
Ilimaquinone (Marine Sponge Metabolite) Induces Apoptosis in HCT-116 Human Colorectal Carcinoma Cells via Mitochondrial-Mediated Apoptosis Pathway

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
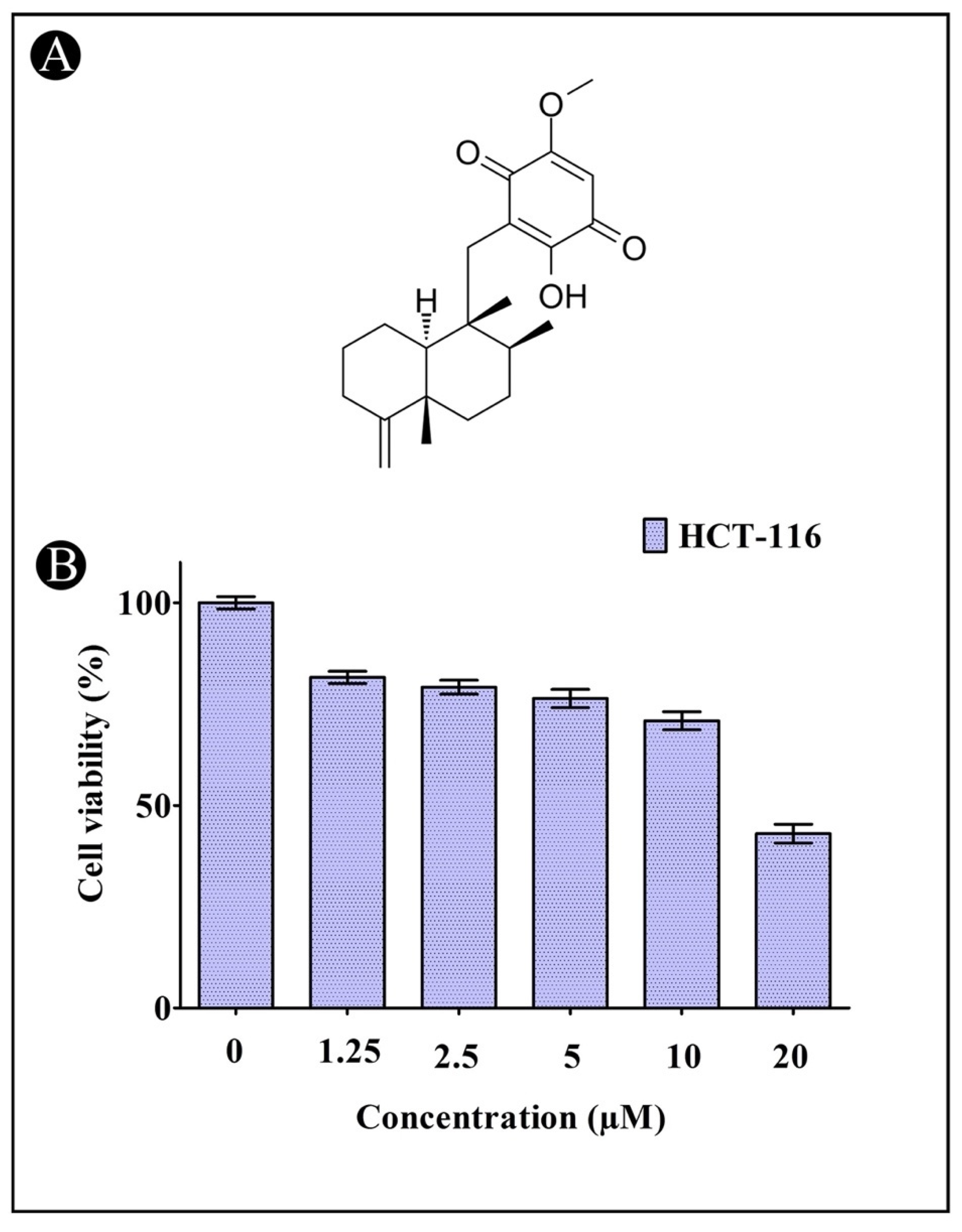
2.1. Induction of Cytotoxicity by IQ in HCT-116 Cells
2.2. Apoptosis Induction by IQ
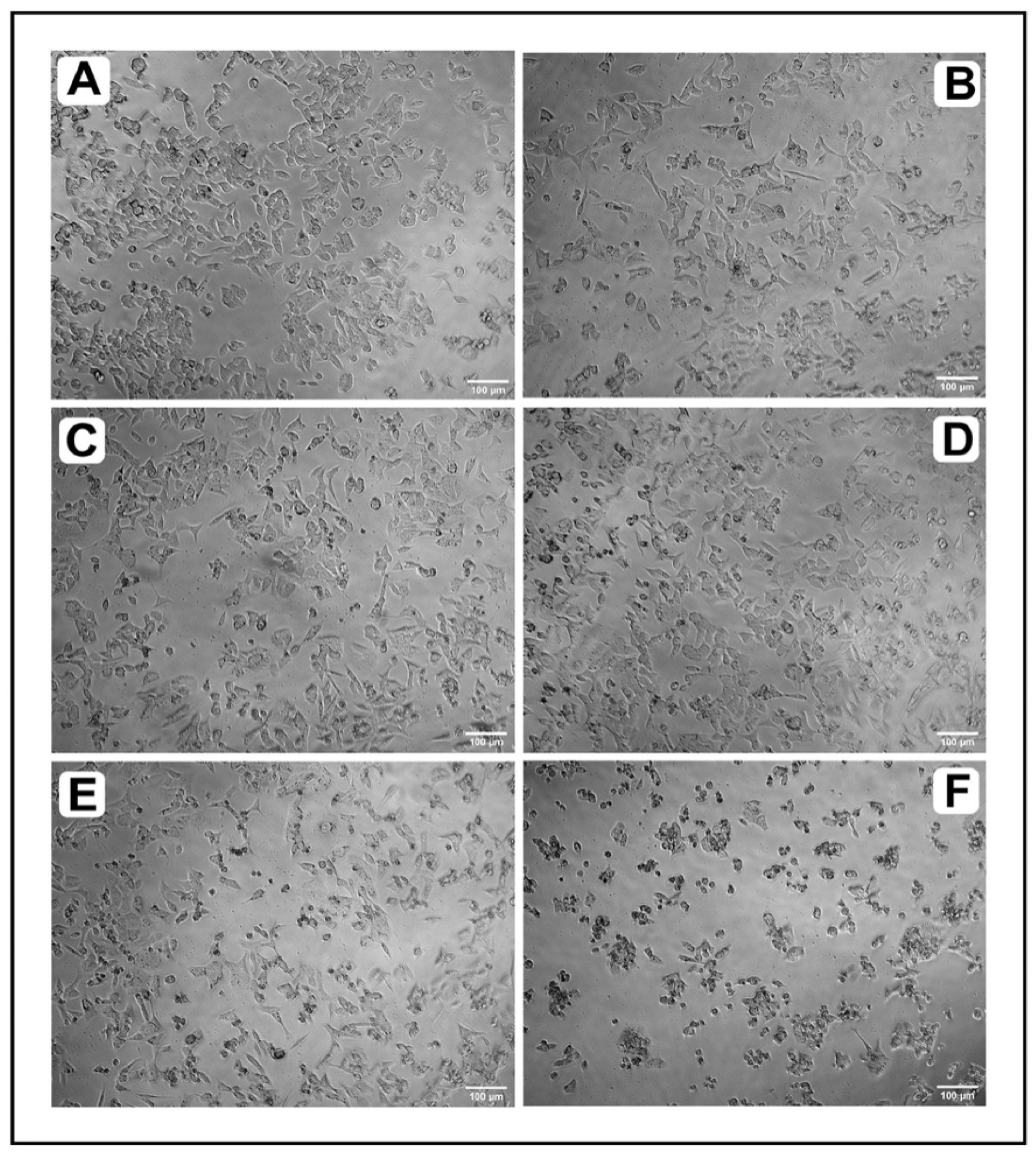
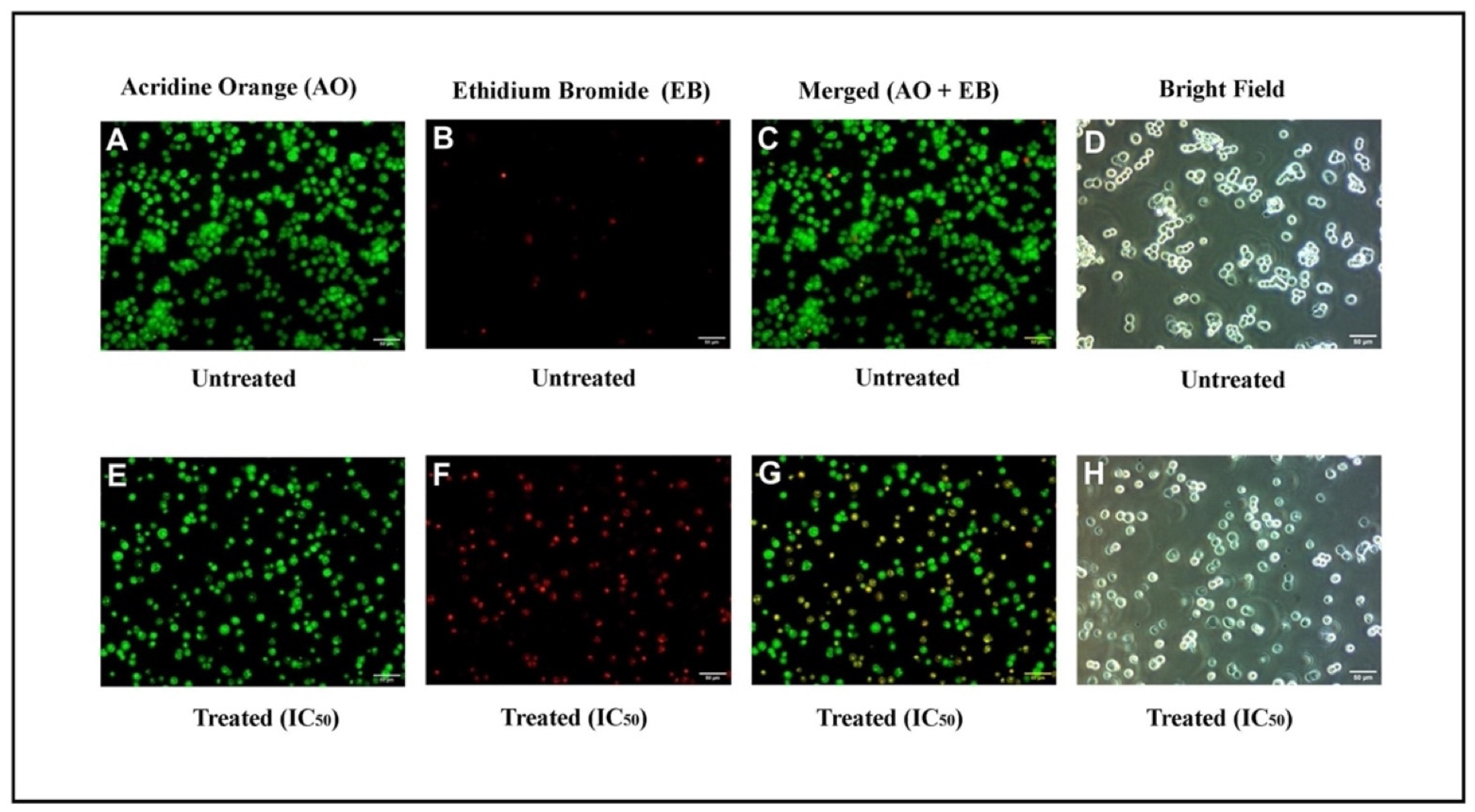
2.2.1. Microscopic Analysis
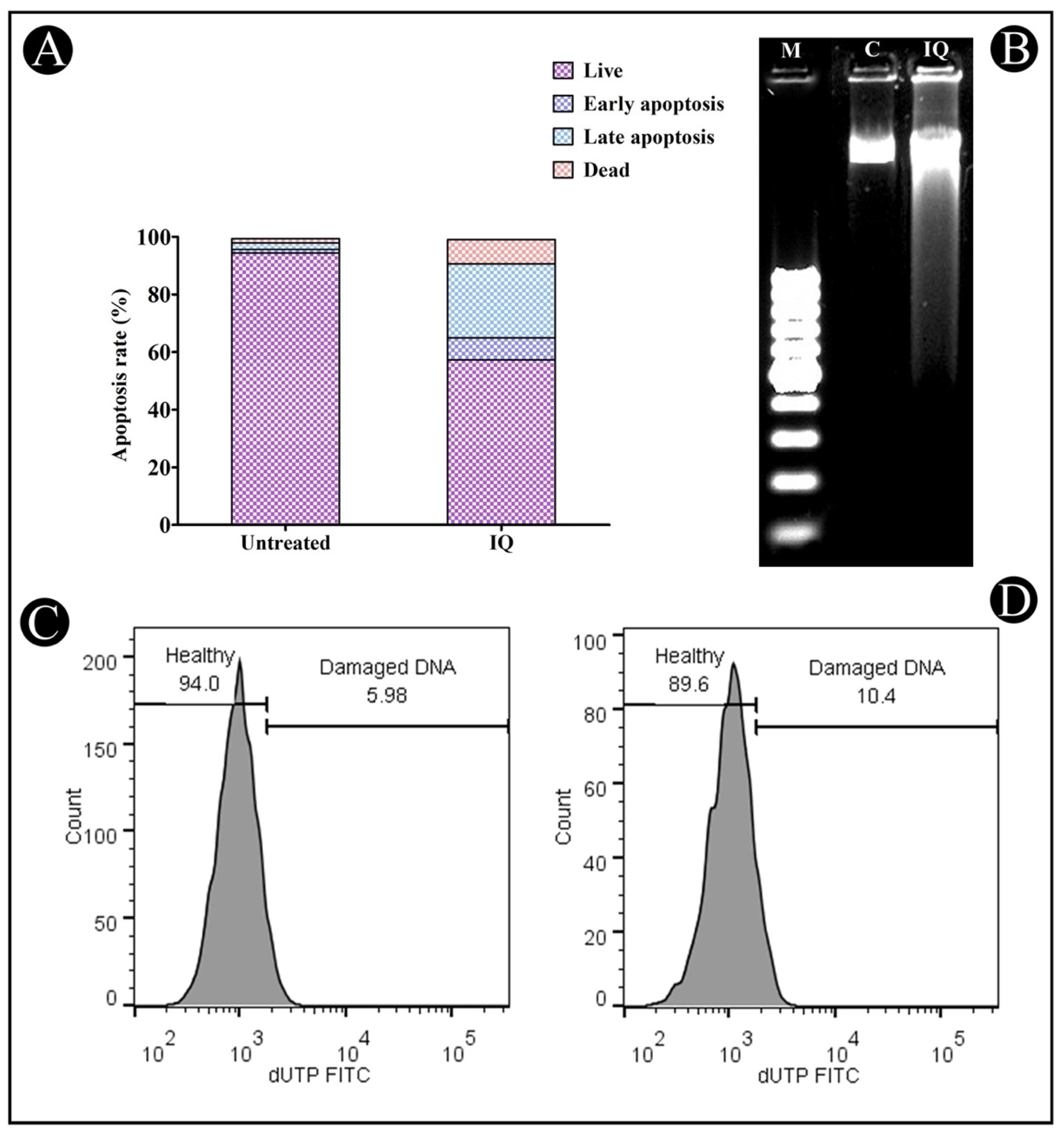
2.2.2. Annexin V–Propidium Iodide (PI) Staining
2.3. Induction of DNA Fragmentation by IQ
TUNEL Assay
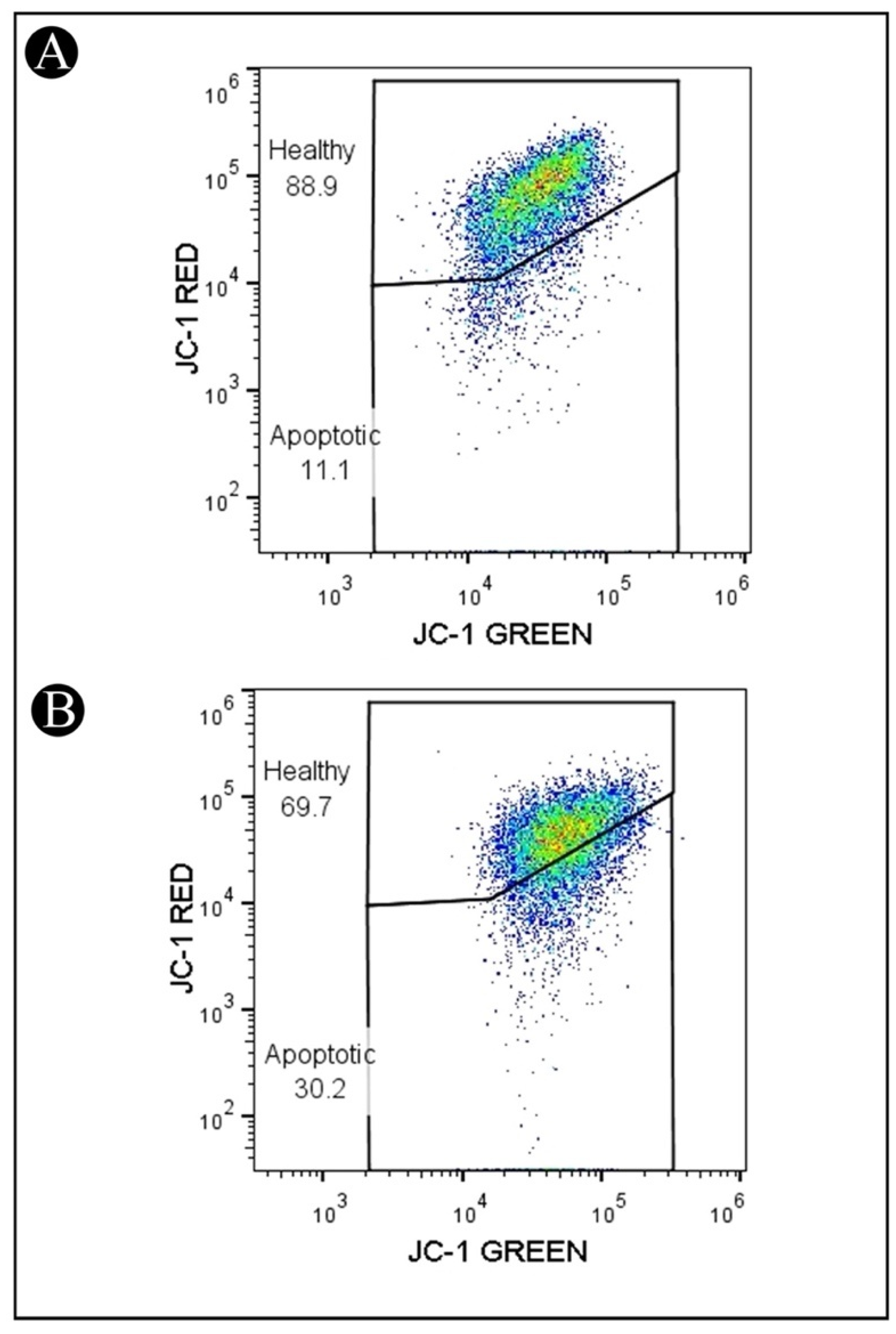
2.4. Reduction in ΔΨm in HCT-116 Cells Treated with IQ
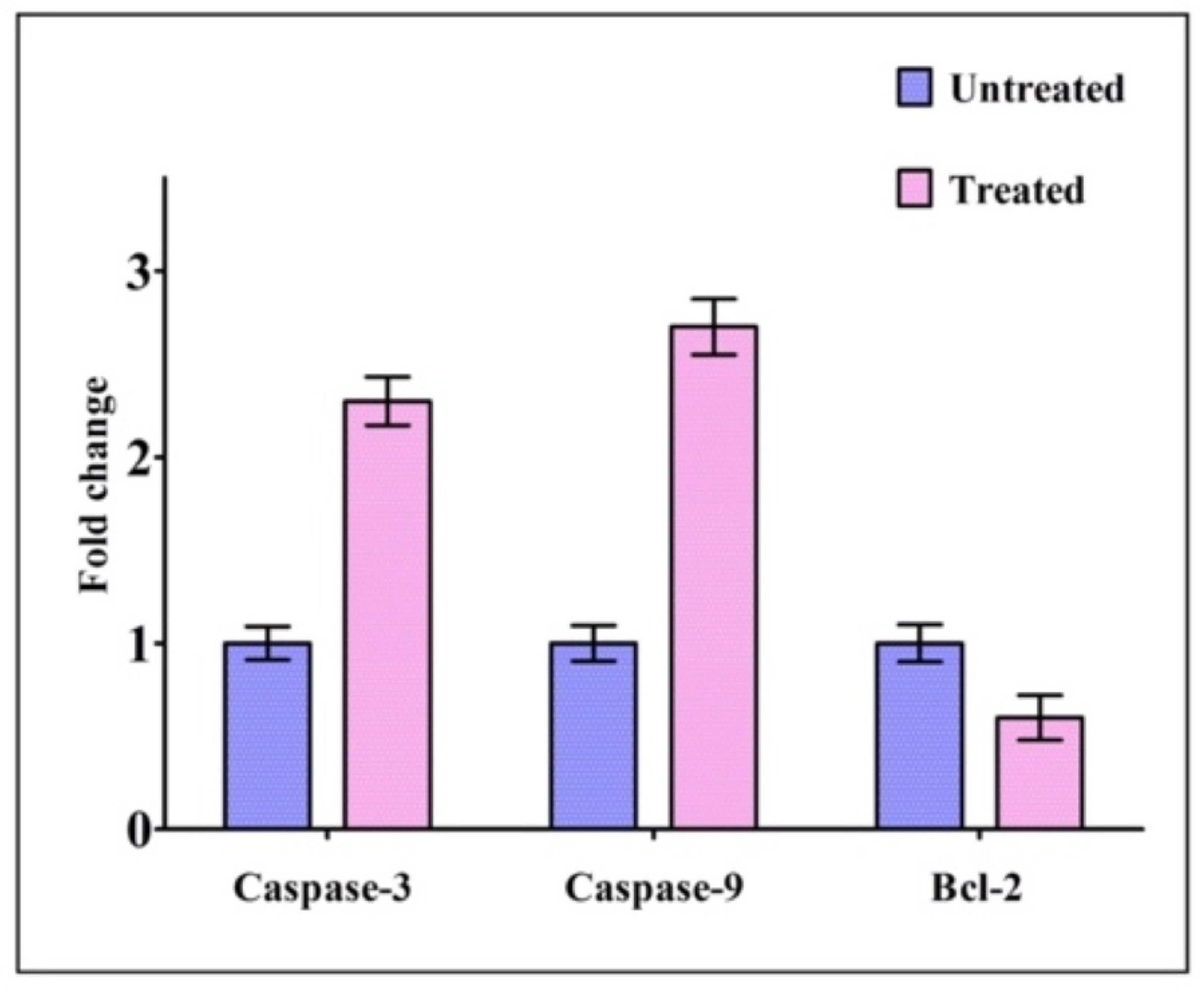
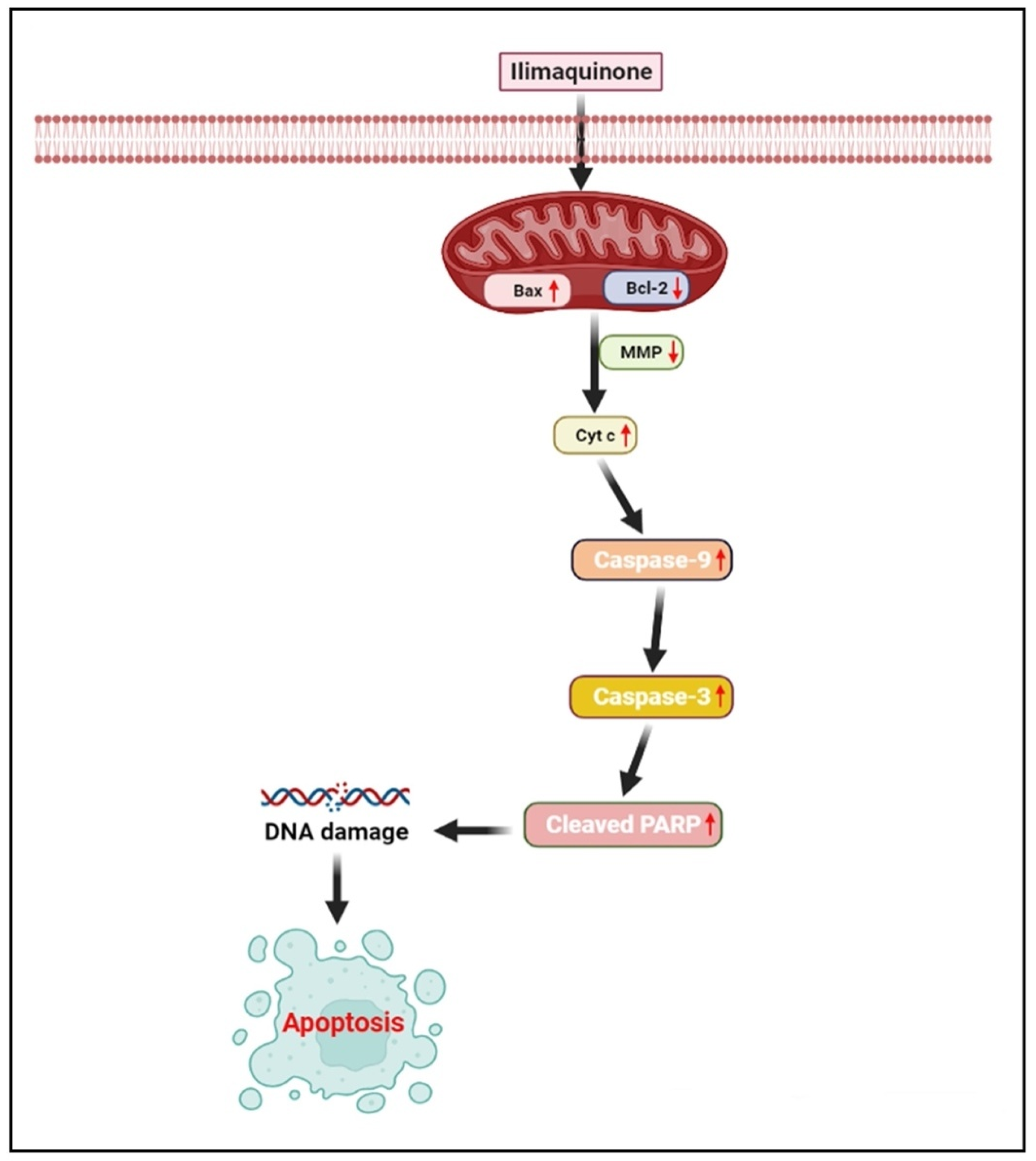
2.5. Expression of Genes Related to Apoptosis
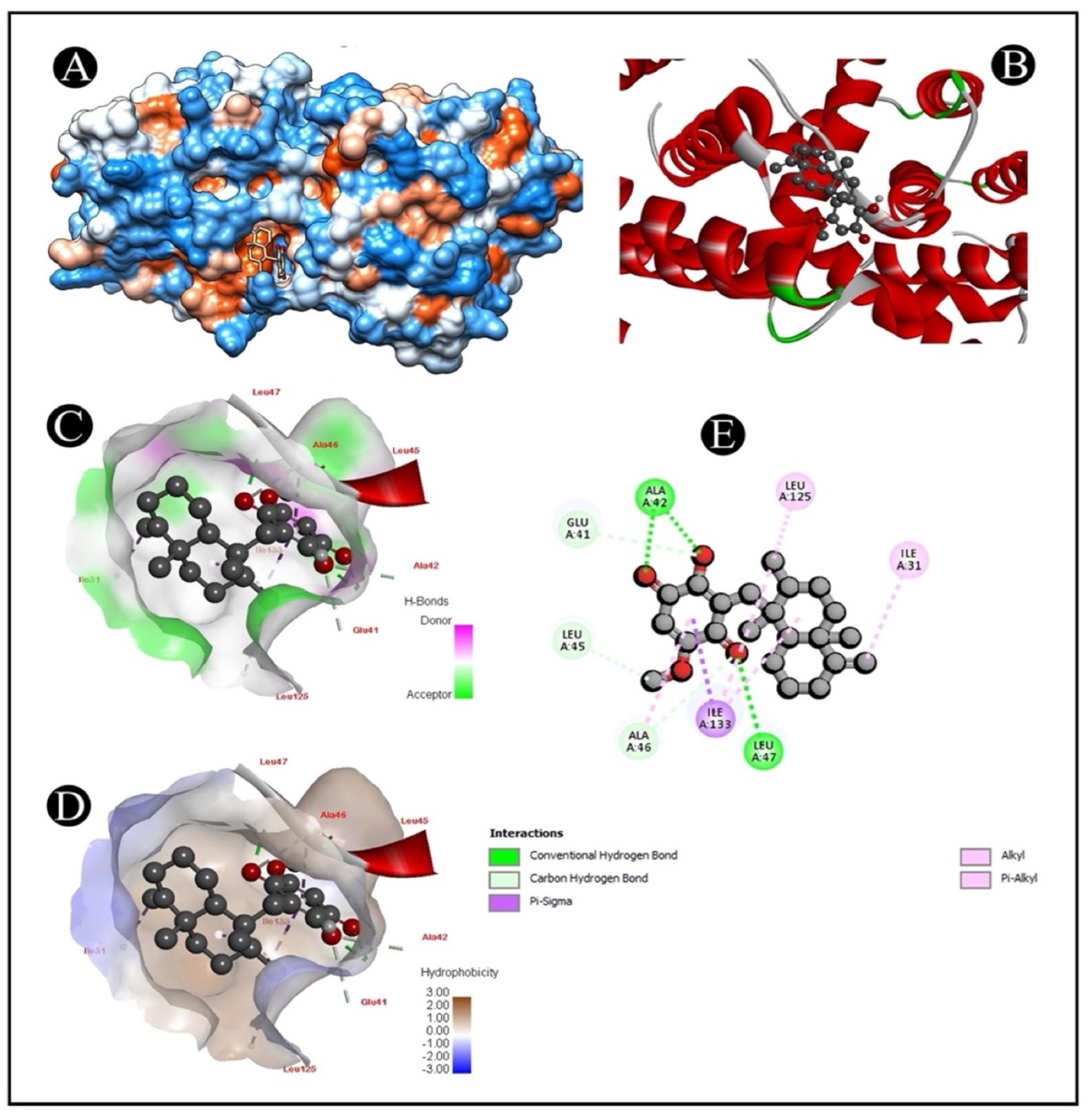
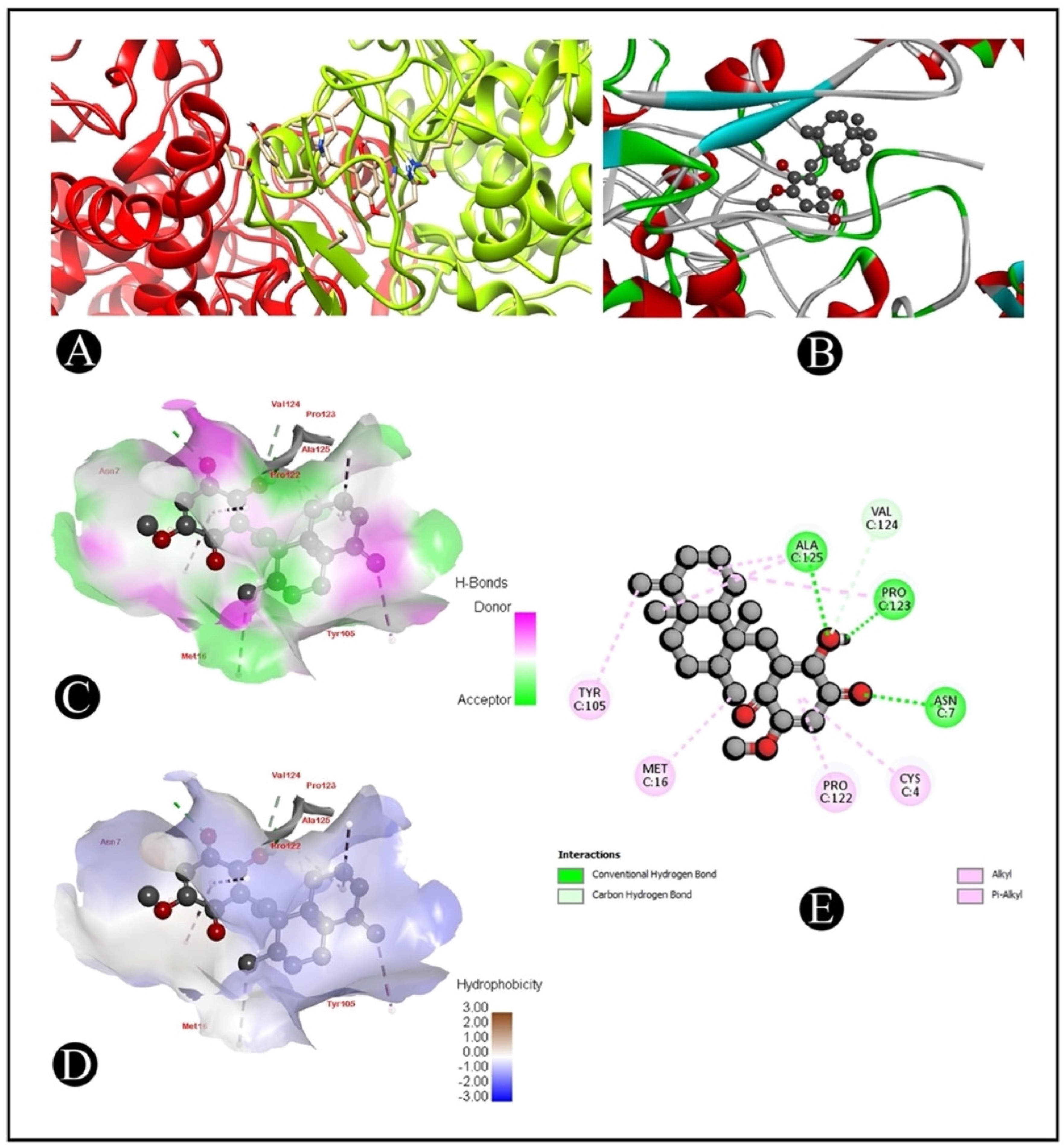
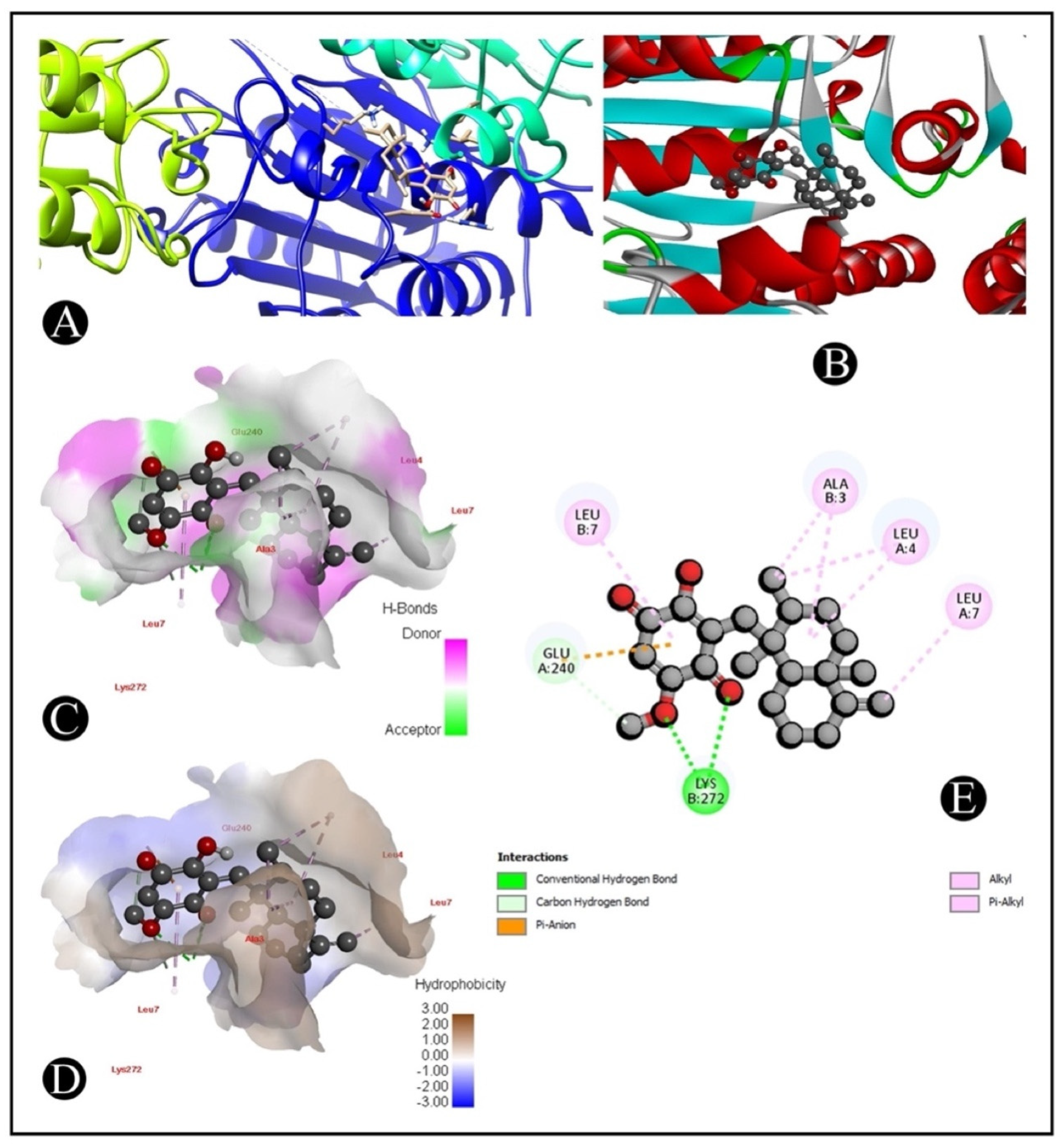
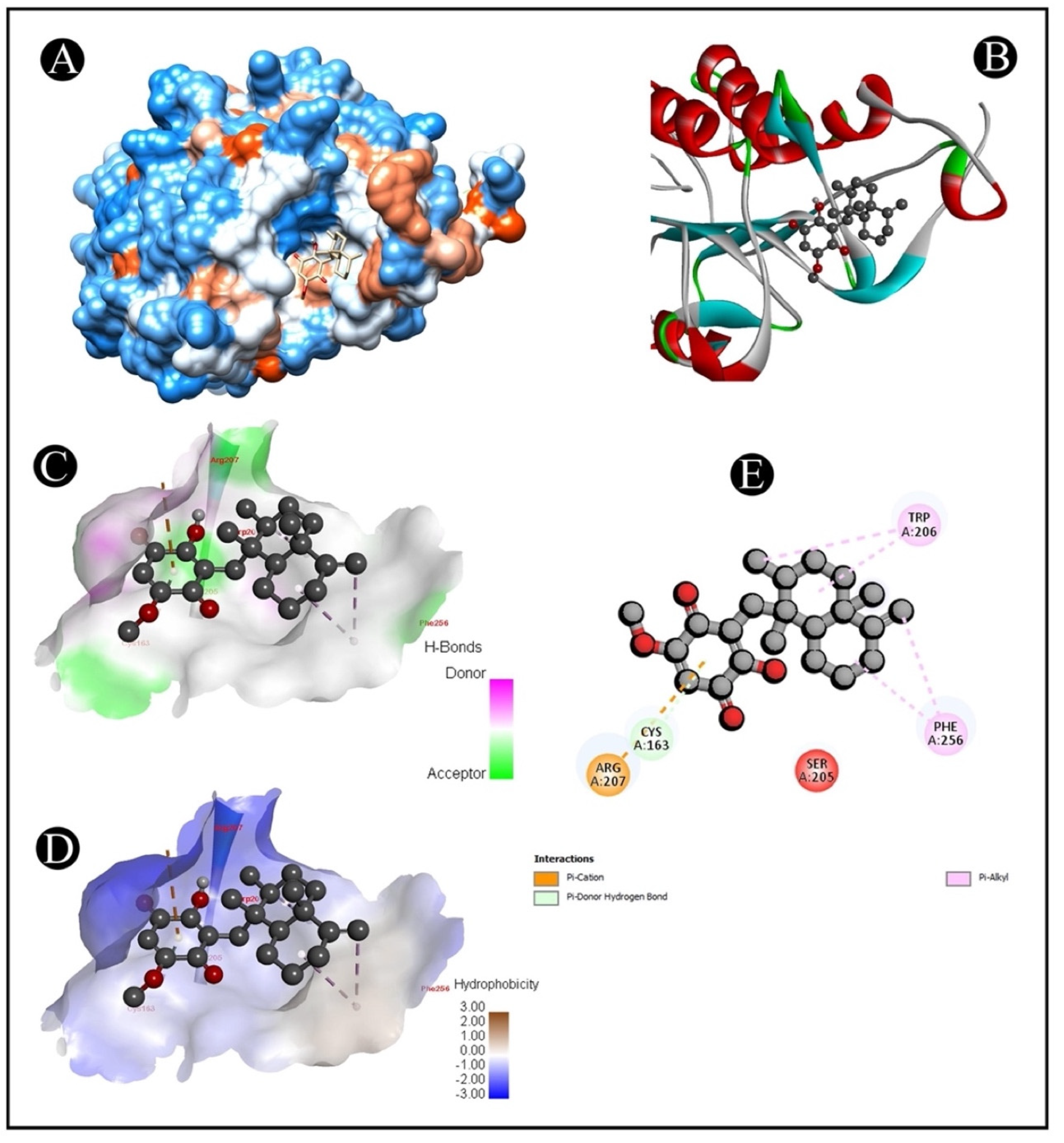
2.6. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability Assay
4.3. Acridine Orange–Ethidium Bromide (AO/EB) Staining
4.4. Annexin V–PI Apoptosis Assay
4.5. DNA Fragmentation Assay
4.6. Apoptotic DNA Degradation Assay by TUNEL (Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling) Assay
4.7. Mitochondrial Membrane Assay
4.8. Analysis of the Expression of Genes Involved in Apoptosis
4.9. Molecular Docking Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Goding Sauer, A.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal Cancer Statistics, 2020. CA. Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Siddiqui, A.J.; Hamadou, W.S.; Snoussi, M.; Badraoui, R.; Ashraf, S.A.; Jamal, A.; Awadelkareem, A.M.; Sachidanandan, M.; Hadi, S. Deciphering the Molecular Mechanism Responsible for Efficiently Inhibiting Metastasis of Human Non-Small Cell Lung and Colorectal Cancer Cells Targeting the Matrix Metalloproteinases by Selaginella Repanda. Plants 2021, 10, 979. [Google Scholar] [CrossRef]
- Ciftci, H.I.; Can, M.; Ellakwa, D.E.; Suner, S.C.; Ibrahim, M.A.; Oral, A.; Sekeroglu, N.; Özalp, B.; Otsuka, M.; Fujita, M. Anticancer Activity of Turkish Marine Extracts: A Purple Sponge Extract Induces Apoptosis with Multitarget Kinase Inhibition Activity. Investig. New Drugs 2020, 38, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Varijakzhan, D.; Loh, J.-Y.; Yap, W.-S.; Yusoff, K.; Seboussi, R.; Lim, S.-H.E.; Lai, K.-S.; Chong, C.-M. Bioactive Compounds from Marine Sponges: Fundamentals and Applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Sorolla, M.A.; Gopal Krishnan, P.D.; Sorolla, A. From Seabed to Bedside: A Review on Promising Marine Anticancer Compounds. Biomolecules 2020, 10, 248. [Google Scholar] [CrossRef]
- Luibrand, R.T.; Erdman, T.R.; Vollmer, J.J.; Scheuer, P.J.; Finer, J.; Clardy, J. Ilimaquinone, a Sesquiterpenoid Quinone from a Marine Sponge. Tetrahedron 1979, 35, 609–612. [Google Scholar] [CrossRef]
- D’yakonov, V.A.; Dzhemileva, L.U.; Dzhemilev, U.M. Advances in the Chemistry of Natural and Semisynthetic Topoisomerase I/II Inhibitors. Stud. Nat. Prod. Chem. 2017, 54, 21–86. [Google Scholar]
- Bourquet-Kondracki, M.-L.; Longeon, A.; Morel, E.; Guyot, M. Sesquiterpene Quinones as Immunomodulating Agents. Int. J. Immunopharmacol. 1991, 13, 393–399. [Google Scholar] [CrossRef]
- Rangel, H.R.; Dagger, F.; Compagnone, R.S. Antiproliferative Effect of Illimaquinone on Leishmania Mexicana. Cell Biol. Int. 1997, 21, 337–339. [Google Scholar] [CrossRef]
- Lu, P.-H.; Chueh, S.-C.; Kung, F.-L.; Pan, S.-L.; Shen, Y.-C.; Guh, J.-H. Ilimaquinone, a Marine Sponge Metabolite, Displays Anticancer Activity via GADD153-Mediated Pathway. Eur. J. Pharmacol. 2007, 556, 45–54. [Google Scholar] [CrossRef]
- Popov, A.M.; Stekhova, S.I.; Utkina, N.K.; Rebachuk, N.M. Antimicrobial and Cytotoxic Activity of Sesquiterpenequinones and Brominated Diphenyl Esters Isolated from Marine Sponges. Pharm. Chem. J. 1999, 33, 71–73. [Google Scholar] [CrossRef]
- Loya, S.; Hizi, A. The Interaction of Illimaquinone, a Selective Inhibitor of the RNase H Activity, with the Reverse Transcriptases of Human Immunodeficiency and Murine Leukemia Retroviruses. J. Biol. Chem. 1993, 268, 9323–9328. [Google Scholar] [CrossRef]
- Son, H.; Noh, K.; Kang, C.; Na, M.; Oh, S.; Song, I.-S.; Kang, W. HPLC-MS/MS Analysis of Ilimaquinone and Its Application in a Pharmacokinetic Study in Rats. J. Pharm. Biomed. Anal. 2019, 166, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Kwak, C.-H.; Jin, L.; Han, J.H.; Han, C.W.; Kim, E.; Cho, M.; Chung, T.-W.; Bae, S.-J.; Jang, S.B.; Ha, K.-T. Ilimaquinone Induces the Apoptotic Cell Death of Cancer Cells by Reducing Pyruvate Dehydrogenase Kinase 1 Activity. Int. J. Mol. Sci. 2020, 21, 6021. [Google Scholar] [CrossRef]
- Do, M.T.; Na, M.; Kim, H.G.; Khanal, T.; Choi, J.H.; Jin, S.W.; Oh, S.H.; Hwang, I.H.; Chung, Y.C.; Kim, H.S. Ilimaquinone Induces Death Receptor Expression and Sensitizes Human Colon Cancer Cells to TRAIL-Induced Apoptosis through Activation of ROS-ERK/P38 MAPK–CHOP Signaling Pathways. Food Chem. Toxicol. 2014, 71, 51–59. [Google Scholar] [CrossRef]
- Carpi, S.; Scoditti, E.; Polini, B.; Brogi, S.; Calderone, V.; Proksch, P.; Ebada, S.S.; Nieri, P. Pro-Apoptotic Activity of the Marine Sponge Dactylospongia Elegans Metabolites Pelorol and 5-Epi-Ilimaquinone on Human 501Mel Melanoma Cells. Mar. Drugs 2022, 20, 427. [Google Scholar] [CrossRef]
- Cetraro, P.; Plaza-Diaz, J.; MacKenzie, A.; Abadía-Molina, F. A Review of the Current Impact of Inhibitors of Apoptosis Proteins and Their Repression in Cancer. Cancers 2022, 14, 1671. [Google Scholar] [CrossRef]
- Ketelut-Carneiro, N.; Fitzgerald, K.A. Apoptosis, Pyroptosis, and Necroptosis—Oh My! The Many Ways a Cell Can Die. J. Mol. Biol. 2022, 434, 167378. [Google Scholar] [CrossRef]
- Liu, Z.-B.; Zhang, T.; Ye, X.; Liu, Z.-Q.; Sun, X.; Zhang, L.-L.; Wu, C.-J. Natural Substances Derived from Herbs or Plants Are Promising Sources of Anticancer Agents against Colorectal Cancer via Triggering Apoptosis. J. Pharm. Pharmacol. 2022, 74, 162–178. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Papavassiliou, A.G. Apoptosis: The Suicide Solution in Cancer Treatment and Chemoprevention. Expert Opin. Investig. Drugs 2006, 15, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.; Mann, V.; Gulabivala, K. A Prospective Study of the Factors Affecting Outcomes of Nonsurgical Root Canal Treatment: Part 1: Periapical Health. Int. Endod. J. 2011, 44, 583–609. [Google Scholar] [CrossRef] [PubMed]
- Poon, I.K.H.; Hulett, M.D.; Parish, C.R. Molecular Mechanisms of Late Apoptotic/Necrotic Cell Clearance. Cell Death Differ. 2010, 17, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Dartsch, D.C.; Schaefer, A.; Boldt, S.; Kolch, W.; Marquardt, H. Comparison of Anthracycline-Induced Death of Human Leukemia Cells: Programmed Cell Death versus Necrosis. Apoptosis 2002, 7, 537–548. [Google Scholar] [CrossRef]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, New and Emerging Functions of Caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef]
- Kitazumi, I.; Tsukahara, M. Regulation of DNA Fragmentation: The Role of Caspases and Phosphorylation. FEBS J. 2011, 278, 427–441. [Google Scholar] [CrossRef]
- Ashkenazi, A.; Dixit, V.M. Death Receptors: Signaling and Modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef]
- Debatin, K.-M.; Krammer, P.H. Death Receptors in Chemotherapy and Cancer. Oncogene 2004, 23, 2950–2966. [Google Scholar] [CrossRef]
- Hakem, R.; Hakem, A.; Duncan, G.S.; Henderson, J.T.; Woo, M.; Soengas, M.S.; Elia, A.; De La Pompa, J.L.; Kagi, D.; Khoo, W. Differential Requirement for Caspase 9 in Apoptotic Pathways in Vivo. Cell 1998, 94, 339–352. [Google Scholar] [CrossRef]
- Green, D.R.; Evan, G.I. A Matter of Life and Death. Cancer Cell 2002, 1, 19–30. [Google Scholar] [CrossRef]
- Heath-Engel, H.M.; Shore, G.C. Mitochondrial Membrane Dynamics, Cristae Remodelling and Apoptosis. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 549–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, K.; Brown, D.G.; Langlais, C.; Cohen, G.M. Caspase Activation Involves the Formation of the Aposome, a Large (∼ 700 KDa) Caspase-Activating Complex. J. Biol. Chem. 1999, 274, 22686–22692. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Li, Y.; Liu, X.; Wang, X. An APAF-1· Cytochrome c Multimeric Complex Is a Functional Apoptosome That Activates Procaspase-9. J. Biol. Chem. 1999, 274, 11549–11556. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.S.; Pati, S.P.; Kumar, P.P.; Pradeep, H.N.; Sastry, G.N. Virtual Screening in Drug Discovery—A Computational Perspective. Curr. Protein Pept. Sci. 2007, 8, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.P.; Rees, S.; Kalindjian, S.B.; Philpott, K.L. Principles of Early Drug Discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef]
- Bishayee, A.; Dhir, N. Resveratrol-Mediated Chemoprevention of Diethylnitrosamine-Initiated Hepatocarcinogenesis: Inhibition of Cell Proliferation and Induction of Apoptosis. Chem. Biol. Interact. 2009, 179, 131–144. [Google Scholar] [CrossRef]
- Bishayee, A.; Mbimba, T.; Thoppil, R.J.; Háznagy-Radnai, E.; Sipos, P.; Darvesh, A.S.; Folkesson, H.G.; Hohmann, J. Anthocyanin-Rich Black Currant (Ribes nigrum L.) Extract Affords Chemoprevention against Diethylnitrosamine-Induced Hepatocellular Carcinogenesis in Rats. J. Nutr. Biochem. 2011, 22, 1035–1046. [Google Scholar] [CrossRef]
- Bhatia, D.; Thoppil, R.J.; Mandal, A.; Samtani, K.A.; Darvesh, A.S.; Bishayee, A. Pomegranate Bioactive Constituents Suppress Cell Proliferation and Induce Apoptosis in an Experimental Model of Hepatocellular Carcinoma: Role of Wnt/β-Catenin Signaling Pathway. Evid.-Based Complement. Altern. Med. 2013, 2013, 371813. [Google Scholar] [CrossRef]
- Haridas, V.; Higuchi, M.; Jayatilake, G.S.; Bailey, D.; Mujoo, K.; Blake, M.E.; Arntzen, C.J.; Gutterman, J.U. Avicins: Triterpenoid Saponins from Acacia Victoriae (Bentham) Induce Apoptosis by Mitochondrial Perturbation. Proc. Natl. Acad. Sci. USA 2001, 98, 5821–5826. [Google Scholar] [CrossRef]
- Mohs, R.C.; Greig, N.H. Alzheimer’s &. Dement. Transl. Res. Clin. Interv. 2017, 3, 651–657. [Google Scholar]
- Cui, W.; Aouidate, A.; Wang, S.; Yu, Q.; Li, Y.; Yuan, S. Discovering Anti-Cancer Drugs via Computational Methods. Front. Pharmacol. 2020, 11, 733. [Google Scholar] [CrossRef] [PubMed]
- Tewari, M.; Quan, L.T.; O’Rourke, K.; Desnoyers, S.; Zeng, Z.; Beidler, D.R.; Poirier, G.G.; Salvesen, G.S.; Dixit, V.M. Yama/CPP32β, a Mammalian Homolog of CED-3, Is a CrmA-Inhibitable Protease That Cleaves the Death Substrate Poly (ADP-Ribose) Polymerase. Cell 1995, 81, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Zaremba, T.; Thomas, H.; Cole, M.; Plummer, E.R.; Curtin, N.J. Doxorubicin-Induced Suppression of Poly (ADP-Ribose) Polymerase-1 (PARP-1) Activity and Expression and Its Implication for PARP Inhibitors in Clinical Trials. Cancer Chemother. Pharmacol. 2010, 66, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Elasbali, A.M.; Al-Soud, W.A.; Al-Oanzi, Z.H.; Qanash, H.; Alharbi, B.; Binsaleh, N.K.; Alreshidi, M.; Patel, M.; Adnan, M. Cytotoxic Activity, Cell Cycle Inhibition, and Apoptosis-Inducing Potential of Athyrium Hohenackerianum (Lady Fern) with Its Phytochemical Profiling. Evid.-Based Complement. Altern. Med. 2022, 2022, 2055773. [Google Scholar] [CrossRef] [PubMed]
- Chelliah, R.; Oh, D.-H. Screening for Anticancer Activity: Dual Staining Method. In Methods in Actinobacteriology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 427–429. [Google Scholar]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutellingsperger, C. A Novel Assay for Apoptosis Flow Cytometric Detection of Phosphatidylserine Expression on Early Apoptotic Cells Using Fluorescein Labelled Annexin V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Jiang, A.-J.; Jiang, G.; Li, L.-T.; Zheng, J.-N. Curcumin Induces Apoptosis through Mitochondrial Pathway and Caspases Activation in Human Melanoma Cells. Mol. Biol. Rep. 2015, 42, 267–275. [Google Scholar] [CrossRef]
- Rahman, H.S.; Rasedee, A.; Abdul, A.B.; Zeenathul, N.A.; Othman, H.H.; Yeap, S.K.; How, C.W.; Hafiza, W.A.G.W.N. Zerumbone-Loaded Nanostructured Lipid Carrier Induces G2/M Cell Cycle Arrest and Apoptosis via Mitochondrial Pathway in a Human Lymphoblastic Leukemia Cell Line. Int. J. Nanomed. 2014, 9, 527. [Google Scholar]
- Ma, W.; Zhao, L.; Yuan, L.; Yu, H.; Wang, H.; Gong, X.; Wei, F.; Xiao, R. Elaidic Acid Induces Cell Apoptosis through Induction of ROS Accumulation and Endoplasmic Reticulum Stress in SH-SY5Y Cells. Mol. Med. Rep. 2017, 16, 9337–9346. [Google Scholar] [CrossRef]
- Abutaha, N.; Nasr, F.A.; Al-Zharani, M.; Alqahtani, A.S.; Noman, O.M.; Mubarak, M.; Abdelhabib, S.; Wadaan, M.A. Effects of Hexane Root Extract of Ferula Hermonis Boiss. On Human Breast and Colon Cancer Cells: An In Vitro and In Vivo Study. Biomed Res. Int. 2019, 2019, 3079895. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open Chemical Toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, K.D.; Barage, S.H. Structural Analysis of Membrane-Bound HECE-1 Dimer Using Molecular Modeling Techniques: Insights into Conformational Changes and Aβ1–42 Peptide Binding. Amino Acids 2015, 47, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Parulekar, R.S.; Sonawane, K.D. Molecular Modeling Studies to Explore the Binding Affinity of Virtually Screened Inhibitor toward Different Aminoglycoside Kinases from Diverse MDR Strains. J. Cell. Biochem. 2018, 119, 2679–2695. [Google Scholar] [CrossRef] [PubMed]
- Nadaf, N.H.; Parulekar, R.S.; Patil, R.S.; Gade, T.K.; Momin, A.A.; Waghmare, S.R.; Dhanavade, M.J.; Arvindekar, A.U.; Sonawane, K.D. Biofilm Inhibition Mechanism from Extract of Hymenocallis Littoralis Leaves. J. Ethnopharmacol. 2018, 222, 121–132. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | 5I9B | 1CX2 | 2AR9 | 4S0O |
|---|---|---|---|---|
| Ilimaquinone | −6.9 | −8.9 | −8.1 | −8.2 |
| Receptor–Ligand | Receptor–Ligand Interactions | Distance in Angstroms |
|---|---|---|
| 2AR9–Ilimaquinone | (B:LYS272) HZ1-O (Ligand) | 2.50 |
| (B:LYS272) HZ3-O (Ligand) | 2.53 | |
| (B:LYS272) HZ3-O (Ligand) | 2.75 | |
| (B:LYS272) CE-O (Ligand) | 3.51 | |
| (A:GLU240) OE1-C (Ligand) | 3.64 | |
| (A:GLU240) OE1-Pi Anion interaction | 3.41 | |
| (A:LEU4) Alkyl–Alkyl interaction | 4.49 | |
| (B:ALA3) Alkyl–Alkyl interaction | 4.74 | |
| (B:ALA3) Alky–C Alkyl interaction | 3.94 | |
| (A:LEU4) Alkyl–C Alkyl interaction | 4.23 | |
| (A:LEU7) Alkyl–C Alkyl interaction | 3.99 | |
| (B:LEU7) Pi-Alkyl interection | 4.77 | |
| 1CX2–Ilimaquinone | (ASN7) HD21-O (Ligand) | 2.56 |
| (ALA125) HN-O (Ligand) | 2.35 | |
| (PRO123) O-H (Ligand) | 1.67 | |
| (VAL124) CA-O (Ligand) | 3.57 | |
| (PRO123) Alkyl–Alkyl interaction | 5.22 | |
| (ALA125) Alkyl–Alkyl interaction | 4.27 | |
| (ALA125) Alkyl–C Alkyl interaction | 3.86 | |
| (MET16) Alkyl–C Alkyl interaction | 4.69 | |
| (TYR105) Pi-Alkyl interaction | 4.06 | |
| (CYS4) Pi-Alkyl interaction | 5.02 | |
| (PRO122) Pi-Alkyl interaction | 4.67 | |
| 4S0O–Ilimaquinone | (ALA42) HN-O (Ligand) | 2.44 |
| (ALA42) HN-O | 2.25 | |
| (LEU47) HN-O | 2.27 | |
| (GLU41) CA-O | 3.61 | |
| (ALA42) CA-O | 3.57 | |
| (ALA46) CA-O | 3.54 | |
| (LEU45) O-C | 3.29 | |
| (ILE133) CD1 Pi Sigma interaction | 3.50 | |
| (ILE133) Alkyl–Alkyl interaction | 4.88 | |
| (LEU125) Alkyl–C Alkyl interaction | 4.78 | |
| (ILE33) Alkyl–C Alkyl interaction | 5.02 | |
| (ILE31) Alkyl–C Alkyl interaction | 3.90 | |
| (ALA46) Pi-Alkyl interaction | 4.47 | |
| 5I9B–Ilimaquinone | (ARG207) NH1-Pi interaction | 4.41 |
| (CYS163) SG-Pi interaction | 3.70 | |
| (TRP206) Pi-Alkyl interaction | 5.06 | |
| (TRP206) Pi-C Alkyl interaction | 4.65 | |
| (PHE256) Pi-Alkyl interaction | 4.11 | |
| (PHE256) Pi-C Alkyl interaction | 3.87 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | 5′ CATGGGGAAGGTGAAGGTCGA 3′ | 5′ TTGGCTCCCCCCTGCAAATGAG 3′ |
| Bcl-2 | 5′ TTCGATCAGGAAGGCTAGAGTT 3′ | 5′ TCGGTCTCCTAAAAGCAGGC 3′ |
| Caspase-3 | 5′ TGCGCTGCTCTGCCTTCT 3′ | 5′ CCATGGGTAGCAGCTCCTTC 3′ |
| Caspase-9 | 5′ CCAGAGATTCGCAAACCAGAGG 3′ | 5′ GAGCACCGACATCACCAAATCC 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surti, M.; Patel, M.; Redhwan, A.; Al-Keridis, L.A.; Adnan, M.; Alshammari, N.; Reddy, M.N. Ilimaquinone (Marine Sponge Metabolite) Induces Apoptosis in HCT-116 Human Colorectal Carcinoma Cells via Mitochondrial-Mediated Apoptosis Pathway. Mar. Drugs 2022, 20, 582. https://doi.org/10.3390/md20090582
Surti M, Patel M, Redhwan A, Al-Keridis LA, Adnan M, Alshammari N, Reddy MN. Ilimaquinone (Marine Sponge Metabolite) Induces Apoptosis in HCT-116 Human Colorectal Carcinoma Cells via Mitochondrial-Mediated Apoptosis Pathway. Marine Drugs. 2022; 20(9):582. https://doi.org/10.3390/md20090582
Chicago/Turabian StyleSurti, Malvi, Mitesh Patel, Alya Redhwan, Lamya Ahmed Al-Keridis, Mohd Adnan, Nawaf Alshammari, and Mandadi Narsimha Reddy. 2022. "Ilimaquinone (Marine Sponge Metabolite) Induces Apoptosis in HCT-116 Human Colorectal Carcinoma Cells via Mitochondrial-Mediated Apoptosis Pathway" Marine Drugs 20, no. 9: 582. https://doi.org/10.3390/md20090582
APA StyleSurti, M., Patel, M., Redhwan, A., Al-Keridis, L. A., Adnan, M., Alshammari, N., & Reddy, M. N. (2022). Ilimaquinone (Marine Sponge Metabolite) Induces Apoptosis in HCT-116 Human Colorectal Carcinoma Cells via Mitochondrial-Mediated Apoptosis Pathway. Marine Drugs, 20(9), 582. https://doi.org/10.3390/md20090582