
Recombinant Analogs of Sea Anemone Kunitz-Type Peptides Influence P2X7 Receptor Activity in Neuro-2a Cells

 , , , , , , , , ,
, , , , , , , , ,  , and
, and 
Abstract
:1. Introduction
2. Results
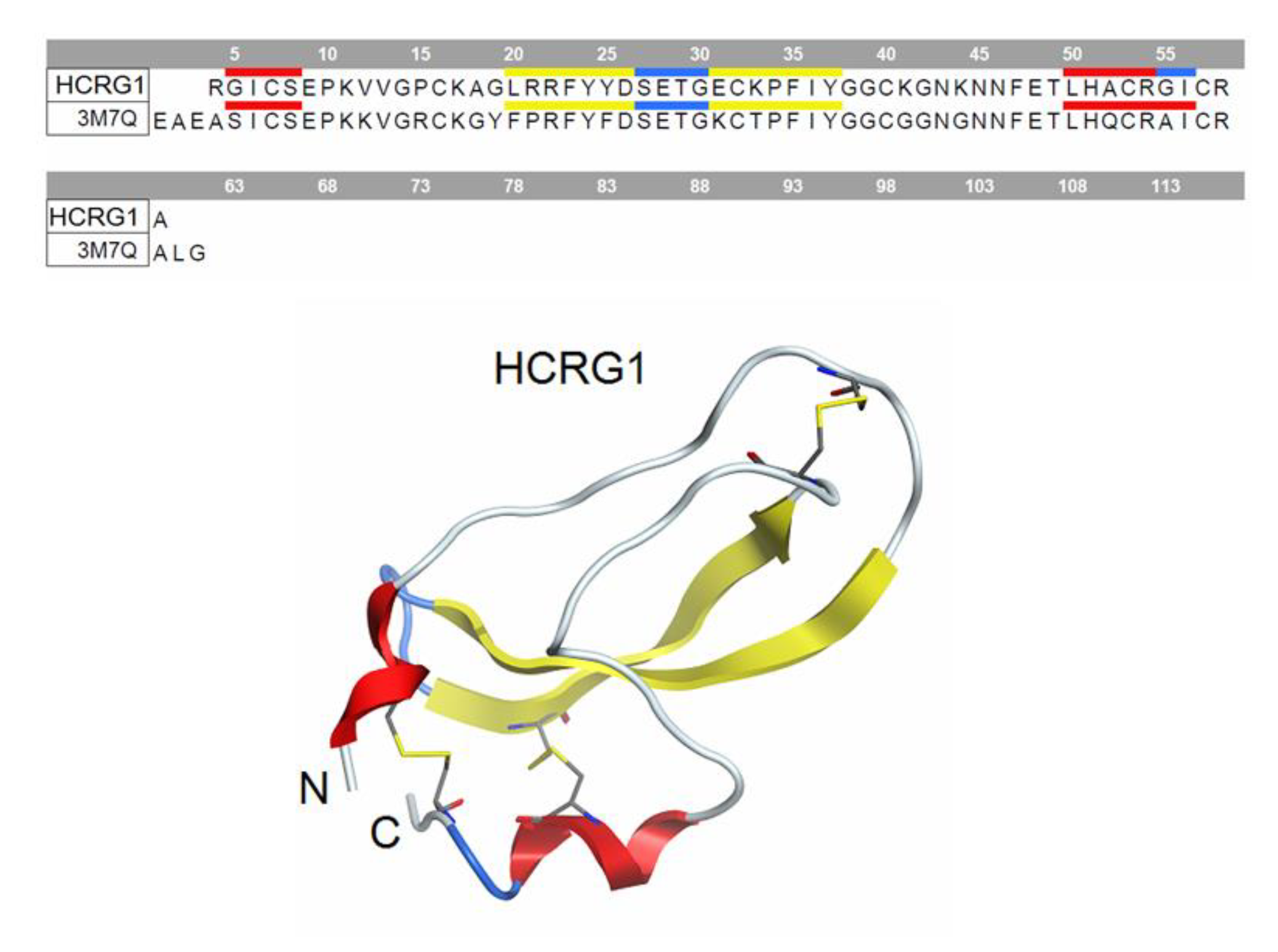
2.1. Expression and Purification of Recombinant Peptides
2.2. Peptides Interaction with Neuronal Cells Neuro-2a
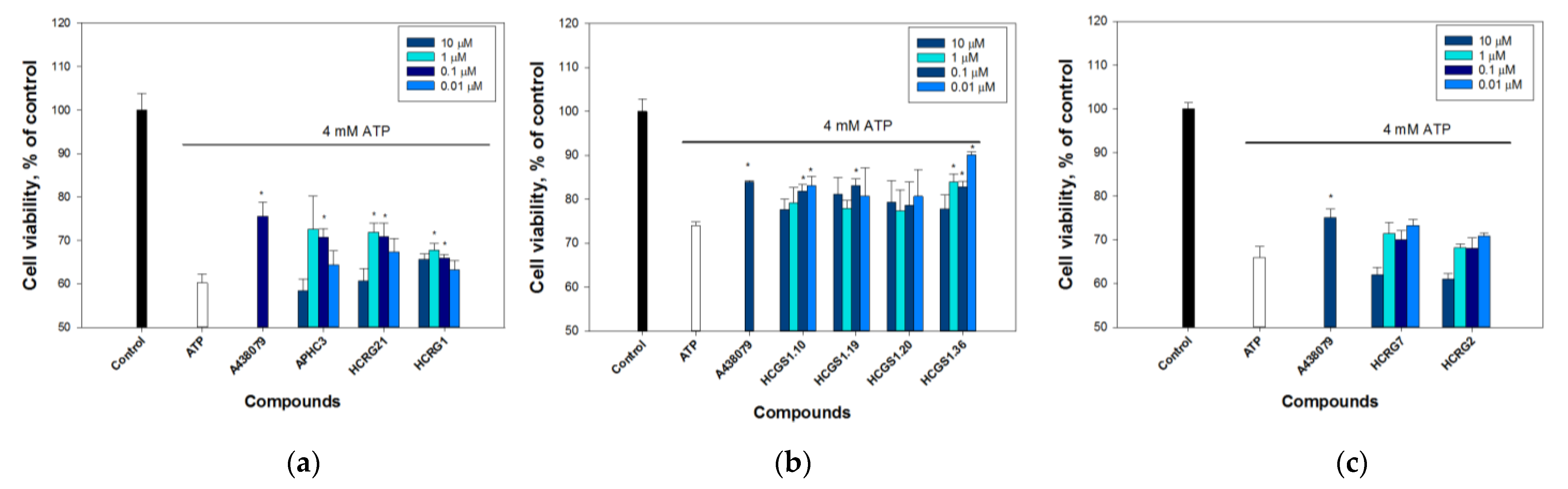
2.2.1. Peptides Protect Cells from ATP-Induced Death
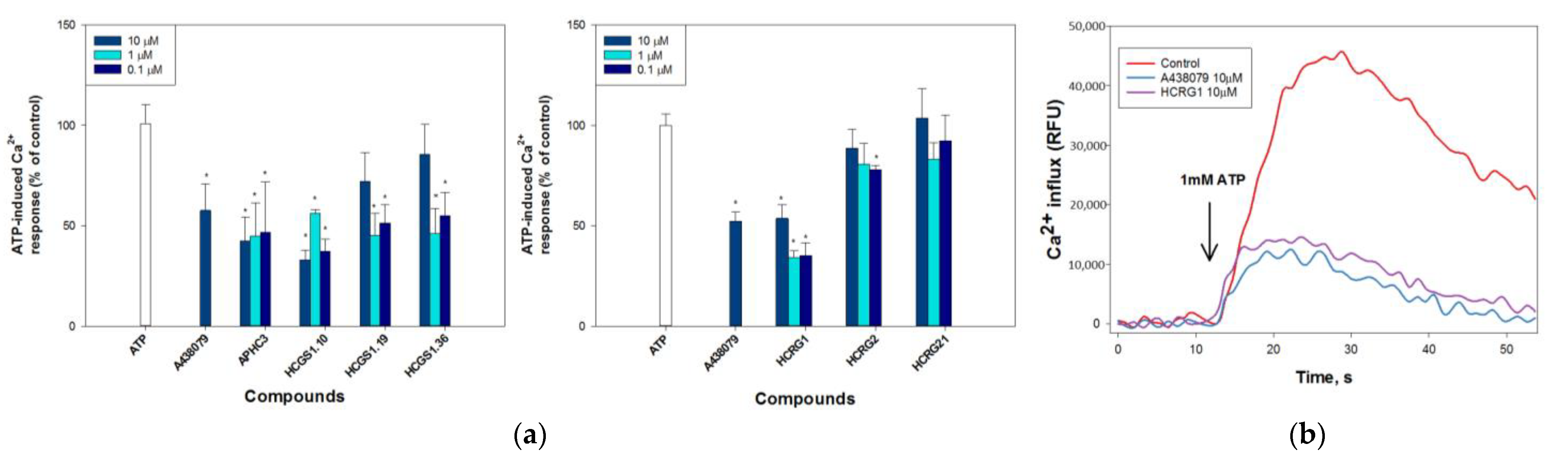
2.2.2. Peptides Regulate ATP-Induced Calcium Influx
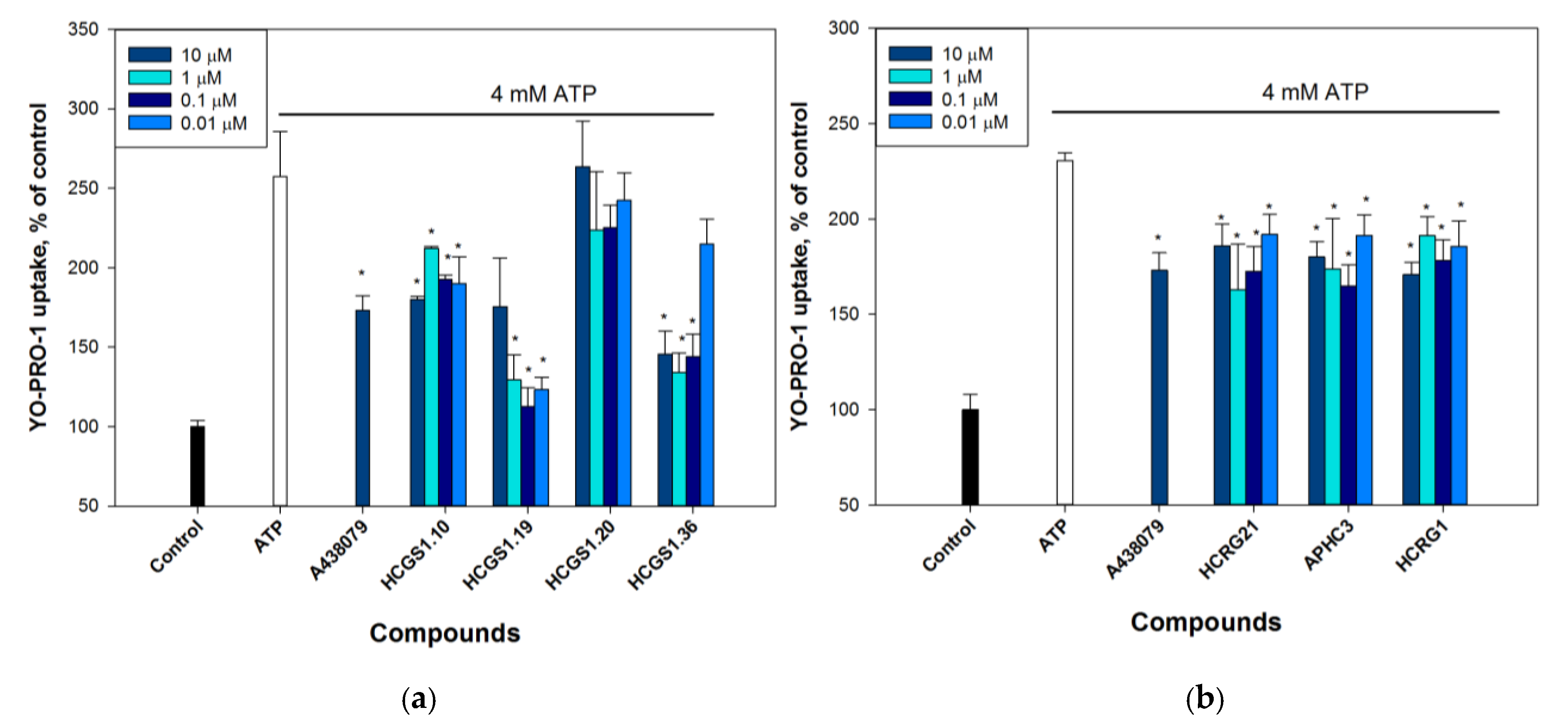
2.2.3. Peptides Reduce P2X7 Mediated Dye Uptake
2.3. Interaction of Peptides with P2X7 Subunit
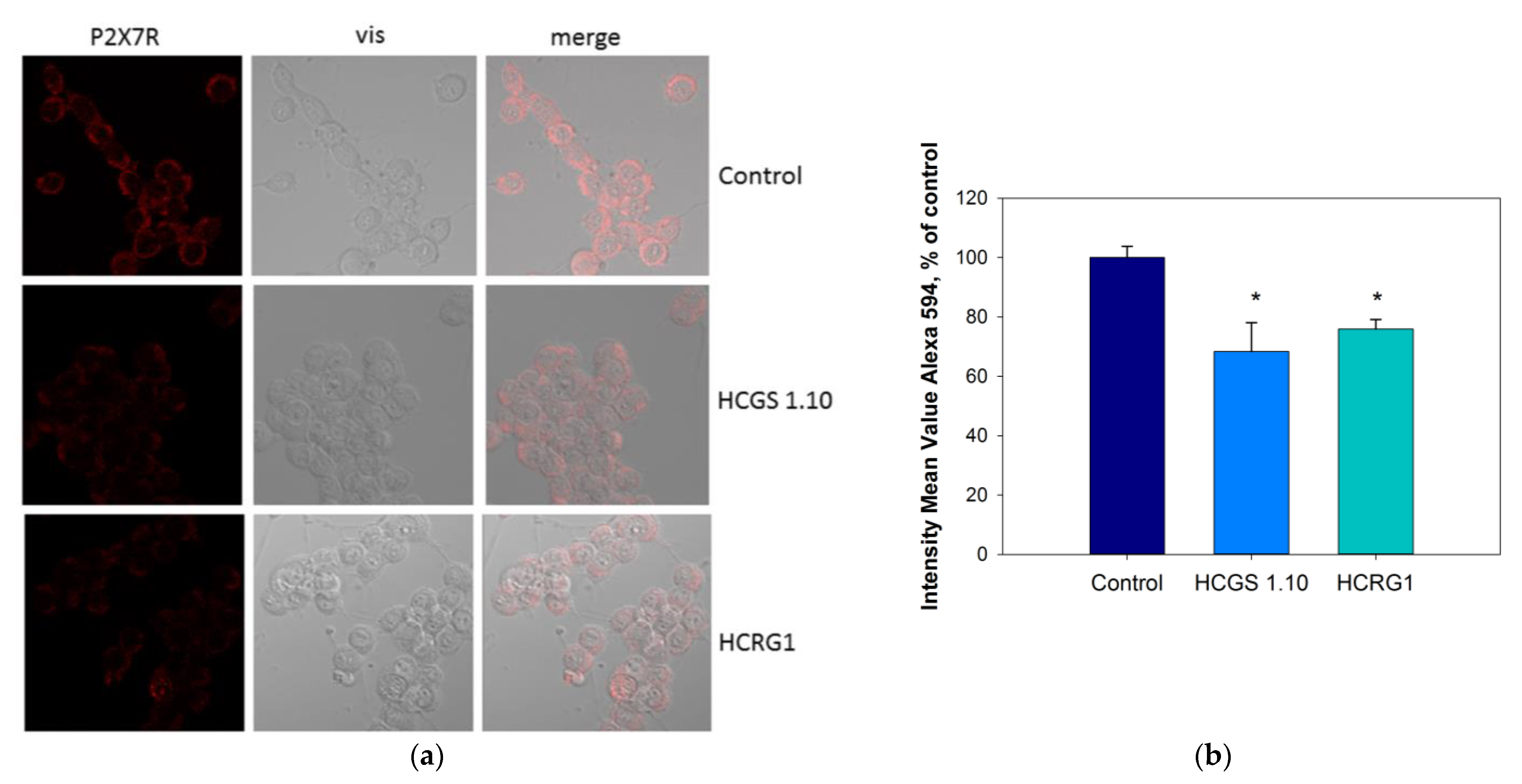
2.4. Influence of Peptides on P2X7 Expression in Neuro-2a Cells
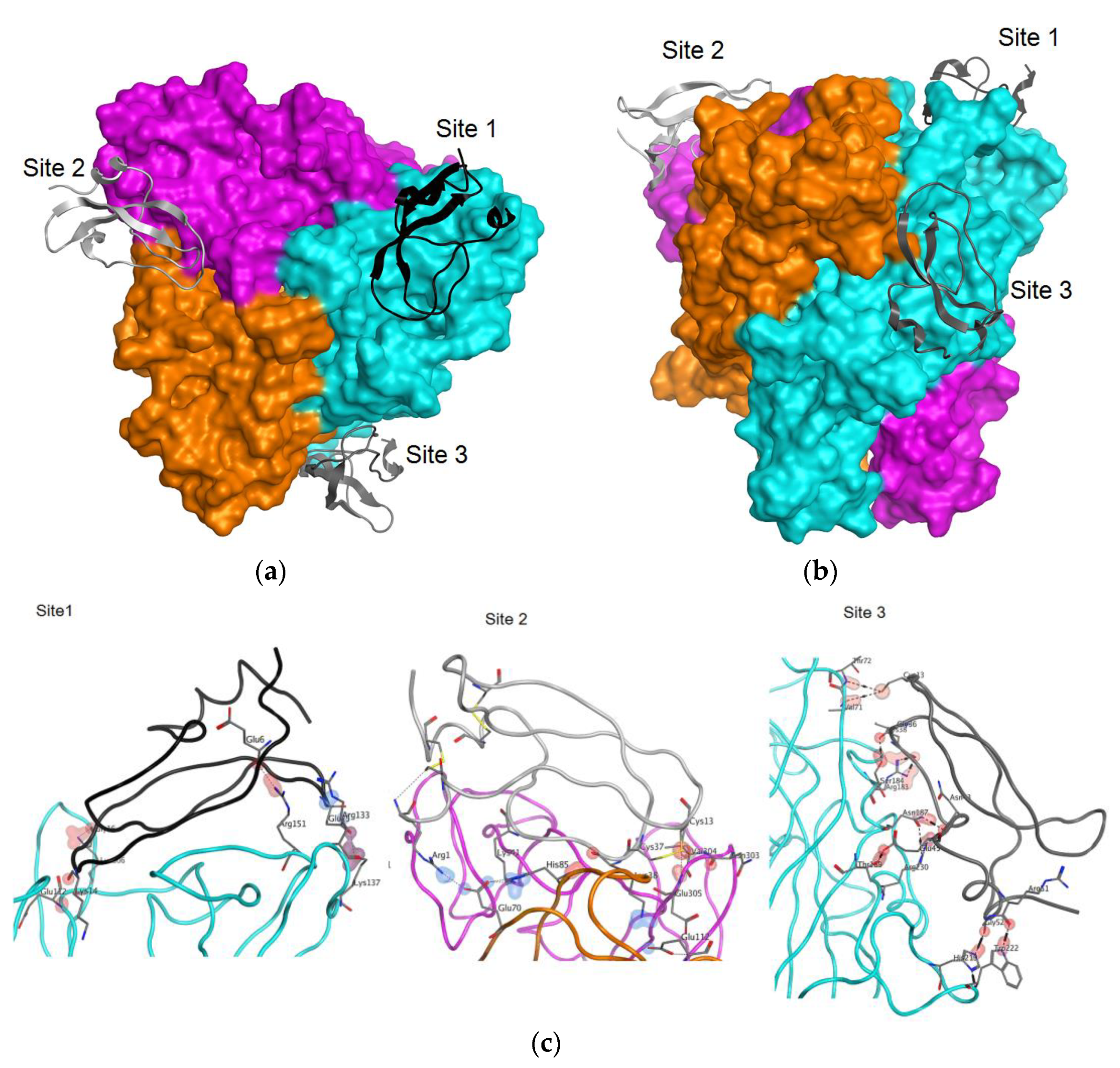
2.5. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Expression and Isolation of Kunitz-Type Recombinant Peptides
4.2. Mass Spectrometry Analysis
4.3. Amino Acid Sequence Determination
4.4. One-Dimensional NMR Spectroscopy
4.5. Cell Culture
4.6. Analysis of Cytotoxic Activity
4.7. Cytoprotection Activity Assay
4.8. Ca2+Influx Measurement
4.9. YO-PRO-1 Uptake Measurement
4.10. Surface Plasmon Resonance
- (1)
- 1:1 (Langmuir) binding:
- (2)
- Reaction of two states (with conformational change):
4.11. Molecular Modeling
4.12. Immunocytochemistry
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linden, J.; Koch-Nolte, F.; Dahl, G. Purine Release, Metabolism, and Signaling in the Inflammatory Response. Annu. Rev. Immunol. 2019, 37, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Schmalzing, G.; Markwardt, F. The Elusive P2X7 Macropore. Trends Cell Biol. 2018, 28, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Stefani, C.; Badiale, G.; Campione, G.; Martini, F.; Tognon, M. The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications. Cancers 2022, 14, 1116. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Knight, G.E. The potential of P2X7 receptors as a therapeutic target, including inflammation and tumour progression. Purinergic Signal. 2018, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Savio, L.E.B.; de Andrade Mello, P.; da Silva, C.G.; Coutinho-Silva, R. The P2X7 Receptor in Inflammatory Diseases: Angel or Demon? Front. Pharmacol. 2018, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Kanellopoulos, J.M.; Delarasse, C. Pleiotropic Roles of P2X7 in the Central Nervous System. Front. Cell. Neurosci. 2019, 13, 401. [Google Scholar] [CrossRef]
- Sperlagh, B.; Illes, P. P2X7 receptor: An emerging target in central nervous system diseases. Trends Pharmacol. Sci. 2014, 35, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Arbeloa, J.; Perez-Samartin, A.; Gottlieb, M.; Matute, C. P2X7 receptor blockade prevents ATP excitotoxicity in neurons and reduces brain damage after ischemia. Neurobiol. Dis. 2012, 45, 954–961. [Google Scholar] [CrossRef]
- Parvathenani, L.K.; Tertyshnikova, S.; Greco, C.R.; Roberts, S.B.; Robertson, B.; Posmantur, R. P2X7 mediates superoxide production in primary microglia and is up-regulated in a transgenic mouse model of Alzheimer’s disease. J. Biol. Chem. 2003, 278, 13309–13317. [Google Scholar] [CrossRef] [Green Version]
- Coelho, G.R.; da Silva, D.L.; Beraldo-Neto, E.; Vigerelli, H.; de Oliveira, L.A.; Sciani, J.M.; Pimenta, D.C. Neglected Venomous Animals and Toxins: Underrated Biotechnological Tools in Drug Development. Toxins 2021, 13, 851. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaeva, M.P.; Chausova, V.E.; Zelepuga, E.A.; Guzev, K.V.; Tabakmakher, V.M.; Monastyrnaya, M.M.; Kozlovskaya, E.P. A new multigene superfamily of Kunitz-type protease inhibitors from sea anemone Heteractis crispa. Peptides 2012, 34, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.; Gladkikh, I.; Chausova, V.; Monastyrnaya, M.; Anastyuk, S.; Chernikov, O.; Yurchenko, E.; Aminin, D.; Isaeva, M.; Leychenko, E.; et al. Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a new beta-defensin alpha-amylase inhibitor. J. Proteom. 2018, 173, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Andreev, Y.A.; Kozlov, S.A.; Korolkova, Y.V.; Dyachenko, I.A.; Bondarenko, D.A.; Skobtsov, D.I.; Murashev, A.N.; Kotova, P.D.; Rogachevskaja, O.A.; Kabanova, N.V.; et al. Polypeptide Modulators of TRPV1 Produce Analgesia without Hyperthermia. Mar. Drugs 2013, 11, 5100–5115. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Fernandez, R.; Peigneur, S.; Pons, T.; Alvarez, C.; Gonzalez, L.; Chavez, M.A.; Tytgat, J. The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels. Toxins 2016, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladkikh, I.; Peigneur, S.; Sintsova, O.; Lopes Pinheiro-Junior, E.; Klimovich, A.; Menshov, A.; Kalinovsky, A.; Isaeva, M.; Monastyrnaya, M.; Kozlovskaya, E.; et al. Kunitz-Type Peptides from the Sea Anemone Heteractis crispa Demonstrate Potassium Channel Blocking and Anti-Inflammatory Activities. Biomedicines 2020, 8, 473. [Google Scholar] [CrossRef]
- Nikolaev, M.V.; Dorofeeva, N.A.; Komarova, M.S.; Korolkova, Y.V.; Andreev, Y.A.; Mosharova, I.V.; Grishin, E.V.; Tikhonov, D.B.; Kozlov, S.A. TRPV1 activation power can switch an action mode for its polypeptide ligands. PLoS ONE 2017, 12, e0177077. [Google Scholar] [CrossRef] [Green Version]
- Monastyrnaya, M.; Peigneur, S.; Zelepuga, E.; Sintsova, O.; Gladkikh, I.; Leychenko, E.; Isaeva, M.; Tytgat, J.; Kozlovskaya, E. Kunitz-Type Peptide HCRG21 from the Sea Anemone Heteractis crispa Is a Full Antagonist of the TRPV1 Receptor. Mar. Drugs 2016, 14, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvetkina, A.; Leychenko, E.; Chausova, V.; Zelepuga, E.; Chernysheva, N.; Guzev, K.; Pislyagin, E.; Yurchenko, E.; Menchinskaya, E.; Aminin, D.; et al. A new multigene HCIQ subfamily from the sea anemone Heteractis crispa encodes Kunitz-peptides exhibiting neuroprotective activity against 6-hydroxydopamine. Sci. Rep. 2020, 10, 4205. [Google Scholar] [CrossRef] [Green Version]
- Kvetkina, A.; Pislyagin, E.; Menchinskaya, E.; Yurchenko, E.; Kalina, R.; Kozlovskiy, S.; Kaluzhskiy, L.; Menshov, A.; Kim, N.; Peigneur, S.; et al. Kunitz-Type Peptides from Sea Anemones Protect Neuronal Cells against Parkinson’s Disease Inductors via Inhibition of ROS Production and ATP-Induced P2X7 Receptor Activation. Int. J. Mol. Sci. 2022, 23, 5115. [Google Scholar] [CrossRef]
- Sintsova, O.; Gladkikh, I.; Monastyrnaya, M.; Tabakmakher, V.; Yurchenko, E.; Menchinskaya, E.; Pislyagin, E.; Andreev, Y.; Kozlov, S.; Peigneur, S.; et al. Sea Anemone Kunitz-Type Peptides Demonstrate Neuroprotective Activity in the 6-Hydroxydopamine Induced Neurotoxicity Model. Biomedicines 2021, 9, 283. [Google Scholar] [CrossRef] [PubMed]
- Esipov, R.S.; Makarov, D.A.; Stepanenko, V.N.; Kostromina, M.A.; Muravyova, T.I.; Andreev, Y.A.; Dyachenko, I.A.; Kozlov, S.A.; Grishin, E.V. Pilot production of the recombinant peptide toxin of Heteractis crispa as a potential analgesic by intein-mediated technology. Protein Expr. Purif. 2018, 145, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.V.; Monastyrnaya, M.M.; Pislyagin, E.A.; Menchinskaya, E.S.; Leychenko, E.V.; Aminin, D.L.; Kozlovskaya, E.P. Anti-inflammatory activity of a polypeptide from the Heteractis crispa sea anemone. Russ. J. Bioorganic Chem. 2015, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.V.; Pislyagin, E.A.; Gladkikh, I.N.; Monastyrnaya, M.M.; Menchinskaya, E.S.; Leychenko, E.V.; Aminin, D.L.; Kozlovskaya, E.P. Kunitz-type peptides of the sea anemone Heteractis crispa: Potential anti-inflammatory compounds. Russ. J. Bioorganic Chem. 2017, 43, 91–97. [Google Scholar] [CrossRef]
- Gladkikh, I.; Monastyrnaya, M.; Zelepuga, E.; Sintsova, O.; Tabakmakher, V.; Gnedenko, O.; Ivanov, A.; Hua, K.F.; Kozlovskaya, E. New Kunitz-Type HCRG Polypeptides from the Sea Anemone Heteractis crispa. Mar. Drugs 2015, 13, 6038–6063. [Google Scholar] [CrossRef]
- García-Fernández, R.; Pons, T.; Perbandt, M.; Valiente, P.A.; Talavera, A.; González-González, Y.; Rehders, D.; Chávez, M.A.; Betzel, C.; Redecke, L. Structural insights into serine protease inhibition by a marine invertebrate BPTI Kunitz-type inhibitor. J. Struct. Biol. 2012, 180, 271–279. [Google Scholar] [CrossRef]
- Andrejew, R.; Oliveira-Giacomelli, Á.; Ribeiro, D.E.; Glaser, T.; Arnaud-Sampaio, V.F.; Lameu, C.; Ulrich, H. The P2X7 Receptor: Central Hub of Brain Diseases. Front. Mol. Neurosci. 2020, 13, 124. [Google Scholar] [CrossRef]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Villafuertes, R.; del Puerto, A.; Díaz-Hernández, M.; Bustillo, D.; Díaz-Hernández, J.I.; Huerta, P.G.; Artalejo, A.R.; Garrido, J.J.; Miras-Portugal, M.T. Ca2+/calmodulin-dependent kinase II signalling cascade mediates P2X7 receptor-dependent inhibition of neuritogenesis in neuroblastoma cells. FEBS J. 2009, 276, 5307–5325. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Martín, Y.; Bustillo, D.; Gómez-Villafuertes, R.; Sánchez-Nogueiro, J.; Torregrosa-Hetland, C.; Binz, T.; Gutiérrez, L.M.; Miras-Portugal, M.T.; Artalejo, A.R. P2X7 receptors trigger ATP exocytosis and modify secretory vesicle dynamics in neuroblastoma cells. J. Biol. Chem. 2011, 286, 11370–11381. [Google Scholar] [CrossRef] [Green Version]
- Giannuzzo, A.; Pedersen, S.F.; Novak, I. The P2X7 receptor regulates cell survival, migration and invasion of pancreatic ductal adenocarcinoma cells. Mol. Cancer 2015, 14, 203. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Wang, M.; Liao, Z.; Camden, J.M.; Yu, S.; Simonyi, A.; Sun, G.Y.; Gonzalez, F.A.; Erb, L.; Seye, C.I.; et al. P2X(7) nucleotide receptors mediate caspase-8/9/3-dependent apoptosis in rat primary cortical neurons. Purinergic Signal. 2005, 1, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, B.C.; Kim, J.S.; Namgung, U.; Ha, H.; Kim, K.T. P2X7 nucleotide receptor mediation of membrane pore formation and superoxide generation in human promyelocytes and neutrophils. J. Immunol. 2001, 166, 6754–6763. [Google Scholar] [CrossRef] [Green Version]
- Thyagarajan, B.; Potian, J.G.; Baskaran, P.; McArdle, J.J. Capsaicin modulates acetylcholine release at the myoneural junction. Eur. J. Pharmacol. 2014, 744, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J. Hormesis: A revolution in toxicology, risk assessment and medicine. EMBO Rep. 2004, 5 (Suppl. S1), S37–S40. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J.; Blain, R.B. The hormesis database: The occurrence of hormetic dose responses in the toxicological literature. Regul. Toxicol. Pharmacol. RTP 2011, 61, 73–81. [Google Scholar] [CrossRef]
- Cookman, C.J.; Belcher, S.M. Classical nuclear hormone receptor activity as a mediator of complex concentration response relationships for endocrine active compounds. Curr. Opin. Pharmacol. 2014, 19, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Logashina, Y.A.; Palikova, Y.A.; Palikov, V.A.; Kazakov, V.A.; Smolskaya, S.V.; Dyachenko, I.A.; Tarasova, N.V.; Andreev, Y.A. Anti-Inflammatory and Analgesic Effects of TRPV1 Polypeptide Modulator APHC3 in Models of Osteo- and Rheumatoid Arthritis. Mar. Drugs 2021, 19, 39. [Google Scholar] [CrossRef]
- Philyppov, I.B.; Paduraru, O.N.; Andreev, Y.A.; Grishin, E.V.; Shuba, Y.M. Modulation of TRPV1-dependent contractility of normal and diabetic bladder smooth muscle by analgesic toxins from sea anemone Heteractis crispa. Life Sci. 2012, 91, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.; Palikov, V.; Palikova, Y.; Klimovich, A.; Gladkikh, I.; Andreev, Y.; Monastyrnaya, M.; Kozlovskaya, E.; Dyachenko, I.; Kozlov, S.; et al. Peptide Blocker of Ion Channel TRPV1 Exhibits a Long Analgesic Effect in the Heat Stimulation Model. Dokl. Biochem. Biophys. 2020, 493, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Kopp, R.; Krautloher, A.; Ramírez Fernández, A.; Nicke, A. P2X7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladkikh, I.; Monastyrnaya, M.; Leychenko, E.; Zelepuga, E.; Chausova, V.; Isaeva, M.; Anastyuk, S.; Andreev, Y.; Peigneur, S.; Tytgat, J.; et al. Atypical reactive center Kunitz-type inhibitor from the sea anemone Heteractis crispa. Mar. Drugs 2012, 10, 1545–1565. [Google Scholar] [CrossRef] [Green Version]
- Hwang, T.L.; Shaka, A.J. Water Suppression That Works. Excitation Sculpting Using Arbitrary Wave-Forms and Pulsed-Field Gradients. J. Magn. Reson. Ser. A 1995, 112, 275–279. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Pislyagin, E.; Kozlovskiy, S.; Menchinskaya, E.; Chingizova, E.; Likhatskaya, G.; Gorpenchenko, T.; Sabutski, Y.; Polonik, S.; Aminin, D. Synthetic 1,4-Naphthoquinones inhibit P2X7 receptors in murine neuroblastoma cells. Bioorganic Med. Chem. 2021, 31, 115975. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | kon1, mole−1c−1 | kon2, mole−1c−1 | koff1, c−1 | koff2, c−1 | Kd, M | Evaluation Model |
|---|---|---|---|---|---|---|
| HCRG1 | (9.20 ± 1.10) × 102 | (8.5 ± 1.0) × 10−3 | (4.37 ± 0.57) × 10−2 | (1.5 ± 0.20) × 10−3 | 7.13 × 10−6 | Binding with two states |
| HCGS1.10 | (1.71 ± 0.19) × 102 | (4.5 ± 0.52) × 10−3 | 2.63 × 10−5 | Binding 1:1 (Langmuir 1:1) | ||
| Site | Type * | ChainA | PosA | SetA | ChainB | PosB | SetB | Energy | Dist |
|---|---|---|---|---|---|---|---|---|---|
| h | mm-trim-.A | 65 | Glu112 | HCRG1 | 14 | Lys14 | −0.700 | 3.354 | |
| i | mm-trim-.A | 86 | Arg133 | HCRG1 | 25 | Glu25 | −1.505 | 3.609 | |
| Site 1 | ih | mm-trim-.A | 90 | Lys137 | HCRG1 | 25 | Glu25 | −8.017 | 2.393 |
| h | mm-trim-.A | 104 | Arg151 | HCRG1 | 6 | Glu6 | −3.600 | 2.821 | |
| h | mm-trim-.A | 259 | Lys306 | HCRG1 | 16 | Gly16 | −7.800 | 2.976 | |
| i | mm-trim-.B | 23 | Glu70 | HCRG1 | 1 | Arg1 | −4.886 | 2.940 | |
| i | mm-trim-.B | 23 | Glu70 | HCRG1 | 41 | Lys41 | −5.965 | 2.800 | |
| h | mm-trim-.B | 38 | His85 | HCRG1 | 38 | Lys38 | −1.700 | 3.268 | |
| Site 2 | i | mm-trim-.B | 65 | Glu112 | HCRG1 | 38 | Lys38 | −2.364 | 3.385 |
| h | mm-trim-.C | 256 | Asn303 | HCRG1 | 13 | Cys13 | 0.300 | 3.246 | |
| h | mm-trim-.C | 257 | Val304 | HCRG1 | 13 | Cys13 | −0.800 | 2.918 | |
| h | mm-trim-.C | 258 | Glu305 | HCRG1 | 13 | Cys13 | −2.600 | 3.125 | |
| h | mm-trim-.A | 24 | Val71 | HCRG1 | 13 | Cys13 | −0.800 | 4.188 | |
| h | mm-trim-.A | 136 | Arg183 | HCRG1 | 38 | Lys38 | −3.600 | 2.968 | |
| h | mm-trim-.A | 137 | Ser184 | HCRG1 | 36 | Gly36 | −1.400 | 2.696 | |
| Site 3 | h | mm-trim-.A | 140 | Asn187 | HCRG1 | 43 | Asn43 | −1.900 | 2.833 |
| h | mm-trim-.A | 142 | Thr189 | HCRG1 | 45 | Glu45 | −1.700 | 2.615 | |
| h | mm-trim-.A | 172 | His219 | HCRG1 | 52 | Gly52 | −3.800 | 2.783 | |
| h | mm-trim-.A | 175 | Trp222 | HCRG1 | 51 | Arg51 | −3.500 | 2.880 | |
| h | mm-trim-.A | 183 | Arg230 | HCRG1 | 43 | Asn43 | −6.900 | 2.809 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pislyagin, E.A.; Menchinskaya, E.S.; Gladkikh, I.N.; Kvetkina, A.N.; Sintsova, O.V.; Popkova, D.V.; Kozlovskiy, S.A.; Gorpenchenko, T.Y.; Likhatskaya, G.N.; Kaluzhskiy, L.A.; et al. Recombinant Analogs of Sea Anemone Kunitz-Type Peptides Influence P2X7 Receptor Activity in Neuro-2a Cells. Mar. Drugs 2023, 21, 192. https://doi.org/10.3390/md21030192
Pislyagin EA, Menchinskaya ES, Gladkikh IN, Kvetkina AN, Sintsova OV, Popkova DV, Kozlovskiy SA, Gorpenchenko TY, Likhatskaya GN, Kaluzhskiy LA, et al. Recombinant Analogs of Sea Anemone Kunitz-Type Peptides Influence P2X7 Receptor Activity in Neuro-2a Cells. Marine Drugs. 2023; 21(3):192. https://doi.org/10.3390/md21030192
Chicago/Turabian StylePislyagin, Evgeny A., Ekaterina S. Menchinskaya, Irina N. Gladkikh, Aleksandra N. Kvetkina, Oksana V. Sintsova, Darya V. Popkova, Sergei A. Kozlovskiy, Tatiana Y. Gorpenchenko, Galina N. Likhatskaya, Leonid A. Kaluzhskiy, and et al. 2023. "Recombinant Analogs of Sea Anemone Kunitz-Type Peptides Influence P2X7 Receptor Activity in Neuro-2a Cells" Marine Drugs 21, no. 3: 192. https://doi.org/10.3390/md21030192
APA StylePislyagin, E. A., Menchinskaya, E. S., Gladkikh, I. N., Kvetkina, A. N., Sintsova, O. V., Popkova, D. V., Kozlovskiy, S. A., Gorpenchenko, T. Y., Likhatskaya, G. N., Kaluzhskiy, L. A., Ivanov, A. S., Andreev, Y. A., Kozlov, S. A., Dmitrenok, P. S., Aminin, D. L., & Leychenko, E. V. (2023). Recombinant Analogs of Sea Anemone Kunitz-Type Peptides Influence P2X7 Receptor Activity in Neuro-2a Cells. Marine Drugs, 21(3), 192. https://doi.org/10.3390/md21030192