
Levels of Tetrodotoxins in Spawning Pufferfish, Takifugu alboplumbeus

 , ,
, ,
Abstract
:1. Introduction
2. Results
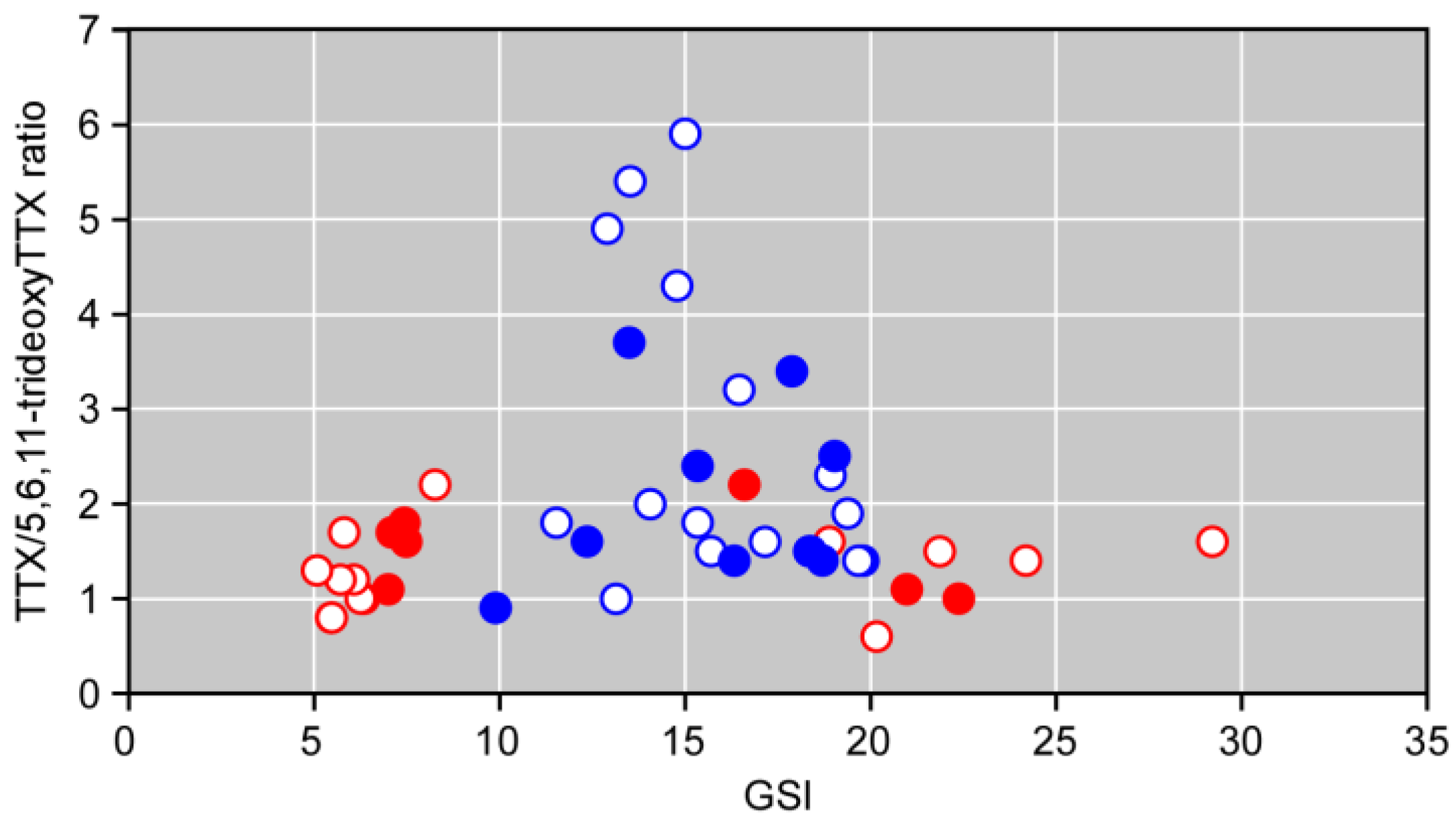
2.1. Sex Ratio and GSI of the Spawning Pufferfish
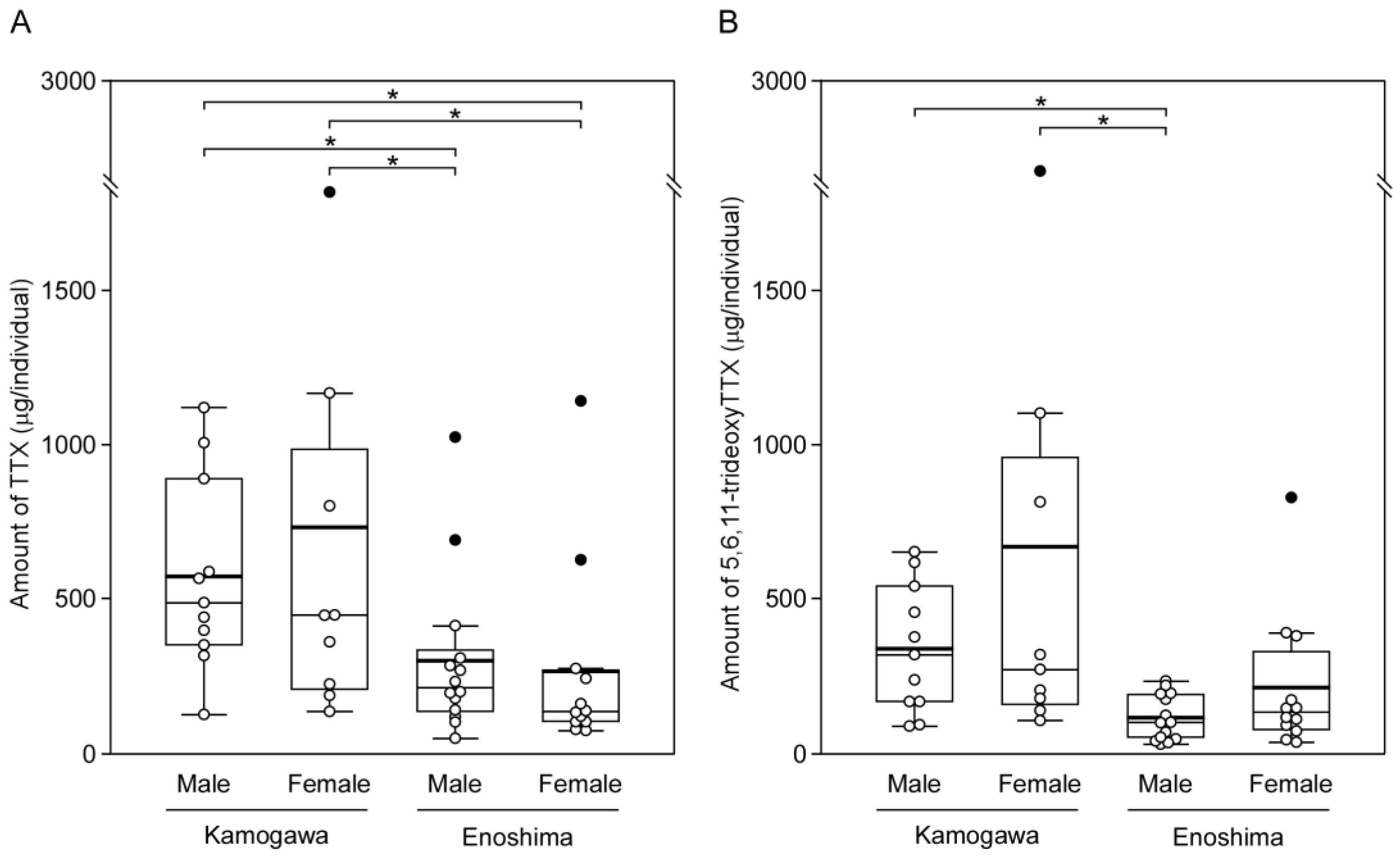
2.2. Amount of TTX and 5,6,11-TrideoxyTTX in the Pufferfish
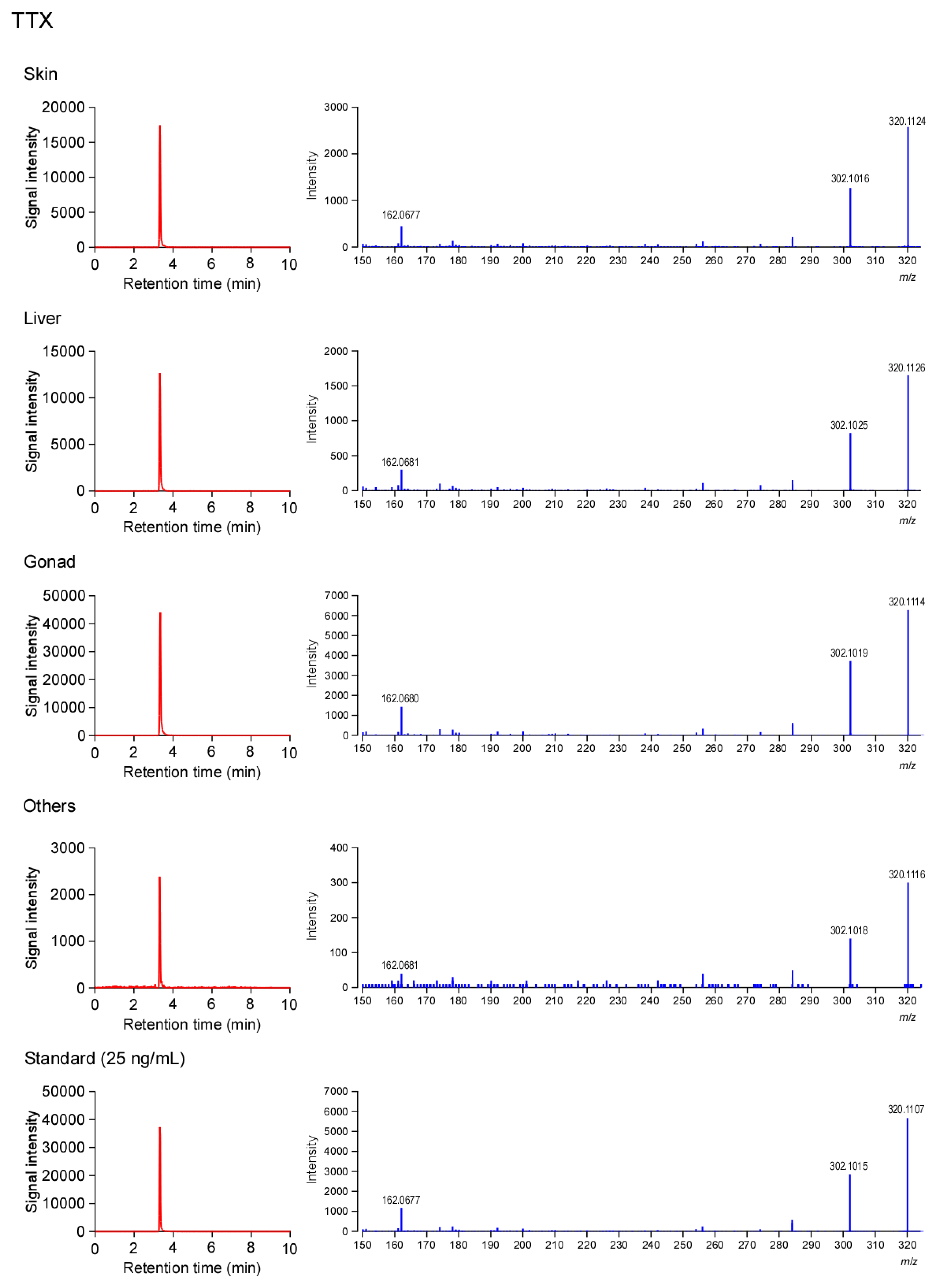
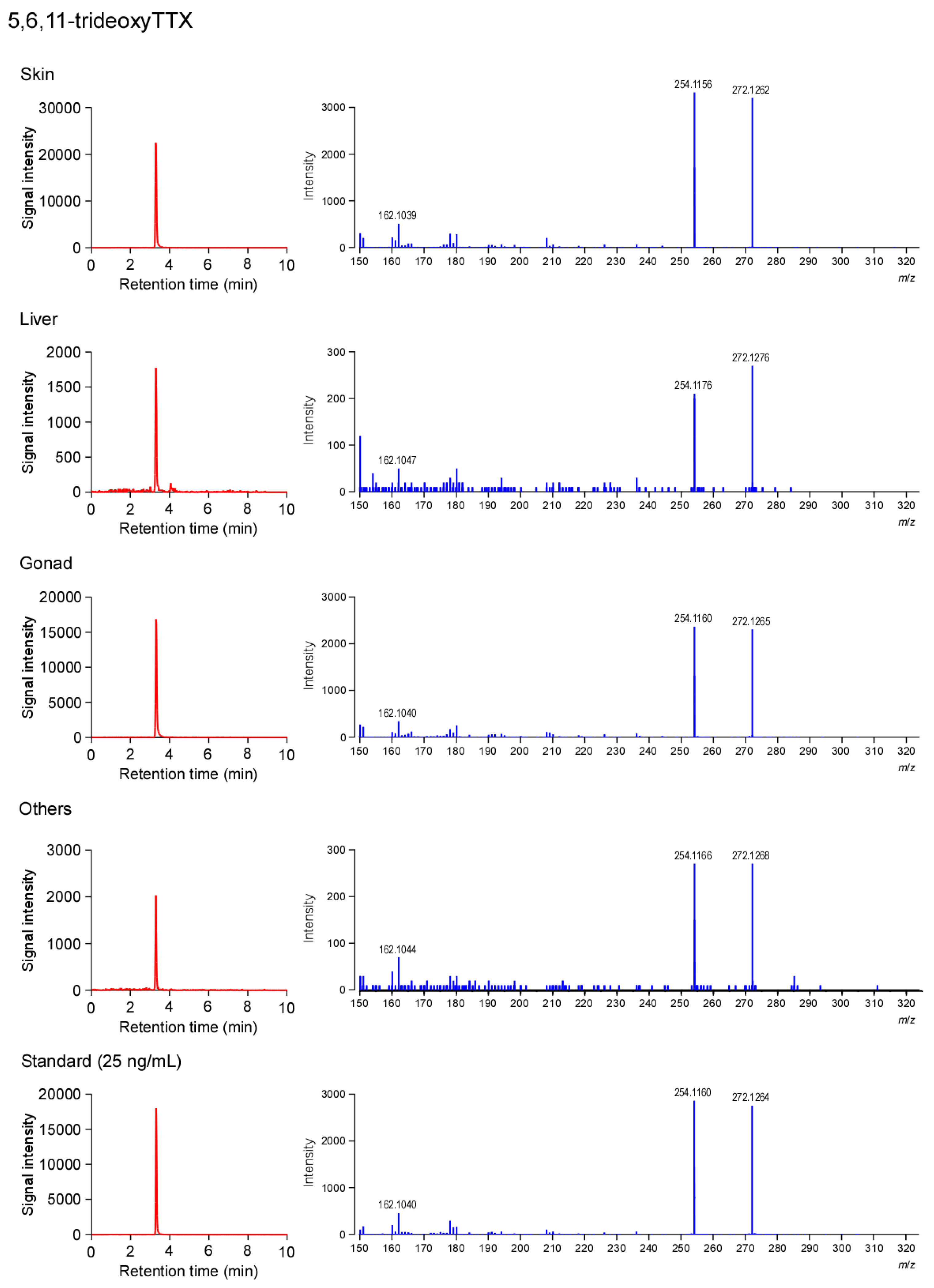
2.3. Tissue Distribution of TTX and 5,6,11-TrideoxyTTX
3. Discussion
4. Materials and Methods
4.1. Pufferfish Specimens
4.2. LC-MS/MS Analysis
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, D. Tetrodotoxin. VII. On the structures of tetrodotoxin and its derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, R.B. The structure of tetrodotoxin. Pure Appl. Chem. 1964, 9, 49–74. [Google Scholar] [CrossRef]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef]
- Noguchi, T.; Kao, H.; Hashimoto, Y. Toxicity of the goby, Gobius criniger. Bull. Jap. Soc. Sci. Fish. 1971, 37, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Hwang, D.F.; Tsai, Y.H.; Chai, T.J.; Jeng, S.S. Occurrence of tetrodotoxin and paralytic shellfish poison in Taiwan crab Zosimus aeneus. Fish. Sci. 1996, 62, 500–501. [Google Scholar] [CrossRef]
- Sheumack, D.D.; Howden, M.E.H.; Spence, I. Maculotoxin: A neurotoxin from the glands of the octopus, Hapalochlaena maculosa identified as tetrodotoxin. Science 1978, 199, 188–189. [Google Scholar] [CrossRef]
- Miyazawa, K.; Jeon, J.K.; Maruyama, J.; Noguchi, T.; Ito, K.; Hashimoto, K. Occurrence of tetrodotoxin in the flatworm Planocera multitentaculata. Toxicon 1986, 24, 645–650. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. USA 2006, 103, 3176–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvitti, L.; Wood, S.A.; Taylor, D.I.; McNabb, P.; Cary, S.C. First identification of tetrodotoxin (TTX) in the flatworm Stylochoplana sp.; a source of TTX for the sea slug Pleurobranchaea maculata. Toxicon 2015, 95, 23–29. [Google Scholar] [CrossRef]
- Ueda, H.; Itoi, S.; Sugita, H. TTX-bearing planocerid flatworm (Platyhelminthes: Acotylea) in the Ryukyu Islands, Japan. Mar. Drugs 2018, 16, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suo, R.; Kashitani, M.; Oyama, H.; Adachi, M.; Nakahigashi, R.; Sakakibara, R.; Nishikawa, T.; Sugita, H.; Itoi, S. First detection of tetrodotoxins in the cotylean flatworm Prosthiostomum trilineatum. Mar. Drugs 2021, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Mosher, H.S.; Fuhrman, G.J.; Fuhrman, F.A.; Fischer, H.G. Tarichatoxin-tetrodotoxin, a potent neurotoxin. Science 1964, 144, 1100–1110. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Mebs, D.; Yasumoto, T. Tetrodotoxin and its analogues in extracts from the toad Atelopus oxyrhynchus (family: Bufonidae). Toxicon 1992, 30, 1489–1492. [Google Scholar] [CrossRef]
- Stokes, A.N.; Ducey, P.K.; Neuman-Lee, L.; Hanifin, C.T.; French, S.S.; Pfrender, M.E.; Brodie, E.D., 3rd; Brodie, E.D., Jr. Confirmation and distribution of tetrodotoxin for the first time in terrestrial invertebrates: Two terrestrial flatworm species (Bipalium adventitium and Bipalium kewense). PLoS ONE 2014, 9, e100718. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Yamagishi, Y.; Yasumoto, T. 5,6,11-Trideoxytetrodotoxin from the puffer fish, Fugu poecilonotus. Tetrahedron Lett. 1995, 36, 9329–9332. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Schimmele, B.; Yasumoto, T. Isolation and structural assignment of 5-deoxytetrodotoxin from the puffer fish Fugu poecilonotus. Biosci. Biotechnol. Biochem. 1999, 63, 961–963. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Finn, J.; Fukushima, K.; Sakugawa, S.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Isolation of 6-deoxytetrodotoxin from the pufferfish, Takifugu pardalis, and a comparison of the effects of the C-6 and C-11 hydroxy groups of tetrodotoxin on its activity. J. Nat. Prod. 2014, 77, 1000–1004. [Google Scholar] [CrossRef]
- Kudo, K.; Yamashita, Y.; Mebs, D.; Cho, Y.; Konoki, K.; Yasumoto, T.; Yotsu-Yamashita, M. C5-C10 directly bonded tetrodotoxin analogues: Possible biosynthetic precursors of tetrodotoxin from newts. Angew Chem. Int. Ed. 2014, 53, 14546–14549. [Google Scholar] [CrossRef]
- Suo, R.; Tanaka, M.; Oyama, H.; Kojima, Y.; Yui, K.; Sakakibara, R.; Adachi, M.; Nishikawa, T.; Sugita, H.; Itoi, S. Tetrodotoxin in the flatworm Planocera multitentaculata. Toxicon 2022, 216, 169–173. [Google Scholar] [CrossRef]
- Ito, M.; Furukawa, R.; Yasukawa, S.; Sato, M.; Oyama, H.; Okabe, T.; Suo, R.; Sugita, H.; Takatani, T.; Arakawa, O.; et al. Local differences in the toxin amount and composition of tetrodotoxin and related compounds in pufferfish (Chelonodon patoca) and toxic goby (Yongeichthys criniger) juveniles. Toxins 2022, 14, 150. [Google Scholar] [CrossRef]
- Oyama, H.; Ito, M.; Suo, R.; Goto-Inoue, N.; Morisasa, M.; Mori, T.; Sugita, H.; Mori, T.; Nakahigashi, R.; Adachi, M.; et al. Changes in tissue distribution of tetrodotoxin and its analogues in association with maturation in the toxic flatworm, Planocera multitentaculata. Mar. Biotechnol. 2022, 24, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. D 2006, 1, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Ogata, T.; Sato, S. External secretion of tetrodotoxin from puffer fishes stimulated by electric shock. Mar. Biol. 1985, 87, 199–202. [Google Scholar] [CrossRef]
- Matsumura, K. Tetrodotoxin as a pheromone. Nature 1995, 378, 563–564. [Google Scholar] [CrossRef]
- Okita, K.; Yamazaki, H.; Sakiyama, K.; Yamane, H.; Niina, S.; Takatani, T.; Arakawa, O.; Sakakura, Y. Puffer smells tetrodotoxin. Ichthyol. Res. 2013, 60, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Ishizuka, K.; Mitsuoka, R.; Takimoto, N.; Yokoyama, N.; Detake, A.; Takayanagi, C.; Yoshikawa, S.; Sugita, H. Seasonal changes in the tetrodotoxin content of the pufferfish Takifugu niphobles. Toxicon 2016, 114, 53–58. [Google Scholar] [CrossRef]
- Itoi, S.; Yoshikawa, S.; Asahina, K.; Suzuki, M.; Ishizuka, K.; Takimoto, N.; Mitsuoka, R.; Yokoyama, N.; Detake, A.; Takayanagi, C.; et al. Larval pufferfish protected by maternal tetrodotoxin. Toxicon 2014, 78, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Itoi, S.; Suzuki, M.; Asahina, K.; Sawayama, E.; Nishikubo, J.; Oyama, H.; Takei, M.; Shiibashi, N.; Takatani, T.; Arakawa, O.; et al. Role of maternal tetrodotoxin in survival of larval pufferfish. Toxicon 2018, 148, 95–100. [Google Scholar] [CrossRef]
- Nozaki, M.; Tsutsumi, T.; Kobayashi, H.; Takei, Y.; Ichikawa, T.; Tsuneki, K.; Miyagawa, K.; Uemura, H.; Tatsumi, Y. Spawning habit of the puffer, Fugu niphobles (Jordan et Snyder) I. Zool. Mag. 1976, 85, 156–168, (in Japanese with English abstract). [Google Scholar]
- Suzuka, M.; Isogai, S. Spawning habit of the puffer, Fugu niphobles (Jordan et Snyder) at Koshigoe Beach of the Miura Peninsula. Sci. Rep. Yokosuka City Mus. 1979, 24, 47–66, pl. 9 (in Japanese with English abstract). [Google Scholar]
- Townsend, K.A.; Altvater, J.; Thomas, M.C.; Schuyler, Q.A.; Nette, G.W. Death in the octopus’ garden: Fatal blue-lined octopus envenomations of adult green sea turtles. Mar. Biol. 2012, 159, 689–695. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Emoto, Y.; Tatsuno, R.; Wang, J.J.; Ngy, L.; Taniyama, S.; Takatani, T.; Arakawa, O. Maturation-associated changes in toxicity of the pufferfish Takifugu poecilonotus. Toxicon. 2010, 55, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Kodama, M.; Sato, S.; Ogata, T.; Suzuki, Y.; Kaneko, T.; Aida, K. Tetrodotoxin secreting glands in the skin of puffer fishes. Toxicon 1986, 24, 819–829. [Google Scholar] [CrossRef]
- Noguchi, Y.; Suzuki, T.; Matsutani, K.; Sakakibara, R.; Nakahigashi, R.; Adachi, M.; Nishikawa, T.; Abe, H. An almost nontoxic tetrodotoxin analog, 5,6,11-trideoxytetrodotoxin, as an odorant for the grass puffer. Sci. Rep. 2022, 12, 15087. [Google Scholar] [CrossRef]
- Suzuki, T.; Nakahigashi, R.; Adachi, M.; Nishikawa, T.; Abe, H. Green spotted puffers detect a nontoxic TTX analog odor using crypt olfactory sensory neurons. Chem. Senses 2022, 47, bjac011. [Google Scholar] [CrossRef]
- Yasumoto, T.; Yotsu-Yamashita, M. Chemical and etiological studies on tetrodotoxin and its analogs. J. Toxicol. Toxin Rev. 1996, 15, 81–90. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kobayashi, H.; Takei, Y.; Nozaki, M. Spawning habit of the puffer, Fugu niphobles (Jordan et Snyder) II. Zool. Mag. 1978, 87, 44–55, (in Japanese with English abstract). [Google Scholar]
- Noguchi, T.; Hashimoto, K. Tetrodotoxin–A review with special reference to the assumed mechanism of toxification of puffers. Food Hyg. Saf. Sci. 1984, 25, 481–487. [Google Scholar] [CrossRef]
- Adachi, M.; Sakakibara, R.; Satake, Y.; Isobe, M.; Nishikawa, T. Synthesis of 5,6,11-trideoxytetrodotoxin. Chem. Lett. 2014, 43, 1719–1721. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Sex | No. of Specimen | TTX 1 | 5,6,11-trideoxyTTX 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total Amount (μg) | Tissue Distribution (%) | Total Amount (μg) | Tissue Distribution (%) | |||||||||
| Skin | Liver | Gonad | Others | Skin | Liver | Gonad | Others | |||||
| Kamogawa | Male | 11 | 573 ± 294 a | 36.8 ± 21.2 | 31.2 ± 13.8 a | 5.7 ± 4.2 b | 26.4 ± 12.8 a | 337 ± 197 a | 69.9 ± 18.8 a | 9.5 ± 7.7 a,b | 2.4 ± 1.9 a,b | 18.2 ± 11.6 a,b |
| Female | 9 | 734 ± 799 a | 25.0 ± 11.1 | 12.6 ± 6.4 b | 54.3 ± 15.0 a | 8.0 ± 6.1 a,b | 667 ± 846 a | 47.2 ± 18.2 a,b | 1.6 ± 0.7 b,c | 46.7 ± 20.8 a | 4.5 ± 5.7 b | |
| Enoshima | Male | 14 | 301 ± 252 b | 44.4 ± 18.4 | 33.7 ± 12.6 a | 1.6 ± 0.7 c | 20.2 ± 8.4 a | 117 ± 69 b | 71.4 ± 17.0 a,b | 6.2 ± 10.7 a | 1.7 ± 1.2 b,c | 20.8 ± 7.9 a |
| Female | 12 | 267 ± 301 b | 30.8 ± 13.5 | 2.0 ± 4.0 c | 59.2 ± 18.3 a | 7.9 ± 10.5 b | 211 ± 216 a,b | 38.8 ± 20.4 b | 0.5 ± 0.6 c | 54.7 ± 23.8 a | 6.1 ± 5.7 b | |
| Locality | Sex | Skin | Liver | Gonad | Others | Whole Body |
|---|---|---|---|---|---|---|
| Kamogawa | Male | 1.1 ± 0.7 | 24.2 ± 25.9 | 3.8 ± 2.1 | 3.5 ± 3.3 | 2.0 ± 0.8 |
| Female | 0.9 ± 0.4 | 9.0 ± 6.8 | 1.8 ± 0.8 | 1.6 ± 0.8 | 1.4 ± 0.5 | |
| Enoshima | Male | 1.7 ± 0.9 | 15.3 ± 11.4 | 2.2 ± 1.5 | 2.6 ± 1.5 | 2.8 ± 1.6 a |
| Female | 1.3 ± 0.9 | 4.4 ± 2.3 | 1.9 ± 1.4 | 1.5 ± 0.7 | 1.3 ± 0.4 b |
| Locality | Sex | No. of Specimen | Body Weight (g) | Tissue Weight (g) | |||
|---|---|---|---|---|---|---|---|
| Skin | Liver | Gonad | Others | ||||
| Kamogawa | Male | 11 | 36 ± 11 b | 4.4 ± 1.0 | 1.3 ± 0.5 | 6.1 ± 2.5 | 25 ± 7.2 |
| Female | 9 | 56 ± 14 a | 7.4 ± 1.8 | 2.0 ± 0.7 | 6.4 ± 4.1 | 40 ± 10 | |
| Enoshima | Male | 14 | 38 ± 12 b | 4.7 ± 1.3 | 1.8 ± 1.3 | 6.0 ± 2.3 | 26 ± 7.6 |
| Female | 12 | 56 ± 16 a | 7.0 ± 2.1 | 2.8 ± 1.5 | 7.5 ± 5.3 | 39 ± 11 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asano, M.; Ishizaki, C.; Tomonou, T.; Kihara, M.; Ito, M.; Yasukawa, S.; Shirai, K.; Oyama, H.; Izawa, S.; Kawamura, R.; et al. Levels of Tetrodotoxins in Spawning Pufferfish, Takifugu alboplumbeus. Mar. Drugs 2023, 21, 207. https://doi.org/10.3390/md21040207
Asano M, Ishizaki C, Tomonou T, Kihara M, Ito M, Yasukawa S, Shirai K, Oyama H, Izawa S, Kawamura R, et al. Levels of Tetrodotoxins in Spawning Pufferfish, Takifugu alboplumbeus. Marine Drugs. 2023; 21(4):207. https://doi.org/10.3390/md21040207
Chicago/Turabian StyleAsano, Masaki, Chihiro Ishizaki, Taiga Tomonou, Masato Kihara, Masaaki Ito, Shino Yasukawa, Kyoko Shirai, Hikaru Oyama, Shin Izawa, Reona Kawamura, and et al. 2023. "Levels of Tetrodotoxins in Spawning Pufferfish, Takifugu alboplumbeus" Marine Drugs 21, no. 4: 207. https://doi.org/10.3390/md21040207
APA StyleAsano, M., Ishizaki, C., Tomonou, T., Kihara, M., Ito, M., Yasukawa, S., Shirai, K., Oyama, H., Izawa, S., Kawamura, R., Saito, K., Suo, R., Nakahigashi, R., Adachi, M., Nishikawa, T., Sugita, H., & Itoi, S. (2023). Levels of Tetrodotoxins in Spawning Pufferfish, Takifugu alboplumbeus. Marine Drugs, 21(4), 207. https://doi.org/10.3390/md21040207