-lift + 0.4 VVM-large bubbles | YEP modified + 2.5% digestate | / | 6 | 2.75 | 2.82 | 1.03 | 0.12 |
| Cylinder-10 L non-axenic | Air-lift + 0.8 VVM-small bubbles | YEP modified + 2.5% digestate | / | 6 | 2.75 | 3.71 | 1.35 | 0.15 |
| Cylinder-800 L non-axenic | Air-lift + 0.4 VVM | YEP modified + 2.5% digestate | / | 500 | 1.58 | 5.21 ± 0.66 | 3.29 | 0.22 |
| Cylinder-800 L non-axenic | Air-lift + 0.4 VVM | YEP modified + 2.5% digestate | +0.3X medium | 500 | 1.58 | 7.15 ± 0.35 | 4.52 | 0.23 |
| Cylinder-800 L non-axenic | Air-lift + 0.4 VVM | YEP modified + 2.5% digestate | +1X medium | 500 | 2.57 | 14.34 ± 0.72 | 5.57 | 0.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Broise, D.; Ventura, M.; Chauchat, L.; Guerreiro, M.; Michez, T.; Vinet, T.; Gautron, N.; Le Grand, F.; Bideau, A.; Goïc, N.L.; et al. Scale-Up to Pilot of a Non-Axenic Culture of Thraustochytrids Using Digestate from Methanization as Nitrogen Source. Mar. Drugs 2022, 20, 499. https://doi.org/10.3390/md20080499
de la Broise D, Ventura M, Chauchat L, Guerreiro M, Michez T, Vinet T, Gautron N, Le Grand F, Bideau A, Goïc NL, et al. Scale-Up to Pilot of a Non-Axenic Culture of Thraustochytrids Using Digestate from Methanization as Nitrogen Source. Marine Drugs. 2022; 20(8):499. https://doi.org/10.3390/md20080499
Chicago/Turabian Stylede la Broise, Denis, Mariana Ventura, Luc Chauchat, Maurean Guerreiro, Teo Michez, Thibaud Vinet, Nicolas Gautron, Fabienne Le Grand, Antoine Bideau, Nelly Le Goïc, and et al. 2022. "Scale-Up to Pilot of a Non-Axenic Culture of Thraustochytrids Using Digestate from Methanization as Nitrogen Source" Marine Drugs 20, no. 8: 499. https://doi.org/10.3390/md20080499
APA Stylede la Broise, D., Ventura, M., Chauchat, L., Guerreiro, M., Michez, T., Vinet, T., Gautron, N., Le Grand, F., Bideau, A., Goïc, N. L., Bidault, A., Lambert, C., & Soudant, P. (2022). Scale-Up to Pilot of a Non-Axenic Culture of Thraustochytrids Using Digestate from Methanization as Nitrogen Source. Marine Drugs, 20(8), 499. https://doi.org/10.3390/md20080499