

Comparing the Ability of Secretory Signal Peptides for Heterologous Expression of Anti-Lipopolysaccharide Factor 3 in Chlamydomonas reinhardtii


Abstract
:1. Introduction
2. Results
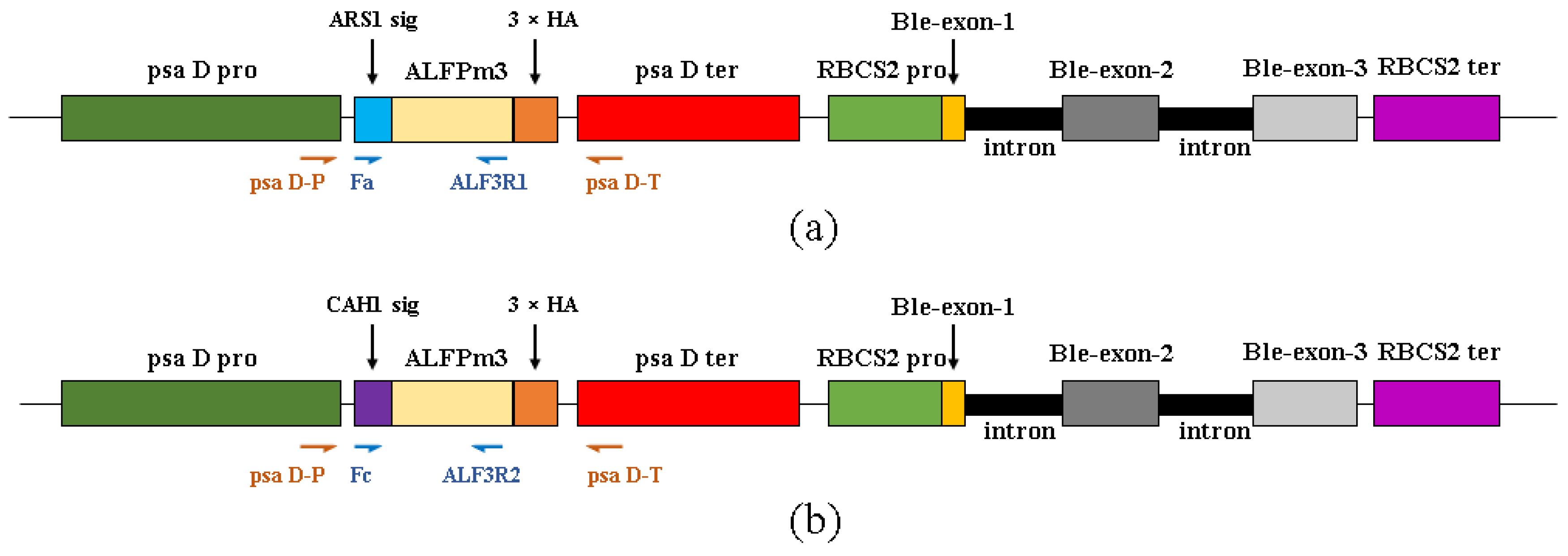
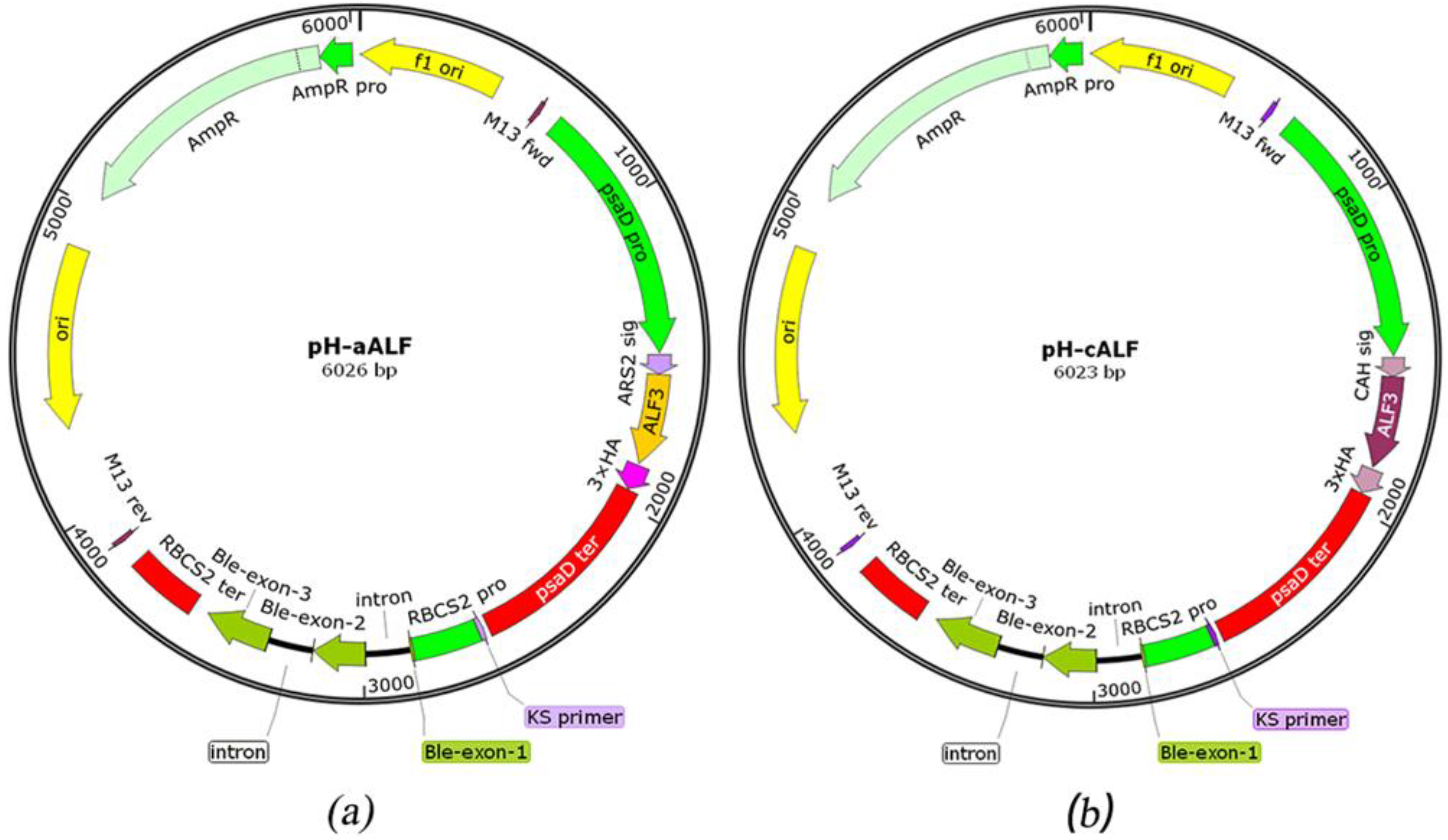
2.1. Design of ALFPm3 Expression Cassette
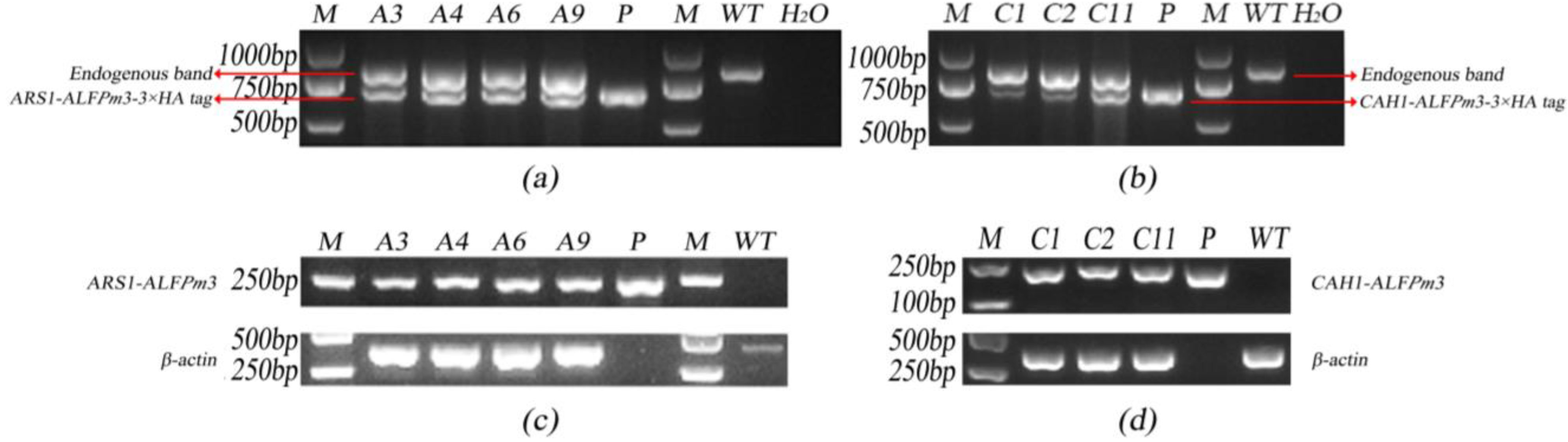
2.2. Screening of Transgenic C. reinhardtii
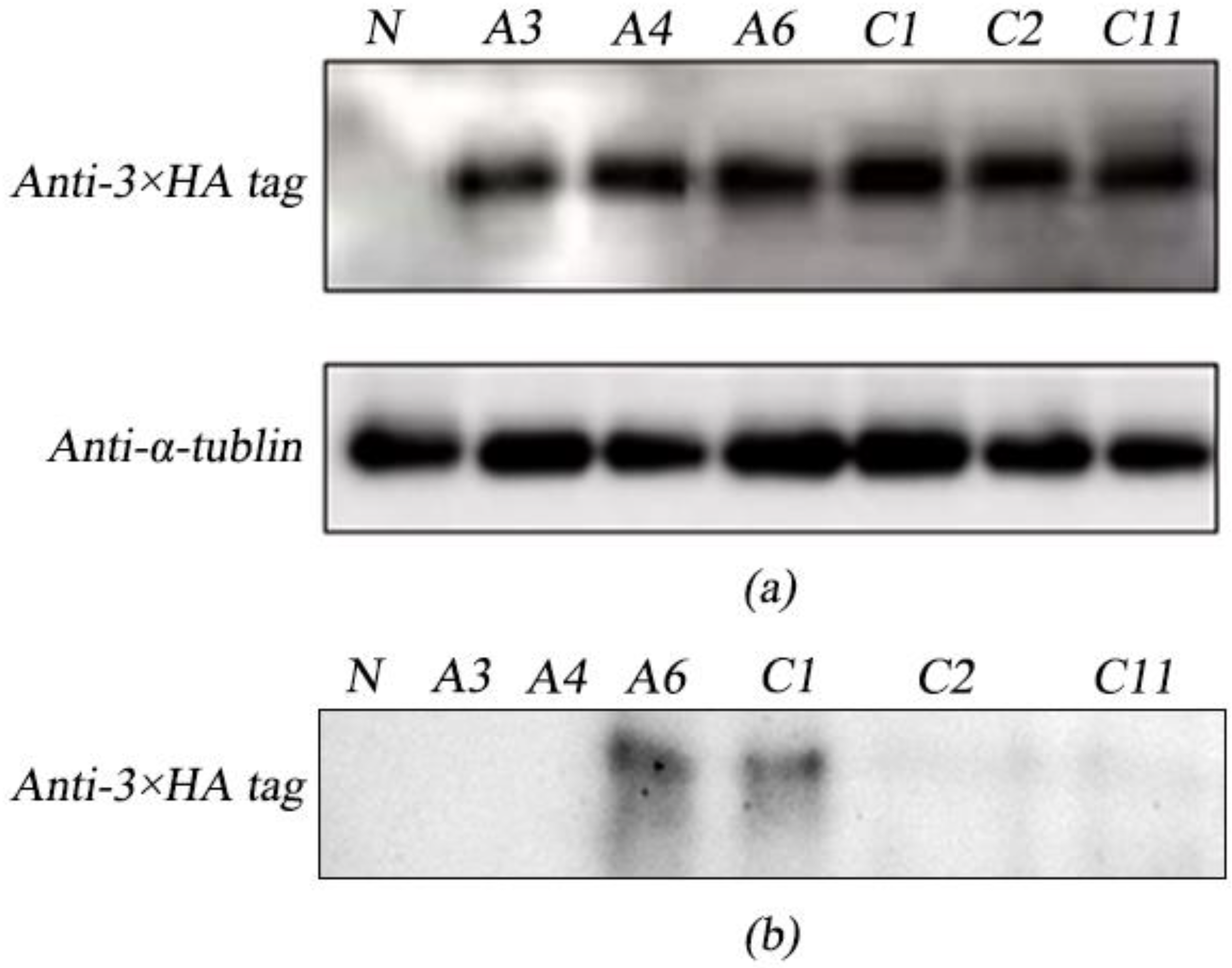
2.3. Analysis of Intracellular and Extracellular Expression of ALFPm3 in Transgenic
C. reinhardtii
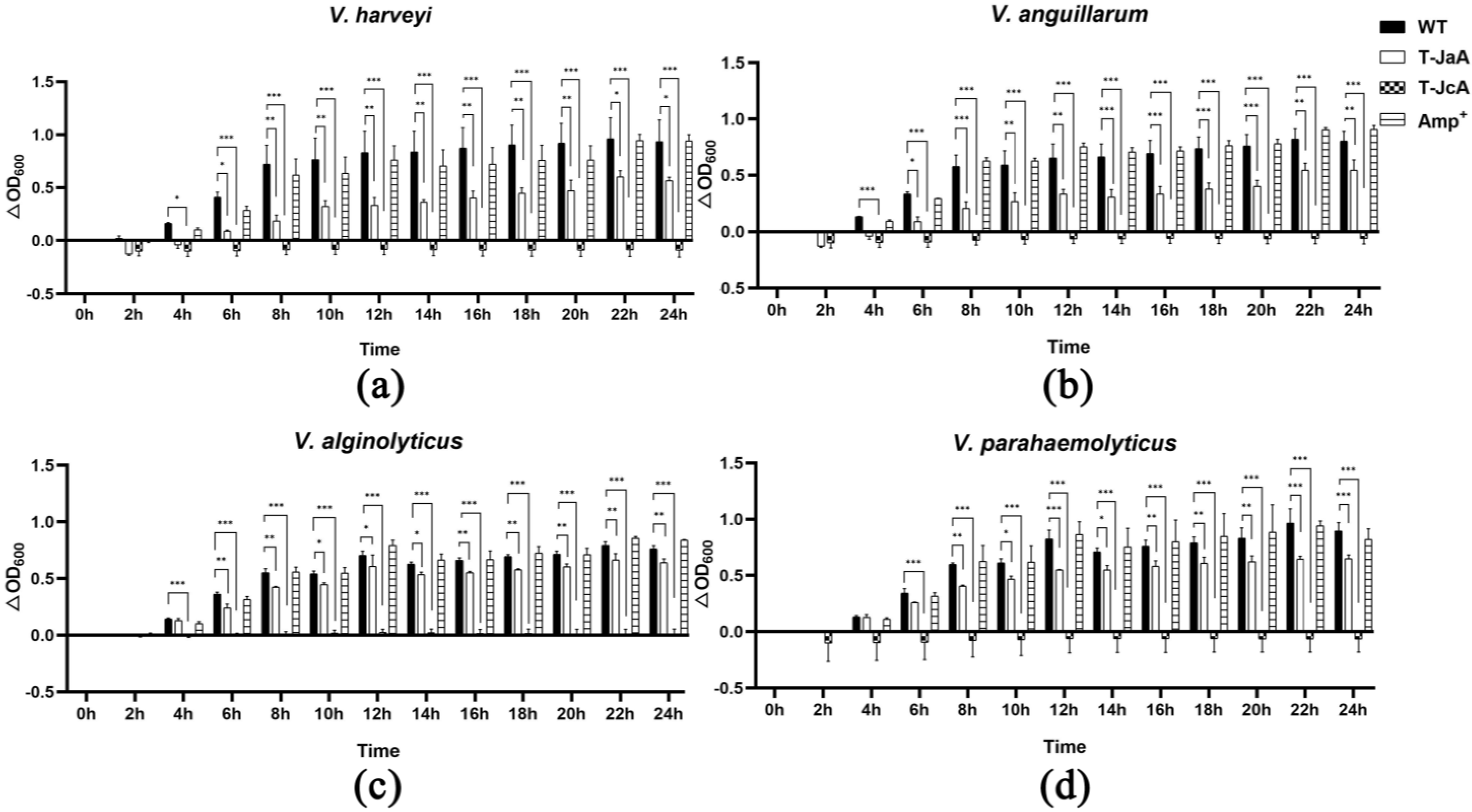
2.4. Both a-ALFPm3 and c-ALFPm3 Showed High Antibacterial Activity
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Algal Strain, and Culture Conditions
4.2. Plasmid Construction and Genetic Transformation
4.3. Genomic PCR and RT-PCR Analysis
4.4. Protein Extraction and Immunoblot Analysis
4.5. Antibacterial Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bombaywala, S.; Mandpe, A.; Paliya, S.; Kumar, S. Antibiotic resistance in the environment: A critical insight on its occurrence, fate, and eco-toxicity. Environ. Sci. Pollut. Res. 2021, 28, 24889–24916. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C.; Godfrey, H.P.; Buschmann, A.H.; Dölz, H.J. Aquaculture as yet another environmental gateway to the development and globalisation of antimicrobial resistance. Lancet Infect. Dis. 2016, 16, e127–e133. [Google Scholar] [CrossRef]
- Boman, H.G.; Faye, I.; Gan, R.; Gudmundsson, G.H.; Lidholm, D.A.; Lee, J.Y.; Xanthopoulos, K.G. Insect immunity: A gene system for antibacterial proteins. Memórias Inst. Oswaldo Cruz 1987, 82, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E.; Brown, K.L.; Mookherjee, N. Host defence peptides from invertebrates—Emerging antimicrobial strategies. Immunobiology 2006, 211, 315–322. [Google Scholar] [CrossRef]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Tanaka, S.; Nakamura, T.; Morita, T.; Iwanaga, S. Limulus anti-LPS factor: An anticoagulant which inhibits the endotoxin-mediated activation of Limulus coagulation system. Biochem. Biophys. Res. Commun. 1982, 105, 717–723. [Google Scholar] [CrossRef]
- Jaree, P.; Tassanakajon, A.; Somboonwiwat, K. Effect of the anti-lipopolysaccharide factor isoform 3 (ALFPm3) from Penaeus monodon on Vibrio harveyi cells. Dev. Comp. Immunol. 2012, 38, 554–560. [Google Scholar] [CrossRef]
- Li, A.; Huang, R.; Wang, C.; Hu, Q.; Li, H.; Li, X. Expression of Anti-Lipopolysaccharide Factor Isoform 3 in Chlamydomonas reinhardtii Showing High Antimicrobial Activity. Mar. Drugs 2021, 19, 239. [Google Scholar] [CrossRef]
- Suraprasit, S.; Methatham, T.; Jaree, P.; Phiwsaiya, K.; Senapin, S.; Hirono, I.; Lo, C.F.; Tassanakajon, A.; Somboonwiwat, K. Anti-lipopolysaccharide factor isoform 3 from Penaeus monodon (ALFPm3) exhibits antiviral activity by interacting with WSSV structural proteins. Antivir. Res. 2014, 110, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Somboonwiwat, K.; Bachère, E.; Rimphanitchayakit, V.; Tassanakajon, A. Localization of anti-lipopolysaccharide factor (ALFPm3) in tissues of the black tiger shrimp, Penaeus monodon, and characterization of its binding properties. Dev. Comp. Immunol. 2008, 32, 1170–1176. [Google Scholar] [CrossRef]
- Puxbaum, V.; Mattanovich, D.; Gasser, B. Quo vadis? The challenges of recombinant protein folding and secretion in Pichia pastoris. Appl. Microbiol. Biotechnol. 2015, 99, 2925–2938. [Google Scholar] [CrossRef] [PubMed]
- Rabert, C.; Weinacker, D.; Pessoa, A., Jr.; Farías, J.G. Recombinants proteins for industrial uses: Utilization of Pichia pastoris expression system. Braz. J. Microbiol. 2013, 44, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Specht, E.; Miyake-Stoner, S.; Mayfield, S. Micro-algae come of age as a platform for recombinant protein production. Biotechnol. Lett. 2010, 32, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Miralles, N.; Saccardo, P.; Corchero, J.L.; Garcia-Fruitós, E. Recombinant Protein Production and Purification of Insoluble Proteins. Methods Mol. Biol. 2022, 2406, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, J.; Adibkia, K. Towards a new avenue for producing therapeutic proteins: Microalgae as a tempting green biofactory. Biotechnol. Adv. 2020, 40, 107499. [Google Scholar] [CrossRef]
- Yang, B.; Liu, J. Chlorella species as hosts for genetic engineering and expression of heterologous proteins: Progress, challenge and perspective. Biotechnol. J. 2016, 11, 1244–1261. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, F. Biology and Industrial Applications of Chlorella: Advances and Prospects. Adv. Biochem. Eng. Biotechnol. 2016, 153, 1–35. [Google Scholar] [CrossRef]
- Lumbreras, V.; Stevens, D.R. Efficient foreign gene expression in Chlamydomonas reinhardtii mediated by an endogenous intron. Plant J. 1998, 14, 441–447. [Google Scholar] [CrossRef]
- Scaife, M.A.; Nguyen, G.T.D.T. Establishing Chlamydomonas reinhardtii as an industrial biotechnology host. Plant J. 2015, 82, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Govea-Alonso, D.O.; Malla, A. An Algae-Made RBD from SARS-CoV-2 Is Immunogenic in Mice. Pharmaceuticals 2022, 15, 1298. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The Potential of Algal Biotechnology to Produce Antiviral Compounds and Biopharmaceuticals. Molecules 2020, 25, 4049. [Google Scholar] [CrossRef]
- El-Ayouty, Y.; El-Manawy, I.; Nasih, S.; Hamdy, E.; Kebeish, R. Engineering Chlamydomonas reinhardtii for Expression of Functionally Active Human Interferon-α. Mol. Biotechnol. 2019, 61, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, J.; Adibkia, K.; Movafeghi, A.; Pourseif, M.M.; Omidi, Y. Designing a new generation of expression toolkits for engineering of green microalgae; robust production of human interleukin-2. Bioimpacts 2020, 10, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.; Ho, S.V.; Farid, S.S. Fed-batch and perfusion culture processes: Economic, environmental, and operational feasibility under uncertainty. Biotechnol. Bioeng. 2013, 110, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Tyo, K.E.J.; Liu, Z.; Magnusson, Y.; Petranovic, D.; Nielsen, J. Impact of protein uptake and degradation on recombinant protein secretion in yeast. Appl. Microbiol. Biotechnol. 2014, 98, 7149–7159. [Google Scholar] [CrossRef]
- Rasala, B.A.; Lee, P.A.; Shen, Z.; Briggs, S.P.; Mendez, M.; Mayfield, S.P. Robust Expression and Secretion of Xylanase1 in Chlamydomonas reinhardtii by Fusion to a Selection Gene and Processing with the FMDV 2A Peptide. PLoS ONE 2012, 7, e43349. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Martinez, E.M.; Fimognari, L.; Sakuragi, Y. High-yield secretion of recombinant proteins from the microalga Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1214–1224. [Google Scholar] [CrossRef] [Green Version]
- Molino, J.V.D.; de Carvalho, J.C.M. Comparison of secretory signal peptides for heterologous protein expression in microalgae: Expanding the secretion portfolio for Chlamydomonas reinhardtii. PLoS ONE 2018, 13, e0192433. [Google Scholar] [CrossRef]
- Kober, L.; Zehe, C.; Bode, J. Optimized signal peptides for the development of high expressing CHO cell lines. Biotechnol. Bioeng. 2013, 110, 1164–1173. [Google Scholar] [CrossRef]
- Lauersen, K.J.; Berger, H.; Mussgnug, J.H.; Kruse, O. Efficient recombinant protein production and secretion from nuclear transgenes in Chlamydomonas reinhardtii. J. Biotechnol. 2013, 167, 101–110. [Google Scholar] [CrossRef]
- Kumar, A.; Falcao, V.R.; Sayre, R.T. Evaluating nuclear transgene expression systems in Chlamydomonas reinhardtii. Algal Res. 2013, 2, 321–332. [Google Scholar] [CrossRef]
- Fischer, N.; Rochaix, J.-D. The flanking regions of PsaD drive efficient gene expression in the nucleus of the green alga Chlamydomonas reinhardtii. Mol. Genet. Genom. 2001, 265, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Baier, T.; Wichmann, J.; Kruse, O.; Lauersen, K. Intron-containing algal transgenes mediate efficient recombinant gene expression in the green microalga Chlamydomonas reinhardtii. Nucleic Acids Res. 2018, 46, 6909–6919. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Chung, C.-P.; Lin, C.-Y.; Sung, H.-H. Function of an anti-lipopolysaccharide factor (ALF) isoform isolated from the hemocytes of the giant freshwater prawn Macrobrachium rosenbergii in protecting against bacterial infection. J. Invertebr. Pathol. 2014, 116, 1–7. [Google Scholar] [CrossRef]
- Ponprateep, S.; Somboonwiwat, K.; Tassanakajon, A. Recombinant anti-lipopolysaccharide factor isoform 3 and the prevention of vibriosis in the black tiger shrimp, Penaeus monodon. Aquaculture 2009, 289, 219–224. [Google Scholar] [CrossRef]
- Anju, M.V.; Archana, K.; Anooja, V.V.; Athira, P.P.; Neelima, S.; Singh, I.S.B.; Philip, R. A novel anti-lipopolysaccharide factor from blue swimmer crab Portunus pelagicus and its cytotoxic effect on the prokaryotic expression host, E. coli on heterologous expression. J. Genet. Eng. Biotechnol. 2023, 21, 22. [Google Scholar] [CrossRef]
- Yang, H.; Li, S.; Li, F.; Yu, K.; Yang, F.; Xiang, J. Recombinant Expression of a Modified Shrimp Anti-Lipopolysaccharide Factor Gene in Pichia pastoris GS115 and Its Characteristic Analysis. Mar. Drugs 2016, 14, 152. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Liu, J.; Li, S.; Li, H.; Liu, F. Recombinant expression of an oriental river prawn anti-lipopolysaccharide factor gene in Pichia pastoris and its characteristic analysis. Fish Shellfish. Immunol. 2020, 98, 414–419. [Google Scholar] [CrossRef]
- Meng, D.-M.; Dai, H.-X.; Gao, X.-F.; Zhao, J.-F.; Guo, Y.-J.; Ling, X.; Dong, B.; Zhang, Z.-Q.; Fan, Z.-C. Expression, purification and initial characterization of a novel recombinant antimicrobial peptide Mytichitin—A in Pichia pastoris. Protein Expr. Purif. 2016, 127, 35–43. [Google Scholar] [CrossRef]
- Meng, D.-M.; Zhao, J.-F.; Ling, X.; Dai, H.-X.; Guo, Y.-J.; Gao, X.-F.; Dong, B.; Zhang, Z.-Q.; Meng, X.; Fan, Z.-C. Recombinant expression, purification and antimicrobial activity of a novel antimicrobial peptide PaDef in Pichia pastoris. Protein Expr. Purif. 2017, 130, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Somboonwiwat, K.; Marcos, M.; Tassanakajon, A.; Klinbunga, S.; Aumelas, A.; Romestand, B.; Gueguen, Y.; Boze, H.; Moulin, G.; Bachère, E. Recombinant expression and anti-microbial activity of anti-lipopolysaccharide factor (ALF) from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2005, 29, 841–851. [Google Scholar] [CrossRef]
- Lai, W.-S.; Kan, S.-C.; Lin, C.-C.; Shieh, C.-J.; Liu, Y.-C. Antibacterial Peptide CecropinB2 Production via Various Host and Construct Systems. Molecules 2016, 21, 103. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, J.-X.; Zhao, X.-F.; Kang, C.-J.; Liu, N.; Xiang, J.-H.; Li, F.-H.; Sueda, S.; Kondo, H. High level expression, purification, and characterization of the shrimp antimicrobial peptide, Ch-penaeidin, in Pichia pastoris. Protein Expr. Purif. 2004, 39, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Rivet, E.; Kiefer-Meyer, M.-C. Protein N-glycosylation in eukaryotic microalgae and its impact on the production of nuclear expressed biopharmaceuticals. Front Plant Sci. 2014, 5, 359. [Google Scholar] [CrossRef] [PubMed]
- Lingg, N.; Zhang, P.; Song, Z.; Bardor, M. The sweet tooth of biopharmaceuticals: Importance of recombinant protein glycosylation analysis. Biotechnol. J. 2012, 7, 1462–1472. [Google Scholar] [CrossRef]
- Gangl, D.; Zedler, J.A.Z.; Rajakumar, P.D.; Martinez, E.M.R.; Riseley, A.; Włodarczyk, A.; Purton, S.; Sakuragi, Y.; Howe, C.J.; Jensen, P.E.; et al. Biotechnological exploitation of microalgae. J. Exp. Bot. 2015, 66, 6975–6990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimpel, J.A.; Hyun, J.S.; Schoepp, N.G.; Mayfield, S.P. Production of recombinant proteins in microalgae at pilot greenhouse scale. Biotechnol. Bioeng. 2015, 112, 339–345. [Google Scholar] [CrossRef]
- Zedler, J.A.Z.; Gangl, D.; Guerra, T.; Santos, E.; Verdelho, V.V.; Robinson, C. Pilot-scale cultivation of wall-deficient transgenic Chlamydomonas reinhardtii strains expressing recombinant proteins in the chloroplast. Appl. Microbiol. Biotechnol. 2016, 100, 7061–7070. [Google Scholar] [CrossRef]
- Murugan, C.; Venkatesan, S.; Kannan, S. Cancer Therapeutic Proficiency of Dual-Targeted Mesoporous Silica Nanocomposite Endorses Combination Drug Delivery. ACS Omega 2017, 2, 7959–7975. [Google Scholar] [CrossRef] [Green Version]
- Lima, S.; Webb, C.L.; Deery, E.; Robinson, C.; Zedler, J.A.Z. Human Intrinsic Factor Expression for Bioavailable Vitamin B12 Enrichment in Microalgae. Biology 2018, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Patena, W.; Armbruster, U.; Gang, S.S.; Blum, S.R.; Jonikas, M.C. High-Throughput Genotyping of Green Algal Mutants Reveals Random Distribution of Mutagenic Insertion Sites and Endonucleolytic Cleavage of Transforming DNA. Plant Cell 2014, 26, 1398–1409. [Google Scholar] [CrossRef] [Green Version]
- Meslet-Cladière, L.; Vallon, O. Novel Shuttle Markers for Nuclear Transformation of the Green Alga Chlamydomonas reinhardtii. Eukaryot. Cell 2011, 10, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, R.M.; Haglund, C.M.; Chin, B.L.; Kobayashi, M.C.; Niyogi, K.K. Functional Genomics of Eukaryotic Photosynthesis Using Insertional Mutagenesis of Chlamydomonas reinhardtii. Plant Physiol. 2005, 137, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, R.; Patena, W.; Gang, S.S.; Blum, S.R.; Ivanova, N.; Yue, R.; Robertson, J.M.; Lefebvre, P.A.; Fitz-Gibbon, S.T.; et al. An Indexed, Mapped Mutant Library Enables Reverse Genetics Studies of Biological Processes in Chlamydomonas reinhardtii. Plant Cell 2016, 28, 367–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, T.; Okamoto, K.; Onai, K.; Niwa, Y.; Shimogawara, K.; Ishiura, M. A systematic forward genetic analysis identified components of the Chlamydomonas circadian system. Genes Dev. 2008, 22, 918–930. [Google Scholar] [CrossRef] [Green Version]
- Cerutti, H.; Johnson, A.M.; Gillham, N.W.; Boynton, J.E. A Eubacterial Gene Conferring Spectinomycin Resistance on Chlamydomonas reinhardtii: Integration into the Nuclear Genome and Gene Expression. Genetics 1997, 145, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, T.; Miyasaka, H.; Ohama, T. Unstable RNAi Effects Through Epigenetic Silencing of an Inverted Repeat Transgene in Chlamydomonas reinhardtii. Genetics 2008, 180, 1927–1944. [Google Scholar] [CrossRef] [Green Version]
- Koblenz, B.; Lechtreck, K.-F. The NIT1 Promoter Allows Inducible and Reversible Silencing of Centrin in Chlamydomonas reinhardtii. Eukaryot. Cell 2005, 4, 1959–1962. [Google Scholar] [CrossRef] [Green Version]
- Eichler-Stahlberg, A.; Weisheit, W.; Ruecker, O.; Heitzer, M. Strategies to facilitate transgene expression in Chlamydomonas reinhardtii. Planta 2009, 229, 873–883. [Google Scholar] [CrossRef]
- Sardis, M.F.; Tsirigotaki, A. Preprotein Conformational Dynamics Drive Bivalent Translocase Docking and Secretion. Structure 2017, 25, 1056–1067.e1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smets, D.; Smit, J.; Xu, Y.; Karamanou, S.; Economou, A. Signal Peptide-rheostat Dynamics Delay Secretory Preprotein Folding. J. Mol. Biol. 2022, 434, 167790. [Google Scholar] [CrossRef]
- Freudl, R. Signal peptides for recombinant protein secretion in bacterial expression systems. Microb. Cell Fact. 2018, 17, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalucki, Y.M.; Gittins, K.L.; Jennings, M.P. Secretory signal sequence non-optimal codons are required for expression and export of β-lactamase. Biochem. Biophys. Res. Commun. 2008, 366, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Zalucki, Y.M.; Jennings, M.P. Experimental confirmation of a key role for non-optimal codons in protein export. Biochem. Biophys. Res. Commun. 2007, 355, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Cai, X.; Liu, Z.; Zheng, Y. High-Throughput Screening of Signal Peptide Library with Novel Fluorescent Probe. Chembiochem 2022, 23, 202100523. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Tian, X. Effects of different signal peptides on the secretion of human-mouse chimeric CMV-IgM. Shengwu Gongcheng Xuebao 2020, 36, 1223. [Google Scholar]
- Neupert, J.; Shao, N.; Lu, Y.; Bock, R. Genetic Transformation of the Model Green Alga Chlamydomonas reinhardtii. Methods Mol. Biol. 2012, 847, 35–47. [Google Scholar] [CrossRef]
- Cao, M.; Fu, Y.; Guo, Y.; Pan, J. Chlamydomonas (Chlorophyceae) colony PCR. Protoplasma 2009, 235, 107–110. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence ( 5′–3′) | Target Gene | Expected Product (bp) |
|---|---|---|---|
| psaD-P | GGGAATTGGAGGTACGACCGAGAT | ARS1-ALFPm3 CAH1-ALFPm3 | 679 676 |
| psaD-T | AGCTCCGATCCCGTATCAATCAGC | ||
| Fa | CGCGCTGGCTGTGTTCG | ARS1-ALFPm3 | 222 |
| ALF3R1 | GTCCAGCCGGGGCACCACATG | ||
| Fc | CGCGCACCGGGGCACTCCTG | CAH1-ALFPm3 | 203 |
| ALF3R2 | ACATGCGGCCCTTGTAGTACACCT | ||
| actin-F | ACCCCGTGCTGCTGACTG | β-actin | 351 |
| actin-R | ACGTTGAAGGTCTCGAACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuang, H.; Ou, Y.; Chen, R.; Huang, D.; Wang, C. Comparing the Ability of Secretory Signal Peptides for Heterologous Expression of Anti-Lipopolysaccharide Factor 3 in Chlamydomonas reinhardtii. Mar. Drugs 2023, 21, 346. https://doi.org/10.3390/md21060346
Zhuang H, Ou Y, Chen R, Huang D, Wang C. Comparing the Ability of Secretory Signal Peptides for Heterologous Expression of Anti-Lipopolysaccharide Factor 3 in Chlamydomonas reinhardtii. Marine Drugs. 2023; 21(6):346. https://doi.org/10.3390/md21060346
Chicago/Turabian StyleZhuang, Huilin, Yaohui Ou, Ruoyu Chen, Danqiong Huang, and Chaogang Wang. 2023. "Comparing the Ability of Secretory Signal Peptides for Heterologous Expression of Anti-Lipopolysaccharide Factor 3 in Chlamydomonas reinhardtii" Marine Drugs 21, no. 6: 346. https://doi.org/10.3390/md21060346
APA StyleZhuang, H., Ou, Y., Chen, R., Huang, D., & Wang, C. (2023). Comparing the Ability of Secretory Signal Peptides for Heterologous Expression of Anti-Lipopolysaccharide Factor 3 in Chlamydomonas reinhardtii. Marine Drugs, 21(6), 346. https://doi.org/10.3390/md21060346