


Structural Characterization and Effects on Insulin Resistance of a Novel Chondroitin Sulfate from Halaelurus burgeri Skin


Abstract
:1. Introduction
2. Results and Discussion
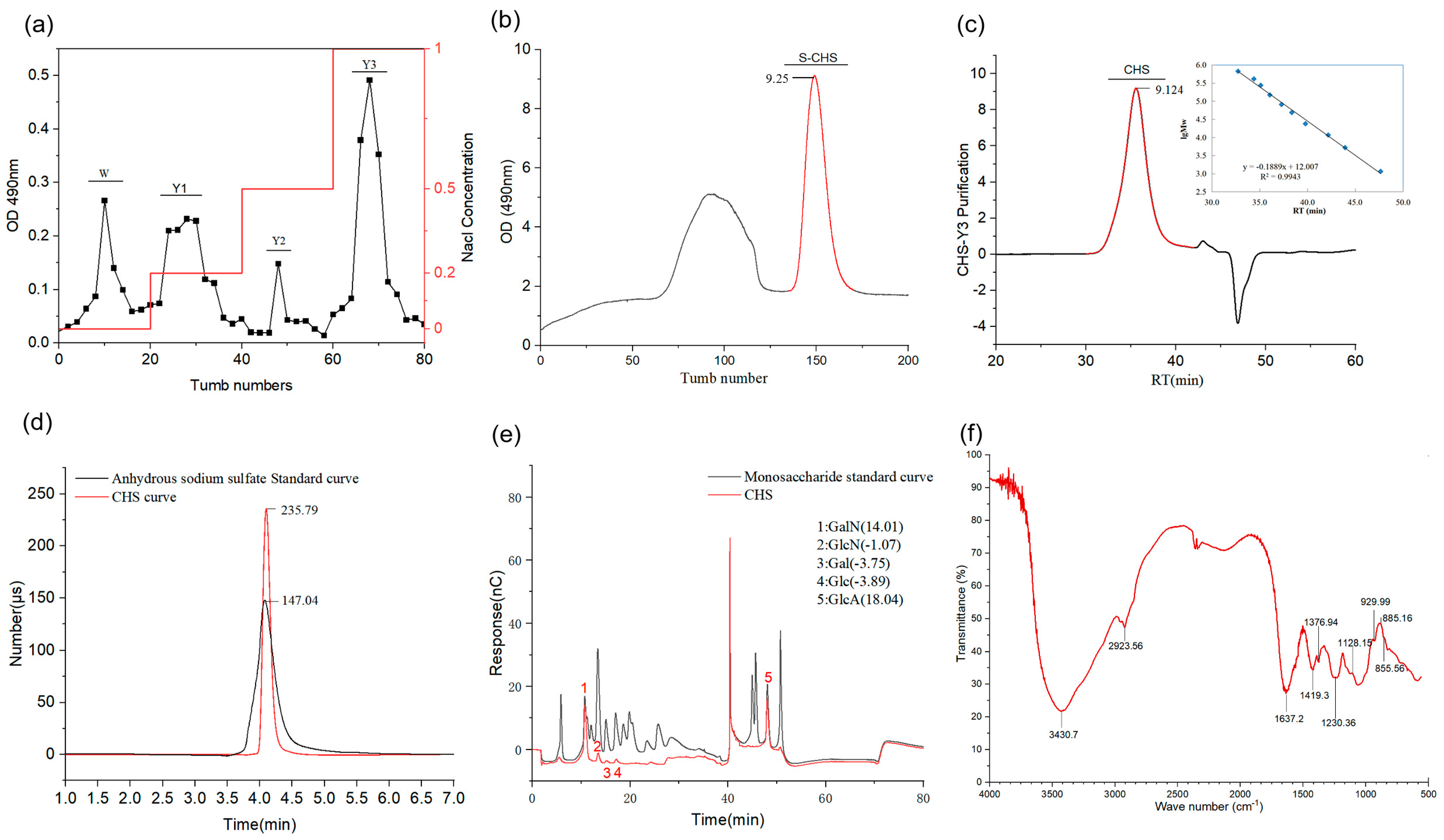
2.1. Isolation and Purification of CHS and General Property Description
2.2. FT-IR Spectroscopy Analysis
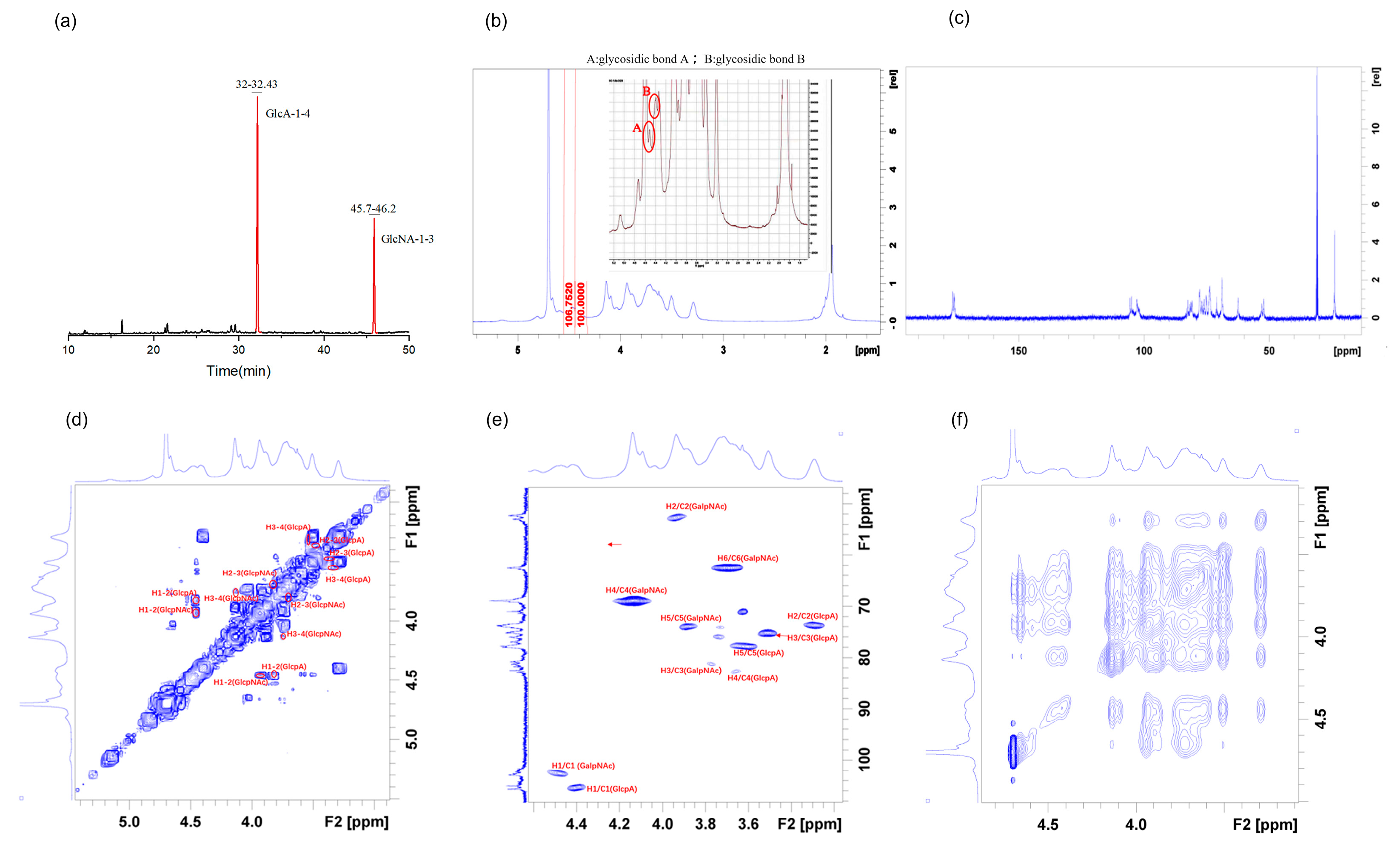
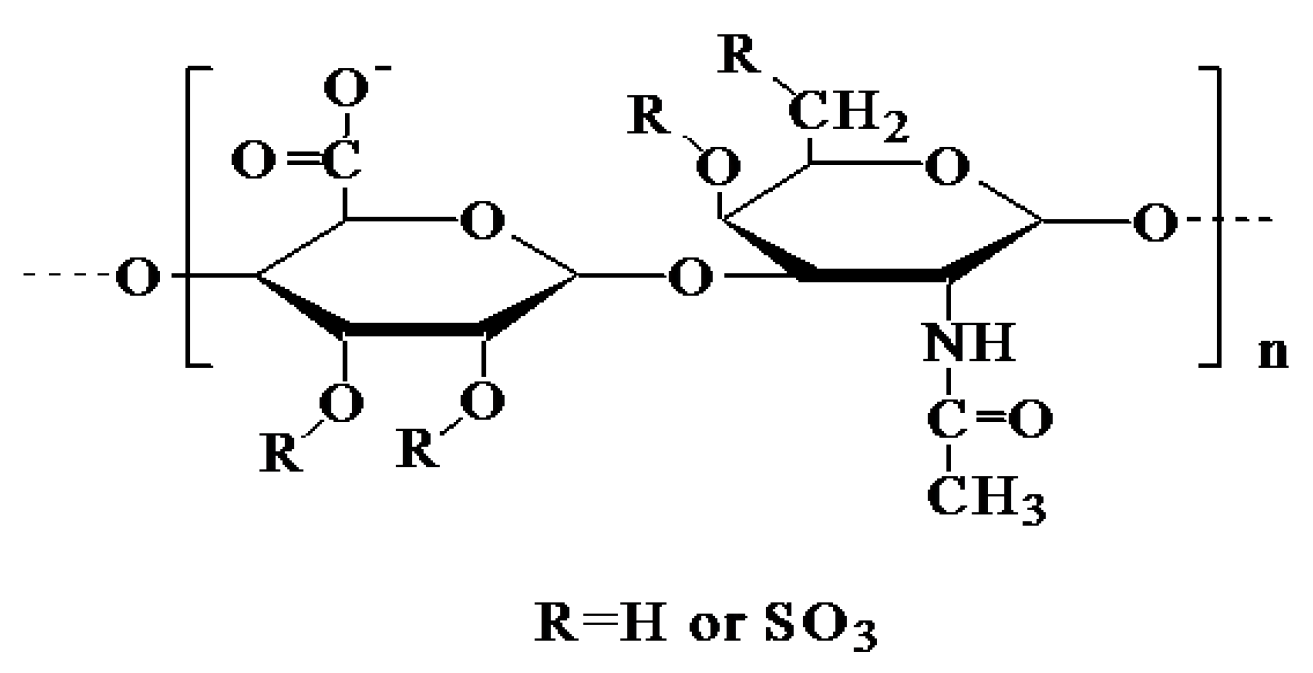
2.3. Methylation and NMR Spectroscopy Analysis
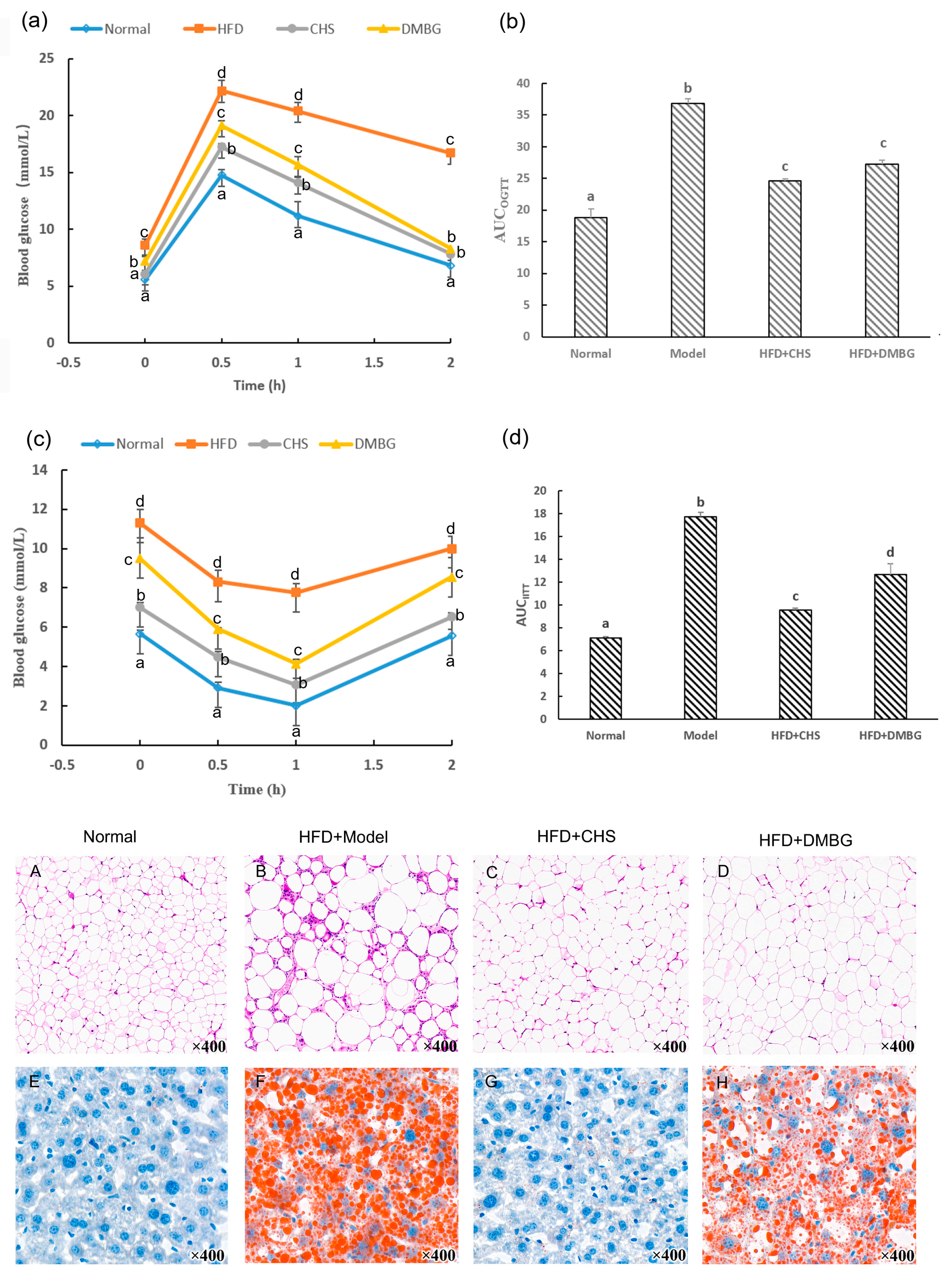
2.4. CHS Reduced Body Weight
2.5. CHS Decreased Blood Glucose, Insulin, and Improvement of Glucose Tolerance and Insulin Tolerance
2.6. CHS Reduces the Abnormal Lipids in Serum and in the Liver
2.7. CHS Attenuated Circulatory Inflammatory Cytokines
3. Materials and Methods
3.1. Extraction and Purification of the CHS
3.2. Determination of General Properties
3.3. FT-IR Spectroscopy Analysis
3.4. Methylation and NMR Analysis
3.5. Animal Experiments
3.6. Insulin-Resistance-Related Parameters in Blood and Liver Measurement
3.7. OGTT and IITT
3.8. H&E and Oil Red O Strain
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Treiber, G.; Guilleux, A.; Huynh, K.; Bonfanti, O.; Flaus-Furmaniuk, A.; Couret, D.; Mellet, N.; Bernard, C.; Le-Moullec, N.; Doray, B.; et al. Lipoatrophic diabetes in familial partial lipodystrophy type 2: From insulin resistance to diabetes. Diabetes Metab. 2023, 49, 101409. [Google Scholar] [CrossRef] [PubMed]
- Roden, M.; Shulman, G.I. The integrative biology of type 2 diabetes. Nature 2019, 576, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.; Chen, C.; Kong, X.; Xia, Z.; Kong, W.; Si, K.; Han, P.; Liu, W.V.; Li, X. Pancreatic fat infiltration, β-cell function and insulin resistance: A study of the young patients with obesity. Diabetes Res. Clin. Pract. 2022, 187, 109860. [Google Scholar] [CrossRef] [PubMed]
- Lydia, A.B.; Du, T. Long-term effects of insulin resistance on appendicular lean muscle. J. Diabetes Complicat. 2019, 33, 13–14. [Google Scholar]
- López-Álvarez, M.; González, P.; Serra, J.; Fraguas, J.; Valcarcel, J.; Vázquez, J.A. Chondroitin sulfate and hydroxyapatite from Prionace glauca shark jaw: Physicochemical and structural characterization. Int. J. Biol. Macromol. 2020, 156, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Meng, Y.; Li, J.; Chen, J.; Liu, Y.; Bai, X. Chondroitin sulfate: Extraction, purification, microbial and chemical synthesis. J. Chem. Technol. Biotechnol. 2014, 89, 1445–1465. [Google Scholar] [CrossRef]
- Urbi, Z.; Azmi, N.S.; Ming, L.C.; Hossain, M.S. A concise review of extraction and characterization of chondroitin sulphate from fish and fish wastes for pharmacological application. Curr. Issues Mol. Biol. 2022, 44, 3905–3922. [Google Scholar] [CrossRef]
- Chen, J.; Chen, X.; Li, J.; Luo, B.; Fan, T.; Li, R.; Liu, X.; Song, B.; Jia, X.; Zhong, S. Preparation and characterization of nano-selenium decorated by chondroitin sulfate derived from shark cartilage and investigation on its antioxidant activity. Mar. Drugs 2022, 20, 172. [Google Scholar] [CrossRef]
- Chilkunda, D.N.; Itoh, N.; Sugahara, K. Novel 70-kDa chondroitin sulfate/dermatan sulfate hybrid chains with a unique heterogenous sulfation pattern from shark skin, which exhibit neuritogenic activity and binding activities for growth factors and neurotrophic factors. J. Biol. Chem. 2005, 280, 4058–4069. [Google Scholar]
- Restaino, O.F.; Schiraldi, C. Chondroitin sulfate: Are the purity and the structural features well assessed? A review on the analytical challenges. Carbohyd. Polym. 2022, 292, 119690. [Google Scholar] [CrossRef]
- Liang, T.; Zhang, Z.; Jing, P. Black rice anthocyanins embedded in self-assembled chitosan/chondroitin sulfate nanoparticles enhance apoptosis in HCT-116 cells. Food Chem. 2019, 301, 125280. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Ren, P.; Wu, Q.; Liu, X.; Zhang, Z.; Hua, M.; Xia, W.; Chen, J. Structure comparison and anti-coagulant effects of the glycosaminoglycans from porcine duodenum and jejunum. Food Chem. 2022, 373, 131609. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Qi, L.; Zhao, L.; Liu, J.; Guo, Y.; Zhang, C. Degradation of chondroitin sulfate: Mechanism of degradation, influence factors, structure-bioactivity relationship and application. Carbohyd. Polym. 2023, 301, 120361. [Google Scholar] [CrossRef] [PubMed]
- Leakey, J.E.A.; Ali, A.A.; Babb, A.R.; Badgley, H.L.; Davis, K.J.; Juliar, B.E.; Leakey, T.I.; Lewis, S.M.; Patton, R.E.; Seng, J.E. Subchronic toxicity evaluation of glucosamine and glucosamine in combination with chondroitin sulfate in obese Zucker rats. Toxicol. Appl. Pharmacol. 2021, 412, 115371. [Google Scholar] [CrossRef] [PubMed]
- Cañas, N.; Valero, T.; Montell, E.; Verges, J.; López, M.G. P2–055: Chondroitin sulphate promotes neuroprotection against free radical–induced damage in vitro. Alzheimers Dement. 2006, 2, S248. [Google Scholar] [CrossRef]
- Krichen, F.; Volpi, N.; Sila, A.; Maccari, F.; Mantovani, V.; Galeotti, F.; Ellouz-Chaabouni, S.; Bougatef, A. Purification, structural characterization and antiproliferative properties of chondroitin sulfate/dermatan sulfate from tunisian fish skins. Int. J. Biol. Macromol. 2017, 95, 32–39. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, S.; Ren, S.; Zhang, X.; Yang, F.; Chen, Y.; Wang, B. Structural characterization and proliferation activity of chondroitin sulfate from the sturgeon, Acipenser schrenckii. Int. J. Biol. Macromol. 2020, 164, 3005–3011. [Google Scholar] [CrossRef]
- Yang, J.; Shen, M.; Wu, T.; Chen, X.; Wen, H.; Xie, J. Physicochemical, structural characterization, and antioxidant activities of chondroitin sulfate from Oreochromis niloticus bones. Food Sci. Hum. Wellness 2023, 12, 1102–1108. [Google Scholar] [CrossRef]
- Dong, F.; Quan, X.; Wang, Q.; Liu, Z.; Cui, T.; Wang, W.; Tang, D.; Zhang, R.; Zhang, C.; Wang, H.; et al. Purification, structural characterization, and anticoagulant activity evaluation of chondroitin sulfate from codfish (Gadus macrocephalus) bones. Int. J. Biol. Macromol. 2022, 210, 759–767. [Google Scholar] [CrossRef]
- Shionoya, K.; Suzuki, T.; Takada, M.; Sato, K.; Onishi, S.; Dohmae, N.; Nishino, K.; Wada, T.; Linhardt, R.J.; Toida, T.; et al. Comprehensive analysis of chondroitin sulfate and aggrecan in the head cartilage of bony fishes: Identification of proteoglycans in the head cartilage of sturgeon. Int. J. Biol. Macromol. 2022, 208, 333–342. [Google Scholar] [CrossRef]
- Meng, D.; Li, W.; Leng, X.; Takagi, Y.; Dai, Z.; Du, H.; Wei, Q. Extraction of chondroitin sulfate and type II collagen from sturgeon (Acipenser gueldenstaedti) notochord and characterization of their hybrid fibrils. Process Biochem. 2023, 124, 180–188. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Fraguas, J.; Novoa-Carballal, R.; Reis, R.L.; Pérez-Martín, R.I.; Valcarcel, J. Optimal isolation and characterisation of chondroitin sulfate from rabbit fish (Chimaera monstrosa). Carbohydr. Polym. 2019, 210, 302–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettolino, F.A.; Walsh, C.; Fincher, G.B.; Bacic, A. Adelaide Research and Scholarship: Determining the polysaccharide composition of plant cell walls. Nat. Prot. 2012, 20, 172. [Google Scholar]
- Nie, K.; Gao, Y.; Chen, S.; Wang, Z.; Wang, H.; Tang, Y.; Su, H.; Lu, F.; Dong, H.; Fang, K. Diosgenin attenuates non-alcoholic fatty liver disease in type 2 diabetes through regulating SIRT6-related fatty acid uptake. Phytomedicine 2023, 111, 154661. [Google Scholar] [CrossRef]
- Afrin, S.; Sabah, M.E.I.; Manzoor, A.; Miyashita-Ishiwata, M.; Reschke, L.; Borahay, M.A. Adipocyte coculture induces a pro-inflammatory, fibrotic, angiogenic, and proliferative microenvironment in uterine leiomyoma cells. BBA Mol. Basis Dis. 2022, 1869, 166564. [Google Scholar] [CrossRef]
- Esmailidehaj, M.; Kuchakzade, F.; Rezvani, M.E.; Farhadi, Z.; Esmaeili, H.; Azizian, H. 17β-Estradiol improves insulin signalling and insulin resistance in the aged female hearts: Role of inflammatory and anti-inflammatory cytokines. Life Sci. 2020, 253, 117673. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gao, Y.; Zhao, Y.; Yang, G.; Wang, C.; Zhao, Z.; Li, S. Chondroitin sulfate stimulates the secretion of H2S by Desulfovibrio to improve insulin sensitivity in NAFLD mice. Int. J. Biol. Macromol. 2022, 213, 631–638. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.; Wang, H.; Fu, X.; Kim, J.; Jeon, Y. Fucoidan isolated from the popular edible brown seaweed Sargassum fusiforme suppresses lipopolysaccharide-induced inflammation by blocking NF-κB signal pathway. J. Appl. Phyccol. 2021, 33, 1845–1852. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Xu, Y.; Yang, H.; Wang, J.; Xue, C.; Yan, X.; Su, L. Anti-inflammation effects of fucosylated chondroitin sulphate from Acaudina molpadioides by altering gut microbiota in obese mice. Food Funct. 2019, 10, 1736–1746. [Google Scholar] [CrossRef]
- Hu, S.; Jiang, W.; Li, S.; Song, W.; Ji, L.; Cai, L.; Liu, X. Fucosylated chondroitin sulphate from sea cucumber reduces hepatic endoplasmic reticulum stress. J. Funct. Food 2015, 16, 352–363. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Q.; Zhang, C.; Jia, W.; Han, L.; Yu, Q. Chicken leg bone as a source of chondroitin sulfate. Carbohyd. Polym. 2019, 207, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Li, P.; Wang, Y.; Su, N.; Xiao, M.; Li, X.; Shang, N. Structural analysis and anti-cancer activity of low-molecular-weight chondroitin sulfate from hybrid sturgeon cartilage. Carbohyd. Polym. 2022, 275, 118700. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Monosaccharide Composition Proportion % | Sulfate | Mp(kDa) | Mw(kDa) | Mn(kDa) | Polydispersity | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| GalN | GlcN | Gal | Gac | GlcA | ||||||
| CHS | 37.60 | 3.92 | 4.17 | 6.95 | 47.36 | 17.40% | 173.75 | 238.35 | 143.72 | 1.66 |
| RT | Methylated Sugar | Mass Fragments (m/z) | Area Ratios | Type of Linkage |
|---|---|---|---|---|
| 32.180 | 2,3,4,6-Me4-Glcp | 43,87,99,101,113,117,129,131,161,173,233 | 69.69 | →4)-β-d-GlcpA-(1→ |
| 45.879 | 2,3,6-Me3-Galp | 43,75,100,117,129,158,171 | 30.31 | →3)-β-d-GalpNAc-(1→ |
| Glycosyl Residues | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | C-Me | C=O | |
|---|---|---|---|---|---|---|---|---|---|
| Glycosidic bond A | →4)-β-D-GlcpA-(1→ | 4.41 | 3.29 | 3.51 | 3.65 | 3.61 | |||
| 105.47 | 73.69 | 75.11 | 82.39 | 77.69 | 175.69 | ||||
| Glycosidic bond B | →3)-β-D-GlcpNAc-(1→ | 4.47 | 3.93 | 3.77 | 4.14 | 3.89 | 3.7 | ||
| 102.56 | 52.29 | 81.20 | 68.85 | 73.90 | 62.36 | 23.90 | 176.33 |
| Normal | HFD | HFD + CHS | HFD + DMBG | |
|---|---|---|---|---|
| Body weight gain (g) | 10.93 ± 0.97 a | 20.62 ± 1.23 a | 9.85 ± 0.98 c | 11.81 ± 0.65 d |
| Liver/body weight (g/100g) | 3.36 ± 0.25 a | 7.32 ± 0.84 b | 4.29 ± 0.41 c | 4.73 ± 0.56 c |
| Kidney/body weight (g/100g) | 1.06 ± 0.09 ab | 1.82 ± 0.12 b | 1.42 ± 0.16 c | 1.28 ± 0.12 c |
| Abdominal fat/body weight (g/g) | 0.47 ± 0.123 c | 2.426 ± 0.43 a | 0.403 ± 0.106 b | 1.376 ± 0.798 d |
| Fasting blood glucose (mmol/L) | 5.21 ± 0.52 a | 13.44 ± 0.32 b | 5.32 ± 0.84 a | 6.07 ± 0.84 a |
| Serum insulin (mIU/L) | 25.99 ± 0.1 a | 32.64 ± 1.1 c | 28.19 ± 0.36 b | 28.47 ± 1.03 b |
| HOMA-IR | 6.36 ± 0.39 a | 12.22 ± 0.72 b | 7.61 ± 0.46 c | 8.77 ± 0.87 d |
| QUICKI | 0.007 ± 0.0004 a | 0.004 ± 0.0002 b | 0.006 ± 0.0004 c | 0.005 ± 0.0005 c |
| Serum TC (mmol/L) | 3.75 ± 0.76 a | 7.35 ± 0.30 c | 4.90 ± 0.16 b | 4.89 ± 0.26 ab |
| Serum TG (mmol/L) | 1.37 ± 0.009 a | 2.66 ± 0.054 c | 1.43 ± 0.152 ab | 1.62 ± 0.061 b |
| Hepatic TC (mmol/L) | 0.005 ± 0.001 a | 0.009 ± 0.001 c | 0.005 ± 0.001 a | 0.006 ± 0.001 b |
| Hepatic TG (mmol/L) | 0.018 ± 0.005 a | 0.026 ± 0.001 b | 0.017 ± 0.004 a | 0.020 ± 0.003 ab |
| Serum HDL-C (mmol/L) | 1.30 ± 0.09 a | 0.31 ± 0.023 b | 1.17 ± 0.08 c | 0.44 ± 0.09 d |
| Serum LDL-C (mmol/L) | 2.50 ± 0.20 a | 5.26 ± 0.37 b | 4.01 ± 0.4 c | 4.64 ± 0.28 d |
| Serum IL-6 (pg/mL) | 55.93 ± 1.05 a | 78.98 ± 1.51 c | 64.48 ± 1.16 b | 57.50 ± 0.83 a |
| Serum IL−1β (pg/mL) | 49.28 ± 1.53 a | 81.17 ± 1.79 b | 73.84 ± 1.53 c | 70.95 ± 2.01 d |
| Serum IL−10 (pg/mL) | 274.06 ± 1.27 a | 256.05 ± 2.36 b | 272.74 ± 2.37 a | 264.82 ± 1.61 c |
| Serum TNF-α (pg/mL) | 19.52 ± 3.29 a | 106.01 ± 3.83 b | 63.83 ± 5.19 c | 35.9 ± 2.39 d |
| Hepatic ALT (U/L) | 50.19 ± 3.65 a | 39.82 ± 1.66 b | 47.41 ± 0.79 a | 40.59 ± 2.84 b |
| Hepatic AST (U/L) | 159.90 ± 9.79 a | 114.49 ± 14.83 c | 136.19 ± 13.17 b | 146.80 ± 8.10 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, S.; Zhu, H.; Chen, S.; Wan, X.; Liu, Y.; Ren, Z.; Gao, S. Structural Characterization and Effects on Insulin Resistance of a Novel Chondroitin Sulfate from Halaelurus burgeri Skin. Mar. Drugs 2023, 21, 221. https://doi.org/10.3390/md21040221
Hu S, Zhu H, Chen S, Wan X, Liu Y, Ren Z, Gao S. Structural Characterization and Effects on Insulin Resistance of a Novel Chondroitin Sulfate from Halaelurus burgeri Skin. Marine Drugs. 2023; 21(4):221. https://doi.org/10.3390/md21040221
Chicago/Turabian StyleHu, Shiwei, Hongli Zhu, Sichun Chen, Xiaofeng Wan, Yishu Liu, Zhaocai Ren, and Shuang Gao. 2023. "Structural Characterization and Effects on Insulin Resistance of a Novel Chondroitin Sulfate from Halaelurus burgeri Skin" Marine Drugs 21, no. 4: 221. https://doi.org/10.3390/md21040221
APA StyleHu, S., Zhu, H., Chen, S., Wan, X., Liu, Y., Ren, Z., & Gao, S. (2023). Structural Characterization and Effects on Insulin Resistance of a Novel Chondroitin Sulfate from Halaelurus burgeri Skin. Marine Drugs, 21(4), 221. https://doi.org/10.3390/md21040221