
Saturated and Polyunsaturated Fatty Acids Production by Aurantiochytrium limacinum PKU#Mn4 on Enteromorpha Hydrolysate

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Sugar Content and Composition of Enteromorpha Hydrolysate
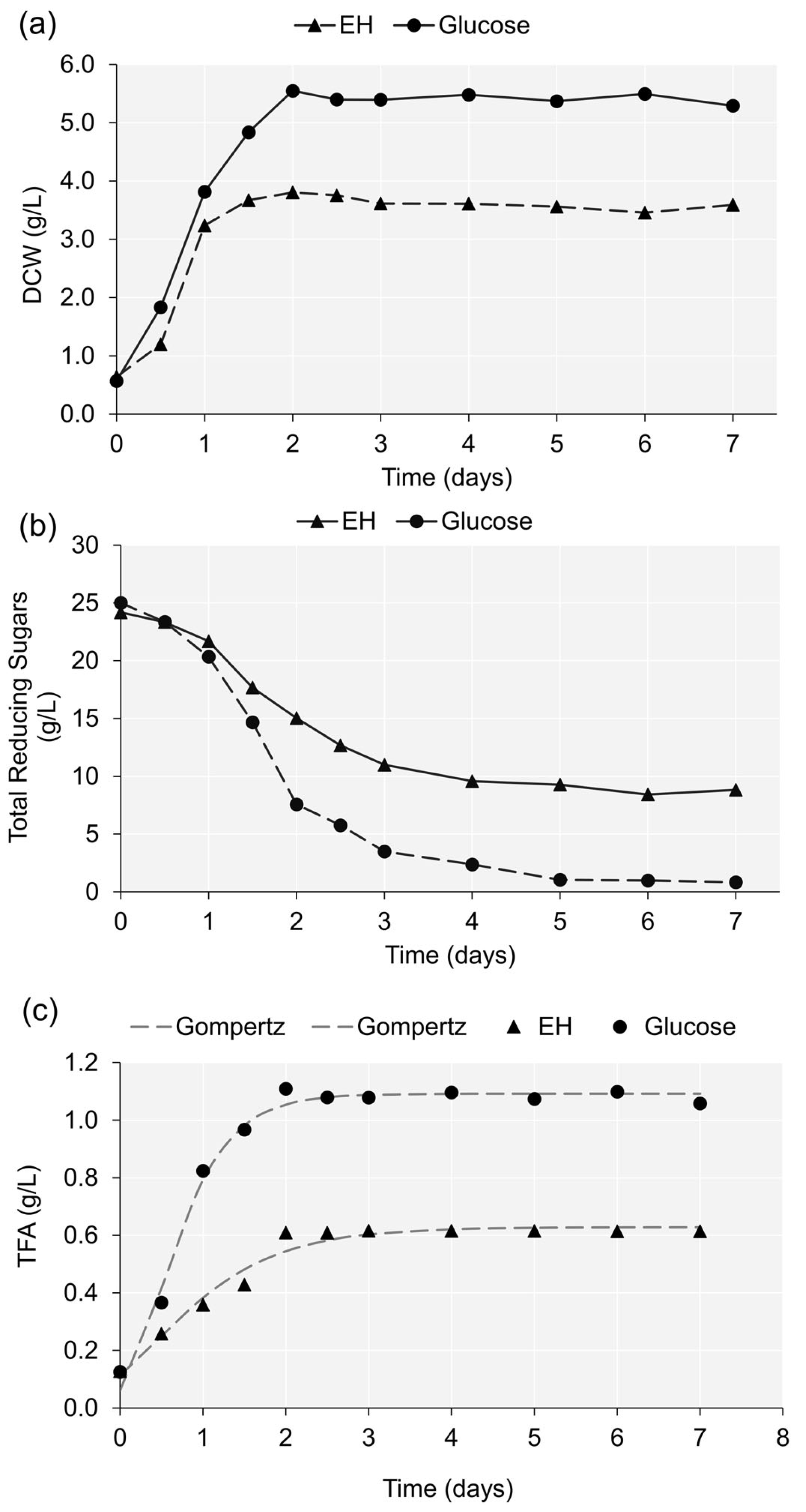
2.2. Lipid Production Potential of Enteromorpha Hydrolysate
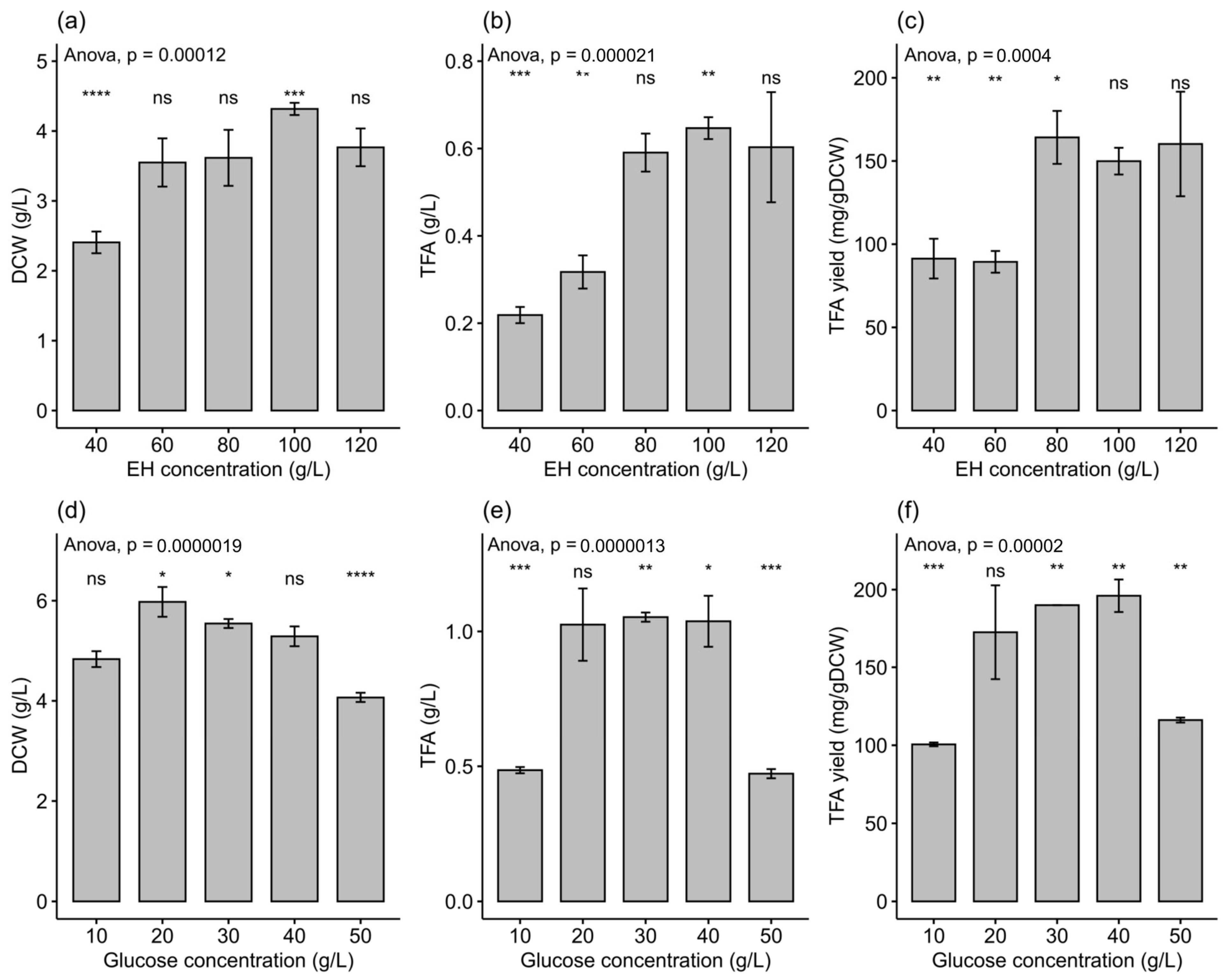
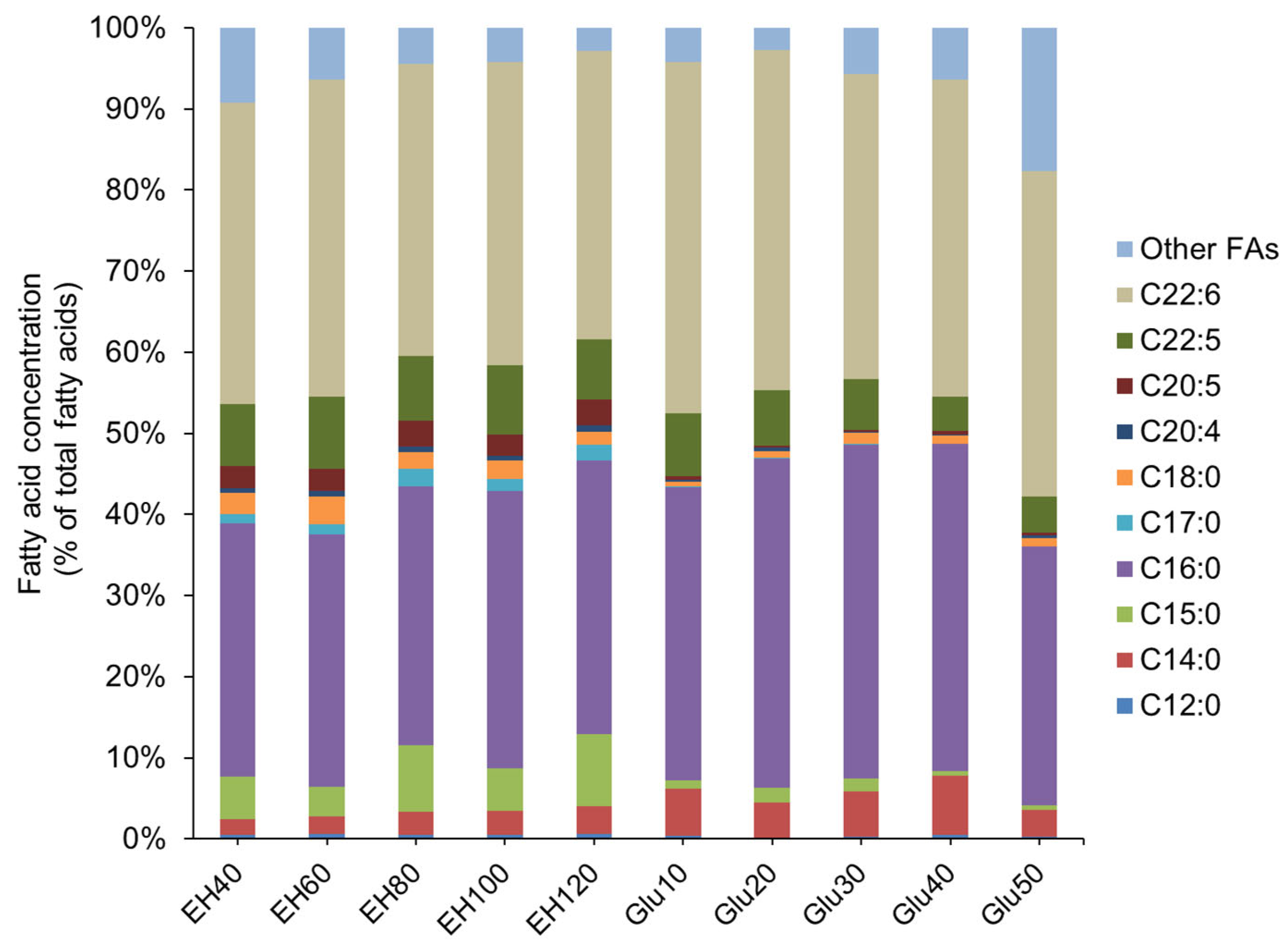
2.3. Effect of Enteromorpha Hydrolysate Concentration on Growth and Fatty Acid Content

{kind=link}
{kind=link}
{kind=link}
| Strain | Carbon Source | Biomass (g/L) | TFA (g/L) | Maximum TFA Yield (g/g Biomass) | Reference |
|---|---|---|---|---|---|
| Schizochytrium sp. HX-308 | Cane molasses | 25.54 | 5.21 | 0.20 | [22] |
| Schizochytrium sp. BCRC33482 | Sugarcane bagasse | 10.45 | 4.72 | 0.45 | [15] |
| Aurantiochytrium sp. YLH70 | Jerusalem artichoke | 32.71 | 19.72 | 0.60 | [16] |
| Aurantiochytrium limacinum SR21 | Sweet sorghum juice (50%) | 9.38 | 6.86 | 0.73 | [17] |
| Aurantiochytrium sp. KRS101 | Empty palm fruit bunches | 34.40 | 12.50 | 0.36 | [61] |
| Aurantiochytrium limacinum PKU#Mn4 | Enteromorpha hydrolysate | 4.32 | 0.65 | 0.16 | This study |
3. Materials and Methods
3.1. Enteromorpha Sampling
3.2. Preparation of Enteromorpha Hydrolysate Medium
3.3. Batch Fermentation
3.4. Analytical Methods
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, A.; Barrow, C.J.; Puri, M. Multiproduct biorefinery from marine thraustochytrids towards a circular bioeconomy. Trends Biotechnol. 2021, 40, 448–462. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of high added-value compounds-a brief review of recent work. Biotechnol. Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Pohnert, G.; Wei, D. Extracellular Metabolites from Industrial Microalgae and Their Biotechnological Potential. Mar. Drugs 2016, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Lim, D.K.Y.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Factories 2012, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, P.S.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Khan, T.M.Y.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae biomass as a sustainable source for biofuel, biochemical and biobased value-added products: An integrated biorefinery concept. Fuel 2022, 307, 121782. [Google Scholar] [CrossRef]
- Morabito, C.; Bournaud, C.; Maës, C.; Schuler, M.; Cigliano, R.A.; Dellero, Y.; Maréchal, E.; Amato, A.; Rébeillé, F. The lipid metabolism in thraustochytrids. Prog. Lipid Res. 2019, 76, 101007. [Google Scholar] [CrossRef]
- Patel, A.; Karageorgou, D.; Katapodis, P.; Sharma, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Bioprospecting of thraustochytrids for omega-3 fatty acids: A sustainable approach to reduce dependency on animal sources. Trends Food Sci. Technol. 2021, 115, 433–444. [Google Scholar] [CrossRef]
- Wang, Q.; Sen, B.; Liu, X.; He, Y.; Xie, Y.; Wang, G. Enhanced saturated fatty acids accumulation in cultures of newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. for large-scale biodiesel production. Sci. Total Environ. 2018, 631, 994–1004. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, H.; Sen, B.; Xie, Y.; He, Y.; Park, S.; Wang, G. Improved production of docosahexaenoic acid in batch fermentation by newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. through bioprocess optimization. Synth. Syst. Biotechnol. 2018, 3, 121–129. [Google Scholar] [CrossRef]
- Chen, X.; Sen, B.; Zhang, S.; Bai, M.; He, Y.; Wang, G. Chemical and physical culture conditions significantly influence the cell mass and docosahexaenoic acid content of Aurantiochytrium limacinum strain PKU#SW8. Mar. Drugs 2021, 19, 671. [Google Scholar] [CrossRef]
- Russo, G.L.; Langellotti, A.L.; Sacchi, R.; Masi, P. Techno-economic assessment of DHA-rich Aurantiochytrium sp. production using food industry by-products and waste streams as alternative growth media. Bioresour. Technol. Rep. 2022, 18, 100997. [Google Scholar] [CrossRef]
- Sun, X.M.; Xu, Y.S.; Huang, H. Thraustochytrid Cell Factories for Producing Lipid Compounds. Trends Biotechnol. 2021, 39, 648–650. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Zhang, M.; Chen, Y.; Jiang, X.; Lin, J.; Cao, X.; Huang, J. A lignocellulosic hydrolysate-tolerant Aurantiochytrium sp. mutant strain for docosahexaenoic acid production. Bioresour. Technol. 2017, 227, 221–226. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Su, C.-H.; Yu, Y.-K.; Huong, D.T.M. Sugarcane bagasse as a novel carbon source for heterotrophic cultivation of oleaginous microalga Schizochytrium sp. Ind. Crops Prod. 2018, 121, 99–105. [Google Scholar] [CrossRef]
- Yu, X.-J.; Liu, J.-H.; Sun, J.; Zheng, J.-Y.; Zhang, Y.-J.; Wang, Z. Docosahexaenoic acid production from the acidic hydrolysate of Jerusalem artichoke by an efficient sugar-utilizing Aurantiochytrium sp. YLH70. Ind. Crops Prod. 2016, 83, 372–378. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y.; Yesuf, J.; Trushenski, J.; Blackburn, J.W. Use of sweet sorghum juice for lipid production by Schizochytrium limacinum SR21. Bioresour. Technol. 2010, 101, 3623–3627. [Google Scholar] [CrossRef]
- Lung, Y.-T.; Tan, C.H.; Show, P.L.; Ling, T.C.; Lan, J.C.-W.; Lam, H.L.; Chang, J.-S. Docosahexaenoic acid production from crude glycerol by Schizochytrium limacinum SR21. Clean Technol. Environ. Policy 2016, 18, 2209–2216. [Google Scholar] [CrossRef]
- Lee Chang, K.J.; Paul, H.; Nichols, P.D.; Koutoulis, A.; Blackburn, S.I. Australian thraustochytrids: Potential production of dietary long-chain omega-3 oils using crude glycerol. J. Funct. Foods 2015, 19, 810–820. [Google Scholar] [CrossRef]
- Abad, S.; Turon, X. Biotechnological Production of Docosahexaenoic Acid Using Aurantiochytrium limacinum: Carbon Sources Comparison And Growth Characterization. Mar. Drugs 2015, 13, 7275–7284. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-K.; Tian, Y.-T.; Dai, Y.-R.; Wang, D.; Liu, K.-C.; Cui, Y.-H. Development of an alternative medium via completely replaces the medium components by mixed wastewater and crude glycerol for efficient production of docosahexaenoic acid by Schizochytrium sp. Chemosphere 2022, 291, 132868. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.W.; Zhu, S.Y.; Guo, D.S.; Ren, L.J.; Ji, X.J.; Huang, H.; Gao, Z. Development of a strategy for the production of docosahexaenoic acid by Schizochytrium sp. from cane molasses and algae-residue. Bioresour. Technol. 2019, 271, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Abraham, R.E.; Barrow, C.J.; Puri, M. Omega-3 fatty acid production from enzyme saccharified hemp hydrolysate using a novel marine thraustochytrid strain. Bioresour. Technol. 2015, 184, 373–378. [Google Scholar] [CrossRef]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Simultaneous production of DHA and squalene from Aurantiochytrium sp. grown on forest biomass hydrolysates. Biotechnol. Biofuels 2019, 12, 255. [Google Scholar] [CrossRef]
- Patel, A.; Liefeldt, S.; Rova, U.; Christakopoulos, P.; Matsakas, L. Co-production of DHA and squalene by thraustochytrid from forest biomass. Sci. Rep. 2020, 10, 1992. [Google Scholar] [CrossRef] [Green Version]
- Thompson, T.M.; Young, B.R.; Baroutian, S. Advances in the pretreatment of brown macroalgae for biogas production. Fuel Process. Technol. 2019, 195, 106151. [Google Scholar] [CrossRef]
- Syrpas, M.; Venskutonis, P.R. Chapter 6—Algae for the Production of Bio-Based Products. In Biobased Products and Industries; Galanakis, C.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 203–243. [Google Scholar]
- Chen, H.; Zhou, D.; Luo, G.; Zhang, S.; Chen, J. Macroalgae for biofuels production: Progress and perspectives. Renew. Sustain. Energy Rev. 2015, 47, 427–437. [Google Scholar] [CrossRef]
- Dussan, K.; Dijkstra, J.W.; Luzzi, S.; van Zandvoort, I.; van Hal, J.W. Seaweed versatility for biorefinery: Blessing or burden? Curr. Opin. Green Sustain. Chem. 2023, 39, 100728. [Google Scholar] [CrossRef]
- Pardilhó, S.; Cotas, J.; Pacheco, D.; Gonçalves, A.M.M.; Bahcevandziev, K.; Pereira, L.; Figueirinha, A.; Dias, J.M. Valorisation of marine macroalgae waste using a cascade biorefinery approach: Exploratory study. J. Clean. Prod. 2023, 385, 135672. [Google Scholar] [CrossRef]
- Abeln, F.; Fan, J.; Budarin, V.L.; Briers, H.; Parsons, S.; Allen, M.J.; Henk, D.A.; Clark, J.; Chuck, C.J. Lipid production through the single-step microwave hydrolysis of macroalgae using the oleaginous yeast Metschnikowia pulcherrima. Algal Res. 2019, 38, 101411. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wang, F.; Li, C.; Qin, S.; Zhou, M.; Ding, L.; Pang, S.; Duan, D.; Wang, G.; Yin, B.; et al. Emerging challenges: Massive green algae blooms in the Yellow Sea. Nat. Preced. 2008. [Google Scholar] [CrossRef]
- Ning, L.; Yao, Z.; Zhu, B. Ulva (Enteromorpha) Polysaccharides and Oligosaccharides: A Potential Functional Food Source from Green-Tide-Forming Macroalgae. Mar. Drugs 2022, 20, 202. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Wan, X.; Wang, D.; Zhao, C.; Liu, D.; Gao, L.; Wang, M.; Wu, C.; Nabavid, S.M.; Daglia, M.; et al. Polysaccharides from Marine Enteromorpha: Structure and function. Trends Food Sci. Technol. 2020, 99, 11–20. [Google Scholar] [CrossRef]
- Su, H.-Y.; Li, J.-M. Attribute non-attendance in choice experiments: A study of residents’ willingness-to-pay for the disposal of Enteromorpha prolifera in Qingdao, China. Ocean. Coast. Manag. 2020, 191, 105184. [Google Scholar] [CrossRef]
- Ren, C.G.; Liu, Z.Y.; Zhong, Z.H.; Wang, X.L.; Qin, S. Integrated biotechnology to mitigate green tides. Environ. Pollut. 2022, 309, 119764. [Google Scholar] [CrossRef]
- Chattopadhyay, K.; Mandal, P.; Lerouge, P.; Driouich, A.; Ghosal, P.; Ray, B. Sulphated polysaccharides from Indian samples of Enteromorpha compressa (Ulvales, Chlorophyta): Isolation and structural features. Food Chem. 2007, 104, 928–935. [Google Scholar] [CrossRef]
- Feng, D.; Liu, H.; Li, F.; Jiang, P.; Qin, S. Optimization of dilute acid hydrolysis of Enteromorpha. Chin. J. Oceanol. Limnol. 2011, 29, 1243. [Google Scholar] [CrossRef]
- Ray, B. Polysaccharides from Enteromorpha compressa: Isolation, purification and structural features. Carbohydr. Polym. 2006, 66, 408–416. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, X.; Zhao, M.; Yu, S.; Qi, H. The immunological and antioxidant activities of polysaccharides extracted from Enteromorpha linza. Int. J. Biol. Macromol. 2013, 57, 45–49. [Google Scholar] [CrossRef]
- Yang, C.; Huang, S.; Lin, Z.; Chen, H.; Xu, C.; Lin, Y.; Sun, H.; Huang, F.; Lin, D.; Guo, F. Polysaccharides from Enteromorpha prolifera alleviate hypercholesterolemia via modulating the gut microbiota and bile acid metabolism. Food Funct. 2022, 13, 12194–12207. [Google Scholar] [CrossRef]
- Nagula, K.; Sati, H.; Trivedi, N.; Reddy, C.R.K. Chapter 17—Biofuels and Bioproducts from Seaweeds. In Advanced Biofuel Technologies; Tuli, D., Kasture, S., Kuila, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 431–455. [Google Scholar] [CrossRef]
- Suganya, T.; Nagendra Gandhi, N.; Renganathan, S. Production of algal biodiesel from marine macroalgae Enteromorpha compressa by two step process: Optimization and kinetic study. Bioresour. Technol. 2013, 128, 392–400. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Priya, A.K.; Dutta, K.; Rajendran, S.; Vasseghian, Y. Engineering strategies and opportunities of next generation biofuel from microalgae: A perspective review on the potential bioenergy feedstock. Fuel 2022, 312, 122827. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, X.; Jiang, X.; Lin, J.; Lin, X.; Zhao, S.; Yang, J. Valorisation of harmful algae bloom (Enteromorpha prolifera) for polysaccharide and crude bio-oil production. Fuel 2022, 324, 124482. [Google Scholar] [CrossRef]
- Wen, Y.; Xue, C.; Ji, D.; Hou, Y.; Li, K.; Li, Y. Eco-friendly Enteromorpha polysaccharides-based hydrogels for heavy metal adsorption: From waste to efficient materials. Colloids Surf. A Physicochem. Eng. Asp. 2023, 656, 130531. [Google Scholar] [CrossRef]
- Wang, Z.; Song, S.; Wang, H.; Yang, W.; Han, J.; Chen, H. Feasibility of Remediation of Heavy-Metal-Contaminated Marine Dredged Sediments by Active Capping with Enteromorpha Biochar. Int. J. Environ. Res. Public Health 2022, 19, 4944. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, Y.; Peng, T.; Zhong, M.; Hu, Z. Enhanced Fermentable Sugar Production from Enteromorpha Polysaccharides by the Crude Enzymes of Vibrio sp. H11. Microb. Physiol. 2019, 29, 66–73. [Google Scholar] [CrossRef]
- Kim, D.-H.; Lee, S.-B.; Jeong, G.-T. Production of reducing sugar from Enteromorpha intestinalis by hydrothermal and enzymatic hydrolysis. Bioresour. Technol. 2014, 161, 348–353. [Google Scholar] [CrossRef]
- Agabo-García, C.; Romero-García, L.I.; Álvarez-Gallego, C.J.; Blandino, A. Valorisation of the invasive alga Rugulopteryx okamurae through the production of monomeric sugars. Appl. Microbiol. Biotechnol. 2023, 107, 1971–1982. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Gustafsson, L.; Niklasson, C.; Lidén, G. Inhibition effects of furfural on aerobic batch cultivation of Saccharomyces cerevisiae growing on ethanol and/or acetic acid. J. Biosci. Bioeng. 2000, 90, 374–380. [Google Scholar] [CrossRef]
- Yan, B.; Chen, Z.S.; Hu, Y.; Yong, Q. Insight in the Recent Application of Polyphenols From Biomass. Front. Bioeng. Biotechnol. 2021, 9, 753898. [Google Scholar] [CrossRef]
- Sen, B. Determination of Factors Affecting the Enzymatic Hydrolysis of Low Severity Acid-steam Pretreated Agro-residue. J. Chin. Chem. Soc. 2014, 61, 809–813. [Google Scholar] [CrossRef]
- Almeida, J.R.; Bertilsson, M.; Gorwa-Grauslund, M.F.; Gorsich, S.; Lidén, G. Metabolic effects of furaldehydes and impacts on biotechnological processes. Appl. Microbiol. Biotechnol. 2009, 82, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.; Allen, M.J.; Abeln, F.; McManus, M.; Chuck, C.J. Sustainability and life cycle assessment (LCA) of macroalgae-derived single cell oils. J. Clean. Prod. 2019, 232, 1272–1281. [Google Scholar] [CrossRef]
- Chi, G.; Xu, Y.; Cao, X.; Li, Z.; Cao, M.; Chisti, Y.; He, N. Production of polyunsaturated fatty acids by Schizochytrium (Aurantiochytrium) spp. Biotechnol. Adv. 2022, 55, 107897. [Google Scholar] [CrossRef]
- Pérez-Alva, A.; MacIntosh, A.J.; Baigts-Allende, D.K.; García-Torres, R.; Ramírez-Rodrigues, M.M. Fermentation of algae to enhance their bioactive activity: A review. Algal Res. 2022, 64, 102684. [Google Scholar] [CrossRef]
- Chi, Z.; Pyle, D.; Wen, Z.; Frear, C.; Chen, S. A laboratory study of producing docosahexaenoic acid from biodiesel-waste glycerol by microalgal fermentation. Process Biochem. 2007, 42, 1537–1545. [Google Scholar] [CrossRef]
- Huang, T.Y.; Lu, W.C.; Chu, I.M. A fermentation strategy for producing docosahexaenoic acid in Aurantiochytrium limacinum SR21 and increasing C22:6 proportions in total fatty acid. Bioresour. Technol. 2012, 123, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Saltarelli, R.; Ceccaroli, P.; Polidori, E.; Citterio, B.; Vallorani, L.; Stocchi, V. A high concentration of glucose inhibits Tuber borchii mycelium growth: A biochemical investigation. Mycol. Res. 2003, 107, 72–76. [Google Scholar] [CrossRef]
- Hong, W.K.; Yu, A.; Heo, S.Y.; Oh, B.R.; Kim, C.H.; Sohn, J.H.; Yang, J.W.; Kondo, A.; Seo, J.W. Production of lipids containing high levels of docosahexaenoic acid from empty palm fruit bunches by Aurantiochytrium sp. KRS101. Bioprocess Biosyst. Eng. 2013, 36, 959–963. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, H.; Xie, Y.; He, Y.; Sen, B.; Wang, G. Culturable Diversity and Lipid Production Profile of Labyrinthulomycete Protists Isolated from Coastal Mangrove Habitats of China. Mar. Drugs 2019, 17, 268. [Google Scholar] [CrossRef] [Green Version]
- Byreddy, A.R. Thraustochytrids as an alternative source of omega-3 fatty acids, carotenoids and enzymes. Lipid Technol. 2016, 28, 68–70. [Google Scholar] [CrossRef]
- Saini, R.K.; Prasad, P.; Sreedhar, R.V.; Akhilender Naidu, K.; Shang, X.; Keum, Y.S. Omega-3 Polyunsaturated Fatty Acids (PUFAs): Emerging Plant and Microbial Sources, Oxidative Stability, Bioavailability, and Health Benefits—A Review. Antioxidants 2021, 10, 1627. [Google Scholar] [CrossRef]
- Shakeri, S.; Amoozyan, N.; Fekrat, F.; Maleki, M. Antigastric Cancer Bioactive Aurantiochytrium Oil Rich in Docosahexaenoic Acid: From Media Optimization to Cancer Cells Cytotoxicity Assessment. J. Food Sci. 2017, 82, 2706–2718. [Google Scholar] [CrossRef] [PubMed]
- Sikka, P.; Behl, T.; Sharma, S.; Sehgal, A.; Bhatia, S.; Al-Harrasi, A.; Singh, S.; Sharma, N.; Aleya, L. Exploring the therapeutic potential of omega-3 fatty acids in depression. Environ. Sci. Pollut. Res. 2021, 28, 43021–43034. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Ravishankar, G.A.; Keum, Y.S. Microalgae and Thraustochytrids are Sustainable Sources of Vegan EPA and DHA with Commercial Feasibility. Indian J. Microbiol. 2023. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable diversity and biochemical features of thraustochytrids from coastal waters of Southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Foundation: Vienna, Austria, 2020. [Google Scholar]
- Tjørve, K.M.C.; Tjørve, E. The use of Gompertz models in growth analyses, and new Gompertz-model approach: An addition to the Unified-Richards family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef]



| Dependent Variable | a (g/L) | R (gL−1d−1) | λ (d) | Residual Standard Error | |
|---|---|---|---|---|---|
| DCW | EH medium | 3.635 *** | 6.201 * | 0.307 ** | 0.251 |
| Glucose medium | 5.469 *** | 4.183 *** | 0.026 | 0.172 | |
| TFA | EH medium | 0.628 *** | 0.287 *** | −0.369 | 0.034 |
| Glucose medium | 1.092 *** | 0.882 *** | 0.028 | 0.039 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Zhu, X.; Ning, Y.; Chen, X.; Sen, B.; Wang, G. Saturated and Polyunsaturated Fatty Acids Production by Aurantiochytrium limacinum PKU#Mn4 on Enteromorpha Hydrolysate. Mar. Drugs 2023, 21, 198. https://doi.org/10.3390/md21040198
He Y, Zhu X, Ning Y, Chen X, Sen B, Wang G. Saturated and Polyunsaturated Fatty Acids Production by Aurantiochytrium limacinum PKU#Mn4 on Enteromorpha Hydrolysate. Marine Drugs. 2023; 21(4):198. https://doi.org/10.3390/md21040198
Chicago/Turabian StyleHe, Yaodong, Xingyu Zhu, Yaodong Ning, Xiaohong Chen, Biswarup Sen, and Guangyi Wang. 2023. "Saturated and Polyunsaturated Fatty Acids Production by Aurantiochytrium limacinum PKU#Mn4 on Enteromorpha Hydrolysate" Marine Drugs 21, no. 4: 198. https://doi.org/10.3390/md21040198
APA StyleHe, Y., Zhu, X., Ning, Y., Chen, X., Sen, B., & Wang, G. (2023). Saturated and Polyunsaturated Fatty Acids Production by Aurantiochytrium limacinum PKU#Mn4 on Enteromorpha Hydrolysate. Marine Drugs, 21(4), 198. https://doi.org/10.3390/md21040198