
Mechanisms of Antitumor Invasion and Metastasis of the Marine Fungal Derivative Epi-Aszonalenin A in HT1080 Cells

 , and
, and 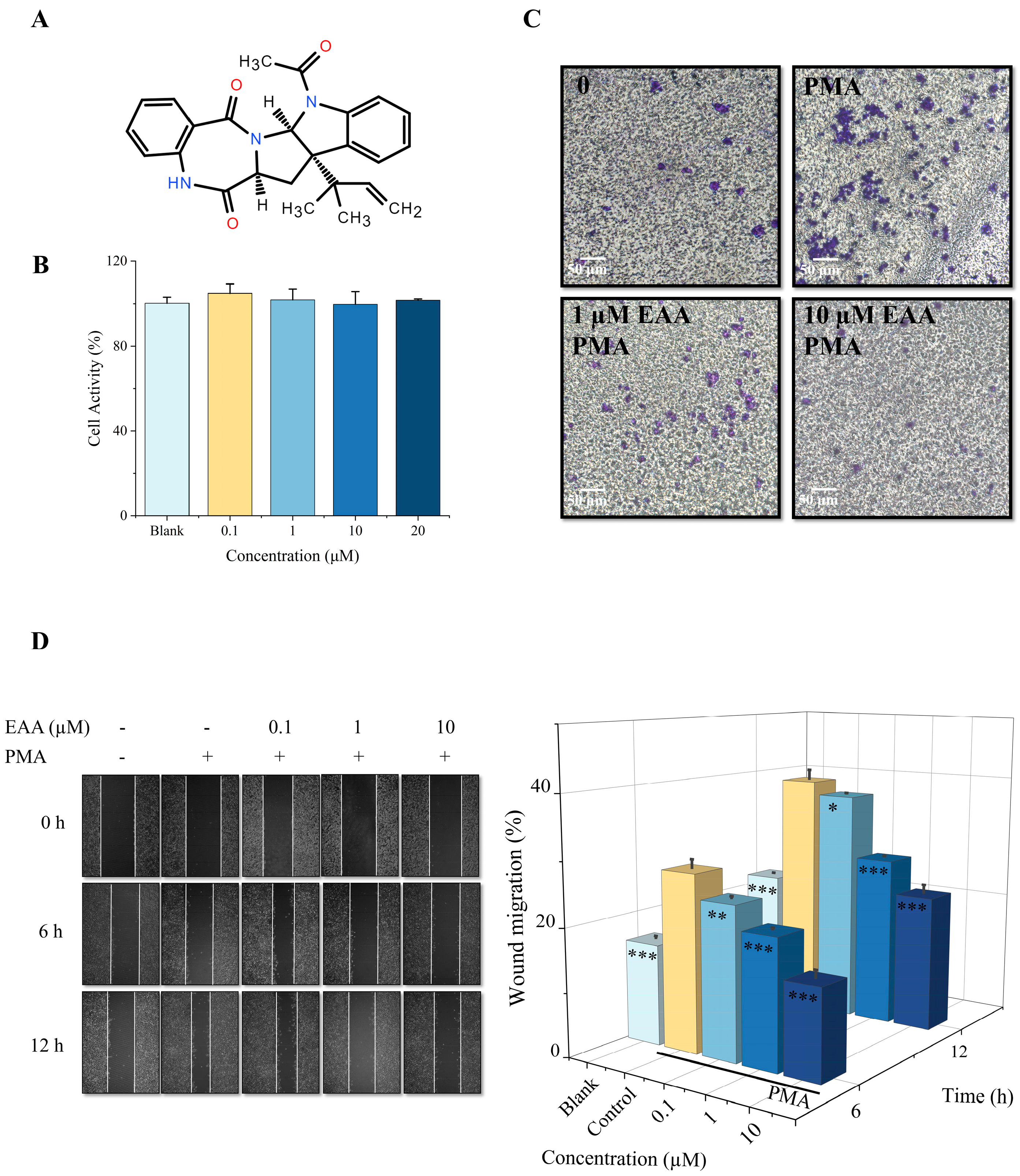{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of EAA on the Viability of HT1080 Cells
2.2. Effect of EAA on Migration Capacity and Invasion of HT1080 Cells
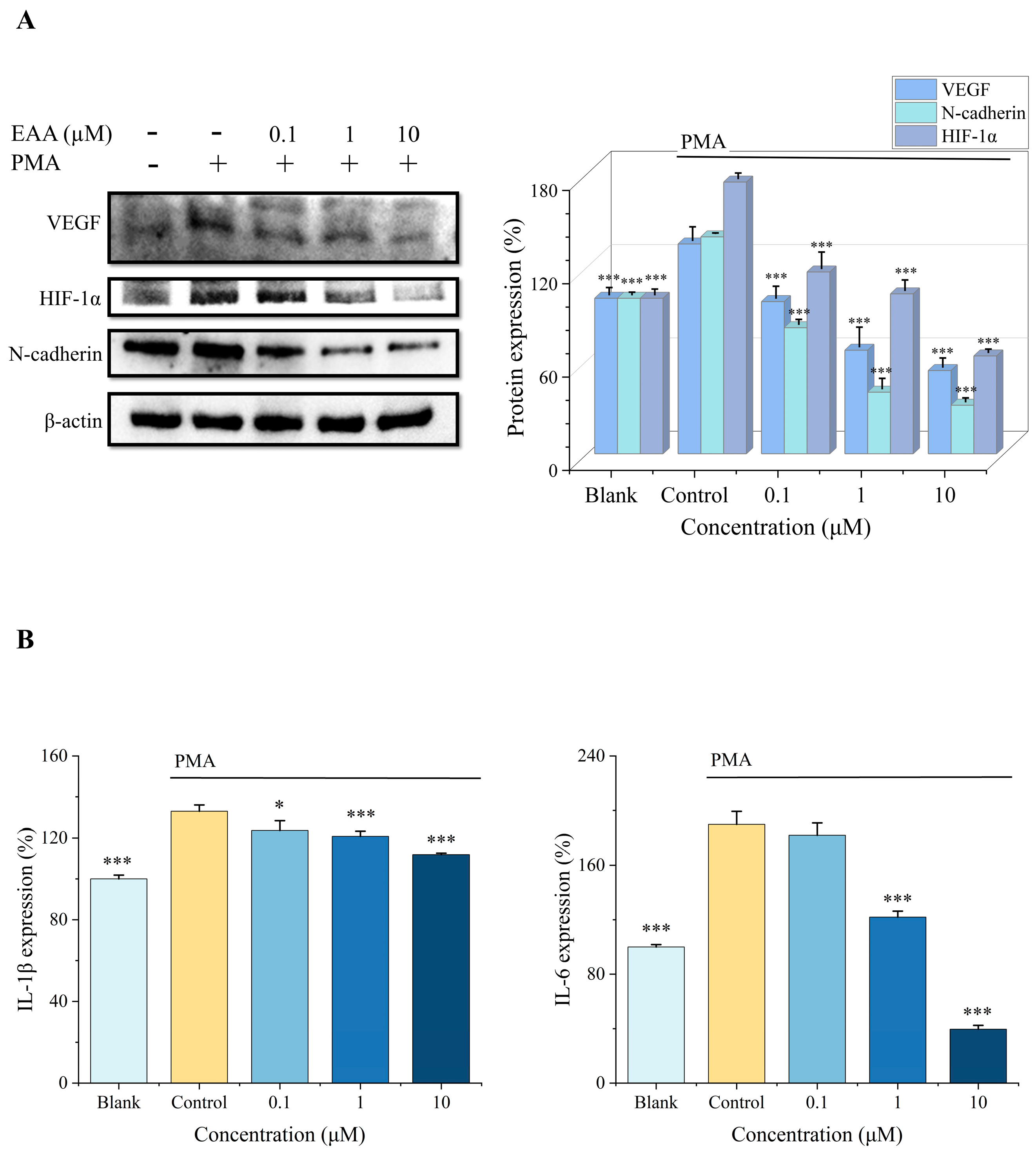
2.3. Effect of EAA on HIF-1α, VEGF, and N-Cadherin Expression in the HT1080 Cells Induced by PMA
2.4. Effect of EAA on IL-1β and IL-6 Expression in HT1080 Cells Induced by PMA
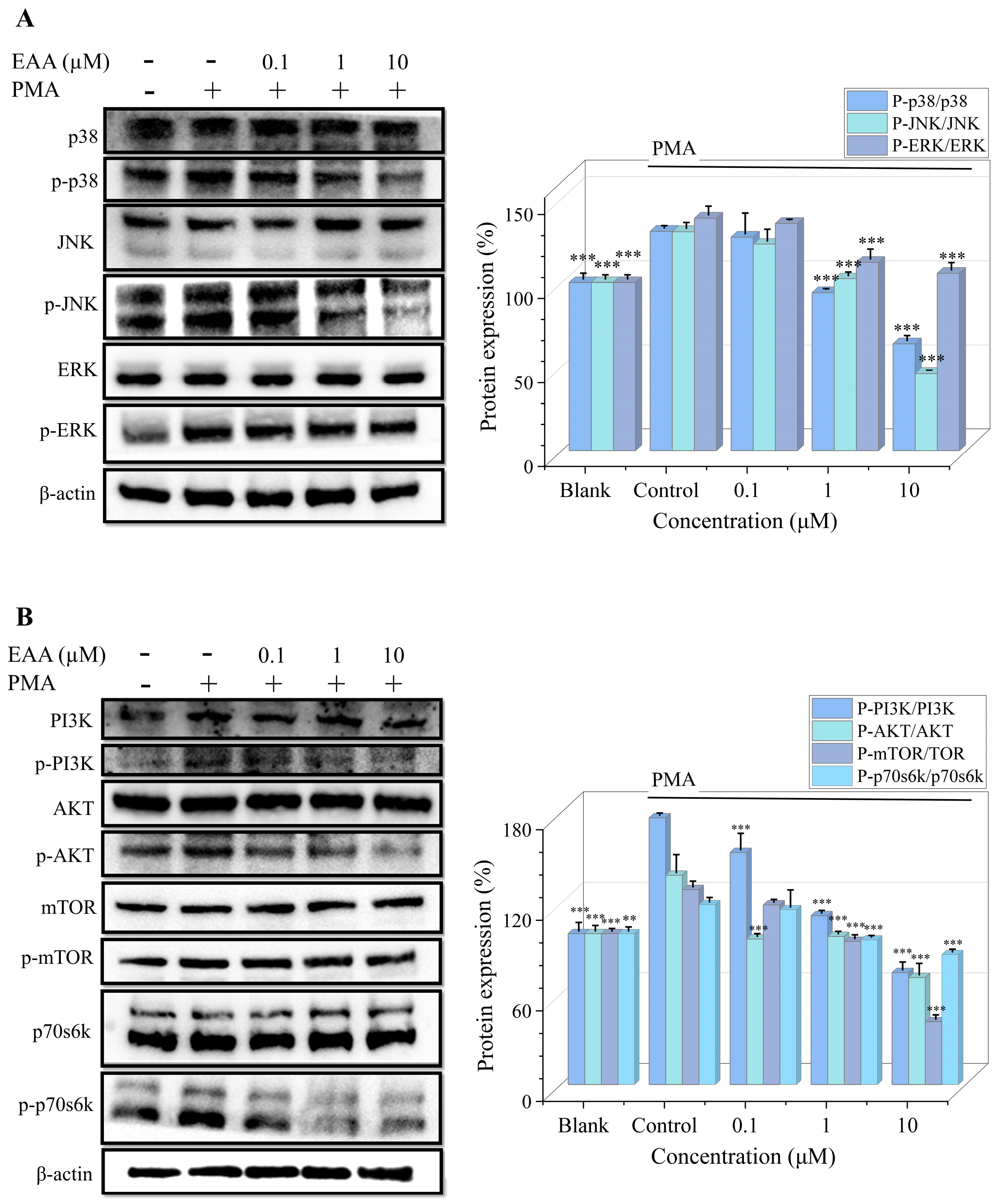
2.5. Effect of EAA on MAPK and PI3K/Akt Pathway in PMA-Induced HT1080 Cells
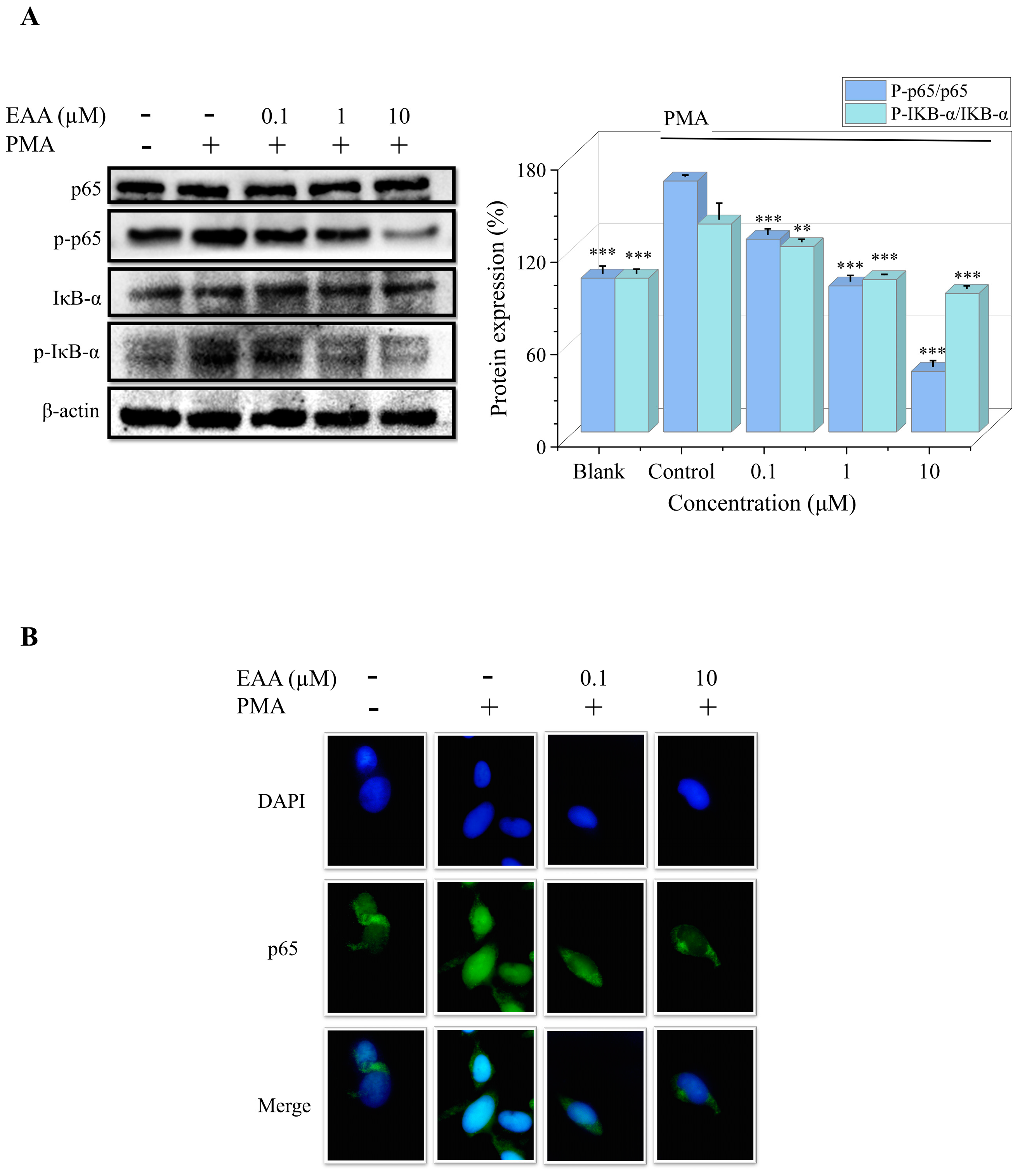
2.6. Effects of EAA on PMA-Induced NF-κB Pathway and DNA-Binding Activity in HT1080 Cells
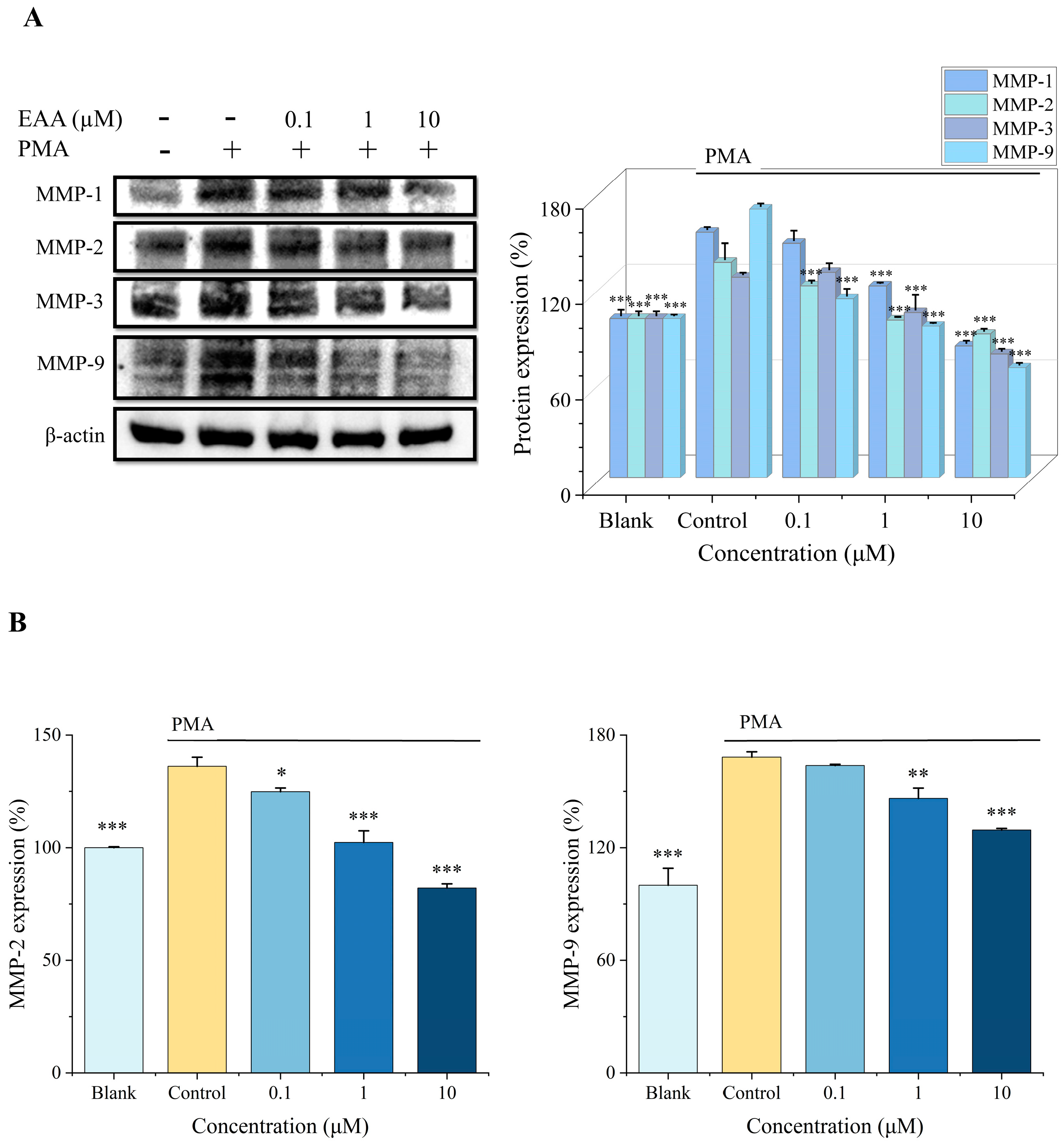
2.7. Effect of EAA on MMPs Activity and Expression in HT1080 Cells Induced by PMA
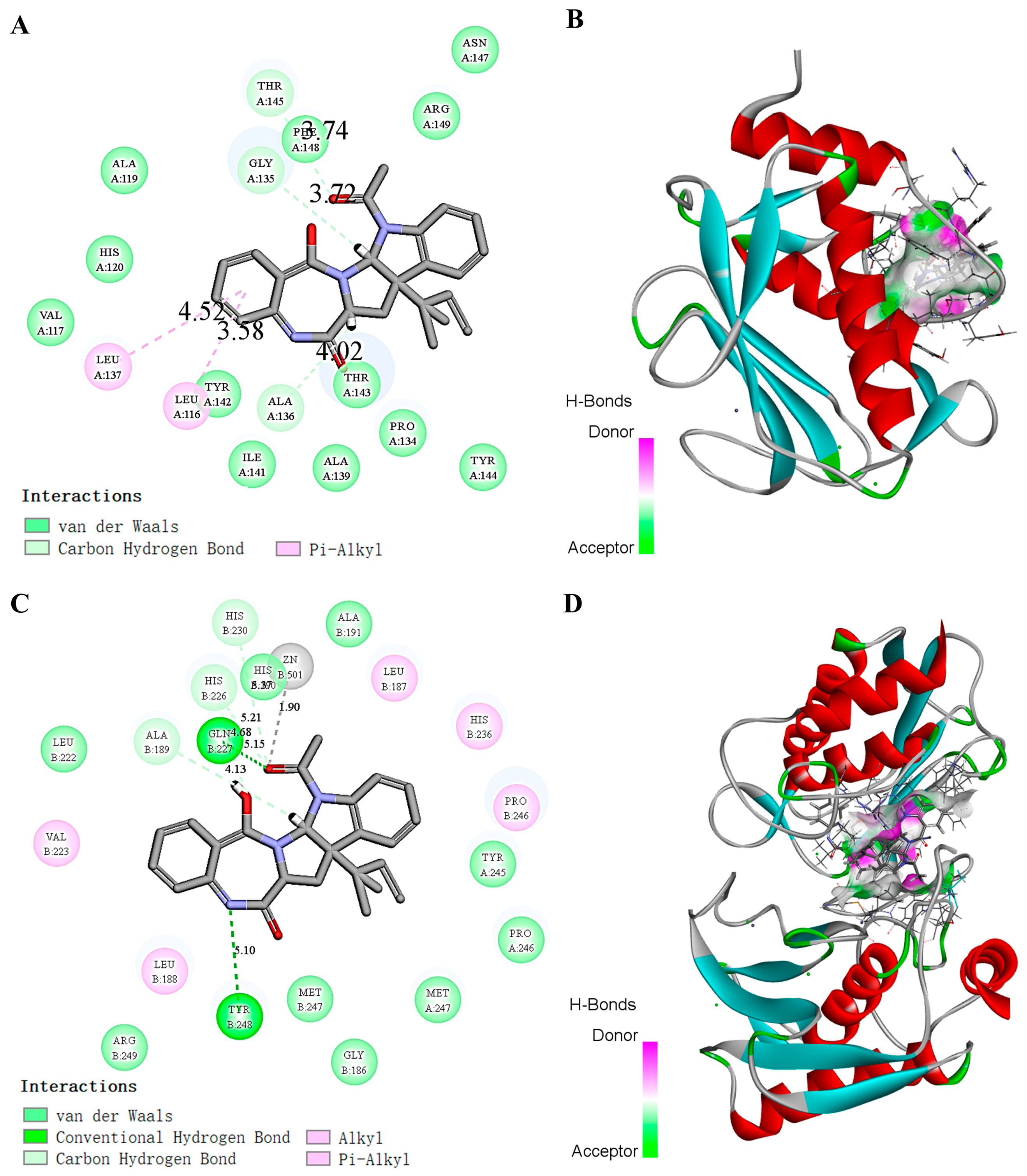
2.8. Molecular Simulation of the Effect of Docking EAA with MMP-2/-9
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Cell Activity Assay (CCK-8)
4.3. Cell Wound Healing Assay
4.4. Cell Invasion by the Transwell Chamber Assay
4.5. Enzyme Linked Immunosorbent Assay (ELISA)
4.6. Western Blot
4.7. Immunocytochemistry
4.8. Molecular Docking
4.9. Statistical Analysis
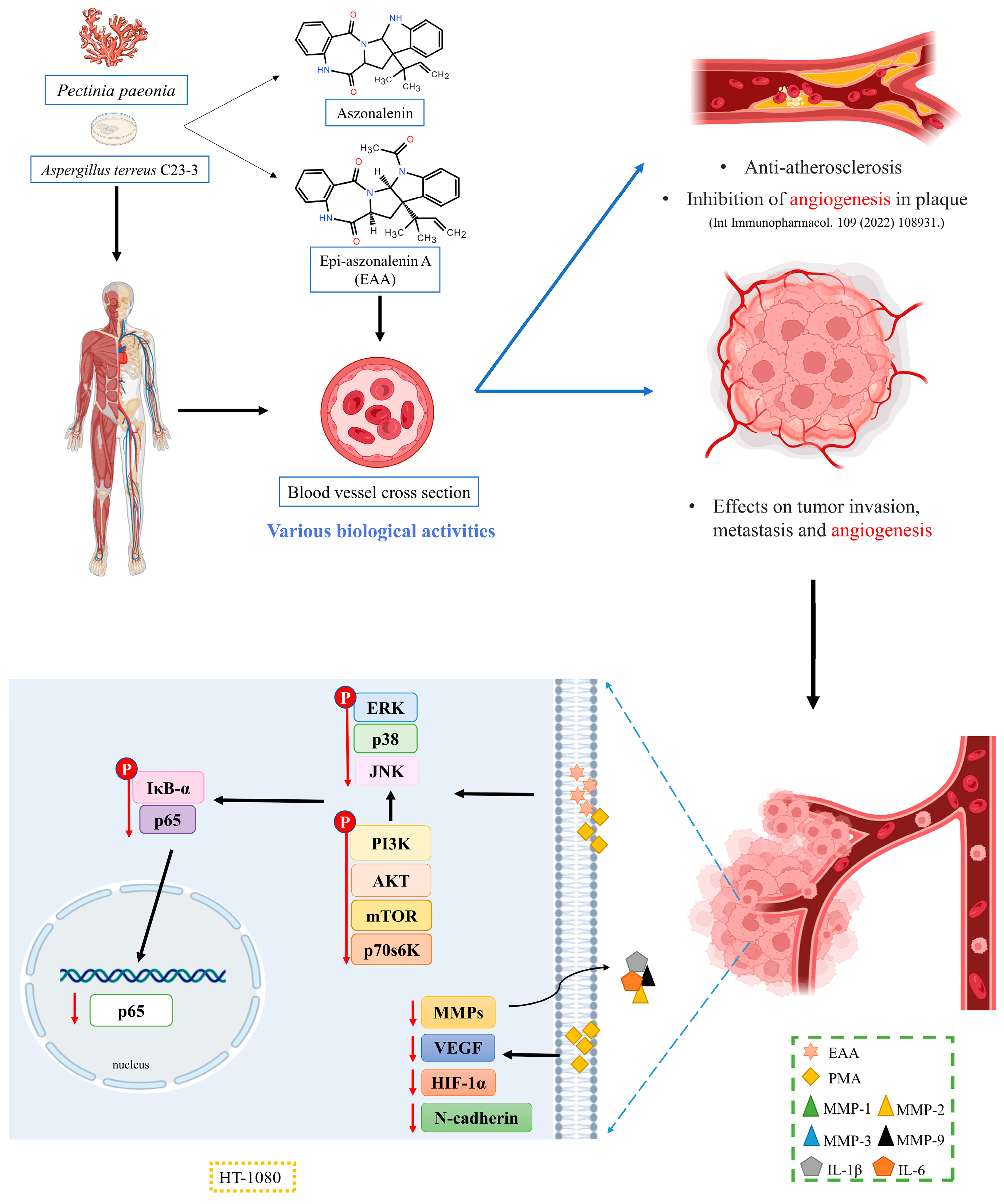
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, W.J.; Gao, J.B. Molecular mechanisms of chemoresistance in gastric cancer. World J. Gastrointest. Oncol. 2016, 8, 673–681. [Google Scholar] [CrossRef]
- Sharma, P.; Allison, J.P. Immune Checkpoint Targeting in Cancer Therapy: Toward Combination Strategies with Curative Potential. Cell 2015, 161, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med. 2019, 8, 5574–5576. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.P.; Massagué, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [Green Version]
- Napoli, C.; Giordano, A.; Casamassimi, A.; Pentimalli, F.; Ignarro, L.J.; De Nigris, F. Directed in vivo angiogenesis assay and the study of systemic neoangiogenesis in cancer. Int. J. Cancer 2011, 128, 1505–1508. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, L.O.; González-Reyes, S.; Marín, L.; González, L.; González, J.M.; Lamelas, M.L.; Merino, A.M.; Rodríguez, E.; Pidal, I.; del Casar, J.M.; et al. Comparative analysis and clinical value of the expression of metalloproteases and their inhibitors by intratumour stromal mononuclear inflammatory cells and those at the invasive front of breast carcinomas. Histopathology 2010, 57, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qiu, P.; Li, J.; Chen, G.; Chen, Y.; Liu, H.; She, Z. Anti-inflammatory polyketides from the mangrove-derived fungus Ascomycota sp. SK2YWS-L. Tetrahedron 2018, 74, 746–751. [Google Scholar] [CrossRef]
- Jamison, M.T.; Molinski, T.F. Antipodal Crambescin A2 Homologues from the Marine Sponge Pseudaxinella reticulata. Antifungal Structure–Activity Relationships. J. Nat. Prod. 2015, 78, 557–561. [Google Scholar] [CrossRef]
- Hasan, S.; Ansari, M.I.; Ahmad, A.; Mishra, M. Major bioactive metabolites from marine fungi: A Review. Bioinformation 2015, 11, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Y.; Chen, M.; Liu, Y.; Liang, J.; Zhang, Y.; Qian, Z.J. Mechanism of two alkaloids isolated from coral endophytic fungus for suppressing angiogenesis in atherosclerotic plaque in HUVEC. Int. Immunopharmacol. 2022, 109, 108931. [Google Scholar] [CrossRef]
- Mischak, H.; Goodnight, J.A.; Kolch, W.; Martiny-Baron, G.; Schaechtle, C.; Kazanietz, M.G.; Blumberg, P.M.; Pierce, J.H.; Mushinski, J.F. Overexpression of protein kinase C-delta and -epsilon in NIH 3T3 cells induces opposite effects on growth, morphology, anchorage dependence, and tumorigenicity. J. Biol. Chem. 1993, 268, 6090–6096. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xiao, Z.; Lin, L.; Tang, Y.; Hong, P.; Sun, S.; Zhou, C.; Qian, Z.-J. Mechanism Analysis of Antiangiogenic d-Isofloridoside from Marine Edible Red algae Laurencia undulata in HUVEC and HT1080 cell. J. Agric. Food Chem. 2021, 69, 13787–13795. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- De Luca, A.; Maiello, M.R.; D’Alessio, A.; Pergameno, M.; Normanno, N. The RAS/RAF/MEK/ERK and the PI3K/AKT signalling pathways: Role in cancer pathogenesis and implications for therapeutic approaches. Expert Opin. Ther. Targets 2012, 16 (Suppl. 2), S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Arkan, M.C.; Greten, F.R. IKK- and NF-κB-mediated functions in carcinogenesis. Curr. Top. Microbiol. Immunol. 2011, 349, 159–169. [Google Scholar] [PubMed]
- Zhao, Y.; Adjei, A.A. Targeting Angiogenesis in Cancer Therapy: Moving Beyond Vascular Endothelial Growth Factor. Oncologist 2015, 20, 660–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efferth, T.; Saeed, M.E.M.; Kadioglu, O.; Seo, E.J.; Shirooie, S.; Mbaveng, A.T.; Nabavi, S.M.; Kuete, V. Collateral sensitivity of natural products in drug-resistant cancer cells. Biotechnol. Adv. 2020, 38, 107342. [Google Scholar] [CrossRef]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [Green Version]
- Tohme, R.; Darwiche, N.; Gali-Muhtasib, H. A journey under the sea: The quest for marine anti-cancer alkaloids. Molecules 2011, 16, 9665–9696. [Google Scholar] [CrossRef]
- Rank, C.; Phipps, R.K.; Harris, P.; Frisvad, J.C.; Gotfredsen, C.H.; Larsen, T.O. epi-Aszonalenins A, B, and C from Aspergillus novofumigatus. Tetrahedron Lett. 2006, 47, 6099–6102. [Google Scholar] [CrossRef]
- Xu, D.; Xu, Z. Indole Alkaloids with Potential Anticancer Activity. Curr. Top. Med. Chem. 2020, 20, 1938–1949. [Google Scholar] [CrossRef] [PubMed]
- Isobe, T.; Aoyagi, K.; Koufuji, K.; Shirouzu, K.; Kawahara, A.; Taira, T.; Kage, M. Clinicopathological significance of hypoxia-inducible factor-1 alpha (HIF-1α) expression in gastric cancer. Int. J. Clin. Oncol. 2013, 18, 293–304. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Hsu, C.C.; Huang, S.F.; Wang, J.S.; Chu, W.K.; Nien, J.E.; Chen, W.S.; Chow, S.E. Interplay of N-Cadherin and matrix metalloproteinase 9 enhances human nasopharyngeal carcinoma cell invasion. BMC Cancer 2016, 16, 800. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.; Frei, R.; Lawson, K.R. The cytoplasmic domain of N-cadherin modulates MMP-9 induction in oral squamous carcinoma cells. Int. J. Oncol. 2014, 45, 1699–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Zhang, Y.; Zhang, W.; Lai, X.; Li, Q.; Zhang, L.; Sun, S. Green tea and black tea inhibit proliferation and migration of HepG2 cells via the PI3K/Akt and MMPs signalling pathway. Biomed. Pharmacother. 2020, 125, 109893. [Google Scholar] [CrossRef]
- Zeng, Z.S.; Cohen, A.M.; Guillem, J.G. Loss of basement membrane type IV collagen is associated with increased expression of metalloproteinases 2 and 9 (MMP-2 and MMP-9) during human colorectal tumorigenesis. Carcinogenesis 1999, 20, 749–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.L.; Dai, N.; Yu, H.G.; Sun, L.M.; Si, J.M. Akt associates with nuclear factor kappaB and plays an important role in chemoresistance of gastric cancer cells. Oncol. Rep. 2010, 24, 113–119. [Google Scholar] [CrossRef]
- Karin, M. Nuclear factor-kappaB in cancer development and progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF-[kappa]B activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef]
- Huang, S.; Pettaway, C.A.; Uehara, H.; Bucana, C.D.; Fidler, I.J. Blockade of NF-kappaB activity in human prostate cancer cells is associated with suppression of angiogenesis, invasion, and metastasis. Oncogene 2001, 20, 4188–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.Q.; Wang, Z.; Leng, P. Aberrant N-cadherin expression in cancer. Biomed. Pharmacother. 2019, 118, 109320. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yang, X.; Geng, M.; Huang, M. Targeting ERK, an Achilles’ Heel of the MAPK pathway, in cancer therapy. Acta Pharm. Sin. B 2018, 8, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Li, X.F.; Jin, L.F.; Zhao, Y.; Zhu, G.J.; Shen, W.Z. Dieckol inhibits non-small-cell lung cancer cell proliferation and migration by regulating the PI3K/AKT signaling pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, W.; Yuan, X.; Yu, L.; Gao, C.; Sun, X.; Wang, D.; Zheng, Q. Licochalcone A-induced human gastric cancer BGC-823 cells apoptosis by regulating ROS-mediated MAPKs and PI3K/AKT signaling pathways. Sci. Rep. 2015, 5, 10336. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tan, L.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.J. Mechanism Analysis of a Novel Angiotensin-I-Converting Enzyme Inhibitory Peptide from Isochrysis zhanjiangensis Microalgae for Suppressing Vascular Injury in Human Umbilical Vein Endothelial Cells. J. Agric. Food Chem. 2020, 68, 4411–4423. [Google Scholar] [CrossRef]
- Zheng, H.; Pei, Y.; Zhou, C.; Hong, P.; Qian, Z.J. Amelioration of atherosclerosis in ox-LDL induced HUVEC by sulfated polysaccharides from Gelidium crinale with antihypertensive activity. Int. J. Biol. Macromol. 2023, 228, 671–680. [Google Scholar] [CrossRef]
- Xiao, Z.; Yang, S.; Chen, J.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.-J. Trehalose against UVB-induced skin photoaging by suppressing MMP expression and enhancing procollagen I synthesis in HaCaT cells. J. Funct. Foods 2020, 74, 104198. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Lin, L.; Zheng, H.; He, Y.-L.; Li, Y.; Zhou, C.; Hong, P.; Sun, S.; Zhang, Y.; Qian, Z.-J. Mechanisms of Antitumor Invasion and Metastasis of the Marine Fungal Derivative Epi-Aszonalenin A in HT1080 Cells. Mar. Drugs 2023, 21, 156. https://doi.org/10.3390/md21030156
Liu Y, Lin L, Zheng H, He Y-L, Li Y, Zhou C, Hong P, Sun S, Zhang Y, Qian Z-J. Mechanisms of Antitumor Invasion and Metastasis of the Marine Fungal Derivative Epi-Aszonalenin A in HT1080 Cells. Marine Drugs. 2023; 21(3):156. https://doi.org/10.3390/md21030156
Chicago/Turabian StyleLiu, Yi, Liyuan Lin, Haiyan Zheng, Yuan-Lin He, Yanmei Li, Chunxia Zhou, Pengzhi Hong, Shengli Sun, Yi Zhang, and Zhong-Ji Qian. 2023. "Mechanisms of Antitumor Invasion and Metastasis of the Marine Fungal Derivative Epi-Aszonalenin A in HT1080 Cells" Marine Drugs 21, no. 3: 156. https://doi.org/10.3390/md21030156
APA StyleLiu, Y., Lin, L., Zheng, H., He, Y. -L., Li, Y., Zhou, C., Hong, P., Sun, S., Zhang, Y., & Qian, Z. -J. (2023). Mechanisms of Antitumor Invasion and Metastasis of the Marine Fungal Derivative Epi-Aszonalenin A in HT1080 Cells. Marine Drugs, 21(3), 156. https://doi.org/10.3390/md21030156