


New Data on the Rhamnose-Binding Lectin from the Colonial Ascidian Botryllus schlosseri: Subcellular Distribution, Secretion Mode and Effects on the Cyclical Generation Change



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
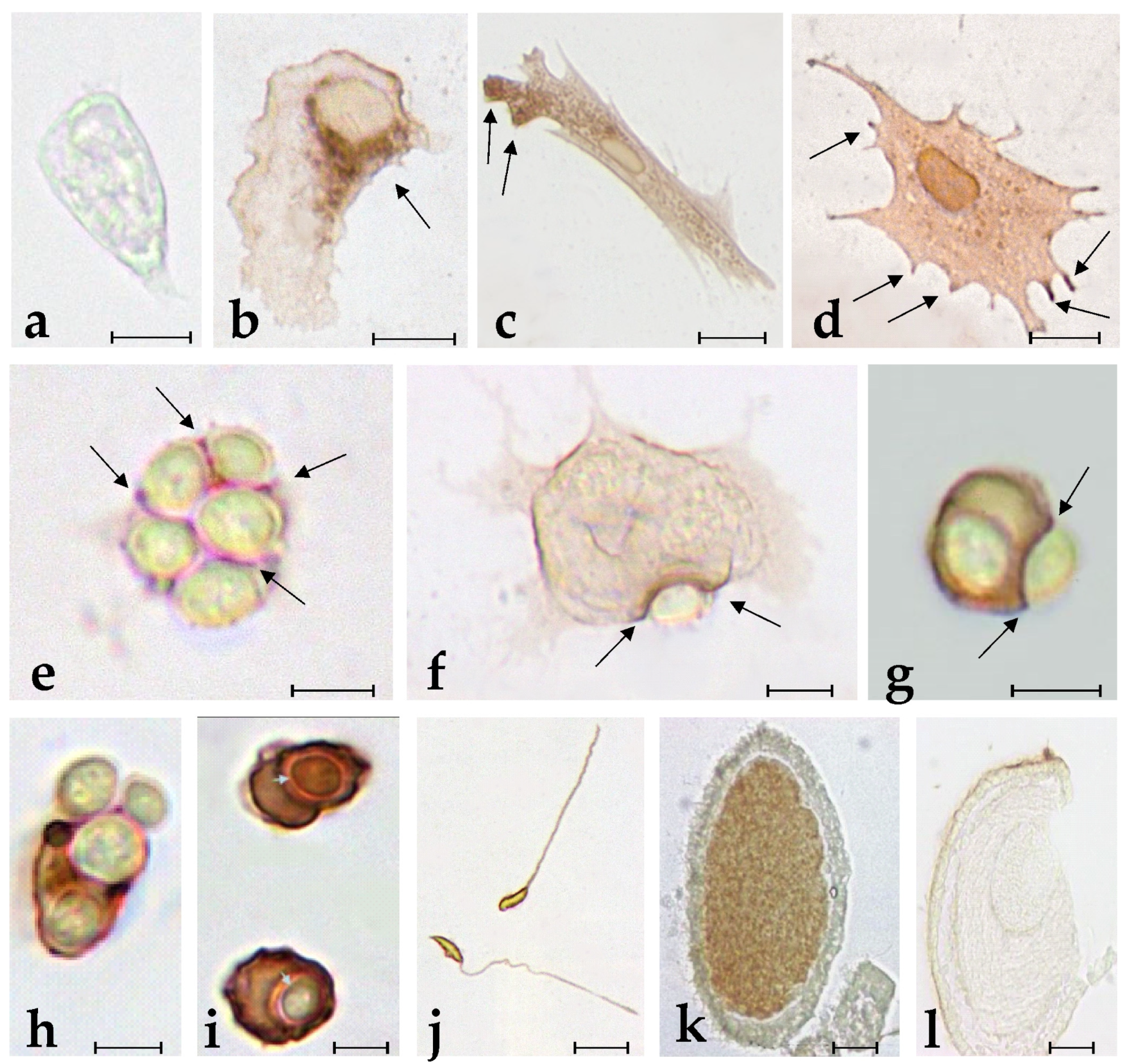
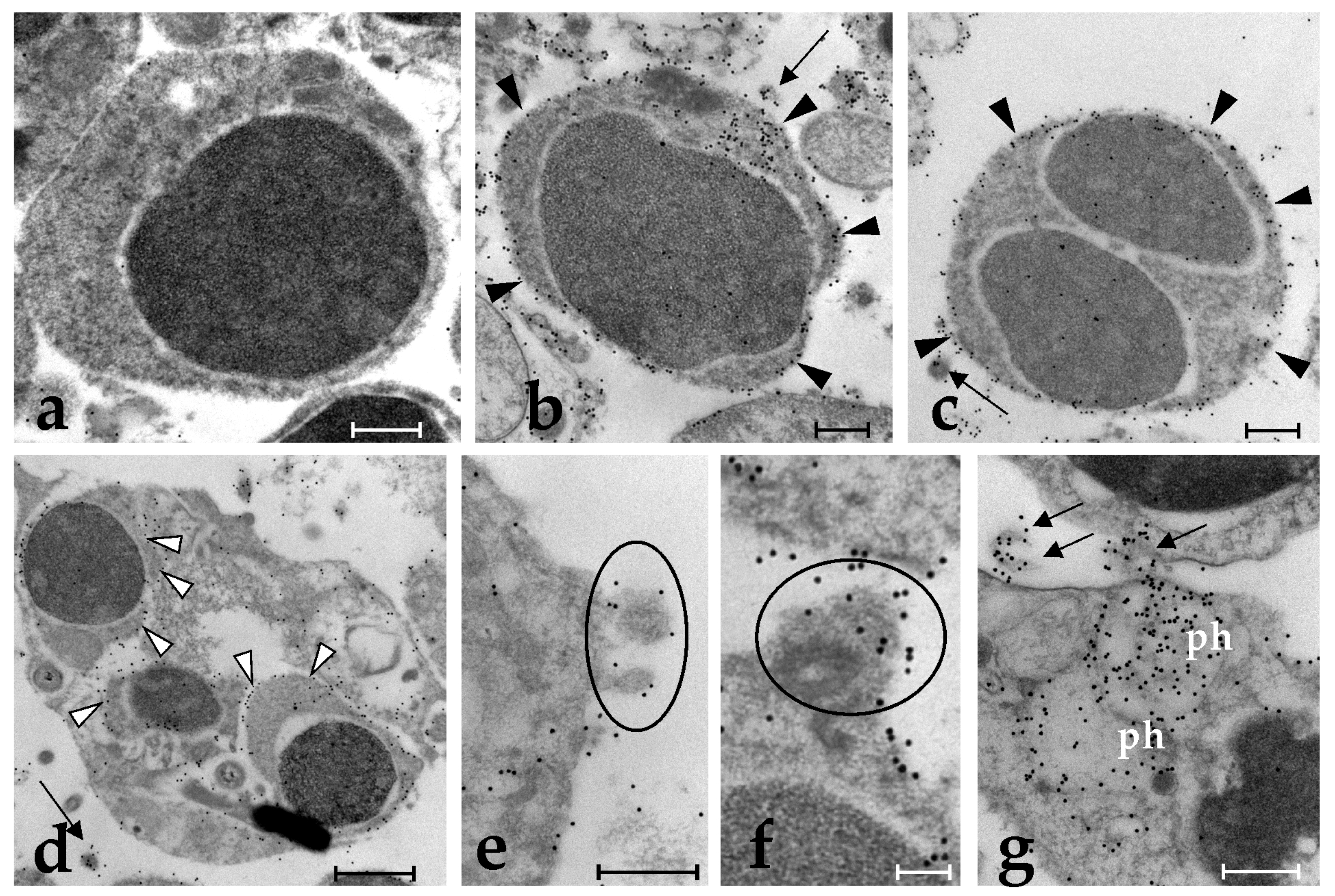
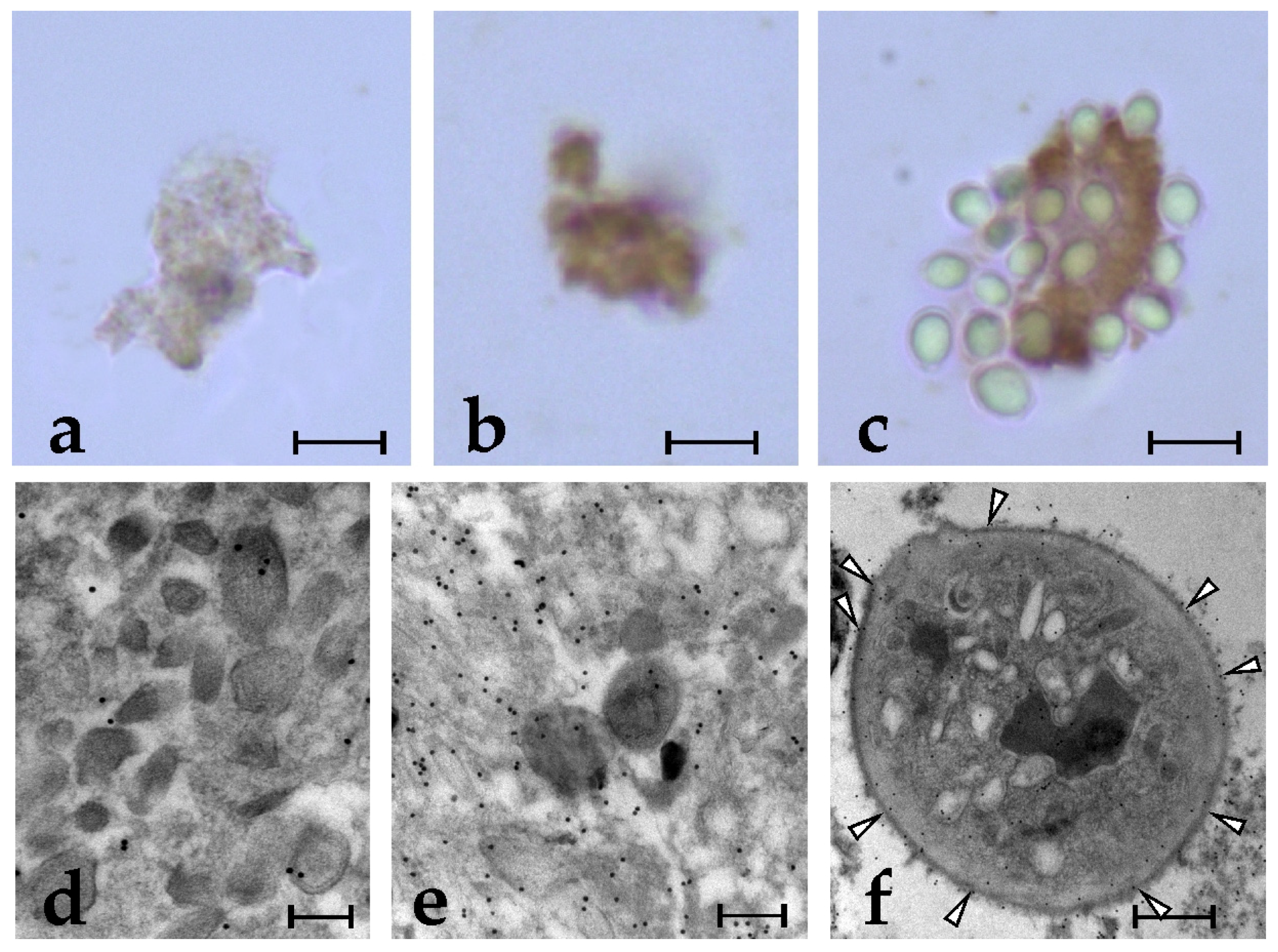
2.1. Specific Anti-BsRBL Antibodies Recognise Agglutinated Yeast Cells, B. schlosseri Phagocytes and Germ Cells
2.2. Anti-BsRBL Antibody Recognises Extracellular Vesicles Released by Phagocytes
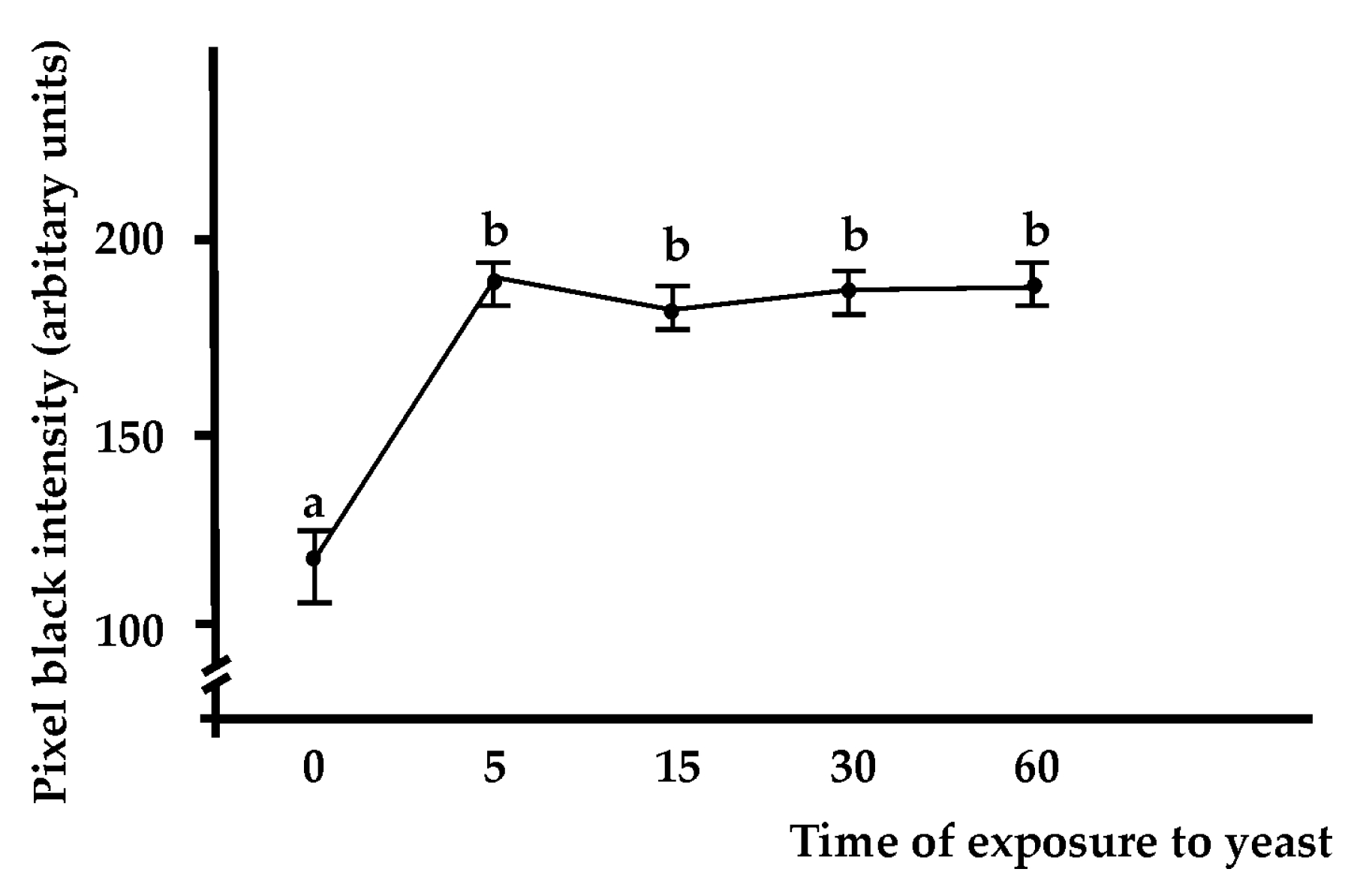
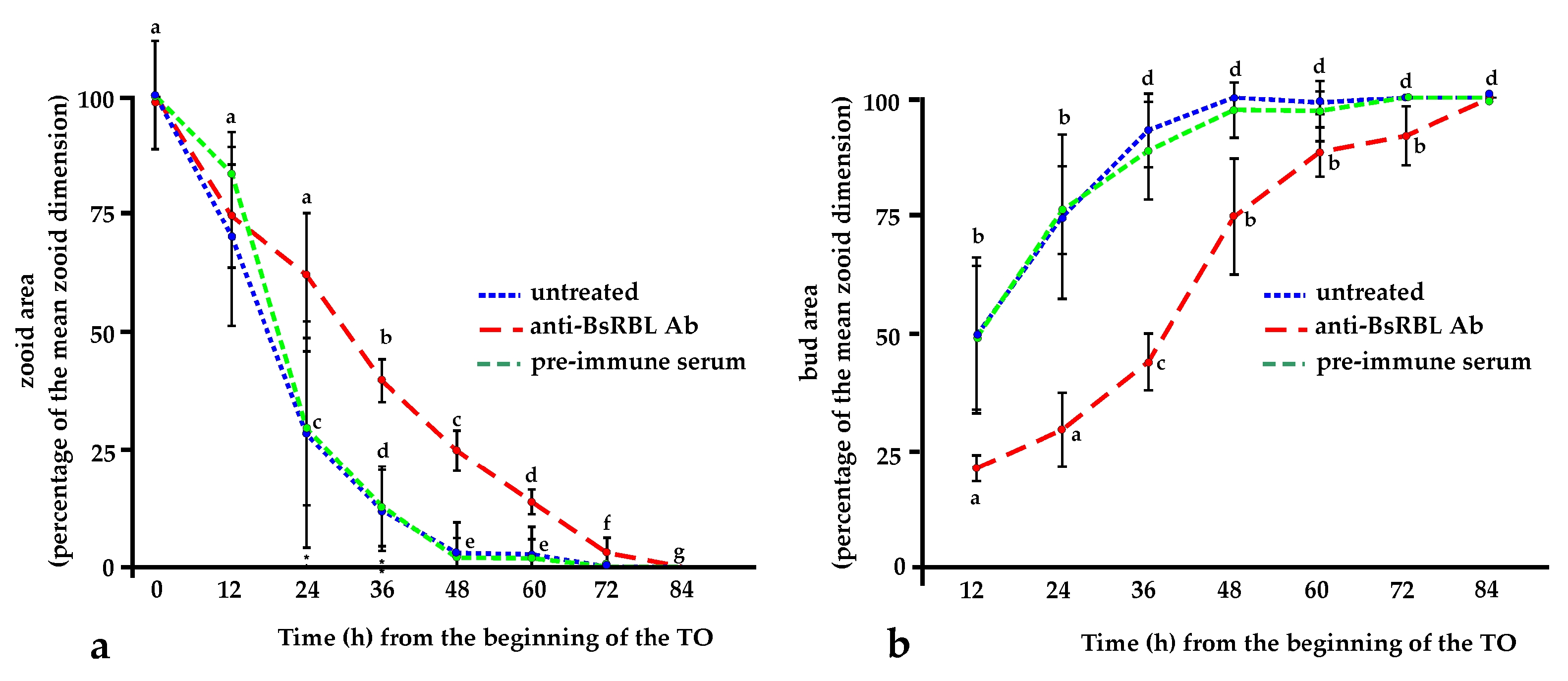
2.3. The Anti-BsRBL Antibody Slows the Progression of the Blastogenetic Cycle
3. Discussion
4. Materials and Methods
4.1. Colonies of B. schlosseri
4.2. Collection of Haemocytes and Phagocytosis Assay
4.3. Immunocytochemistry and Immunohistochemistry
4.4. Extracellular Vesicles Isolation and Characterisation
4.5. Effects of the Anti-BsRBL Antibody on the Blastogenetic Cycle
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kilpatrick, D.C. Animal lectins: A historical introduction and overview. Biochim. Biophys. Acta 2002, 1572, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Watanabe, M.; Naganuma, T.; Muramoto, K. Diversified carbohydrate-binding lectins from marine resources. J. Amino Acids 2011, 2011, 838914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballarin, L.; Cammarata, M.; Franchi, N.; Parrinello, N. Routes in innate immunity evolution: Galectins and rhamnose-binding lectins in ascidians. In Marine Proteins and Peptides: Biological Activities and Applications; Kim, S.-K., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 185–205. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, S.; Pallan, A.R.; Balakrishnan, A.; Paul, E.J.; Elumalai, P. Animal lectin. In Lectins, Innate Immune Defense and Therapeutics; Elumalai, P., Lakshmi, S., Eds.; Springer Nature: Singapore, 2021; pp. 89–106. [Google Scholar] [CrossRef]
- Radhakrishnan, A.; Chellapandian, H.; Ramasamy, P.; Jeyachandran, S. Back2Basics: Animal lectins: An insight into a highly versatile recognition protein. J. Proteins Proteom. 2023, 14, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Kaltner, H.; Stierstorfer, B. Animal lectins as cell adhesion molecules. Acta Anat. 1998, 161, 162–179. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, J.; Pang, X.; Liu, T.; Ning, Z.; Cheng, G. The roles of direct recognition by animal lectins in antiviral immunity and viral pathogenesis. Molecules 2015, 20, 2272–2295. [Google Scholar] [CrossRef] [Green Version]
- Vasta, G.R.; Quesenberry, M.S.; Hafiz, A.; O’Leary, N. Lectins from tunicates: Structure-function relationships in innate immunity. In Phylogenetic Perspectives on the Vertebrate Immune System. Advances in Experimental Medicine and Biology; Beck, G., Sugumaran, M., Cooper, E.L., Eds.; Springer: Boston, MA, USA, 2001; Volume 484, pp. 275–287. [Google Scholar] [CrossRef]
- Quesenberry, M.S.; Ahmed, H.; Elola, M.T.; O’Leary, N.; Vasta, G.R. Diverse lectin repertoires in tunicates mediate broad recognition and effector innate immune responses. Integr. Comp. Biol. 2003, 43, 323–330. [Google Scholar] [CrossRef]
- Lam, Y.W.; Ng, T.B. Purification and characterization of a rhamnose-binding lectin with immunoenhancing activity from grass carp (Ctenopharyngodon idellus) ovaries. Protein Expr. Purif. 2002, 26, 378–385. [Google Scholar] [CrossRef]
- Tateno, H.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Saneyoshi, M. Rhamnose binding lectins from steelhead trout (Oncorhynchus mykiss) eggs recognize bacterial lipopolysaccharides and lipoteichoic acid. Biosci. Biotechnol. Biochem. 2002, 66, 604–612. [Google Scholar] [CrossRef] [Green Version]
- Tateno, H.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Saneyoshi, M. Distribution and molecular evolution of rhamnose-binding lectins in Salmonidae: Isolation and characterization of two lectins from white-spotted charr (Salvelinus leucomaenis) eggs. Biosci. Biotechnol. Biochem. 2002, 66, 1356–1365. [Google Scholar] [CrossRef]
- Terada, T.; Watanabe, Y.; Tateno, H.; Naganuma, T.; Ogawa, T.; Muramoto, K.; Kamiya, H. Structural characterization of a rhamnose binding glycoprotein (lectin) from Spanish mackerel (Scomberomorous niphonius) eggs. Biochim. Biophys. Acta 2007, 1770, 617–629. [Google Scholar] [CrossRef]
- Hosono, M.; Matsuda, K.; Kawauchi, H.; Takayanagi, Y.; Shiokawa, H.; Mineki, R.; Murayama, K.; Nitta, K. Comparison of N-terminal amino acid sequence of fish roe rhamnose-binding lectins. Biomed. Res. 1992, 13, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Tateno, H.; Nakamura-Tsuruta, S.; Kominami, J.; Hirabayashi, J.; Nakamura, O.; Watanabe, T.; Kamiya, H.; Naganuma, T.; Ogawa, T.; et al. The function of rhamnose-binding lectin in innate immunity by restricted binding to Gb3. Dev. Comp. Immunol. 2009, 33, 187–197. [Google Scholar] [CrossRef]
- Cammarata, M.; Parisi, M.G.; Benenati, G.; Vasta, G.R.; Parrinello, N. A rhamnose-binding lectin from sea bass (Dicentrarchus labrax) plasma agglutinates and opsonizes pathogenic bacteria. Dev. Comp. Immunol. 2014, 44, 332–340. [Google Scholar] [CrossRef]
- Gao, C.; Su, B.; Zhang, D.; Yang, N.; Song, L.; Fu, Q.; Zhou, S.; Tan, F.; Li, C. L-rhamnose-binding lectins (RBLs) in turbot (Scophthalmus maximus L.): Characterization and expression profiling in mucosal tissues. Fish Shellfish Immunol. 2018, 80, 264–273. [Google Scholar] [CrossRef]
- Ozeki, Y.; Matsui, T.; Suzuki, M.; Titani, K. Amino acid sequence and molecular characterization of a D-galactoside-specific lectin purified from sea urchin (Anthocidaris crassispina) eggs. Biochemistry 1991, 30, 2391–2394. [Google Scholar] [CrossRef]
- Naganuma, T.; Ogawa, T.; Hirabayashi, J.; Kasai, K.; Kamiya, H.; Muramoto, K. Isolation, characterization and molecular evolution of a novel pearl shell lectin from a marine bivalve, Pteria penguin. Mol. Divers. 2006, 10, 607–618. [Google Scholar] [CrossRef]
- Carneiro, R.F.; Teixeira, C.S.; de Melo, A.A.; de Almeida, A.S.; Cavada, B.S.; de Sousa, O.V.; da Rocha, B.A.; Nagano, C.S.; Sampaio, A.H. L-Rhamnose-binding lectin from eggs of the Echinometra lucunter: Amino acid sequence and molecular modeling. Int. J. Biol. Macromol. 2015, 78, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, T.; Ichise, A.; Unno, H.; Goda, S.; Oda, T.; Tateno, H.; Hirabayashi, J.; Sakai, H.; Nakagawa, H. Carbohydrate recognition by the rhamnose-binding lectin SUL-I with a novel three-domain structure isolated from the venom of globiferous pedicellariae of the flower sea urchin Toxopneustes pileolus. Protein Sci. 2017, 26, 1574–1583. [Google Scholar] [CrossRef] [Green Version]
- Araújo, C.A.C.; Pacheco, J.P.F.; Waniek, P.J.; Geraldo, R.B.; Sibajev, A.; Dos Santos, A.L.; Evangelho, V.G.O.; Dyson, P.J.; Azambuja, P.; Ratcliffe, N.A.; et al. A rhamnose-binding lectin from Rhodnius prolixus and the impact of its silencing on gut bacterial microbiota and Trypanosoma cruzi. Dev. Comp. Immunol. 2021, 114, 103823. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L. Putative rhamnose-binding lectin in the solitary ascidian Ciona intestinalis. ISJ—Invertebr. Surviv. J. 2013, 10, 18. [Google Scholar]
- Gasparini, F.; Franchi, N.; Spolaore, B.; Ballarin, L. Novel rhamnose-binding lectins from the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2008, 32, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Schiavon, F.; Carletto, M.; Gasparini, F.; Bertoloni, G.; Tosatto, S.C.; Ballarin, L. Immune roles of a rhamnose-binding lectin in the colonial ascidian Botryllus schlosseri. Immunobiology 2011, 216, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Manni, L.; Zaniolo, G.; Cima, F.; Burighel, P.; Ballarin, L. Botryllus schlosseri: A model ascidian for the study of asexual reproduction. Dev. Dyn. 2007, 236, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Ballarin, L.; Tonello, C.; Sabbadin, A. Humoral opsonin from the colonial ascidian Botryllus schlosseri as a member of the galectin family. Mar. Biol. 2000, 136, 813–822. [Google Scholar] [CrossRef]
- Rudd, P.M.; Elliott, T.; Cresswell, P.; Wilson, I.A.; Dwek, R.A. Glycosylation and the immune system. Science 2001, 291, 2370–2376. [Google Scholar] [CrossRef]
- Marth, J.; Grewal, P. Mammalian glycosylation in immunity. Nat. Rev. Immunol. 2008, 8, 874–887. [Google Scholar] [CrossRef] [Green Version]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Šebestík, J.; Reiniš, M.; Ježek, J. Sugar code (glycocode). In Biomedical Applications of Peptide-, Glyco- and Glycopeptide Dendrimers, and Analogous Dendrimeric Structures; Sebestik, J., Reinis, M., Jezek, J., Eds.; Springer: Vienna, Austria, 2012; pp. 23–27. [Google Scholar] [CrossRef]
- Rodríguez, E.; Schetters, S.; van Kooyk, Y. The tumour glyco-code as a novel immune checkpoint for immunotherapy. Nat. Rev. Immunol. 2018, 18, 204–211. [Google Scholar] [CrossRef]
- Lepenies, B.; Lang, R. Editorial: Lectins and their ligands in shaping immune responses. Front. Immunol. 2019, 10, 2379. [Google Scholar] [CrossRef]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [Green Version]
- Ballarin, L.; Tonello, C.; Guidolin, L.; Sabbadin, A. Purification and characterization of a humoral opsonin, with specificity for D-galactose, in the colonial ascidian Botryllus schlosseri. Comp. Biochem. Physiol. 1999, 123B, 115–123. [Google Scholar] [CrossRef]
- Ballarin, L.; Menin, A.; Franchi, N.; Bertoloni, G.; Cima, F. Morula cells and non-self recognition in the compound ascidian Botryllus schlosseri. ISJ—Invertebr. Surviv. J. 2005, 2, 1–5. [Google Scholar]
- Menin, A.; Del Favero, M.; Cima, F.; Ballarin, L. Release of phagocytosis-stimulating factor(s) by morula cells in a colonial ascidian. Mar. Biol. 2005, 148, 225–230. [Google Scholar] [CrossRef]
- Menin, A.; Ballarin, L. Immunomodulatory molecules in the compound ascidian Botryllus schlosseri: Evidence from conditioned media. J. Invertebr. Pathol. 2008, 99, 275–280. [Google Scholar] [CrossRef]
- Watanabe, Y.; Shiina, N.; Shinozaki, F.; Yokoyama, H.; Kominami, J.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Sugahara, K.; Kamiya, H.; Matsubara, H.; et al. Isolation and characterization of l-rhamnose-binding lectin, which binds to microsporidian Glugea plecoglossi, from ayu (Plecoglossus altivelis) eggs. Dev. Comp. Immunol. 2008, 32, 487–499. [Google Scholar] [CrossRef]
- Hosono, M.; Sugawara, S.; Tatsuta, T.; Hikita, T.; Kominami, J.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Kawsar, S.M.; Ozeki, Y.; Hakomori, S.I.; et al. Domain composition of rhamnose-binding lectin from shishamo smelt eggs and its carbohydrate-binding profiles. Fish Physiol. Biochem. 2013, 39, 1619–1630. [Google Scholar] [CrossRef]
- Shiina, N.; Tateno, H.; Ogawa, T.; Muramoto, K.; Saneyoshi, M.; Kamiya, H. Isolation and characterization of L-rhamnose-binding lectins from chum salmon (Oncorhynchus keta) eggs. Fish. Sci. 2002, 68, 1352–1366. [Google Scholar] [CrossRef]
- Ozeki, Y.; Yokota, Y.; Kato, K.H.; Titani, K.; Matsui, T. Developmental expression of D-galactoside-binding lectin in sea urchin (Anthocidaris crassispina) eggs. Exp. Cell Res. 1995, 216, 318–324. [Google Scholar] [CrossRef]
- Cima, F.; Manni, L.; Basso, G.; Fortunato, E.; Accordi, B.; Schiavon, F.; Ballarin, L. Hovering between death and life: Haemocytes and natural apoptosis in the blastogenetic cycle of the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2010, 34, 272–285. [Google Scholar] [CrossRef]
- Franchi, N.; Ballin, F.; Manni, L.; Schiavon, F.; Basso, G.; Ballarin, L. Recurrent phagocytosis-induced apoptosis in the cyclical generation change of the compound ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2016, 62, 8–16. [Google Scholar] [CrossRef]
- Voskoboynik, A.; Rinkevich, B.; Weiss, A.; Moiseeva, E.; Reznick, A.Z. Macrophage involvement for successful degeneration of apoptotic organs in the colonial urochordate Botryllus schlosseri. J. Exp. Biol. 2004, 207, 2409–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bovo, G.; Ballarin, L. New Data on the Rhamnose-Binding Lectin from the Colonial Ascidian Botryllus schlosseri: Subcellular Distribution, Secretion Mode and Effects on the Cyclical Generation Change. Mar. Drugs 2023, 21, 171. https://doi.org/10.3390/md21030171
Bovo G, Ballarin L. New Data on the Rhamnose-Binding Lectin from the Colonial Ascidian Botryllus schlosseri: Subcellular Distribution, Secretion Mode and Effects on the Cyclical Generation Change. Marine Drugs. 2023; 21(3):171. https://doi.org/10.3390/md21030171
Chicago/Turabian StyleBovo, Giacomo, and Loriano Ballarin. 2023. "New Data on the Rhamnose-Binding Lectin from the Colonial Ascidian Botryllus schlosseri: Subcellular Distribution, Secretion Mode and Effects on the Cyclical Generation Change" Marine Drugs 21, no. 3: 171. https://doi.org/10.3390/md21030171
APA StyleBovo, G., & Ballarin, L. (2023). New Data on the Rhamnose-Binding Lectin from the Colonial Ascidian Botryllus schlosseri: Subcellular Distribution, Secretion Mode and Effects on the Cyclical Generation Change. Marine Drugs, 21(3), 171. https://doi.org/10.3390/md21030171