
Identification of ACE I-Inhibitory Peptides Released by the Hydrolysis of Tub Gurnard (Chelidonichthys lucerna) Skin Proteins and the Impact of Their In Silico Gastrointestinal Digestion


 and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Analysis of Raw Material
2.2. Degree of Hydrolysis (DH)
2.3. MW Distribution of the Hydrolysates
2.4. ACE-Inhibitory Activity
2.5. Isolation and Purification of ACE-Inhibitory Peptides
2.6. Identification of the Peptide Composition of the Most Potent ACE-Inhibiting Fractions
2.7. In Silico Gastrointestinal Digestion of the Peptides in the Most Potent ACE-Inhibiting Fractions
3. Materials and Methods
3.1. Reagents
3.2. Sample Preparation
3.3. Proteolytic Activity
3.4. Analysis of the Proximate Composition of the Raw Material
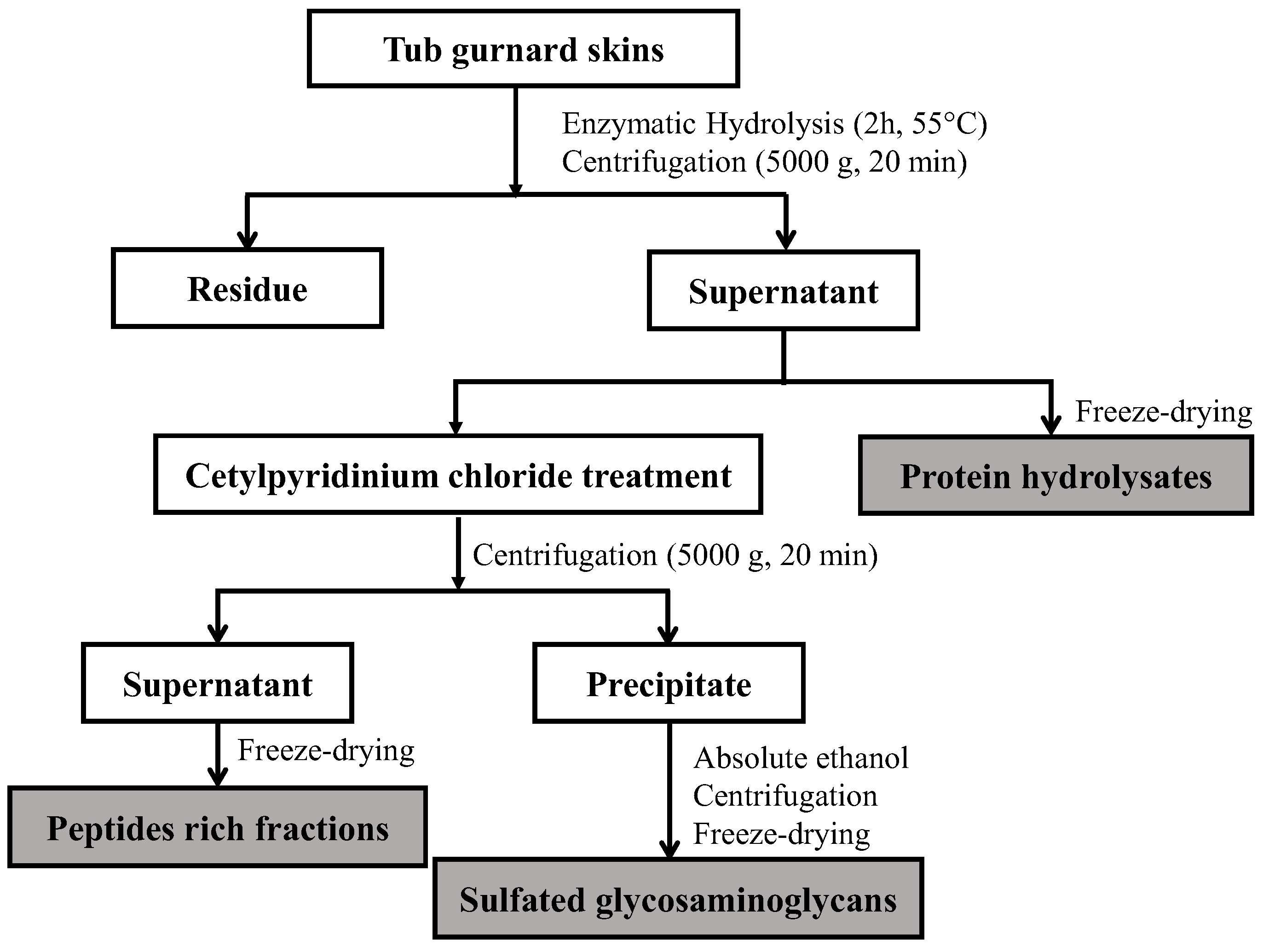
3.5. Extraction Procedure
3.5.1. Preparation of Enzymatic Hydrolysates
3.5.2. Separation and Purification of Glycosaminoglycans
3.6. Determination of the Degree of Hydrolysis
3.7. Determination of the Molecular Weight (MW) Profile
3.8. Analysis of Amino Acid Compositions
3.9. Determination of GAGs Content
3.10. Evaluation of the ACE-Inhibitory Activity
3.11. Fractionation and Purification of Active Peptides
3.12. Identification of the Peptide Composition of the ACE-Inhibiting Fraction
3.12.1. Analysis by RP-LC-ESI-MS/MS
3.12.2. Analysis of MS/MS Spectra
3.13. In Silico Gastrointestinal Digestion of the ACE-Inhibiting Fractions
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller-Nurasyid, M. Worldwide Trends in Hypertension Prevalence and Progress in Treatment and Control from 1990 to 2019: A Pooled Analysis of 1201 Population-Representative Studies with 104 Million Participants. Lancet 2021, 398, 957–980. [Google Scholar] [CrossRef]
- Messerli, F.H.; Bangalore, S.; Bavishi, C.; Rimoldi, S.F. Angiotensin-Converting Enzyme Inhibitors in Hypertension. J. Am. Coll. Cardiol. 2018, 71, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Alvarez, O.; Batista, I.; Ramos, C.; Montero, P. Enhancement of ACE and Prolyl Oligopeptidase Inhibitory Potency of Protein Hydrolysates from Sardine and Tuna By-Products by Simulated Gastrointestinal Digestion. Food Funct. 2016, 7, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Angiotensin I-Converting Enzyme (ACE) Inhibitory Activity, Antioxidant Properties, Phenolic Content and Amino Acid Profiles of Fucus spiralis L. Protein Hydrolysate Fractions. Mar. Drugs 2017, 15, 311. [Google Scholar] [CrossRef]
- Fan, H.; Yu, W.; Liao, W.; Wu, J. Spent Hen Protein Hydrolysate with Good Gastrointestinal Stability and Permeability in Caco-2 Cells Shows Antihypertensive Activity in SHR. Foods 2020, 9, 1384. [Google Scholar] [CrossRef]
- Salampessy, J.; Reddy, N.; Phillips, M.; Kailasapathy, K. Isolation and Characterization of Nutraceutically Potential ACE-Inhibitory Peptides from Leatherjacket (Meuchenia sp.) Protein Hydrolysates. LWT 2017, 80, 430–436. [Google Scholar] [CrossRef]
- Phadke, G.G.; Rathod, N.B.; Ozogul, F.; Elavarasan, K.; Karthikeyan, M.; Shin, K.-H.; Kim, S.-K. Exploiting of Secondary Raw Materials from Fish Processing Industry as a Source of Bioactive Peptide-Rich Protein Hydrolysates. Mar. Drugs 2021, 19, 480. [Google Scholar] [CrossRef]
- Caruso, G.; Abdelhamid, M.T.; Kalisz, A.; Sekara, A. Linking Endophytic Fungi to Medicinal Plants Therapeutic Activity. A Case Study on Asteraceae. Agriculture 2020, 10, 286. [Google Scholar] [CrossRef]
- Stergiou, K.I.; Machias, A.; Somarakis, S.; Kapantagakis, A. Can We Define Target Species in Mediterranean Trawl Fisheries? Fish. Res. 2003, 59, 431–435. [Google Scholar] [CrossRef]
- Venugopal, V. Valorization of Seafood Processing Discards: Bioconversion and Bio-Refinery Approaches. Front. Sustain. Food Syst. 2021, 5, 611835. [Google Scholar] [CrossRef]
- Nag, M.; Lahiri, D.; Dey, A.; Sarkar, T.; Pati, S.; Joshi, S.; Bunawan, H.; Mohammed, A.; Edinur, H.; Nirmal, N.; et al. Seafood Discards: A Potent Source of Enzymes and Biomacromolecules with Nutritional and Nutraceutical Significance. Front. Nutr. 2022, 9, 879929. [Google Scholar] [CrossRef] [PubMed]
- Sarmadi, B.; Ismail, A.; Hamid, M. Antioxidant and Angiotensin Converting Enzyme (ACE) Inhibitory Activities of Cocoa (Theobroma cacao L.) Autolysates. Food Res. Int. 2011, 44, 290–296. [Google Scholar] [CrossRef]
- Mutalipassi, M.; Esposito, R.; Ruocco, N.; Viel, T.; Costantini, M.; Zupo, V. Bioactive Compounds of Nutraceutical Value from Fishery and Aquaculture Discards. Foods 2021, 10, 1495. [Google Scholar] [CrossRef]
- Roncarati, A.; Felici, A.; Mariotti, F.; Melotti, P. Flesh Qualitative Traits of Tub Gurnard (Chelidonichthys Lucerna L.), A Promising Species Candidate for Aquaculture, Captured in the Middle Adriatic Sea in Different Seasons. Ital. J. Anim. Sci. 2014, 13, 3159. [Google Scholar] [CrossRef] [Green Version]
- Sila, A.; Bougatef, A. Antioxidant Peptides from Marine By-Products: Isolation, Identification and Application in Food Systems. A Review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Meng, D.; Tanaka, H.; Kobayashi, T.; Hatayama, H.; Zhang, X.; Ura, K.; Yunoki, S.; Takagi, Y. The Effect of Alkaline Pretreatment on the Biochemical Characteristics and Fibril-Forming Abilities of Types I and II Collagen Extracted from Bester Sturgeon by-Products. Int. J. Biol. Macromol. 2019, 131, 572–580. [Google Scholar] [CrossRef]
- Truong, T.M.T.; Nguyen, V.M.; Tran, T.T.; Le, T.M.T. Characterization of Acid-Soluble Collagen from Food Processing By-Products of Snakehead Fish (Channa Striata). Processes 2021, 9, 1188. [Google Scholar] [CrossRef]
- Byun, H.-G.; Kim, S.-K. Purification and Characterization of Angiotensin I Converting Enzyme (ACE) Inhibitory Peptides from Alaska Pollack (Theragra chalcogramma) Skin. Process Biochem. 2001, 36, 1155–1162. [Google Scholar] [CrossRef]
- Raikos, V.; Hays, H.; Stead, D.; Ni, H. Angiotensin-Converting Enzyme Inhibitory Activity of Hydrolysates Generated from Whey Protein Fortified with Salal Fruits (Galtheria shallon) by Enzymatic Treatment with Pronase from Streptomyces Griseus. Int. J. Food Sci. Technol. 2019, 54, 2975–2982. [Google Scholar] [CrossRef]
- Nuchprapha, A.; Paisansak, S.; Sangtanoo, P.; Srimongkol, P.; Saisavoey, T.; Reamtong, O.; Choowongkomon, K.; Karnchanatat, A. Two Novel ACE Inhibitory Peptides Isolated from Longan Seeds: Purification, Inhibitory Kinetics and Mechanisms. RSC Adv. 2020, 10, 12711–12720. [Google Scholar] [CrossRef] [Green Version]
- Prydz, K. Determinants of Glycosaminoglycan (GAG) Structure. Biomolecules 2015, 5, 2003–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimzadeh, K. Antihypertensive and anticoagulant properties of glycosaminoglycans extracted from the sturgeon (Acipenser persicus) cartilage. Curr. Issues Pharm. Med. Sci. 2018, 31, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Krichen, F.; Bougatef, H.; Capitani, F.; Amor, I.B.; Koubaa, I.; Gargouri, J.; Maccari, F.; Mantovani, V.; Galeotti, F.; Volpi, N.; et al. Purification and Structural Elucidation of Chondroitin Sulfate/Dermatan Sulfate from Atlantic Bluefin Tuna (Thunnus thynnus) Skins and Their Anticoagulant and ACE Inhibitory Activities. RSC Adv. 2018, 8, 37965–37975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmalek, B.E.; Sila, A.; Krichen, F.; Karoud, W.; Martinez-Alvarez, O.; Ellouz-Chaabouni, S.; Ayadi, M.A.; Bougatef, A. Sulfated Polysaccharides from Loligo Vulgaris Skin: Potential Biological Activities and Partial Purification. Int. J. Biol. Macromol. 2015, 72, 1143–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosquera, M.; Giménez, B.; Ramos, S.; López-Caballero, M.E.; del Carmen Gómez-Guillén, M.; Montero, P. Antioxidant, ACE-Inhibitory, and Antimicrobial Activities of Peptide Fractions Obtained From Dried Giant Squid Tunics. J. Aquat. Food Prod. Technol. 2016, 25, 444–455. [Google Scholar] [CrossRef]
- Ghalamara, S.; Silva, S.; Brazinha, C.; Pintado, M. Valorization of Fish By-Products: Purification of Bioactive Peptides from Codfish Blood and Sardine Cooking Wastewaters by Membrane Processing. Membranes 2020, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Mongkonkamthorn, N.; Malila, Y.; Regenstein, J.M.; Wangtueai, S. Enzymatic Hydrolysis Optimization for Preparation of Tuna Dark Meat Hydrolysate with Antioxidant and Angiotensin I-Converting Enzyme (ACE) Inhibitory Activities. J. Aquat. Food Prod. Technol. 2021, 30, 1090–1108. [Google Scholar] [CrossRef]
- Heffernan, S.; Giblin, L.; O’Brien, N. Assessment of the Biological Activity of Fish Muscle Protein Hydrolysates Using in Vitro Model Systems. Food Chem. 2021, 359, 129852. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-Converting Enzyme (ACE) Inhibitors from Marine Resources: Prospects in the Pharmaceutical Industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [Green Version]
- Shi, A.; Liu, H.; Liu, L.; Hu, H.; Wang, Q.; Adhikari, B. Isolation, Purification and Molecular Mechanism of a Peanut Protein-Derived ACE-Inhibitory Peptide. PLoS ONE 2014, 9, e111188. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Wang, Y.; Hao, Y.; Zhang, W.; Zhou, G. Antihypertensive Effects in Vitro and in Vivo of Novel Angiotensin-Converting Enzyme Inhibitory Peptides from Bovine Bone Gelatin Hydrolysate. J. Agric. Food Chem. 2020, 68, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE Inhibitory Peptides Derived from Simulated Gastrointestinal Digestion in Vitro of Sesame (Sesamum indicum L.) Protein and Molecular Docking Study. Int. J. Mol. Sci. 2020, 21, 1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure-Activity Relationship Modeling of Peptides Containing 4-10 Amino Acid Residues. QSAR Comb. Sci. 2006, 25, 873–880. [Google Scholar] [CrossRef]
- Qian, Z.-J.; Jung, W.-K.; Lee, S.-H.; Byun, H.-G.; Kim, S.-K. Antihypertensive Effect of an Angiotensin I-Converting Enzyme Inhibitory Peptide from Bullfrog (Rana Catesbeiana Shaw) Muscle Protein in Spontaneously Hypertensive Rats. Process Biochem. 2007, 42, 1443–1448. [Google Scholar] [CrossRef]
- Lajmi, K.; Gómez-Estaca, J.; Hammami, M.; Martínez-Alvarez, O. Upgrading Collagenous Smooth Hound By-Products: Effect of Hydrolysis Conditions, in Vitro Gastrointestinal Digestion and Encapsulation on Bioactive Properties. Food Biosci. 2019, 28, 99–108. [Google Scholar] [CrossRef]
- Hamed, M.; Bougatef, H.; Karoud, W.; Krichen, F.; Haddar, A.; Bougatef, A.; Sila, A. Polysaccharides Extracted from Pistachio External Hull: Characterization, Antioxidant Activity and Potential Application on Meat as Preservative. Ind. Crops Prod. 2020, 148, 112315. [Google Scholar] [CrossRef]
- Ben Mansour, M.; Majdoub, H.; Bataille, I.; Roudesli, M.S.; Hassine, M.; Ajzenberg, N.; Chaubet, F.; Maaroufi, R.M. Polysaccharides from the Skin of the Ray Raja Radula. Partial Characterization and Anticoagulant Activity. Thromb. Res. 2009, 123, 671–678. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: London, UK, 1986; ISBN 978-0-85334-386-8. [Google Scholar]
- Martínez-Alvarez, O.; Ravallec, R.; Cudennec, B.; Guimas, L.; Delannoy, C.; Fouchereau-Peron, M. Effect of Daily Gavage with a Collagen Hydrolysate Containing Calcitonin Gene-Related Peptide (CGRP)-like Molecules on Plasma CGRP-Levels in Rats. J. Funct. Foods 2012, 4, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lawrence, R.; Schwartz, J.J.; Bai, X.; Wei, G.; Esko, J.D.; Rosenberg, R.D. The Effect of Precursor Structures on the Action of Glucosaminyl 3-O-Sulfotransferase-1 and the Biosynthesis of Anticoagulant Heparan Sulfate. J. Biol. Chem. 2001, 276, 28806–28813. [Google Scholar] [CrossRef] [Green Version]
- Sentandreu, M.Á.; Toldrá, F. A Rapid, Simple and Sensitive Fluorescence Method for the Assay of Angiotensin-I Converting Enzyme. Food Chem. 2006, 97, 546–554. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Amino Acids | ‰ |
|---|---|
| Asp + Asn (D + N) | 67 |
| Thr (T) | 34 |
| Ser (S) | 56 |
| Glu + Gln (E + Q) | 85 |
| Gly (G) | 269 |
| Ala (A) | 111 |
| Cys (C) | 4 |
| Val (V) | 29 |
| Met (M) | 19 |
| Ile (I) | 17 |
| Leu (L) | 41 |
| Tyr (Y) | 10 |
| Phe (F) | 21 |
| Hyl | 7 |
| His (H) | 10 |
| Lys (K) | 38 |
| Arg (R) | 48 |
| Pro (P) | 80 |
| Hyp | 54 |
| TEAA * | 209 |
| THAA * | 587 |
| Pro + Hyp | 134 |
| Enzyme | Enzymatic Conditions | MW at 214 nm (Da) | MW at 280 nm (Da) | ACE-Inhibition (%) | |
|---|---|---|---|---|---|
| pH | Temperature (°C) | ||||
| Esperase | 8.5 | 55 | 729 | 358 | 93.92 ± 2.04 a |
| Alcalase | 8.5 | 55 | 927 | 545 (48%) | 86.16 ± 2.29 b |
| 375 (52%) | |||||
| Trypsin | 8.5 | 40 | 2052 | 1646 | 90.92 ± 2.04 a |
| Pronase E | 8.5 | 55 | 3700 | 2165 | 83.34 ± 0.73 b |
| Sample | ACE Inhibition | ||
|---|---|---|---|
| Inhibition (%) | IC50 Value (μg/mL) | ||
| PRFs | Esperase | 91.44 ± 0.69 a | 47 ± 2 |
| Alcalase | 91.12 ± 0.63 a | 68 ± 2 | |
| Trypsin | 89.37 ± 0.44 a | 89 ± 2 | |
| GAGs | Esperase | 49.81 ± 2.32 d | - |
| Alcalase | 52.34 ± 0.23 b | - | |
| Trypsin | 62.99 ± 2.09 c | - | |
| Peptide | −10lgP | Mass (Da) | ppm | ALC (%) | Relative Abundance (%) | Parental Protein |
|---|---|---|---|---|---|---|
| (1) GPPGS(sub P)PGPAGPP(+15.99)GPPGSGM | 23.48 | 1586.7 | −0.2 | 37.5 | Collagen (1) | |
| (2) GM(+15.99)P(+15.99)GERGAAGLP(+15.99)GLR | 23.16 | 1485.7 | 1.1 | 24.6 | Collagen (2) | |
| (3) GPAGIVGPP(+15.99)GPAGPA | 28.06 | 1229.6 | 1.6 | 9.9 | Collagen (3) | |
| (4) GVEDELDKY | 1066.5 | −0.6 | 97 | 4.5 | ||
| (5) VNPVYEGY | 939.4 | 0.6 | 86 | 4.0 | ||
| (6) GPP(+15.99)GSP(+15.99)GLPGPPGPA | 23.71 | 1285.6 | 1.8 | 1.7 | Collagen (4) | |
| (7) TDGLDGPYDELK | 1321.6 | −0.5 | 90 | 1.0 | ||
| (8) TGFPDPVAGN | 973.4 | 0.2 | 85 | 1.0 | ||
| (9) TGFPPDVAGN | 973.4 | 0.2 | 86 | 1.0 | ||
| (10) TGFPPVDAGN | 973.4 | 0.2 | 89 | 1.0 | ||
| (11) TGFPPDVAGN | 973.4 | 0.2 | 94 | 1.0 | ||
| (12) SADAPM(+15.99)FVM(+15.99) | 999.4 | −1.2 | 94 | 0.9 | ||
| (13) QSLFPLQ | 831.4 | 0.1 | 82 | 0.9 |
| Peptide | −10lgP | Mass (Da) | ppm | ALC (%) | Relative Abundance (%) | Parental Protein |
|---|---|---|---|---|---|---|
| (1) LLAPPERKY | 1085.6 | −0.7 | 86 | 30.84 | ||
| (2) ELEEELEAE | 1089.5 | −1.6 | 94 | 15.44 | ||
| (3) GPRGPAGPL | 820.4 | 1.2 | 92 | 11.89 | Collagen (1) | |
| (4) SLDDKVELE | 1046.5 | 0.8 | 96 | 8.49 | ||
| (5) KYPLEHALLTK | 1311.8 | −1.8 | 84 | 5.80 | ||
| (6) AGPP(+15.99)GP(+15.99)AGPP(+15.99)APGAPGGGFD | 23.2 | 1690.8 | 1.6 | 2.17 | Collagen (2) | |
| (7) AGPP(+15.99)GFP(+15.99)GGPGP(+15.99)KGEIGPA | 27.77 | 1706.8 | 1.5 | 2.00 | Collagen (3) | |
| (8) AGPP(+15.99)GFP(+15.99)GGPGP(+15.99)KGELGPA | 27.77 | 1706.8 | 1.5 | 2.00 | Collagen (4) | |
| (9) AGP(+15.99)PGFP(+15.99)GGP(+15.99)GPKGEIGPA | 27.46 | 1706.8 | 1.5 | 2.00 | Collagen (5) | |
| (10) AGP(+15.99)PGFP(+15.99)GGP(+15.99)GPKGELGPA | 27.46 | 1706.8 | 1.5 | 2.00 | Collagen (6) | |
| (11) LEHEEGKLL | 1066.6 | −1.9 | 98 | 1.77 | ||
| (12) KTDDGKLFT | 1023.5 | −1.5 | 90 | 1.00 | ||
| (13) QP(+15.99)GNTGLPGMT | 1087.5 | 2 | 91 | 0.99 | Collagen (7) | |
| (14) NIGFP(+15.99)GPK(+14.02)GASGDPGKP(+15.99)GDKGATGPS | 24.18 | 2411.1 | −0.6 | 0.98 | Collagen (8) |
| Sequence | Mass (Da) | Times Released |
|---|---|---|
| GY | 238 | 1 |
| PGL | 285 | 2 |
| PL | 228 | 1 |
| AF | 236 | 1 |
| GL | 188 | 1 |
| DF | 280 | 3 |
| VM | 248 | 2 |
| Sequence | Mass (Da) | Times Released |
|---|---|---|
| GY | 238 | 3 |
| PL | 228 | 5 |
| IW | 317 | 1 |
| VK | 245 | 1 |
| GM | 206 | 2 |
| GL | 188 | 1 |
| GK | 203 | 2 |
| VR | 273 | 1 |
| EK | 275 | 1 |
| TF | 266 | 1 |
| AGDDAPR | 700 | 1 |
| VM | 248 | 1 |
| ER | 303 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bougatef, H.; de la Vega-Fernández, C.; Sila, A.; Bougatef, A.; Martínez-Alvarez, O. Identification of ACE I-Inhibitory Peptides Released by the Hydrolysis of Tub Gurnard (Chelidonichthys lucerna) Skin Proteins and the Impact of Their In Silico Gastrointestinal Digestion. Mar. Drugs 2023, 21, 131. https://doi.org/10.3390/md21020131
Bougatef H, de la Vega-Fernández C, Sila A, Bougatef A, Martínez-Alvarez O. Identification of ACE I-Inhibitory Peptides Released by the Hydrolysis of Tub Gurnard (Chelidonichthys lucerna) Skin Proteins and the Impact of Their In Silico Gastrointestinal Digestion. Marine Drugs. 2023; 21(2):131. https://doi.org/10.3390/md21020131
Chicago/Turabian StyleBougatef, Hajer, Cristina de la Vega-Fernández, Assaad Sila, Ali Bougatef, and Oscar Martínez-Alvarez. 2023. "Identification of ACE I-Inhibitory Peptides Released by the Hydrolysis of Tub Gurnard (Chelidonichthys lucerna) Skin Proteins and the Impact of Their In Silico Gastrointestinal Digestion" Marine Drugs 21, no. 2: 131. https://doi.org/10.3390/md21020131
APA StyleBougatef, H., de la Vega-Fernández, C., Sila, A., Bougatef, A., & Martínez-Alvarez, O. (2023). Identification of ACE I-Inhibitory Peptides Released by the Hydrolysis of Tub Gurnard (Chelidonichthys lucerna) Skin Proteins and the Impact of Their In Silico Gastrointestinal Digestion. Marine Drugs, 21(2), 131. https://doi.org/10.3390/md21020131