The total FA composition in corals stems from a blend of the host organism’s FAs, FAs from intercellular symbiotic microalgae (when present), and associated organisms such as bacteria, filamentous algae, sponges, and marine fungi. This FA composition is shaped by several factors: the specific attributes of the host organism’s FA biosynthesis, the quantity of zooxanthellae, the composition and volume of associated organisms, the FA composition of the food source, environmental influences, and the coral’s developmental stage. Many members of the symbiotic coral association, as well as some coral food sources, possess unique FAs that can serve as markers for these organisms.
4.3.1. Marker Lipids and FAs of Zooxanthellae and the Host in Corals
The composition of total lipids and FAs has been determined for numerous coral species [
68,
208,
209]. However, there are limited data on the composition of lipids and FAs of zooxanthellae extracted directly from corals [
101,
112,
115]. Only a few studies, such as those by Papina and colleagues [
116], Treignier and team [
121], and Imbs and associates [
134], have concurrently identified the FA composition of both components (algae and animal) from the same hard coral colony. The initial data regarding the FA composition of pure zooxanthellae from soft corals have only recently been gathered [
69,
134].
Although the FA compositions of pure zooxanthellae vary depending on their source (see
Tables S38–S43), a comparative analysis reveals certain FAs typical for all symbiotic dinoflagellates. These can be viewed as markers for zooxanthellae.
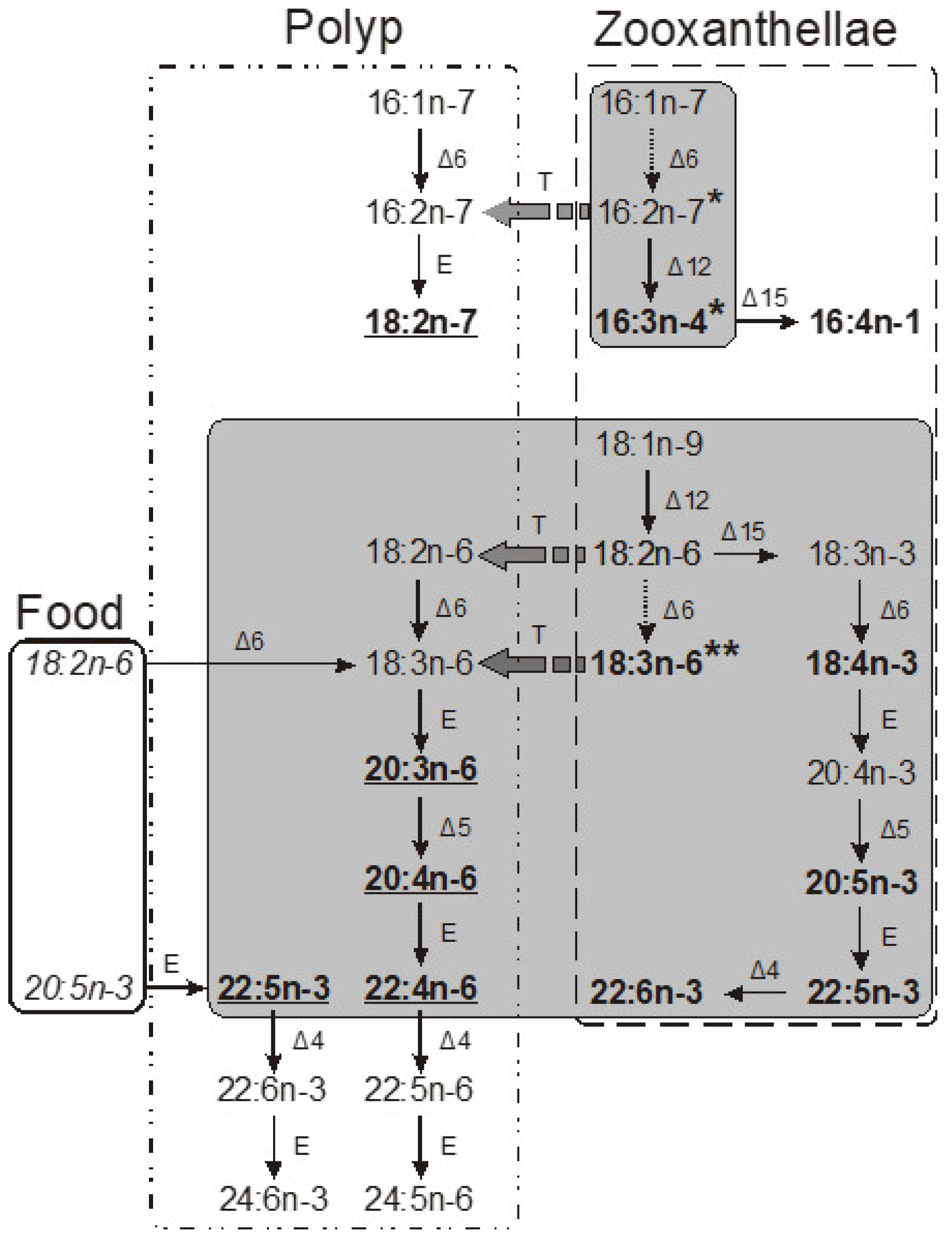
A universally recognized marker for zooxanthellae is 18:4n-3 (octadeca-6,9,12,15-tetraenoic acid). Found in significant amounts in zooxanthellae lipids from hydrocorals and hard and soft corals, this PUFA is predominantly present in the glycolipids of zooxanthellae. These glycolipids are essential for the functioning of the thylakoid membranes in the photosynthetic machinery of zooxanthellae [
118]. In hard coral zooxanthellae, the content of 18:4n-3 in total lipids and glycolipids can reach up to 15% and 40% of total FAs, respectively. For hydrocorals from the genus
Millepora and soft corals from the
Sinularia genus, the percentages are around 17% and 13%, respectively. Among ten strains of zooxanthellae cultured in labs, the 18:4n-3 content varied between 6 and 30%, averaging around 15%. In the case of the hard coral
Stylophora subseriata and the surrounding zooplankton, 18:4n-3 made up 12.7%, 6.6%, and 1.4% of their total FAs, respectively [
38], suggesting that zooxanthellae are the primary source of 18:4n-3 in
S. subseriata.
Almost all of the 18:4n-3 is localized within the zooxanthellae lipids. The proportion of 18:4n-3 in polyp tissues is significantly reduced, being 15–20 times lower than in zooxanthellae. Hence, by determining the percentage of 18:4n-3 in the lipids of entire coral colonies, we can estimate a crucial aspect: the proportion of zooxanthellae lipids in the total lipids of the colony. This factor represents the extent of the influence of zooxanthellae on the coral’s lipid composition. This is crucial for discerning coral chemotaxonomy via lipid markers and also vital when investigating coral bleaching and nutrition. However, it is worth noting that the 18:4n-3 concentration in the zooxanthellae is variable, making the assessment of the zooxanthellae lipid portion only an approximation.
The 18:4n-3 concentration in whole colonies of both hard corals and the soft corals from the Sinularia genus typically ranges between 2 and 4% of total FAs. The mean concentration of 18:4n-3 in the zooxanthellae FAs of these corals stands at around 15%. A rough calculation indicates that the zooxanthellae lipids account for roughly 20% of the coral’s total lipids. Interestingly, the 18:4n-3 content in entire colonies of soft coral species is roughly double that found in whole hard coral colonies. Nevertheless, the absence of data on the 18:4n-3 concentration in pure zooxanthellae from these soft coral species prevents us from precisely determining the zooxanthellae lipid portion.
Azooxanthellate corals’ total FA contains up to 0.5% of 18:4n-3, suggesting that zooxanthellae are not the sole source of 18:4n-3. This fatty acid might also be sourced from free-living dinoflagellates or algae that are associated with the coral colonies.
The acid octadeca-6,9,12-trienoic (18:3n-6) can be recognized as a marker for hard coral zooxanthellae [
68]. In pure zooxanthellae from hard corals, its content in total lipids varied between 5 and 23%, averaging about 10% of total FA. For the FA of zooxanthellae glycolipids, it averaged 13%. Cultivated zooxanthellae strains had just 2–4% of 18:3n-6 [
118]. The 18:3n-6 content in hard coral polyp tissues was considerably higher (2–4% of total FA) than that of 18:4n-3. This indicates a possible transfer of this PUFA from the zooxanthellae to the host or an uptake of 18:3n-6 by the coral from food, or its own biosynthesis from 18:2n-6. Consequently, accurately determining zooxanthellae lipid shares using 18:3n-6 content, similar to the approach with 18:4n-3, is not feasible.
The fatty acid 18:3n-6 was virtually absent in both entire colonies and pure zooxanthellae fractions and polyp tissues of the Millepora genus hydrocorals.
For zooxanthellate hard corals, the combined amounts of the marker PUFAs 18:4n-3 and 18:3n-6 fluctuated based on the coral’s taxonomic classification and environmental conditions. Even partial bleaching (loss of zooxanthellae) led to a notable reduction in the concentration of these markers in the coral’s total FAs. In the hermatypic coral
Pavona frondifera, a reduction in zooxanthellae led to a drop in the combined content of 18:3n-6 and 18:4n-3 from 12.2% to 1.8% [
210]. In comparison, healthy symbiotic hard corals had these acids ranging from 2 to 15% of their total FA [
68,
100,
103]. The levels of 18:3n-6 and 18:4n-3 were minimal in non-symbiotic alcyonarians [
79], gorgonians [
111], and certain hard corals [
68,
100].
In summary, if the sum of 18:3n-6 and 18:4n-3 exceeds 2% of a coral colony’s total FAs, it can indicate the presence of zooxanthellae within that coral. The 18:3n-6 content in zooxanthellae extracted from various
Sinularia species was minimal. Notably, an inverse correlation between 18:3n-6 and 16:2n-7 contents was identified in soft corals, particularly within the
Sinularia genus [
68]. Some
Sinularia species exhibited a low concentration of 18:3n-6 (0.5%) and a high concentration of 16:2n-7 (8.1%). In contrast, other
Sinularia species showed a low concentration of 16:2n-7 (0.4%) and a high concentration of 18:3n-6 (6.4%). Those specific
Sinularia colonies from which pure zooxanthellae were extracted had a very minimal amount of 18:3n-6. Consequently, their zooxanthellae naturally lacked this marker acid. Nonetheless, it is plausible that zooxanthellae extracted from soft corals with a high concentration of 18:3n-6 might also possess a similar level of 18:3n-6 to that found in zooxanthellae from hard corals. Thus, the acid 18:3n-6 can be regarded as a zooxanthellae marker for alcyonarians from the genera
Carijoa,
Alcyonium,
Cladiella,
Lemnalia,
Nephthea, specific
Sinularia species, gorgonians from the
Hicksonella genus, and the blue coral
Heliopora coerulea.
Although all species within the
Lobophytum and
Sarcophyton genera contain zooxanthellae, the 18:3n-6 content in these soft corals was low. As a result, 18:3n-6 should not be considered a definitive lipid marker for zooxanthellae in soft corals from these two genera. The two C16 PUFAs, 16:3n-4 (hexadeca-6,9,12-trienoic acid) and 16:4n-1 (hexadeca-6,9,12,15-tetraenoic acid), are indicative of soft coral zooxanthellae [
69,
134]. The biosynthesis of C16 PUFAs (16:2n-7 → 16:3n-4 → 16:4n-1) is restricted in hard coral zooxanthellae and represents a primary biochemical trait of soft coral zooxanthellae. On average, the total content of 16:3n-4 and 16:4n-1 in pure zooxanthellae lipids from
Sinularia genus soft corals constituted 4.3% and 3.2% of total FA, respectively. The ratio of these markers between zooxanthellae and polyp tissues was between 7 and 10. Unlike plants, animals lack Δ12 and Δ15 desaturases, meaning they cannot produce 16:3n-4 and 16:4n-1 independently. Primary sources of these PUFAs in soft coral polyp tissues, aside from zooxanthellae, are likely plant-based foods such as phytoplankton.
Certain dinoflagellate FAs, such as 20:5n-3 and 22:6n-3, were notably more abundant in pure zooxanthellae lipids compared to the lipids of an entire coral colony. While these PUFAs are characteristic of zooxanthellae, their use as definitive lipid markers presents challenges. For instance, the content of 22:6n-3 in zooxanthellae lipids, whole
Stylophora subseriata hard coral colonies, and zooplankton collected from the same region showed little variation [
38]. In addition to obtaining 20:5n-3 and 22:6n-3 from zooxanthellae, corals also acquire these FAs from phyto- and zooplankton lipids, which form a significant portion of their diet. Currently, methods to differentiate between autotrophic and heterotrophic lipid sources are still in development [
211]. Likely, a combined approach using both lipid and isotope marker techniques will be necessary to ascertain the amount of 20:5n-3 and 22:6n-3 introduced into corals via zooxanthellae.
Initially, in total lipids of hard corals of pure zooxanthellae, the 18:5n-3 acid was found in quantities ranging from 2 to 8%, and up to 24% in zooxanthellae glycolipid FA [
112]. Subsequent studies confirmed the presence of this acid in zooxanthellae FA composition [
115,
116,
117].
Millepora polar lipid FA had an 18:5n-3 content of 8.7% [
115]. As 18:5n-3 is distinctively indicative of dinoflagellates and is nearly absent in other sources, it offers potential as a unique zooxanthellae lipid marker. However, many studies on dinoflagellate FA composition used alkaline conditions to extract FA from total lipids, resulting in the degradation of the fragile 18:5n-3 acid and leading to inaccurate measurements of its content [
192].
The processes of PUFA biosynthesis in animals and plants differ. Typically, animal desaturases cannot insert a double bond beyond the C-9 atom of the FA molecule’s carbon chain, meaning animals cannot produce 18:2n-6 and 18:3n-3 [
210]. This is a primary reason for the distinction between the PUFA compositions of endosymbiotic dinoflagellates (plants) and their hosts (animals). All previously mentioned zooxanthellae FAs as lipid markers are either n-3 series PUFAs or products of specific Δ12 and Δ15 plant desaturases. Animals typically exhibit a dominance of n-6 series PUFAs. Some of these PUFAs, such as 20:4n-6 (eicosa-5,8,11,14-tetraenoic acid) and 22:4n-6 (docosa-7,10,13,16-tetraenoic acid), which are concentrated in polyp tissues, have been suggested as marker indicators for the host organism in hard corals [
70]. Many studies have found that the percentages of 20:4n-6 and 22:4n-6 in the hard coral host organism are 2–3 times greater than those in zooxanthellae. In some hard coral species, the 22:4n-6 content constituted over 20% of the total FAs. Hashimoto and co-workers [
120] observed that pure zooxanthellae from the soft coral
Clavularia viridis lacked arachidonic acid (20:4n-6) entirely. Yet, for soft corals of the
Sinularia genus [
69,
70], no notable difference was found in 20:4n-6 content between polyps and zooxanthellae, and the 22:4n-6 content was less than 1% of the total FAs.
Very-long-chain fatty acids, specifically 24:5n-6 (tetracosa-6,9,12,15,18-pentaenoic acid) and 24:6n-3 (tetracosa-6,9,12,15,18,21-hexaenoic acid), serve as distinctive markers for the hosts of soft corals. These PUFAs act as chemotaxonomic indicators for octocorals. Tetracosapolyenoic acids (TPAs) have been detected in both zooxanthellate and azooxanthellate soft corals. However, the concentrations of these PUFAs are notably higher in non-symbiotic species compared to their zooxanthellate counterparts. This suggests that the biosynthesis of 24:5n-6 and 24:6n-3 occurs within the polyp tissues, and the zooxanthellae do not contribute to this process [
92].
The fatty acid eicosa-4,7,10,13,16-pentaenoic acid (22:5n-6) has been suggested as a host marker for hydrocorals of the genus Millepora. This acid is found in minimal amounts in both hard and soft corals. The complexity arises from the similar percentages of 22:4n-6 in both zooxanthellae and hydrocoral polyp tissues. It is theorized that the biosynthesis of n-6 series PUFAs occurs within the host tissues of Millepora, following the sequence 18:3n-6 → 20:3n-6 → 20:4n-6 → 22:4n-6 → 22:5n-6. The low concentrations of intermediates (18:3n-6, 20:3n-6, and 20:4n-6) suggest a rapid biosynthesis pathway in Millepora. There is a possibility that 22:5n-6 gets transferred from the host to the zooxanthellae. More research is essential to understand the biosynthesis and transfer processes of 22:5n-6 in Millepora.
Galactolipids, specifically MGDG and DGDG, which are typical of plant photosynthetic apparatus biomembranes, should be recognized as distinct lipid markers of zooxanthellae in the symbiotic coral community. The PUFAs of these galactolipids primarily mark zooxanthellae [
112]. The absence of galactolipids in the polar lipids of polyp tissue fractions from the soft coral
Clavularia viridis has been cited as proof of this fraction’s high purity [
120]. It is likely that Forssman antigen-like glycosphingolipids (gangliosides) of zooxanthellae have a significant role in the symbiosis between corals and Symbiodinium. The lectin SLL-2, extracted from the octocoral
Sinularia lochmodes, attaches to the glycosphingolipids on zooxanthellae’s surface. Consequently, Symbiodinium cells transition from a flagellated swimming form to a non-flagellated coccoid form, which resembles the symbiotic stage observed in corals [
212].
Another unique marker, MADAG, was found in soft coral host lipids. This marker was not present in zooxanthellae lipids but was identified in about 35% of the polyp tissue total lipids [
69]. This uncommon lipid class is typical for cnidarians [
43]. MADAG content is lower in hard corals compared to soft corals, with zooxanthellate coral species characterized by a higher MADAG content.
4.3.2. Marker FAs of Associated Organisms in Corals
In addition to symbiotic microalgae (zooxanthellae), coral colonies harbor a diverse group of associated organisms, including bacteria, filamentous algae, sea fungi, sponges, and various biofouling entities that often settle on the colony’s exterior. While these associated organisms do not establish true symbiotic relationships with the host coral polyps, their biological activities significantly influence the coral’s nutrition and the exchange of inorganic and organic substances between the corals and their environment. For instance, certain filamentous algae have been identified to form genuine symbiotic bonds with hard corals. The composition, both quantitative and qualitative, of these associated organisms tends to vary and is only somewhat influenced by the coral species. Much of the research on marine associates is qualitative, focusing primarily on the species composition of the identified flora and fauna. Chemical markers, particularly lipid markers, can be employed to determine the presence or absence of specific associates and to provide a rough estimate of their total abundance within coral colonies. Regrettably, research in this direction is in its infancy, and there is a dearth of data on the composition and utilization of lipid markers in coral-associated organisms.
A consistent bacterial community is found in most marine symbiotic animals, and corals are no exception. A comparative study of microbes in healthy and disease-afflicted coral colonies (specifically those affected by white and yellow spot diseases) of 11 coral species from the reefs of Nha Trang Bay, Vietnam, was conducted [
213]. This study identified 59 strains of heterotrophic bacteria in the mucus covering the coral colonies. The bacterial composition was not species specific relative to the examined corals and typically included 2–4 primary heterotrophic bacterial species. The fatty acid composition of these bacteria’s lipids was also characterized (refer to
Table S45).
Among the microbial composition of healthy corals, γ–Proteobacteria, notably halomonads, were predominant. Additionally, Gram-negative bacteria such as
Pseudomonas and
Vibrio spp., which are part of the Cytophaga–Flavobacterium–Bacteroides (CFB) phylogenetic cluster, were identified [
213]. The Gram-positive bacteria detected included species of
Bacillus,
Staphylococcus,
Halococcus,
Micrococcus, and coryneformic bacteria. The fatty acids (FAs) of these bacteria mainly consisted of saturated and monoenoic FAs, both with unbranched and branched carbon structures, ranging from 10 to 19 carbon atoms (see
Table S45). Hydroxy- and cyclopropane acids were also present. Primary FAs in the polar lipids of
Pseudomonas and
Halomonas were 16:0, 16:1n-7, and 18:1n-7. Pseudomonas specifically had cy-17:0.
Vibrio sp. was characterized by a content of over 40% of 16:1n-7. For the genera
Bacillus,
Planococcus, and
Micrococcus, the predominant FAs were ai-15:0 and ai-17:0.
Numerous studies on coral–microbe interactions have revealed a diverse microbial community residing on coral surfaces and potentially within coral tissues [
44,
190]. However, it remains uncertain whether these microbes play a specific role in coral biology or if the observed association merely represents an opportunistic collaboration between corals and marine water bacteria. Bacteria stand out from eukaryotes in their ability to synthesize a broad range of branched saturated and unsaturated FAs, as well as acids with an odd number of carbon atoms [
183]. They also produce 18:1n-7. Acids with an odd number of carbon atoms, branched acids, and 18:1n-7 are considered bacterial markers. These markers have been utilized to identify bacterial presence [
68].
In comparison to symbiotic coral species, the content of “bacterial” FAs was significantly higher (
p < 0.05) in non-symbiotic species, indicating a more robust bacterial community [
68]. A notable increase (
p < 0.01) in bacterial FA levels within the total FAs of non-symbiotic soft corals from the
Dendronephthya genus was observed [
92]. The bacterial FA content showed a significant difference (
p < 0.05) between non-symbiotic and symbiotic gorgonian corals (refer to
Figure 45). This indicates a more prominent bacterial community in azooxanthellate gorgonians [
111].
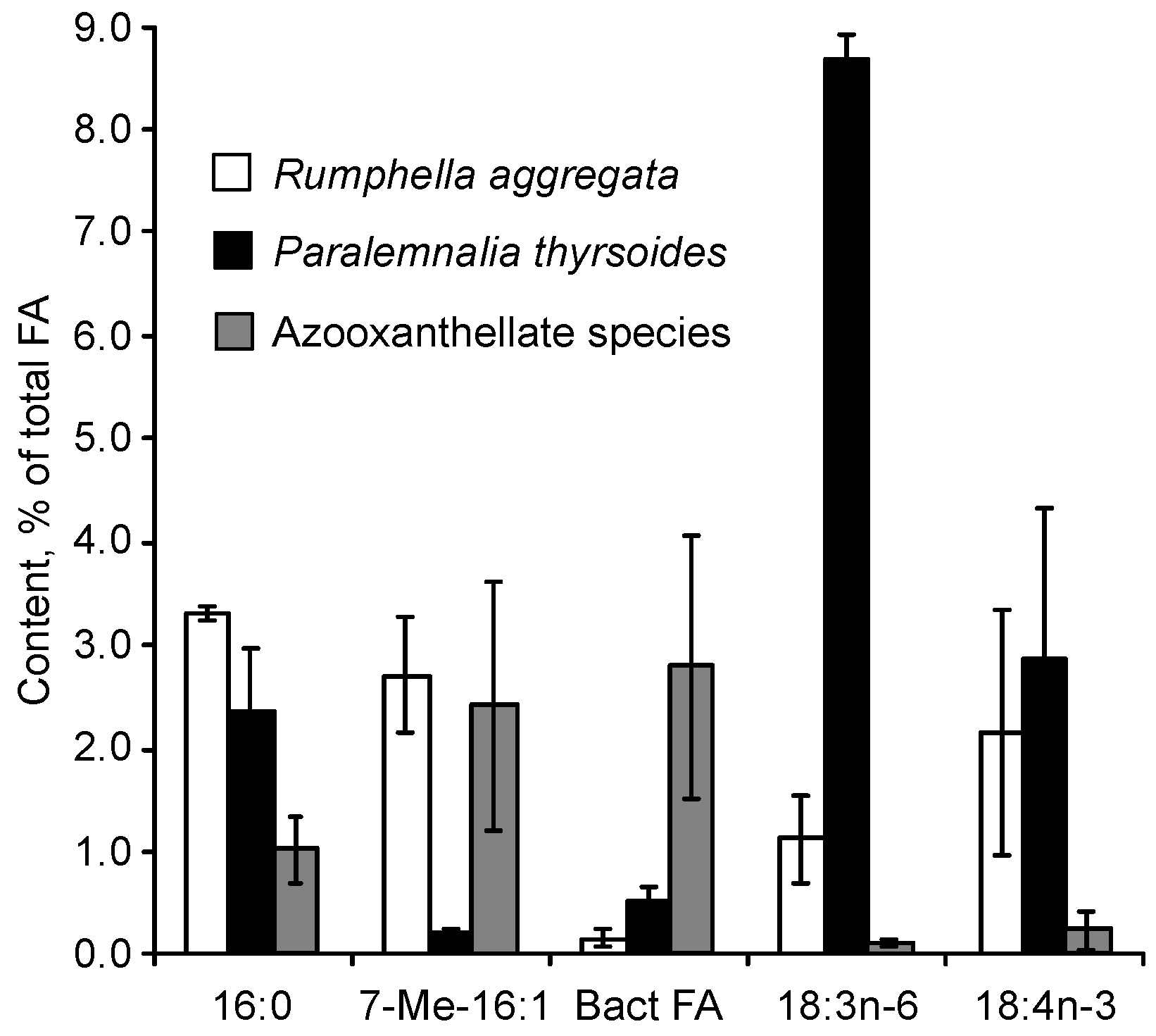
Another intriguing compound, the 7-methyl-6-hexadecenoic acid (7-Me-16:1n-10), was found in all examined soft corals. The concentration of this acid was significantly higher in azooxanthellate alcyonarians from the
Dendronephthya genus compared to other zooxanthellate alcyonarians (
p < 0.01) [
92]. In
Dendronephthya, the 7-Me-16:1n-10 content reached up to 7.3% of the total FAs. In contrast, the average content of 16:1n-10 was only 1.1 ± 0.7% across 13 zooxanthellate alcyonarian species. Moreover, the level of 7-Me-16:1n-10 was significantly higher (
p < 0.01) in non-symbiotic gorgonians at 2.4 ± 1.2%, compared to their symbiotic counterparts [
111]. On average, this FA constituted 2.9% of the polar lipid FAs in
Gorgonia corals [
45]. An increase in the 7-Me-16:1n-10 content was also observed in hard corals lacking zooxanthellae. The concentration of this FA was 2.3% of the total FAs in azooxanthellate species such as
Balanophyllia sp. and
Tubastrea aurea [
68], while it made up no more than 0.6% of the total FAs in zooxanthellate hard corals.
The fatty acid 7-Me-16:1n-10 has been identified in marine invertebrates such as sponges, actiniae, and gorgonians. Carballeira and colleagues [
45] suggested that either bacteria or zooxanthellae might produce this FA. However, a comparison between the fatty acids of zooxanthellate and azooxanthellate soft corals revealed that zooxanthellae likely are not the primary producers of 7-Me-16:1n-10. Notably, marine sponges, which harbor a significant bacterial population and exhibit high levels of 7-Me-16:1n-10 [
214,
215], do not host zooxanthellae. Consequently, bacteria appear to be the most likely source of 7-Me-16:1n-10 in soft corals.
Significant fluctuations in the levels of 7-Me-16:1n-10, and the presence of 2.7% of this acid in the total fatty acids of the symbiotic gorgonian
R. aggregate (referenced in
Figure 46), challenge the idea of a direct correlation between elevated 7-Me-16:1n-10 levels and the absence of zooxanthellae in corals [
111]. It is possible that 7-Me-16:1n-10 marks the presence not of the entire bacterial community, but of specific bacterial groups associated with corals [
92].
Zooxanthellae serve as the primary source of organic carbon for their host organisms. The rise in bacterial populations within azooxanthellate corals might represent an adaptive strategy to compensate for the absence of zooxanthellae, enabling the corals to obtain organic carbon from their associated bacteria [
92].
A unique group of furan acids (F-acids), which have potential as biomarkers, was identified in gorgonian corals [
111]. These acids appear to be widespread across various life forms, having been found in sea sponges, algae, crabs, other invertebrates, and even fish [
216]. Both higher plants and algae have been demonstrated to form the carbon skeleton of F-acids from linoleic acid (18:2n-6). Later stages see the elongation of F-acids with acetate residues to achieve their higher homologs. Several studies have reported the presence and biosynthesis of F-acids in phototrophic bacteria [
217,
218], which are common to many marine creatures. As such, the F-acids discovered in these animals are believed to originate either from marine bacteria or algae [
217]. F-acids with chain lengths of 18–22 carbon atoms (43–49) were identified in six azooxanthellate gorgonian coral species, with the exception of
A. isoxia [
111]. Their contribution to the total FA ranged between 0 and 9.7%. The same compounds were previously noted in the aforementioned biological entities [
216,
217,
218].
The source of F-acids in octocorals [
111] could be microalgae consumed as food or the microorganisms that live in association with corals, such as bacteria or other microalgae. We are inclined to support the idea that azooxanthellate soft corals form a temporary, non-specific association with phototrophic bacteria or certain microalgae. In this context, the quantity of these associated microorganisms within the coral colony might be estimated based on the levels of their markers, the F-acids.
Furthermore, the presence of C25–28 Δ5,9 unsaturated very-long-chain fatty acids (51–54) or demospongic acids was initially identified in a
Bebrice studeri sample from near Den Is. (Vietnam) and subsequently in other samples of the same species from Nha Trang Bay (Vietnam) [
111]. These acids are characteristic of sponge FAs [
219]. Excluding the aforementioned acids, demospongic acids were absent in all other coral species studied. Although the collected
Bebrice colonies were free from fouling, the existence of an endosymbiotic sponge from this genus had been previously documented [
220]. Consequently, the discovery of demospongic acids in the total FAs of
B. studeri gorgonians attests to the presence of a symbiotic sponge. These acids can serve as specific markers for the presence of symbiotic sponges in other coral species. Fatty acids comprise around 25% and 6% of the dry weight of corals and sponges, respectively. In
B. studeri, demospongic acids constituted 20.3 ± 4.5% of the total FAs. A rough calculation indicates that the symbiotic sponge could make up as much as 50% of the overall dry weight of the
B. studeri coral [
111,
214,
215].
4.3.3. Lipid and FA Transfer between Host Organism and Zooxanthellae in Corals
Corals are marine organisms, and many species obtain organic carbon derived from photosynthesis through their symbiotic relationship with intracellular dinoflagellates known as zooxanthellae [
221]. For zooxanthellate corals, phototrophic nutrition is a primary carbon source. In some of these coral species, over 90% of photosynthetic production can be relayed from zooxanthellae to the host cells [
222,
223]. In return, the host provides zooxanthellae with carbon dioxide and water for photosynthetic processes, as well as the necessary substrates for the synthesis of their cellular components, including the photosynthetic apparatus. The zooxanthellae produce low-molecular-weight compounds, such as glycerol, organic acids, glucose, and amino acids, which are then integrated into the coral tissues [
224]. A portion of the organic substances transferred from zooxanthellae to the host consists of lipids. It was initially believed that this transfer took place in the form of “lipid droplets”, observable under microscopic examination of zooxanthellate corals. However, later research indicated that these droplets contain minimal lipids [
222].
A study on
Pocillopora damicornis in laboratory tanks with seawater enriched with dissolved inorganic nitrogen (N) evaluated the rate at which zooxanthellae were lost [
225]. For the first two weeks, the lipid content in the N-enriched corals was comparable to that of the controls. However, after this period, it decreased to about two-thirds of the control levels, suggesting that N enrichment leads to a decreased rate of photosynthate transfer to the host.
The mechanism of lipid transfer between the members of the symbiotic community in corals remains an area of investigation. Some studies have indicated that carbon synthesized by zooxanthellae may be passed to the host not just as glycerol, but also in the form of acyl lipids. An analysis comparing the metabolism of labeled
14C-bicarbonate and 1-
14C-acetate in two hard coral species proposed that carbon, once photosynthetically fixed by zooxanthellae, is promptly used for lipid acyl residue synthesis before being transferred to the host organism [
33].
Short-term incubations of the zooxanthellate hard coral,
Stylophora pistillata, with
14C-bicarbonate were conducted [
226]. Zooxanthellae primarily incorporated
14C into a chloroform-methanol fraction rather than water-soluble compounds (88–94% vs. 4–7%). In contrast, the host displayed a greater preference for incorporating
14C into water-soluble compounds over the chloroform-methanol fraction (44–67% vs. 33–56%). In vivo, the average translocation rate, defined as the amount of
14C found in the host fraction as a percentage of the total
14C fixed, was 60.4%. This study affirms that the host takes up a portion of the photosynthetic products from its symbionts. However, the specifics of the compounds transferred, as well as the composition of lipids within the mixture of low-polar compounds in the chloroform-methanol fractions, remain elusive.
For the hard coral
Montipora digitata, there was documentation of lipid biosynthesis from
14C-glucose, which the zooxanthellae transferred to the host [
83]. Treignier and co-workers [
211] hypothesized that certain lipids are directly transferred from zooxanthellae to the host in the scleractinia
Turbinaria reniformis.
It was noted that the fatty acid (FA) profile of zooxanthellae is distinct from that of coral polyp tissues. When analyzing the overall FA composition of an entire coral colony, several marker FAs are detected in both the zooxanthellae and the host. However, the origins of these acids and the potential for their transfer between the zooxanthellae and the host are ambiguous. Meyers [
97,
98,
99] theorized that corals obtain saturated FAs from zooxanthellae and unsaturated FAs from external sources, such as zooplankton. Yet, the current literature lacks direct evidence showcasing the transfer of individual FAs from zooxanthellae to the host. Some studies [
116,
121] alluded to experiments that support the transport of both saturated and unsaturated FAs from zooxanthellae to the host in hard corals. However, these references do not offer experimental data directly evidencing the transfer of FAs from symbionts to the host within corals.
The marker FA method was employed to validate the hypothesis of PUFA transfer from zooxanthellae to the host. The aim was to identify the presence of distinct zooxanthellae marker PUFAs within host tissues. Several studies [
116,
121] have examined the FA composition of isolated zooxanthellae and pure polyp tissue from identical hard coral colonies. Notably, a modest quantity of PUFAs, specific to zooxanthellae, were identified within the FA profile of the host’s total lipids. PUFAs from the n-3 series, such as 18:4n-3, 22:5n-3, and 22:6n-3, constituted a minor fraction of the total FAs in polyp tissues from the hard coral
Montipora digitata [
116]. Of these, 18:4n-3 is especially significant as a primary zooxanthellae marker, given that corals could acquire 22:5n-3 and 22:6n-3 from dietary sources or through intrinsic biosynthesis. The ratio of 18:4n-3 percentages between pure zooxanthellae and
M. digitata polyp tissues was 3.92 (as per Papina and co-workers [
116]). PUFAs from the n-3 series, namely 18:4n-3, 20:5n-3, and 22:6n-3, were also identified in pure polyp tissues of the hard coral
Turbinaria reniformis [
121]. The percentage compositions of 18:4n-3 in the total FAs of pure zooxanthellae and polyp tissues from
T. reniformis were 7.9% and 1.4%, respectively. Given that these zooxanthellae marker PUFAs were detected in unadulterated polyp tissues, it is inferred that these PUFAs were sourced from symbionts.
A recent study [
122] simultaneously determined the FA compositions of total lipids from zooxanthellae and polyp tissues extracted from various hard corals:
Montipora faliosa,
M. digitata,
Pocillopora damicornis,
Acropora intermedia,
A. muricata,
Porites cylindrica, and
Pavona decussata, as well as the hydrocoral
Millepora platyphylla (as outlined in
Table S43). There were marked differences in the FA patterns between zooxanthellae and their hosts. Zooxanthellae predominantly contained n-3 series PUFAs, while polyp tissues were enriched in n-6 series PUFAs. Notably, the symbionts from all examined species demonstrated an elevated presence of 18:4n-3 (up to 29.5% in
P. decussata). Meanwhile, this marker acid was detectable in the polyp tissues of all analyzed cnidarian species, albeit at a considerably reduced proportion (not exceeding 1.3% of total FAs). Conversely, the proportion of 18:3n-6 in polyp tissues was slightly elevated (2–3% of total FAs) compared to the 18:4n-3 levels (as seen in
Table 23). Thus, the partition coefficient (the ratio of acid percentages in zooxanthellae to polyp tissues) for 18:4n-3 was substantially higher (ranging between 11.5 and 22.7) than that for 18:3n-6 (between 1.1 and 9.8). The heightened partition coefficient value for 18:4n-3 underscores the high purity of the zooxanthellae fraction.
The modest presence of zooxanthellae marker FAs in polyp tissues supports the hypothesis that these PUFAs are transferred from the symbionts to their hard coral hosts. It is worth highlighting that the concentration of 18:3n-6 in the zooxanthellae of M. platyphylla was notably low, and this acid was also absent in the polyp tissues of this hydrocoral.
When examining the distribution of zooxanthellae marker FAs between the symbionts and the host in soft corals from the genus
Sinularia, it was observed that 18:4n-3 was predominantly found in zooxanthellae, similar to its concentration in hard corals (as shown in
Table 24) [
69,
134]. Only a slight presence of 18:4n-3 (comprising 1.2–1.3% of total FAs) was detected in the soft coral host tissue. The partition coefficient for 18:4n-3 in Sinularia varied between 7.0 and 10.2. Meanwhile, the partition coefficients for two other marker acids, 16:3n-4 and 16:4n-1—both characteristic of soft coral zooxanthellae—were comparatively lower, ranging from 3.8 to 11.0. The 16:2n-7 concentration, which acts as a precursor to C16 PUFA in zooxanthellae, was relatively consistent in both the zooxanthellae and soft coral polyp tissues, with a partition coefficient between 1.2 and 1.5. Interestingly, the concentration of 18:2n-7, which derives from 16:2n-7, was found to be greater in the polyp tissues (as detailed in
Table 24).
The minor presence of zooxanthellae marker FAs in polyp tissues aligns with the hypothesis suggesting the transfer of PUFAs from symbionts to the host in soft corals. However, the proportion of these marker PUFAs in the total FAs of the host was relatively low for both soft and hard corals. It is plausible to assume that the presence of zooxanthellae PUFAs in the polyp tissues could be attributed to contamination by zooxanthellae lipids during polyp tissue extraction. Additionally, the host could intake C16 and C18 PUFAs from their food sources.
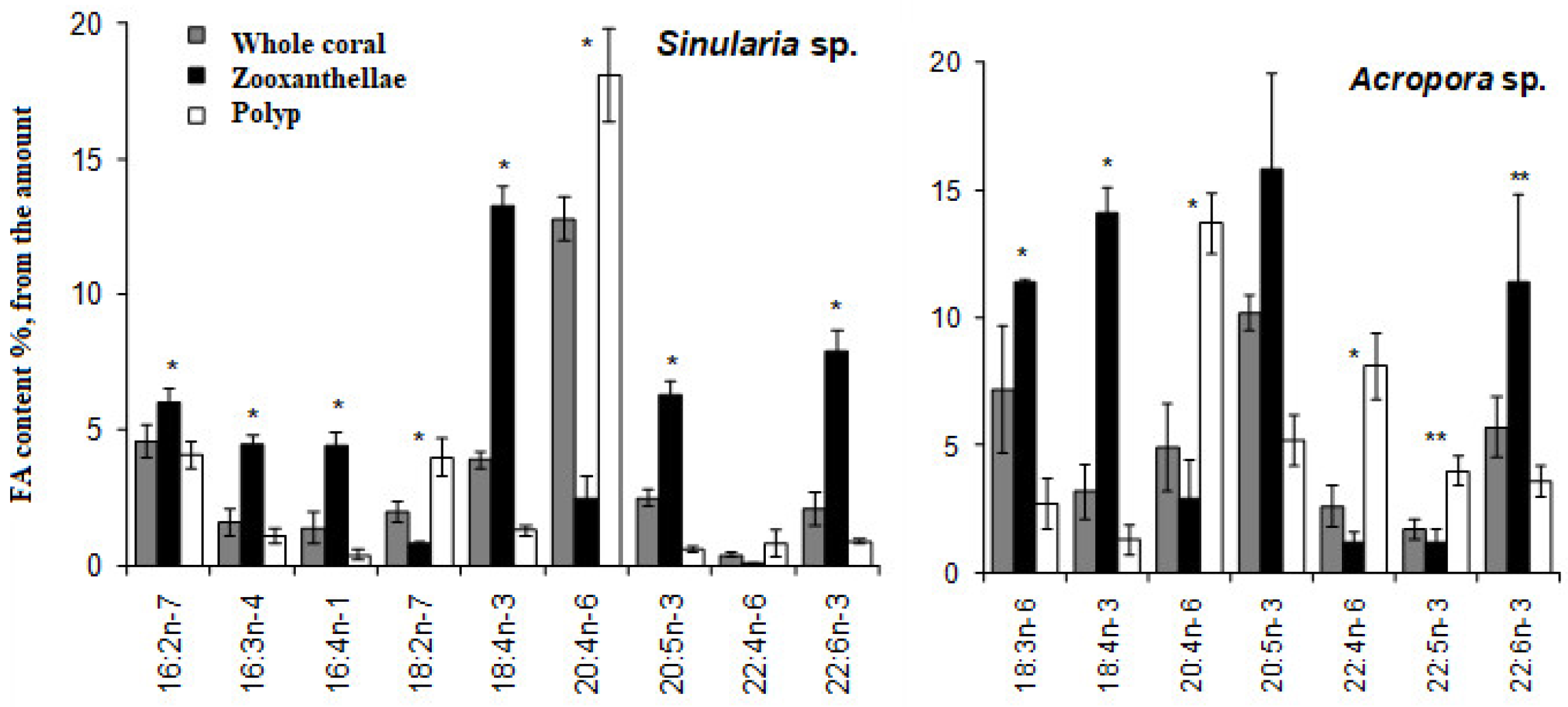
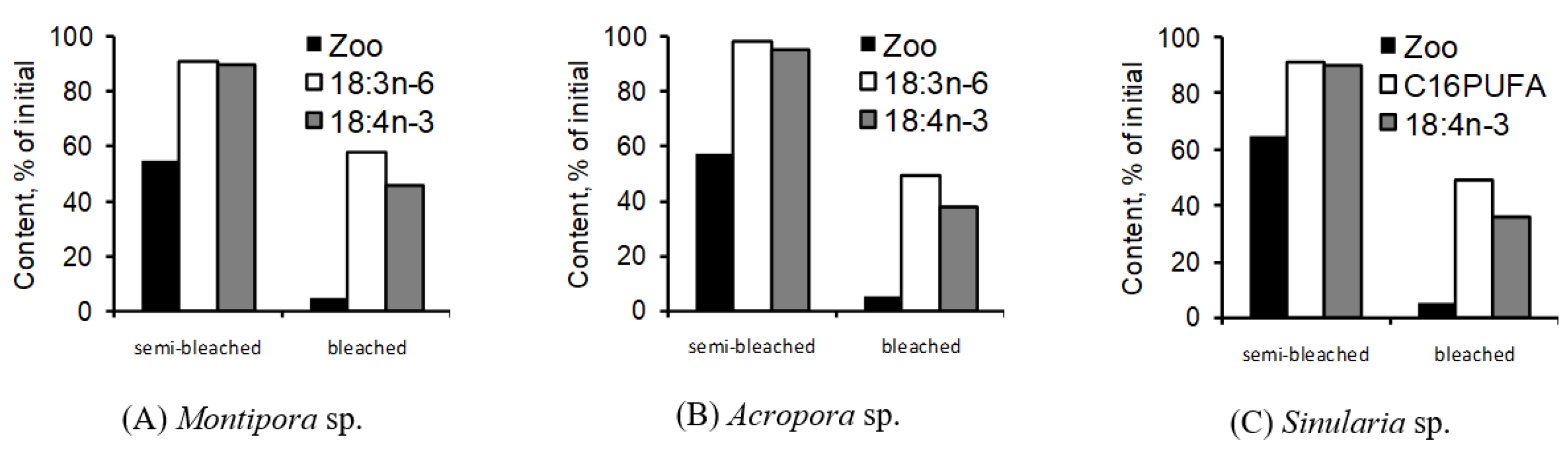
A detailed study assessed the dynamics of zooxanthellae and their marker PUFAs in whole colonies of both hard (specifically
Montipora sp. and
Acropora sp.) and soft (
Sinularia sp.) corals during coral bleaching [
134]. In this study, the marker PUFAs’ content was gauged in unfractionated, intact coral colonies to prevent potential contamination by zooxanthellae lipids. For the hard corals studied, 18:3n-6 and 18:4n-3 were identified as the zooxanthellae marker PUFAs, while for the soft corals, C16 PUFAs and 18:4n-3 were the markers. A 50% reduction in zooxanthellae led to only a 1.1-fold decrease in their marker PUFA levels in
Montipora sp. (as shown in
Figure 46A) and a 1.3-fold decrease in
Sinularia sp. (
Figure 46C). Surprisingly, even with a substantial 65% reduction in initial zooxanthellae levels in
Acropora sp., there was not a significant dip in marker PUFA levels (
Figure 46B). By the experiment’s conclusion, when coral bleaching was nearly complete (with a 90–95% loss of zooxanthellae), the levels of 18:3n-6 were 28% and 49% of their original values in
Montipora sp. and
Acropora sp., respectively (as shown in
Figure 46A,B). As for the 18:4n-3 levels, they were 46%, 38%, and 36% of their starting levels in
Montipora sp.,
Acropora sp., and
Sinularia sp., respectively, by the study’s end. Furthermore, the C16 PUFA level was at 45% of its initial value in
Sinularia sp. (illustrated in
Figure 46C).
The comparative analysis of zooxanthellae dynamics and their marker PUFAs revealed that a high concentration of zooxanthellae marker PUFAs remained in corals, even when approximately 95% of zooxanthellae had been lost from these colonies. This indicates that the marker PUFAs were retained within the host, rather than being lost alongside the symbionts. Logically, the concentration of symbiont marker PUFAs in the host would decrease when their primary source, the zooxanthellae, is diminished. This decline, however, seems to be a gradual process, likely spanning several days [
134]. For instance, when half of the zooxanthellae were depleted, the concentration of symbiont marker PUFAs in the overall FAs of the intact coral colony experienced only a minor reduction. As a specific example, experimental temperature stress led to a 1.5-fold decrease in the zooxanthellae density within hard coral
Turbinaria reniformis colonies, yet the levels of zooxanthellae marker FAs, such as 18:3n-6 and 18:4n-3, remained relatively stable [
227]. The stable carbon isotopic values (δ13C) of 18:3n-6, 20:4n-6, and 22:4n-6 from
T. reniformis colonies did not show significant variance when external food sources were excluded. This observation aligns with the notion that 18:3n-6 may be transferred from zooxanthellae to the host, serving as a precursor for the synthesis of 20:4n-6 and 22:4n-6 in polyp tissues.
Coral bleaching experiments have indirectly shown that certain symbiotic marker PUFAs are transferred to the host in both hard and soft corals [
134,
227]. The exploration of the transfer of C20-22 PUFAs, such as 20:5n-3, 22:5n-3, and 22:6n-3, from symbionts to the host is challenging using the marker acid method. This is because these PUFAs, while not unique to zooxanthellae, are scarcely found in the coral’s diet and are predominantly synthesized in the coral tissues themselves.
Beyond the marker FA method, the stable carbon isotope (δ
13C) composition of individual FAs, determined using gas chromatography-isotope ratio mass spectrometry (GC-IRMS), has proven effective in examining coral FA acquisition through translocation from zooxanthellae or via heterotrophic plankton consumption. The δ
13C values of FAs in corals would align with those of the same compound in its dietary source, as the isotope composition reflects the FA’s synthetic pathway and source. The δ
13C values for FAs produced by zooxanthellae and zooplankton differ significantly, by over 5‰ [
211].
Comparing the δ
13C values of individual FAs in zooxanthellae and coral hosts from two hard coral species,
Montastraea faveolata and
Porites astreoides, revealed that most energy-rich saturated FAs in coral colonies come from their zooxanthellae [
228]. The study also suggested that individual coral colonies source their essential PUFAs from both autotrophic and heterotrophic inputs, with the feeding mechanism specific to each colony. For instance, in
P. astreoides, the δ
13C values of 18:4n-3 in the host ranged between −12.2‰ and −15.8‰, which closely resembled those in zooxanthellae (from −11.5‰ to −13.8‰). However, these values differed significantly from those in zooplankton (from −22.0‰ to −25.0‰), indicating that zooxanthellae are the most likely source of 18:4n-3 in the host [
228].
In another study using
13C and
15N isotope tracers, Tanaka and co-workers [
229] delved into the carbon (C) and nitrogen (N) metabolism within the coral–zooxanthellae symbiotic system of
Acropora pulchra. They posited that the symbiotic system selectively consumed organic compounds with high C:N ratios, such as lipids and carbohydrates, faster than those with low C:N ratios such as proteins and nucleic acids.
The study explored the use of carbon in the coral tissue and zooxanthellae of
Stylophora pistillata and
Favia favus from the Red Sea, extending down a depth gradient of up to 60 m. Additionally, the isotopic carbon composition of the lipid fraction, taken from both the coral tissue and algal symbionts, was measured [
230]. The findings revealed that for both species, the δ
13C values in algal lipids were higher than those in tissue lipids. As depth increased, δ
13C values diminished by 7–8‰ in lipid fractions. Moreover, algal lipids had δ
13C values that were, on average, ~2‰ lower than those of the entire zooxanthellae at all depths. This suggests a highly efficient carbon recycling process between the coral host and its algal partners.
In another study, the stable isotope (
13C) uptake from dissolved inorganic carbon (NaH
13CO
3) in the coral
Acropora millepora and a clade B dinoflagellate culture was documented [
231]. Using the symbiotic anemone
Aiptasia pulchella (Anthozoa, Cnidaria) as a primary model system, the research delved into free FA (FFA) synthesis in both the dinoflagellate symbionts and cnidarian host and the lipid translocation from zooxanthellae to the host. Notably, no discernible evidence was found of the symbiont-derived enriched isotope FA or the direct utilization of catabolized
13C derivatives in host long-chain FA lipogenesis. Such results do not concur with the widely accepted translocation model concerning the employment of translocated symbiont photo-assimilates in host long-chain FA lipogenesis. It is crucial to understand that the findings of Dunn and colleagues [
231] were exclusively based on free FA, which only constituted a small percentage of the total lipids. Such lipids can stem from both non-specifically hydrolyzed lipids and unidentified associated microorganisms. Therefore, the
13C values recorded for FFA might not accurately portray the inorganic carbon integration in the total lipids and their FA. They also do not shed light on the lipid translocation process from zooxanthellae to the host in
A. pulchella.
In summary, a potential mechanism for the PUFA transfer from symbionts to the host in corals can be postulated. Acids such as 18:4n-3, 16:3n-4, and 16:4n-1 are produced in zooxanthellae and are either transferred in limited quantities to the host or are quickly metabolized by the host. Meanwhile, 18:3n-6 is transferred in larger volumes, especially in hard and certain soft coral species, and may serve as a precursor for n-6 series PUFAs in animal tissues. An interchange of 16:2n-7 likely occurs between the zooxanthellae and the host. The elongation of 16:2n-7 to 18:2n-7 predominantly happens within animal tissues. The methods and chemical forms of PUFA transfer from symbionts to hosts remain subjects of ongoing research and discussion.
A review of studies on zooxanthellae lipids reveals that zooxanthellae derived from different cnidarian species exhibit diverse FA compositions. For instance, tetracosapolyenoic FA is found in zooxanthellae lipids from soft corals, while 22:5n-6 is identified in hydrocoral lipids. Many researchers believe that corals are colonized by distinct zooxanthellae species, each with its unique, species-specific FA composition. However, there has not been direct taxonomic identification of these zooxanthellae species in certain studies [
112,
115]. FAs that are not typical of microalgae are suspected to have originated from contamination by animal lipids (such as polyp tissues) during the extraction of symbiont fractions [
69].
The taxonomy of coral zooxanthellae remains underdeveloped. All potential coral zooxanthellae species are generally categorized under the Simbiodinium group [
232]. The prevalent classification of coral zooxanthellae relies on genetic analysis of 16S ribosomal DNA. Currently, eight distinct zooxanthellae clades have been identified (A–H) [
233], and of these, five (A–D, F) are symbiotic with corals. The lipid composition of individual coral zooxanthellae clades has not been extensively studied. Yet, it is puzzling that if all coral zooxanthellae fall under the same genus, their FA compositions would vary so significantly. This variation in zooxanthellae FA composition can be more comprehensibly explained if one assumes that lipid transfer is bidirectional: not only from zooxanthellae to polyps, but also in the reverse, from the host organism back to the symbionts in corals.
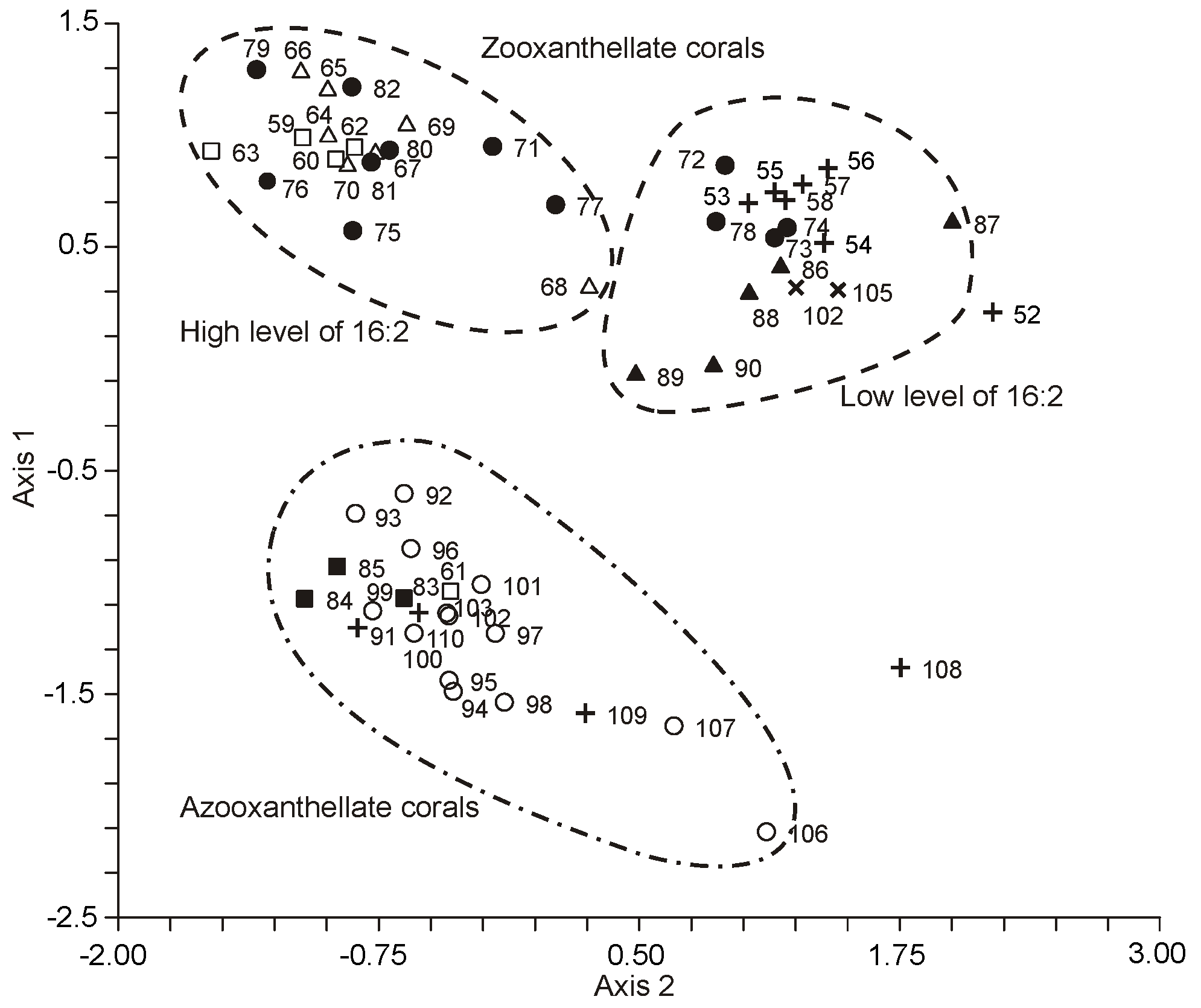
A principal component analysis (PCA) of the PUFA content in whole colony lipids, zooxanthellae, and polyp tissues from 10 cnidarian species, which included both soft and hard corals, as well as
Millepora platyphylla (referenced in
Table 21,
Figure 47 and
Figure 48), was conducted to validate the aforementioned hypothesis [
122]. Eight PUFAs (namely 16:3n-4, 16:4n-1, 18:3n-6, 18:4n-3, 20:4n-6, 20:5n-3, 22:5n-6, and 22:6n-3) were selected as variables for the PCA.
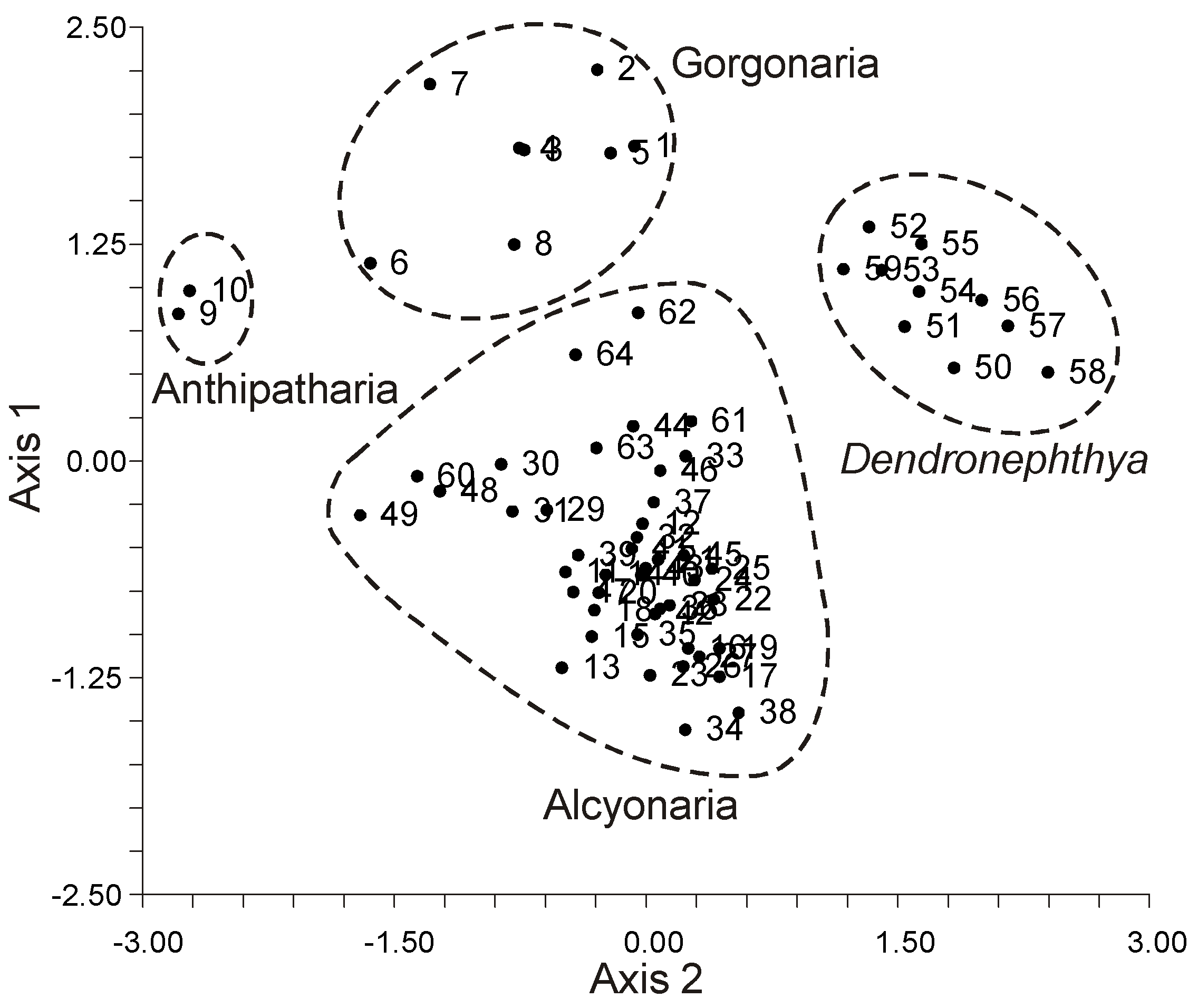
The intact colonies of hard corals, soft corals, and hydrocorals, along with their polyp tissues, were distinctly segregated into three groups within a two-dimensional space defined by axes 1 and 2, as illustrated in
Figure 47. Likewise, zooxanthellae were categorized into three groups that aligned with the taxonomic classifications of their hosts. Notably, zooxanthellae from hard corals were distinctly separated from those of soft corals and hydrocorals (as seen in
Figure 47). Clearly, if all zooxanthellae shared identical FA compositions, they would not cluster into these distinct, tight groupings.
Interestingly, no correlation was observed between the FA composition and the phylogenetic classification of the investigated zooxanthellae (as referenced in
Table 21 and
Table S43). This suggests that the variations in FA composition among zooxanthellae from different cnidarian groups are not influenced by the phylogenetic classifications of these zooxanthellae [
122].
Zooxanthellae derived from soft corals (Octocorallia) were distinctly set apart from those of hard corals and hydrocorals (Hexacorallia) based on their PUFA composition, as shown in
Figure 47. To clarify the differences in the FA composition of zooxanthellae across different coral types, the distribution of polyp marker PUFAs between the symbionts and their hosts was analyzed (refer to
Table S43 and
Figure 6). The host’s signature markers are n-6 series PUFAs, such as 20:4n-6 and 22:4n-6 [
69,
121]. Tetracosapolyenoic FAs, specifically 24:5n-6 and 24:6n-3, are indicative of octocoral polyp tissues, whereas 22:5n-6 is characteristic of Milleporidae polyp tissues [
68].
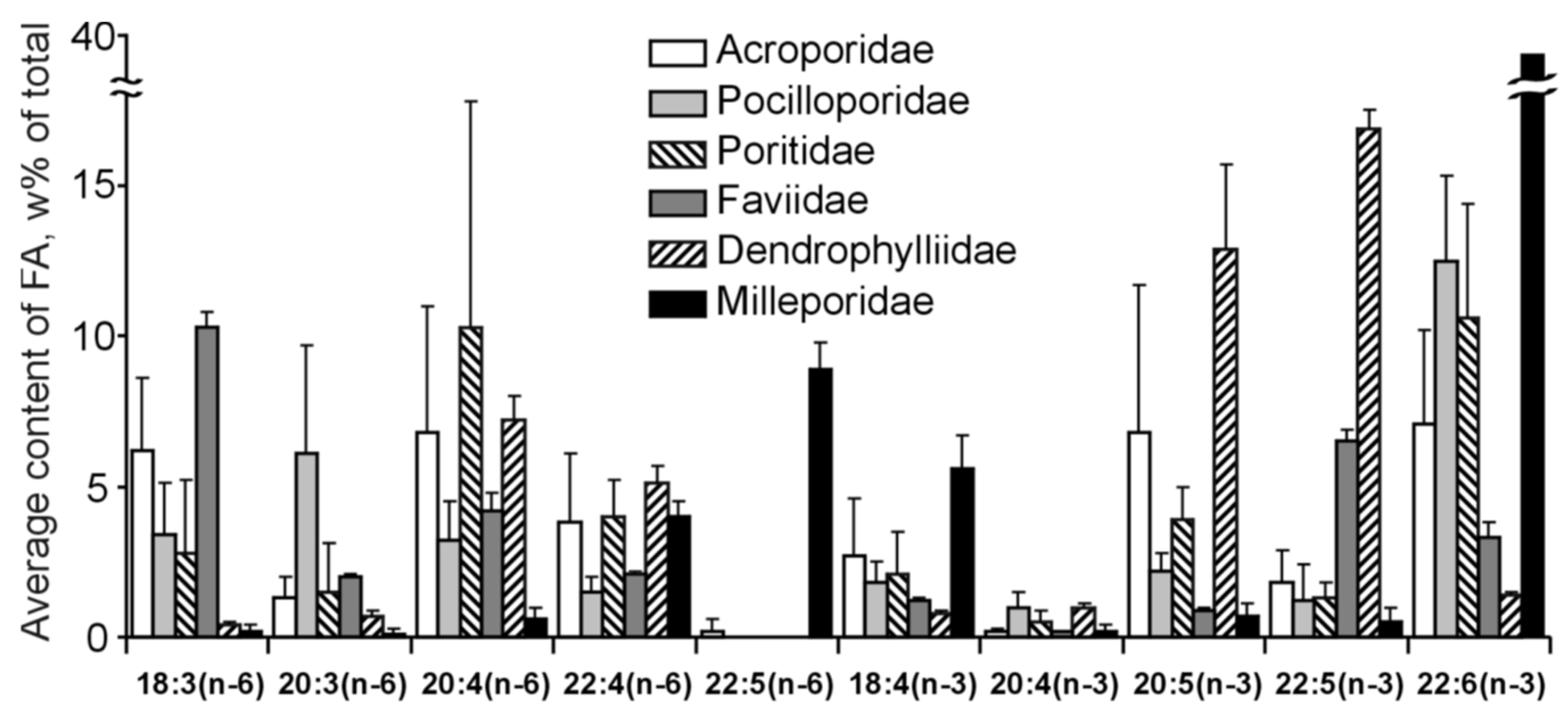
A considerable quantity of both 20:4n-6 and 22:4n-6 was present in all zooxanthellae (referenced in
Table S43 and
Figure 38). Additionally, the concentration of 20:4n-6 in both polyps and their respective zooxanthellae was notably similar in certain coral species, particularly in
P. damicornis and
P. cylindrica. No significant disparities were found in the levels of 22:4n-6 and 22:5n-6 between the zooxanthellae and polyps of
M. platyphylla. FAs 24:5n-6 and 24:6n-3 were identified in the zooxanthellae of soft corals; these fatty acids serve as markers for the host organism (as illustrated in
Table S43 and
Figure 38). The detection of distinct animal-specific PUFA markers in zooxanthellae may be attributable to the transfer of these PUFAs from the host to their symbiotic partners. Such a transfer could alter the inherent FA pattern of zooxanthellae and might account for the observed variations in the FA compositions of zooxanthellae across different cnidarian classifications.
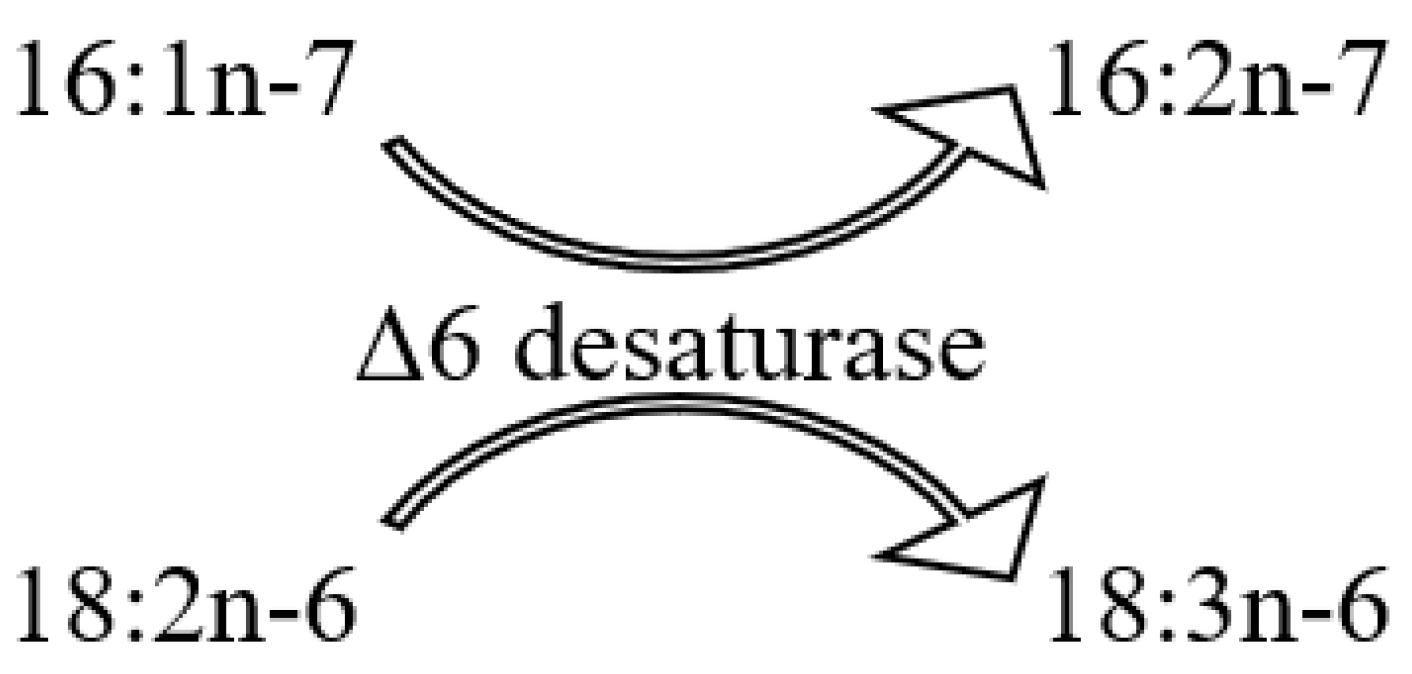
In the 10 cnidarian species studied, the zooxanthellae of hard and soft corals predominantly contained 18:3n-6 and C16 PUFAs, respectively. Neither 18:3n-6 nor C16 PUFAs were present in hydrocorals. Clearly, PUFA biosynthesis in these zooxanthellae groups follows distinct pathways [
70]. Crucially, both 18:3n-6 and 16:2n-7 are synthesized from 18:2n-6 and 16:1n-7, respectively, employing the same enzyme, Δ6 desaturase. It is plausible that the host organism exerts some level of regulation or modulation over PUFA biosynthesis within zooxanthellae, possibly by supplying the Δ6 desaturase with varied substrates (either 18:2n-6 or 16:1n-7). This regulatory influence exerted by the host on PUFA synthesis within the zooxanthellae might also explain the dependency of zooxanthellae FA composition on the host’s taxonomic classification.
Consequently, the impact of the host’s nature on the FA composition of their symbionts has been demonstrated. The transfer of host PUFAs to zooxanthellae and/or the modulation of PUFA biosynthesis in zooxanthellae by the host might be contributing factors to this effect. In their interaction, zooxanthellae occupy almost all the internal space of the host cell, reducing its cytoplasm and cell membrane to a thin external layer. Such spatial organization might facilitate the diffusion and transport of lipids and FAs between the host and the zooxanthellae. However, the exact mechanism of this transfer requires further study.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
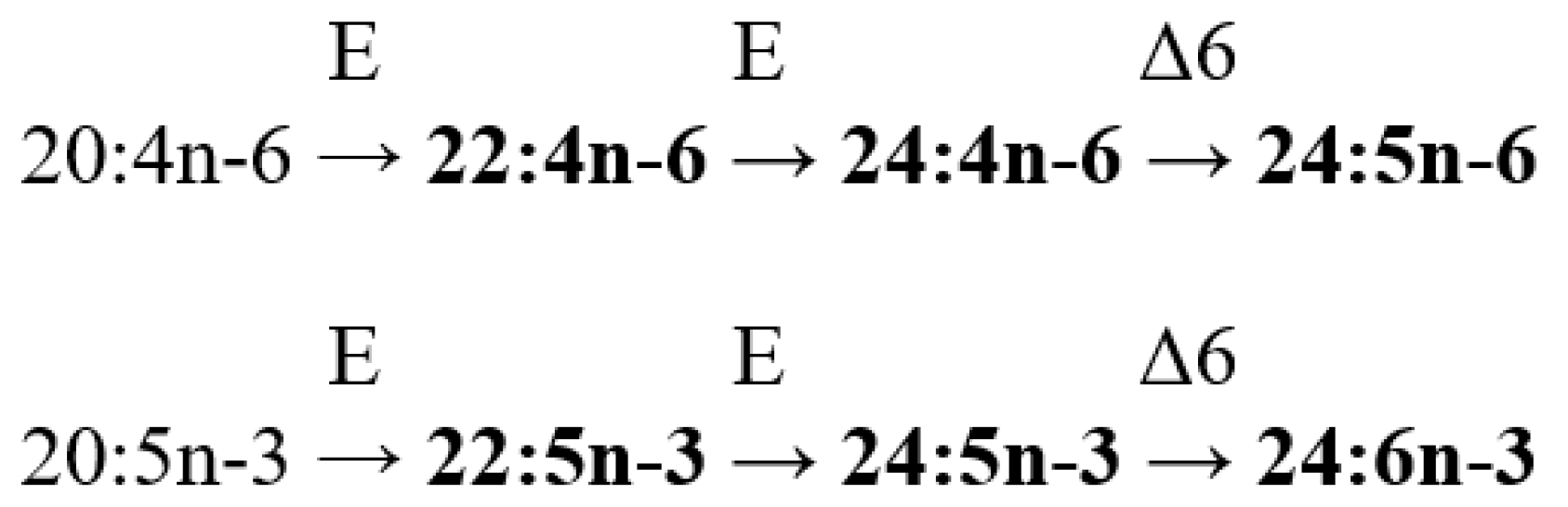{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}















































