
Identification and Functional Analysis of Two Novel Genes—Geranylgeranyl Pyrophosphate Synthase Gene (AlGGPPS) and Isopentenyl Pyrophosphate Isomerase Gene (AlIDI)—from Aurantiochytrium limacinum Significantly Enhance De Novo β-Carotene Biosynthesis in Escherichia coli


Abstract
:
1. Introduction
2. Results
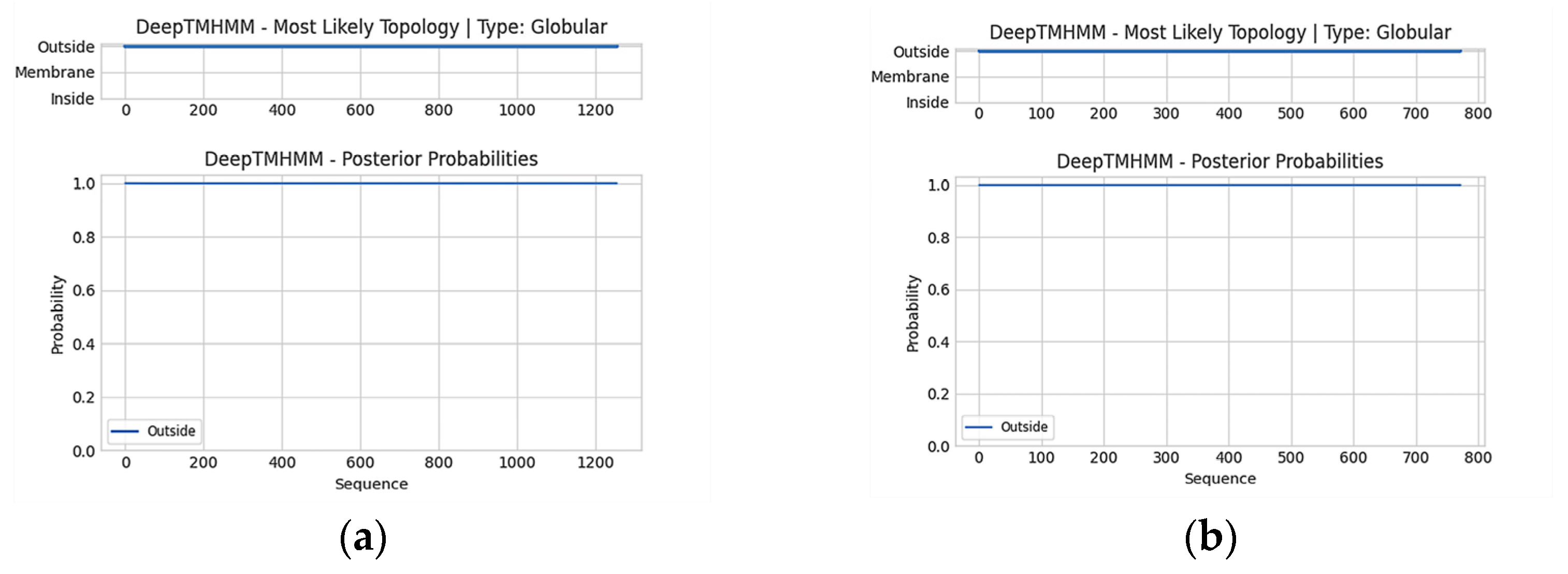
2.1. Bioinformatic Analysis of AlGGPPS and AlIDI
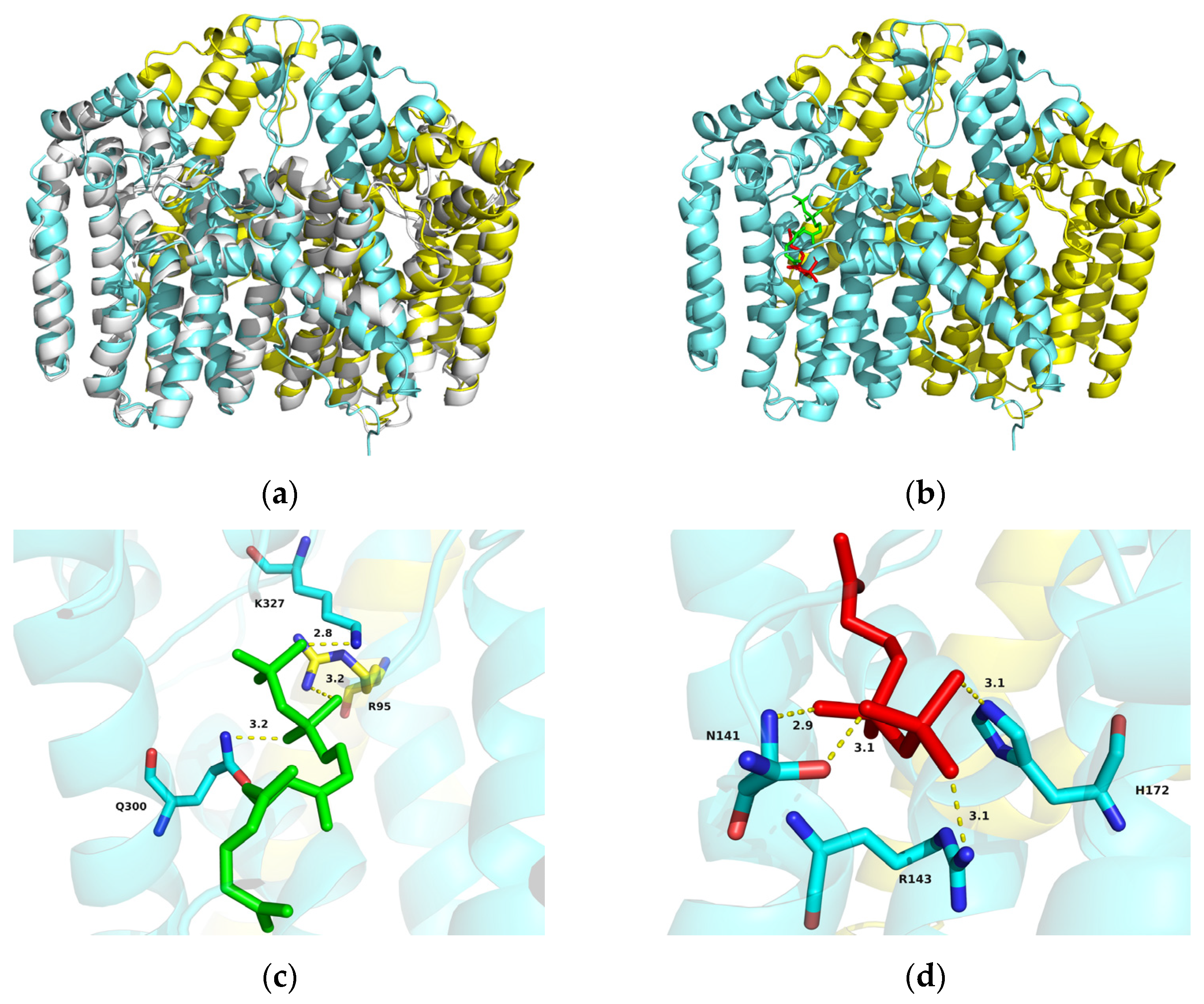
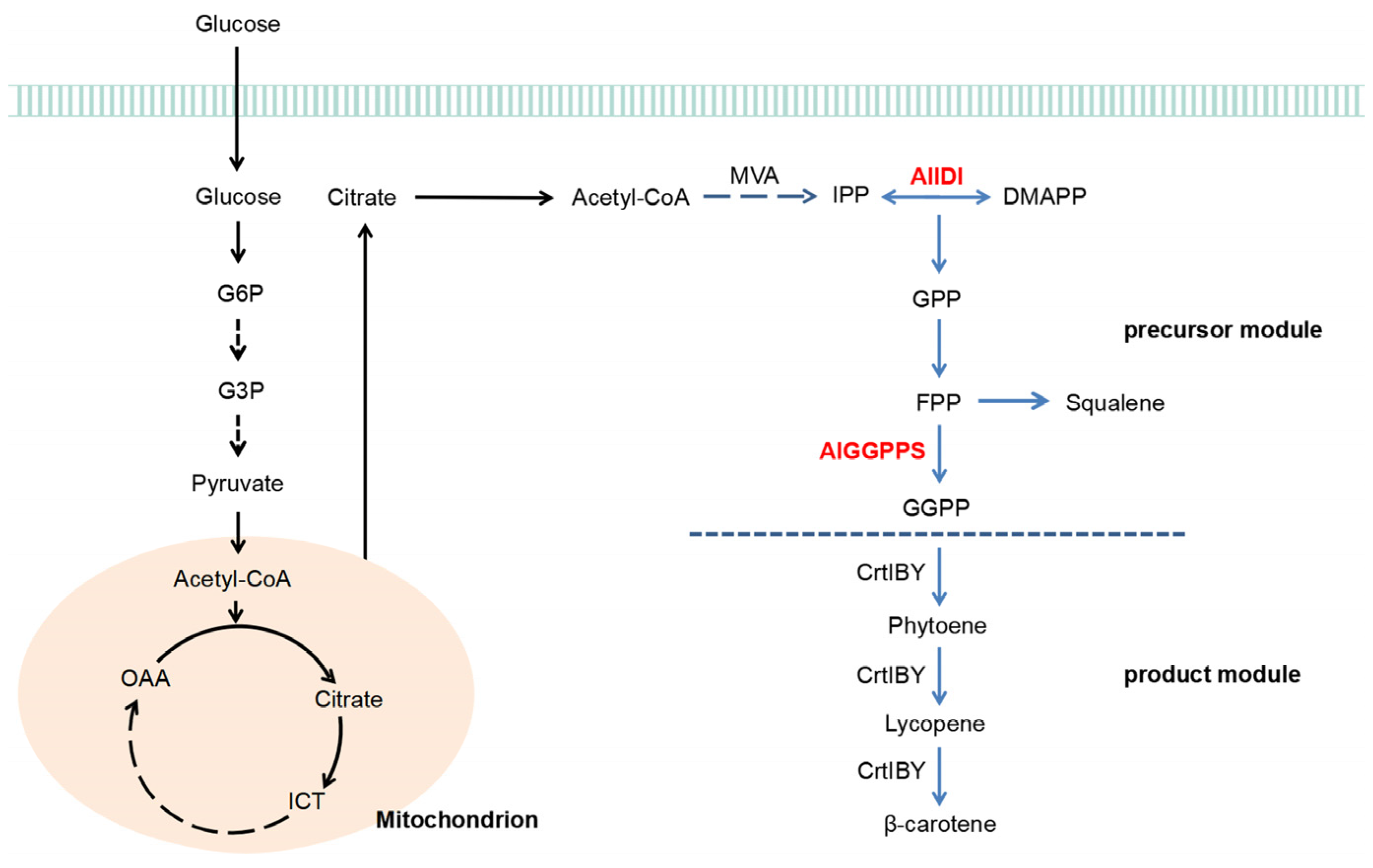
2.2. Identification of AlGGPPS for β-Carotene Biosynthesis
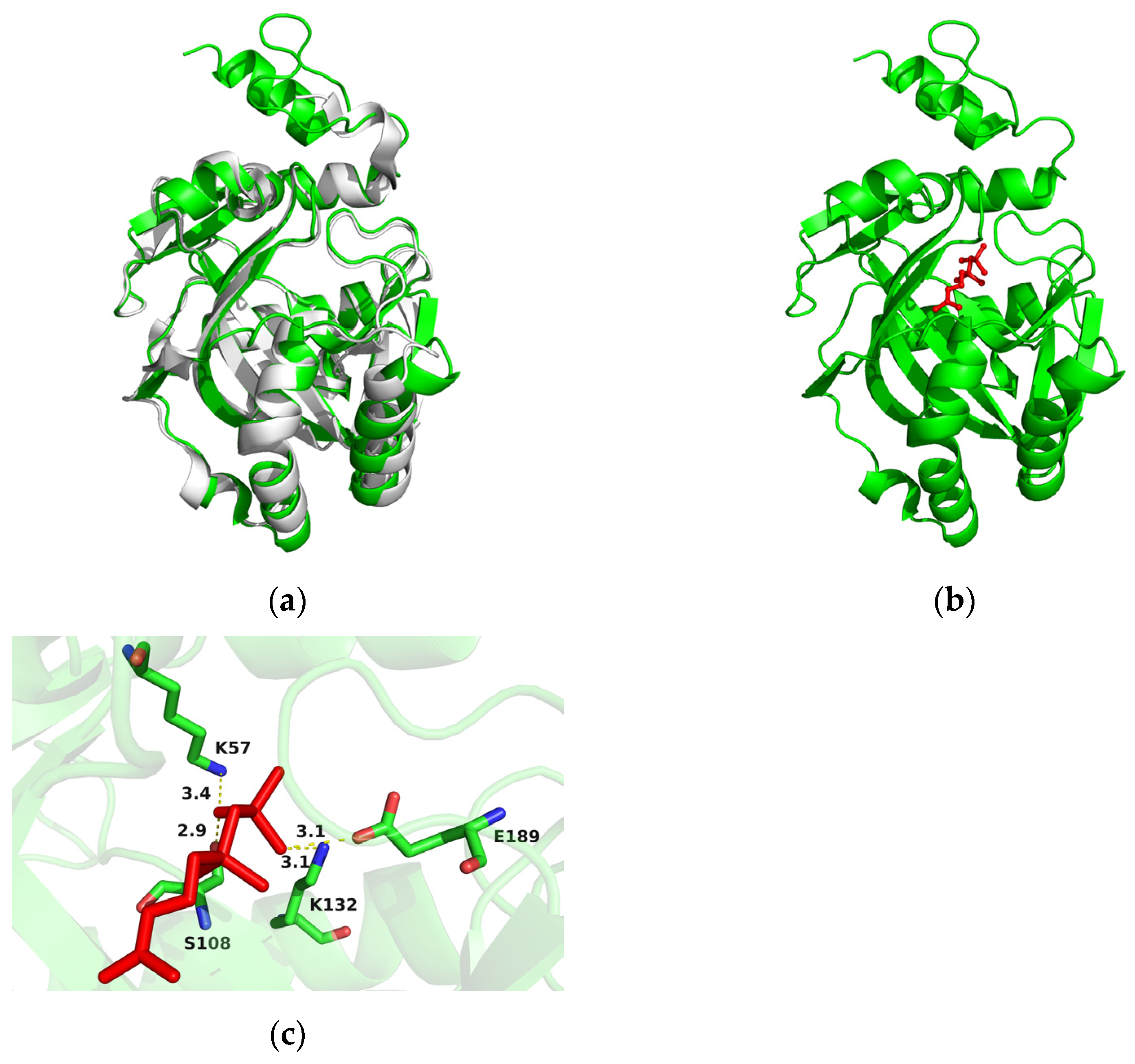
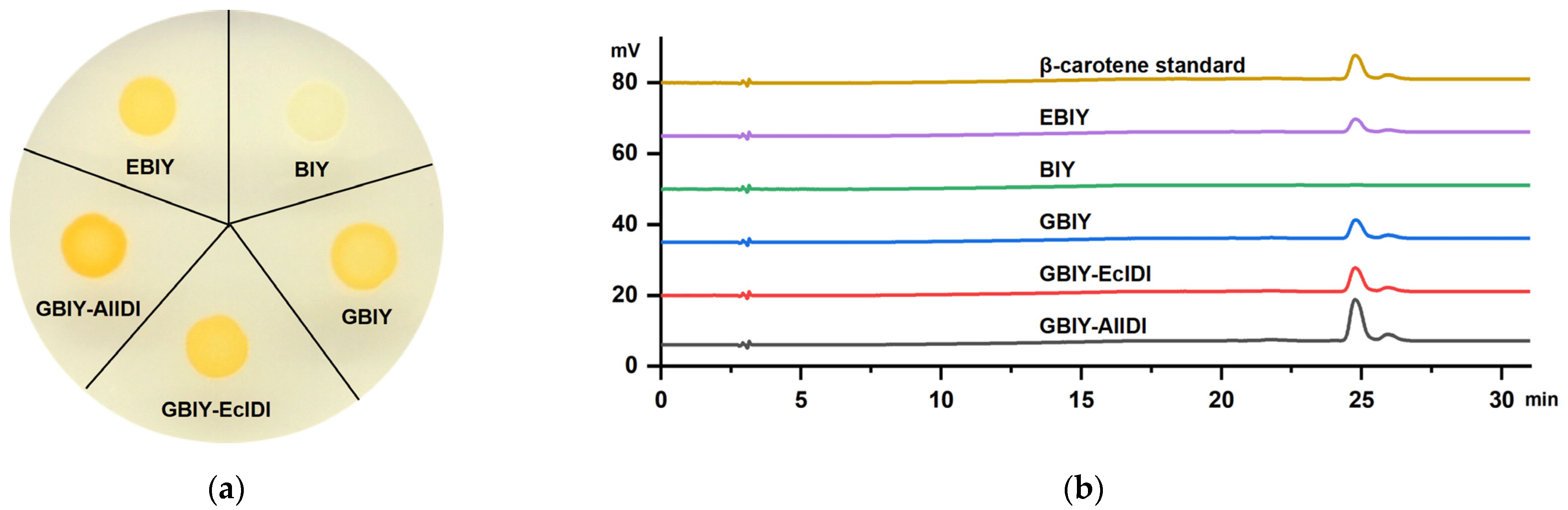
2.3. Overexpression of AlIDI for Improving β-Carotene Production
3. Discussion
4. Materials and Methods
4.1. Strains, Media and Growth Conditions
4.2. Bioinformatic Analysis
4.3. RNA Extraction, cDNA Synthesis and Genomic DNA Extraction
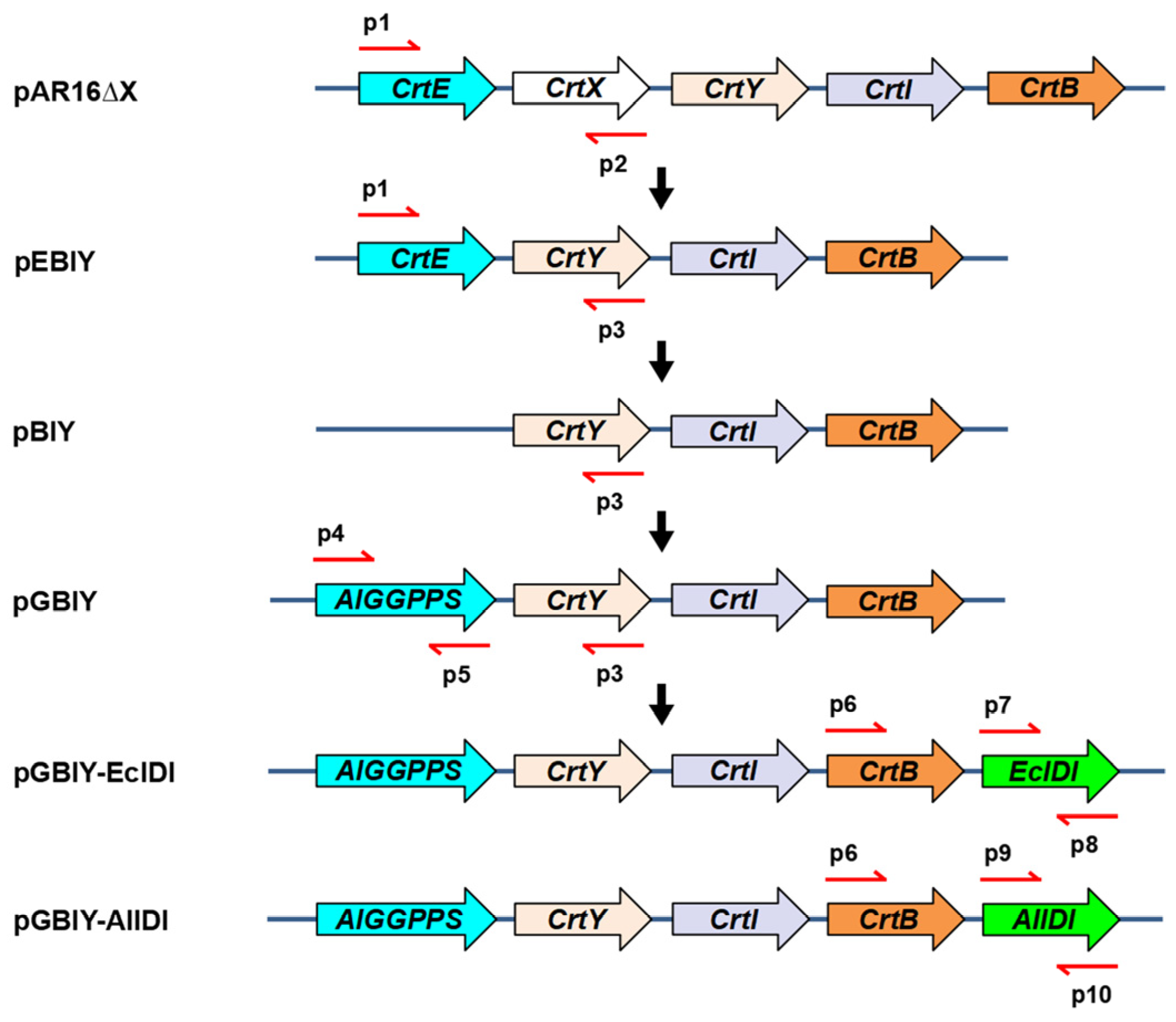
4.4. Gene Cloning and Construction of Plasmid and Strain
4.5. Extraction and Measurement of β-Carotene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Wang, Y.; He, Y.; Liu, B.; Mou, H.; Chen, F.; Yang, S. Microalgae-derived pigments for the food industry. Mar. Drugs 2023, 21, 82. [Google Scholar] [CrossRef] [PubMed]
- Morone, J.; Lopes, G.; Morais, J.; Neves, J.; Vasconcelos, V.; Martins, R. Cosmetic application of cyanobacteria extracts with a sustainable vision to skincare: Role in the antioxidant and antiaging process. Mar. Drugs 2022, 20, 761. [Google Scholar] [CrossRef]
- Giani, M.; Gervasi, L.; Loizzo, M.R.; Martinez-Espinosa, R.M. Carbon source influences antioxidant, antiglycemic, and antilipidemic activities of haloferax mediterranei carotenoid extracts. Mar. Drugs 2022, 20, 659. [Google Scholar] [CrossRef] [PubMed]
- Kandy, S.K.; Nimonkar, M.M.; Dash, S.S.; Mehta, B.; Markandeya, Y.S. Astaxanthin protection against neuronal excitotoxicity via glutamate receptor inhibition and improvement of mitochondrial function. Mar. Drugs 2022, 20, 645. [Google Scholar] [CrossRef]
- Capelli, B.; Bagchi, D.; Cysewski, G.R. Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods 2013, 12, 145–152. [Google Scholar] [CrossRef]
- Ye, L.; Zhu, X.; Wu, T.; Wang, W.; Zhao, D.; Bi, C.; Zhang, X. Optimizing the localization of astaxanthin enzymes for improved productivity. Biotechnol. Biofuels 2018, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Binkley, R.M.; Kim, W.J.; Lee, M.H.; Lee, S.Y. Metabolic engineering of Escherichia coli for high-level astaxanthin production with high productivity. Metab. Eng. 2018, 49, 105–115. [Google Scholar] [CrossRef]
- Zhang, C.; Seow, V.Y.; Chen, X.; Too, H.P. Multidimensional heuristic process for high-yield production of astaxanthin and fragrance molecules in Escherichia coli. Nat. Commun. 2018, 9, 1858. [Google Scholar] [CrossRef]
- Jiang, G.; Yang, Z.; Wang, Y.; Yao, M.; Chen, Y.; Xiao, W.; Yuan, Y. Enhanced astaxanthin production in yeast via combined mutagenesis and evolution. Biochem. Eng. J. 2020, 156, 107519. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, W.; Zhang, X.; Fan, X.; Xu, D.; Ye, N.; Su, Z.; Yu, J.; Yang, Q. Isolation and expression analyses of methyl-d-erythritol 4-phosphate (MEP) pathway genes from Haematococcus pluvialis. J. Appl. Phycol. 2016, 28, 209–218. [Google Scholar] [CrossRef]
- Li, M.; Xia, Q.; Zhang, H.; Zhang, R.; Yang, J. Metabolic engineering of different microbial hosts for lycopene production. J. Agric. Food Chem. 2020, 68, 14104–14122. [Google Scholar] [CrossRef]
- Rinaldi, M.A.; Ferraz, C.A.; Scrutton, N.S. Alternative metabolic pathways and strategies to high-titre terpenoid production in Escherichia coli. Nat. Prod. Rep. 2022, 39, 90–118. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.H. Reaction Kinetics, Catalytic Mechanisms, Conformational Changes, and Inhibitor Design for Prenyltransferases. Biochemistry 2009, 48, 6562–6570. [Google Scholar] [CrossRef]
- Gwak, Y.; Hwang, Y.; Wang, B.; Kim, M.; Jeong, J.; Lee, C.G.; Hu, Q.; Han, D.; Jin, E. Comparative analyses of lipidomes and transcriptomes reveal a concerted action of multiple defensive systems against photooxidative stress in Haematococcus pluvialis. J. Exp. Bot. 2014, 65, 4317–4334. [Google Scholar] [CrossRef] [PubMed]
- Coman, D.; Altenhoff, A.; Zoller, S.; Gruissem, W.; Vranova, E. Distinct evolutionary strategies in the GGPPS family from plants. Front. Plant Sci. 2014, 5, 230. [Google Scholar] [CrossRef]
- Alcaino, J.; Romero, I.; Niklitschek, M.; Sepulveda, D.; Cecilia Rojas, M.; Baeza, M.; Cifuentes, V. Functional Characterization of the Xanthophyllomyces dendrorhous Farnesyl Pyrophosphate Synthase and Geranylgeranyl Pyrophosphate Synthase Encoding Genes That Are Involved in the Synthesis of Isoprenoid Precursors. PLoS ONE 2014, 9, e96626. [Google Scholar] [CrossRef]
- Higuera-Ciapara, I.; Felix-Valenzuela, L.; Goycoolea, F.M. Astaxanthin: A review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 2006, 46, 185–196. [Google Scholar] [CrossRef]
- Lao, Y.; Xiao, L.; Ye, Z.; Jiang, J.; Zhou, S. In silico analysis of phytoene synthase and its promoter reveals hints for regulation mechanisms of carotenogenesis in Duanliella bardawil. Bioinformatics 2011, 27, 2201–2208. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercia products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Song, X.; Zhang, L.; Cui, J.; Chen, L.; Zhang, W. Tailoring cyanobacteria as a new platform for highly efficient synthesis of astaxanthin. Metab. Eng. 2020, 61, 275–287. [Google Scholar] [CrossRef]
- Dong, C.; Qu, G.; Guo, J.; Wei, F.; Gao, S.; Sun, Z.; Jin, L.; Sun, X.; Rochaix, J.D.; Miao, Y.; et al. Rational design of geranylgeranyl diphosphate synthase enhances carotenoid production and improves photosynthetic efficiency in Nicotiana tabacum. Sci. Bull. 2022, 67, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yuan, Y.; Chen, Q.; Nie, S.; Guo, J.; Ou, Z.; Huang, M.; Deng, Z.; Liu, T.; Ma, T. Metabolic pathway assembly using docking domains from type I cis-AT polyketide synthases. Nat. Commun. 2022, 13, 5541. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Peters-Wendisch, P.; Beekwilder, J.; Wendisch, V.F. IdsA is the major geranylgeranyl pyrophosphate synthase involved in carotenogenesis in Corynebacterium glutamicum. FEBS. J. 2014, 281, 4906–4920. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, X.; Cao, T.; Zhuang, Z.; Wang, R.; Lu, S. Heteromeric geranylgeranyl diphosphate synthase contributes to carotenoid biosynthesis in ripening fruits of red pepper (Capsicum annuum var. conoides). J. Agric. Food Chem. 2018, 66, 11691–11700. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, W.; Li, A.; Wang, C.; Hu, Z. Discovery of geranylgeranyl pyrophosphate synthase (GGPPS) paralogs from Haematococcus pluvialis based on iso-seq analysis and their function on astaxanthin biosynthesis. Mar. Drugs 2019, 17, 696. [Google Scholar] [CrossRef]
- Wang, J.; Lin, H.; Su, P.; Chen, T.; Guo, J.; Gao, W.; Huang, L. Molecular cloning and functional characterization of multiple geranylgeranyl pyrophosphate synthases (ApGGPPS) from Andrographis paniculata. Plant. Cell. Rep. 2019, 38, 117–128. [Google Scholar] [CrossRef]
- Zhou, F.; Pichersky, E. The complete functional characterisation of the terpene synthase family in tomato. New. Phytol. 2020, 226, 1341–1360. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, H.; Zong, Y.; Tu, Z.; Li, H. Isolation, expression, and functional analysis of the geranylgeranyl pyrophosphate synthase (GGPPS) gene from Liriodendron tulipifera. Plant. Physiol. Bioch. 2021, 166, 700–711. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Peters-Wendisch, P.; Netzer, R.; Stafnes, M.; Brautaset, T.; Wendisch, V.F. Production and glucosylation of C-50 and C-40 carotenoids by metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biot. 2014, 98, 1223–1235. [Google Scholar] [CrossRef]
- Yang, L.; Huang, X.; Lu, Q.; Zhu, J.; Lu, S. Cloning and characterization of the geranylgeranyl diphosphate synthase (GGPS) responsible for carotenoid biosynthesis in Pyropia umbilicalis. J. Appl. Phycol. 2016, 28, 671–678. [Google Scholar] [CrossRef]
- Johnson, M.B.; Wen, Z. Production of biodiesel fuel from the microalga Schizochytrium limacinum by direct transesterification of algal biomass. Energy. Fuels 2009, 23, 5179–5183. [Google Scholar] [CrossRef]
- Gupta, A.; Barrow, C.J.; Puri, M. Omega-3 biotechnology: Thraustochytrids as a novel source of omega-3 oils. Biotechnol. Adv. 2012, 30, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Valdebenito, D.; Urrutia, S.; Leyton, A.; Chisti, Y.; Asenjo, J.A.; Shene, C. Nitrogen sources affect the long-chain polyunsaturated fatty acids content in Thraustochytrium sp. RT2316-16. Mar. Drugs 2023, 21, 15. [Google Scholar] [CrossRef] [PubMed]
- Hien, H.T.M.; Thom, L.T.; Ha, N.C.; Tam, L.T.; Thu, N.T.H.; Nguyen, T.V.; Loan, V.T.; Dan, N.T.; Hong, D.D. Characterization and optimization of culture conditions for Aurantiochytrium sp. SC145 isolated from Sand Cay (Son Ca) Island, Vietnam, and antioxidative and neuroprotective activities of its polyunsaturated fatty acid mixture. Mar. Drugs 2022, 20, 780. [Google Scholar] [CrossRef]
- Ji, X.; Ren, L.; Huang, H. Omega-3 biotechnology: A green and sustainable process for omega-3 fatty acids production. Front. Bioeng. Biotech. 2015, 3, 158. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, H.; Zhang, K.; Gao, F.; Chen, S.; Li, D. Physicochemical properties and storage stability of microencapsulated DHA-rich oil with different wall materials. Appl. Biochem. Biotech. 2016, 179, 1129–1142. [Google Scholar] [CrossRef]
- Cheng, Y.; Sun, Z.; Cui, G.; Song, X.; Cui, Q. A new strategy for strain improvement of Aurantiochytrium sp. based on heavy-ions mutagenesis and synergistic effects of cold stress and inhibitors of enoyl-ACP reductase. Enzyme. Microb. Tech. 2016, 93–94, 182–190. [Google Scholar] [CrossRef]
- Raghukumar, S. Thraustochytrid marine protists: Production of PUFAs and other emerging technologies. Mar. Biotechnol. 2008, 10, 631–640. [Google Scholar] [CrossRef]
- Chang, G.; Gao, N.; Tian, G.; Wu, Q.; Chang, M.; Wang, X. Improvement of docosahexaenoic acid production on glycerol by Schizochytrium sp. S31 with constantly high oxygen transfer coefficient. Bioresour. Technol. 2013, 142, 400–406. [Google Scholar] [CrossRef]
- Guo, D.; Ji, X.; Ren, L.; Li, G.; Sun, X.; Chen, K.; Gao, S.; Huang, H. Development of a scale-up strategy for fermentative production of docosahexaenoic acid by Schizochytrium sp. Chem. Eng. Sci. 2018, 176, 600–608. [Google Scholar] [CrossRef]
- Quilodran, B.; Hinzpeter, I.; Hormazabal, E.; Quiroz, A.; Shene, C. Docosahexaenoic acid (C22:6n-3, DHA) and astaxanthin production by Thraustochytriidae sp. AS4-A1 a native strain with high similitude to Ulkenia sp.: Evaluation of liquid residues from food industry as nutrient sources. Enzyme. Microb. Tech. 2010, 47, 24–30. [Google Scholar] [CrossRef]
- Singh, P.; Liu, Y.; Li, L.; Wang, G. Ecological dynamics and biotechnological implications of thraustochytrids from marine habitats. Appl. Microbiol. Biot. 2014, 98, 5789–5805. [Google Scholar] [CrossRef] [PubMed]
- Aasen, I.M.; Ertesvag, H.; Heggeset, T.M.B.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biot. 2016, 100, 4309–4321. [Google Scholar] [CrossRef]
- Ren, L.; Sun, X.; Zhang, L.; Huang, H.; Zhao, Q. Exergy analysis for docosahexaenoic acid production by fermentation and strain improvement by adaptive laboratory evolution for Schizochytrium sp. Bioresour. Technol. 2020, 298, 122562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Chen, L.; Liu, J.; Gao, F.; He, R.; Chen, W.; Guo, W.; Chen, S.; Li, D. Effects of butanol on high value product production in Schizochytrium limacinum B4D1. Enzyme. Microb. Tech. 2017, 102, 9–15. [Google Scholar] [CrossRef]
- Du, H.; Liao, X.; Gao, Z.; Li, Y.; Lei, Y.; Chen, W.; Chen, L.; Fan, X.; Zhang, K.; Chen, S.; et al. Effects of methanol on carotenoids as well as biomass and fatty acid biosynthesis in Schizochytrium limacinum B4D1. Appl. Environ. Microb. 2019, 85, e01243-19. [Google Scholar] [CrossRef]
- Zhu, X.; Meng, C.; Du, H.; Chen, L.; Sun, F.; Chen, W.; Wei, Z.; Ren, J.; Gao, Z.; Li, D. Enhancement of astaxanthin production in Schizochytrium limacinum B4D1 under ethanol induction. Algal. Res. 2022, 61, 102537. [Google Scholar] [CrossRef]
- Tang, X.; Man, Y.; Hu, X.; Xu, X.; Ren, L. Identification of carotenoids biosynthesis pathway in Schizochytrium sp. and utilization in astaxanthin biosynthesis. Enzyme. Microb. Tech. 2022, 156, 110018. [Google Scholar] [CrossRef]
- Iwasaka, H.; Koyanagi, R.; Satoh, R.; Nagano, A.; Watanabe, K.; Hisata, K.; Satoh, N.; Aki, T. A possible trifunctional beta-carotene synthase gene identified in the draft genome of Aurantiochytrium sp. strain KH105. Genes 2018, 9, 200. [Google Scholar] [CrossRef]
- Lacbay, C.M.; Waller, D.D.; Park, J.; Palou, M.G.; Vincent, F.; Huang, X.; Ta, V.; Berghuis, A.M.; Sebag, M.; Tsantrizos, Y.S. Unraveling the prenylation-cancer paradox in multiple myeloma with novel geranylgeranyl pyrophosphate synthase (GGPPS) inhibitors. J. Med. Chem. 2018, 61, 6904–6917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, L.; Xu, H.; Wei, Z.; Wang, Y.; Lin, Y.; Gong, W. Crystal structures of human IPP isomerase: New insights into the catalytic mechanism. J. Mol. Biol. 2007, 366, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, D.; Chen, H.; Wang, Q. Advances in Genetic Engineering in Improving Photosynthesis and Microalgal Productivity. Int. J. Mol. Sci. 2023, 24, 1898. [Google Scholar] [CrossRef]
- Sun, Z.; Chen, H.; Sun, L.; Wang, Q. Converting carbon dioxide to high value-added products: Microalgae-based green biomanufacturing. GCB Bioenergy 2023, 15, 386–398. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Q.; Sun, T.; Zhu, X.; Xu, H.; Tang, J.; Zhang, X.; Ma, Y. Engineering central metabolic modules of Escherichia coli for improving beta-carotene production. Metab. Eng. 2013, 17, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, M.; Liu, C.; Zhang, H.; Xian, M.; Liu, H. Enhancement of the catalytic activity of Isopentenyl diphosphate isomerase (IDI) from Saccharomyces cerevisiae through random and site-directed mutagenesis. Microb. Cell. Fact. 2018, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Bettiga, M.; Rova, U.; Christakopoulos, P.; Matsakas, L.; Patel, A. Structural and molecular characterization of squalene synthase belonging to the marine Thraustochytrid species Aurantiochytrium limacinum using bioinformatics approach. Mar. Drugs 2022, 20, 180. [Google Scholar] [CrossRef]
- Ignea, C.; Trikka, F.A.; Nikolaidis, A.K.; Georgantea, P.; Ioannou, E.; Loupassaki, S.; Kefalas, P.; Kanellis, A.K.; Roussis, V.; Makris, A.M.; et al. Efficient diterpene production in yeast by engineering Erg20p into a geranylgeranyl diphosphate synthase. Metab. Eng. 2015, 27, 65–75. [Google Scholar] [CrossRef]
- Ding, M.; Yan, H.; Li, L.; Zhai, F.; Shang, L.; Yin, Z.; Yuan, Y. Biosynthesis of taxadiene in Saccharomyces cerevisiae: Selection of geranylgeranyl diphosphate synthase directed by a computer-aided docking Strategy. PLoS ONE 2014, 9, e109348. [Google Scholar] [CrossRef]
- Engels, B.; Dahm, P.; Jennewein, S. Metabolic engineering of taxadiene biosynthesis in yeast as a first step towards Taxol (Paclitaxel) production. Metab. Eng. 2008, 10, 201–206. [Google Scholar] [CrossRef]
- Apel, A.R.; d’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.J.; et al. A Cas9-based toolkit to program gene expression in Saccharomyces cerevisiae. Nucleic. Acids. Res. 2017, 45, 496–508. [Google Scholar]
- Nowrouzi, B.; Li, R.A.; Walls, L.E.; d’Espaux, L.; Malci, K.; Liang, L.; Jonguitud-Borrego, N.; Lerma-Escalera, A.I.; Morones-Ramirez, J.R.; Keasling, J.D.; et al. Enhanced production of taxadiene in Saccharomyces cerevisiae. Microb. Cell. Fact. 2020, 19, 200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Gao, W.; Rong, Q.; Jin, G.; Chu, H.; Liu, W.; Yang, W.; Zhu, Z.; Li, G.; Zhu, G.; et al. Modular pathway engineering of diterpenoid synthases and the mevalonic acid pathway for miltiradiene production. J. Am. Chem. Soc. 2012, 134, 3234–3241. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, W.; Li, A.; Hu, Z.; Wang, J.; Wang, C. Cloning and identification of a novel beta-carotene hydroxylase gene from Haematococcus pluvialis and its function in Escherichia coli. Algal. Res. 2021, 55, 102245. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid/Strain | Description |
|---|---|
| pEBIY | CmR; p15A ori; concise CrtE-CrtB-CrtI-CrtY gene cluster from Pantoea ananatis |
| pBIY | CmR; p15A ori; CrtB-CrtI-CrtY gene cluster |
| pGBIY | CmR; p15A ori; AlGGPPS-CrtB-CrtI-CrtY gene cluster |
| pGBIY-EcIDI | CmR; p15A ori; AlGGPPS-CrtB-CrtI-CrtY gene cluster with E. coli IDI |
| pGBIY-AlIDI | CmR; p15A ori; AlGGPPS-CrtB-CrtI-CrtY gene cluster with Aurantiochytrium IDI |
| EBIY | BL21(DE3) stain bearing plasmid pEBIY |
| BIY | BL21(DE3) stain bearing plasmid pBIY |
| GBIY | BL21(DE3) stain bearing plasmid pGBIY |
| GBIY-EcIDI | BL21(DE3) stain bearing plasmid pGBIY-EcIDI |
| GBIY-AlIDI | BL21(DE3) stain bearing plasmid pGBIY-AlIDI |
| Primer | Sequence |
|---|---|
| P1 | ATGACGGTCTGCGCAAAAAAACACG |
| P2 | TGACCGGTGCACATAACCTGCTC |
| P3 | TTAACGATGAGTCGTCATAATGGCT |
| P4 | ATGCCAAGTGCAGGACCGG |
| P5 | CTAAATCTCGCATTCGTCAACCTCCT |
| P6 | ATGGCAGTTGGCTCGAAAAGTT |
| P7 | ATGCAAACGGAACACGTCATTTTAT |
| P8 | TTAATTGTGCTGCGCGAAAGCAGAC |
| P9 | ATGCGCATGACTTACTCCAACTT |
| P10 | CTAAGCGTGGGCAGGGGCGATAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, S.; Chang, Y.; Yu, J.; Chen, H.; Wang, Q.; Bi, Y. Identification and Functional Analysis of Two Novel Genes—Geranylgeranyl Pyrophosphate Synthase Gene (AlGGPPS) and Isopentenyl Pyrophosphate Isomerase Gene (AlIDI)—from Aurantiochytrium limacinum Significantly Enhance De Novo β-Carotene Biosynthesis in Escherichia coli. Mar. Drugs 2023, 21, 249. https://doi.org/10.3390/md21040249
Shi S, Chang Y, Yu J, Chen H, Wang Q, Bi Y. Identification and Functional Analysis of Two Novel Genes—Geranylgeranyl Pyrophosphate Synthase Gene (AlGGPPS) and Isopentenyl Pyrophosphate Isomerase Gene (AlIDI)—from Aurantiochytrium limacinum Significantly Enhance De Novo β-Carotene Biosynthesis in Escherichia coli. Marine Drugs. 2023; 21(4):249. https://doi.org/10.3390/md21040249
Chicago/Turabian StyleShi, Shitao, Yi Chang, Jinhui Yu, Hui Chen, Qiang Wang, and Yuping Bi. 2023. "Identification and Functional Analysis of Two Novel Genes—Geranylgeranyl Pyrophosphate Synthase Gene (AlGGPPS) and Isopentenyl Pyrophosphate Isomerase Gene (AlIDI)—from Aurantiochytrium limacinum Significantly Enhance De Novo β-Carotene Biosynthesis in Escherichia coli" Marine Drugs 21, no. 4: 249. https://doi.org/10.3390/md21040249
APA StyleShi, S., Chang, Y., Yu, J., Chen, H., Wang, Q., & Bi, Y. (2023). Identification and Functional Analysis of Two Novel Genes—Geranylgeranyl Pyrophosphate Synthase Gene (AlGGPPS) and Isopentenyl Pyrophosphate Isomerase Gene (AlIDI)—from Aurantiochytrium limacinum Significantly Enhance De Novo β-Carotene Biosynthesis in Escherichia coli. Marine Drugs, 21(4), 249. https://doi.org/10.3390/md21040249