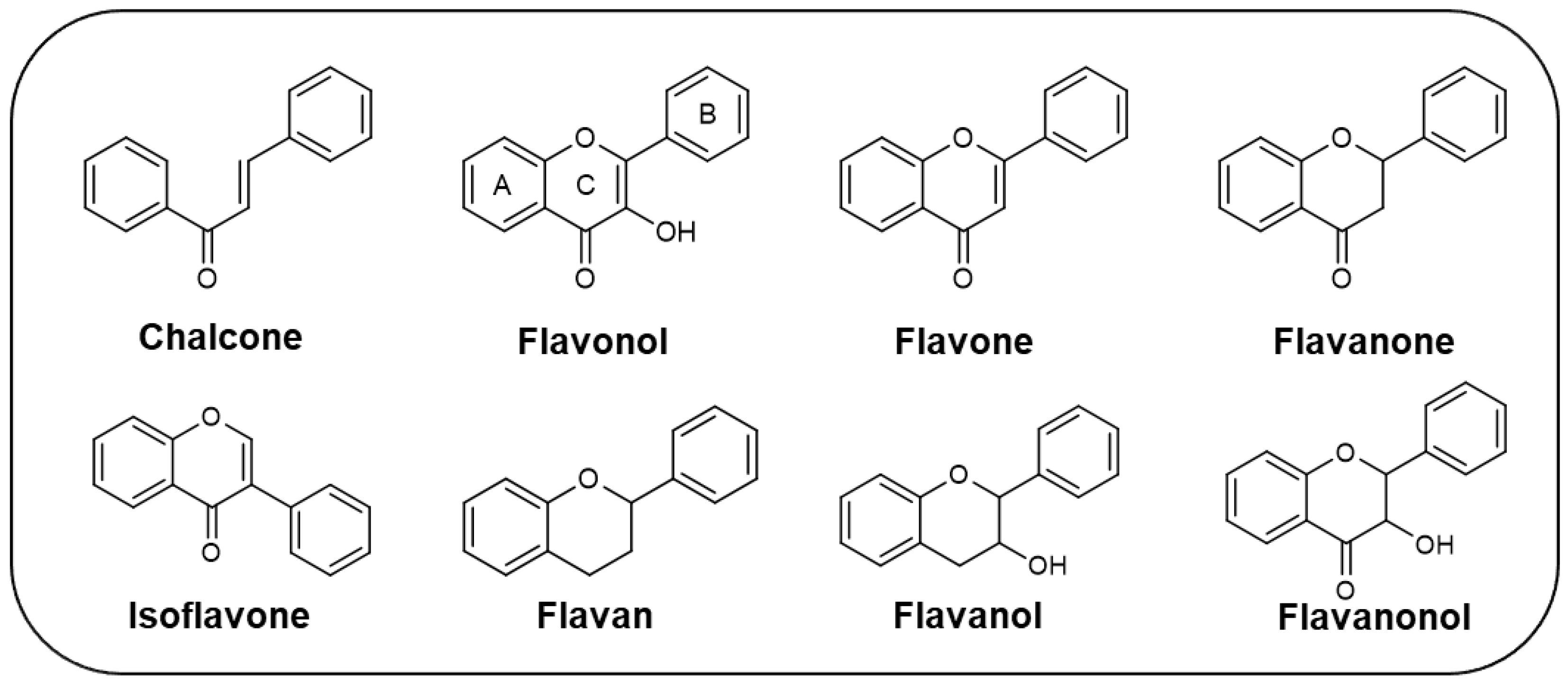
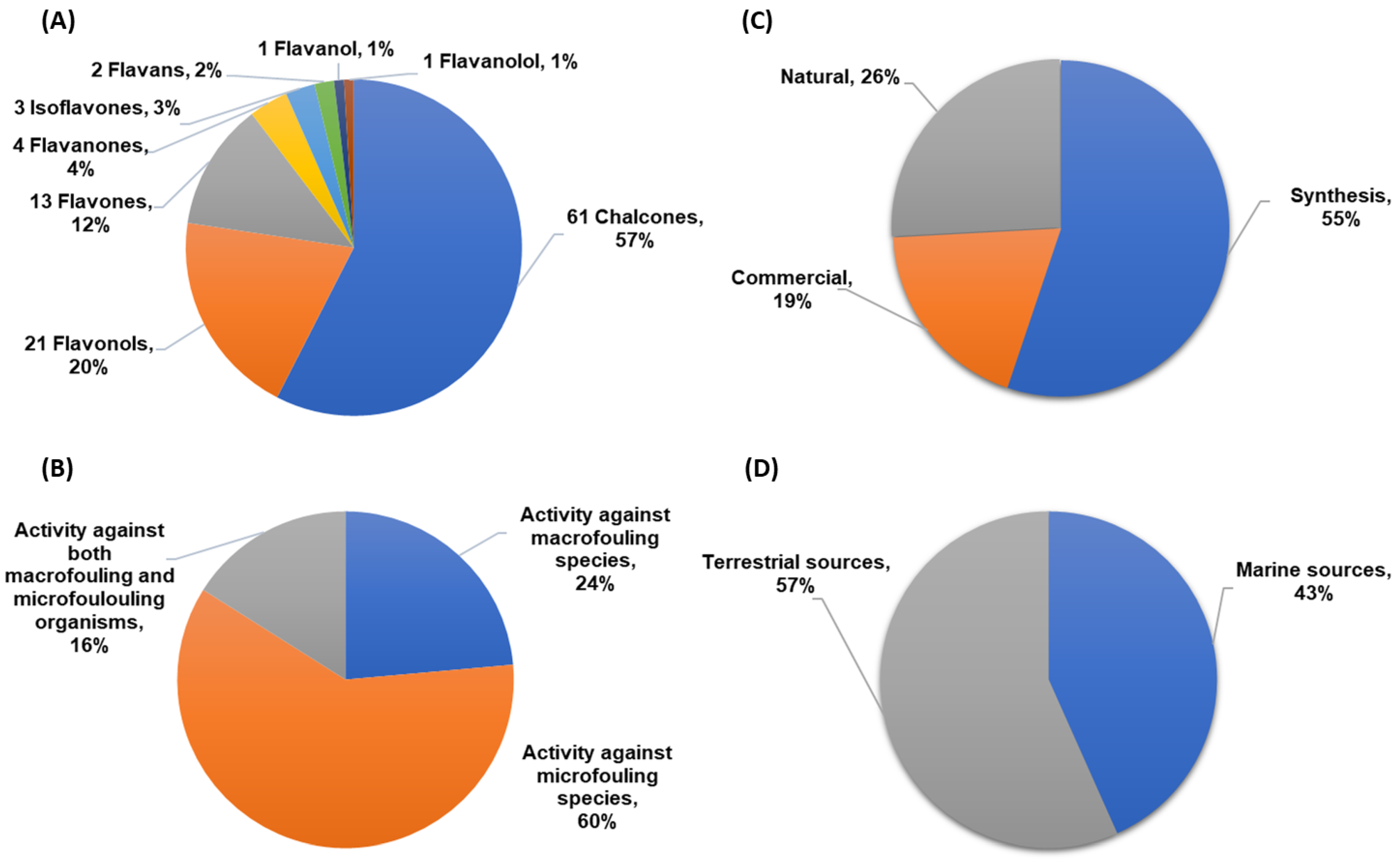
A total of 106 flavonoids with antifouling activity against macrofouling (mussels, barnacles, algae) and microfouling (diatoms, bacteria, protozoans) species have been reported, including mainly chalcones, flavonols, and flavones. Other classes of flavonoids with antifouling potential were also reported but less represented, namely flavanone, isoflavone, flavan, flavanol, and flavanonol derivatives (
Figure 2A). Most of the studies (60%) were performed using microfouling species (namely marine bacteria and diatoms), and only 24% of the compounds were evaluated against macrofouling organisms, mainly mussel and barnacle species. Nonetheless, 16% of the tested compounds were evaluated against both macrofouling and microfouling organisms (
Figure 2B). Considering the origin of the flavonoids reported in this review, as shown in
Figure 2C, most of the compounds were obtained through synthesis in a laboratory (55%), followed by natural flavonoids (26%) and commercial compounds (19%). Among natural flavonoids, the percentage of antifouling flavonoids obtained from marine sources is almost as high as those from terrestrial sources, as shown in
Figure 2D. The structure, origin, and antifouling effects of flavonoids are presented in
Table S1 (Supplementary Material), which is organized in chronological order of the year of the first report of the antifouling activity.
2.1. Chalcones
A total of 61 chalcones with antifouling potential have been reported. Although this class of flavonoids is common in nature, all chalcones reported to have antifouling activity were obtained by chemical synthesis. The structures of chalcones with antifouling potential are shown in
Figure 3.
Sivakumar and colleagues evaluated the antifouling activity of 47 synthetic chalcone derivatives (
1–
47) against three marine bacteria (
Bacillus flexus,
Pseudomonas fluorescens, and
Vibrio natriegens) obtained from biofilms formed on polymer and metal surfaces immersed in ocean water [
14]. All compounds were shown to be highly active against
B. flexus bacterial growth, with minimum inhibitory concentration (MIC) values between 0.002 and 0.466 µM,
P. fluorescens (MIC values between 0.004 and 0.133 µM), and
V. natriegens (MIC values between 0.024 and 0.249 µM) [
14]. Among tested compounds, chalcones
5,
7,
16,
23, and
36 were the most active against
B. flexus, with MIC values of 0.002–0.031 µM. All these chalcones presented a hydroxylated ring on their A ring, which seems to be an important substituent for their antibacterial activity against
B. flexus. For
V. natriegens, chalcones
31 and
32 presented the highest activity, with MIC values of 0.024 µM. All compounds showed potent activity regarding
P. fluorescens.
The same research group showed that 2-methoxy-2′,4′-dichlorochalcone (
45), mixed with a marine paint and coated on polycarbonate (PC), polymethylmethacrylate (PMMA), and a glass-fiber-reinforced plastic (GFRP) surface, inhibited the marine bacterium
V. natriegens biofilm [
15]. This coated surface was also compared with surfaces coated with marine paint without any additive and the one mixed with copper (1%), and it was found that surfaces coated with the paint containing the 0.423 µM of 2-methoxy-2′,4′-dichlorochalcone were the most effective amongst these three in reducing the amount of biofilm as well as its thickness. The biofilm formation can be attributed to the presence of fimbria structures in the
V. natriegens bacteria, which are used for attachment on surfaces, and these structures are highly enriched with proteins, which is one possible reason for its hydrophobicity. Since the coating with dichlorochalcone
45 increases the hydrophilicity of the polymeric surfaces, it decreases the formation of biofilm, which can explain the activity of this coating. Another assay showed that only about 15% of chalcone
45 was lost from the surface after 28 days, maintaining its activity in a sustained manner [
15].
A series of synthetic chalcone derivatives were evaluated by us to determine their antifouling potential against marine macro- and microfouling organisms, including mussel
M. galloprovincialis larvae, marine bacteria
Vibrio harveyi,
Cobetia marina,
Halomonas aquamarina,
Pseudoalteromonas atlantica, and
Roseobacter litoralis, and marine diatoms
Cylindrotheca sp.,
Halamphora sp.,
Nitzschia sp., and
Navicula sp. [
16]. Among the 16 compounds tested, three methoxylated chalcones,
48,
49, and
50, showed effective anti-settlement activity against the
Mytilus galloprovincialis mussel larvae, with EC
50 values ranging from 7.24 to 34.63 µM. Chalcones
49 and
50 also showed growth-inhibitory activity against the bacterial strains
H. aquamarina (with EC
50 values of 18.67 and 18.78, respectively) and
R. litoralis (with EC
50 values of 4.09 and 12.34 µM, respectively). Moreover, chalcone
50, with a prenyl group, also displayed inhibitory activity against diatoms
Cylindrotheca sp.,
Halamphora sp.,
Nitzschia sp., and
Navicula sp., with EC
50 values ranging from 6.75 to 20.31 µM. An ecotoxicity assay performed against non-target organism
Artemia salina showed that these compounds were not toxic at 50 µM [
16].
Sathicq et al. reported the synthesis and antifouling evaluation of a series of seven chalcones, including the non-substituted chalcone (
4) and six furylchalcones (
51–
56) [
17]. These compounds were incorporated into a rosin-based antifouling coating, and acrylic panels were painted with these paints and further immersed in the sea for 45 days, and then the results were compared with a control paint (without the incorporation of chalcones). Concerning the evaluation of their antifouling activity against macrofouling species, paints containing non-substituted chalcone
4 did not show significant differences compared with the control paint, with only a reduction of approximately 20% in the presence of macrofouling organisms compared to the control paint, whereas furylchalcones
51–
56 displayed a strong anti-macrofouling effect, showing a high reduction in the settlement of algae (filamentous red alga
Polysiphonia sp., green alga
Ulva sp., filamentous brown alga
Ectocarpus sp.) as well as calcareous tubeworms (
Hydroides sp. and
Spirorbis sp.). In fact, paints with furylchalcones
53 and
55 showed an inhibition of almost 100% of the settlement of macrofouling species, and compounds
51,
52,
54, and
56 showed only about 10%–25% of settlement of these organisms compared to the control paint. Regarding their anti-microfouling effect, all paints containing chalcones
4 and
51–
56 were able to inhibit the settlement and growth of diatoms, namely
Achnanthes sp.,
Coscinodiscus sp.,
Grammatophora sp.,
Licmophora sp.,
Navicula sp.,
Nitzschia longissima,
Pinnularia sp.,
Pleurosigma sp.,
Synedra sp., and
Skeletonema costatum, and protozoans such as
Vorticella sp. and
Zoothamnium sp. Compared to the control paint, which was 100% covered with diatoms and protozoans, for paints with unsubstituted chalcone and furylchalcones only from about 20% to 40% of the painted plate was covered with these fouling organisms, with furylchalcone
51 being the most active with less than 20% of the plate being covered with microfouling organisms. SAR studies showed that, in general, compounds
55 and
56, with the presence of a 2′-hydroxy group on their A ring, showed higher activity than compounds
51–
54 in the inhibition of the settlement of macrofouling species. Compounds
53–
54, with a methyl group on their A ring, also showed higher activity than compounds
51–
52 against the settlement of macrofouling species, suggesting that the presence of methyl and hydroxy groups is beneficial to the antimacrofouling activity of this series of compounds [
17].
More recently, our group reported a synthesis and antifouling activity evaluation for a series of eight triazole-linked flavonoid glycosides, as well as intermediate compounds. Among them, chalcone
57, as well as structure-related non-glycosylated chalcones
58 and
59 and the 4′-propargylated chalcones obtained as synthetic intermediates (
60 and
61), displayed antifouling activity against macro- and/or microfouling species [
18]. The hybridization of intermediate chalcone with a glycosyl moiety was performed through the click chemistry reaction, obtaining a 1,2,3-triazole ring as a linker, which has been much highlighted as a promising strategy in medicinal chemistry, giving rise to hybrids with a wide range of biological activities [
19]. Moreover, as the triazole ring was proven to be an effective scaffold for marine biofouling prevention, the use of flavonoids with a triazole ring could result in promising antifouling agents [
20]. To assess their antifouling activity, the compounds were studied concerning their effect on the settlement of the macrofouling species
M. galloprovincialis mussel larvae and on the growth of five biofilm-forming marine bacteria, namely
Vibrio harveyi,
Cobetia marina,
H. aquamarina,
Pseudoalteromonas atlantica, and
R. litoralis. Among the tested compounds, chalcones
57 (EC
50 = 3.28 µM; 2.43 µg/mL),
58 (EC
50 = 18.10 µM; 5.44 µg/mL), and
59 (EC
50 = 9.64 µM; 3.18 µg/mL) displayed effective anti-settlement activity (EC
50 values < 25 µg/mL), with triazolyl glycosylated chalcone
57 being the most potent and having the highest therapeutic ratio (>60.98). Concerning the antimicrofouling activity, only propargylated chalcones
60 and
61 presented some activity towards the growth of
R. litoralis (EC
30 values of 135 and 83.5 µM, respectively). Later, chalcone
57 was shown to also be able to inhibit the biofilm formation of two strains of bacteria
S. aureus [
21]. The compounds showing the highest anti-settlement effect towards the mussel larvae (
57–
59) were further evaluated against the biofilm-forming marine diatom
Navicula sp. Only chalcone
57 was able to display some activity against this microalga’s growth (EC
50 = 41.76 µM; 30.94 µg/mL). An ecotoxicity assay performed against the marine non-target organism
Artemia salina showed no toxicity for the chalcones
57–
59 (less than 10% mortality at 50 μM) [
18]. In this study, it was highlighted that the presence of a 3,4,5-trimethoxy group on the B ring of the chalcone scaffold seems to be beneficial for antifouling activity. Following the promising results obtained for compound
57, the toxicity of compound
57 to humans was preliminarily assessed with an immortalized human retinal pigment epithelial cell line (hTERT-RPE-1) and compared with the commercial biocide Econea®. While the commercial biocide caused a 50% decrease in the cell viability at the lowest concentration tested (1 µM), chalcone
57 was only able to decrease the cell viability by 50% at the highest concentration tested (100 µM), which allows us to hypothesize that chalcone
57 may present a more safe profile [
22]. Overall, the results obtained for triazole-linked chalcone glycoside
57 suggest that the conjugation of flavonoids with a glycosyl moiety through a click chemistry approach might be a potential tool for the development of new antifouling agents.
2.2. Flavonols
Flavonols comprise the second most reported class of flavonoids with an effect in the prevention of marine biofouling. The structures of flavonols with antifouling potential are presented in
Figure 4.
The Kaempferol glucoside (kaempferol 3-
O-(2″,6″-di-
O-(
E)-
p-coumaroyl-β-
d-glucopyranoside,
62), isolated for the first time from the leaves of
Quercus dentata, was the first flavonol reported for antifouling evaluation [
13]. This flavonol showed high repellent activity against the settlement of blue mussel
Mytilus edulis at 0.22 µmol/cm
2, whereas the positive control, copper sulfate, showed similar activity at higher concentration (0.50 µmol/cm
2), indicating a twofold more potent effect of this flavonol than copper sulfate. Authors also compared the activity of compound
62 with kaempferol (
63) (concentration of 6.0 µmol/cm
2 to exert the same activity) and
p-coumaric acid (7.3 µmol/cm
2 to exert the same activity), as well as a mixture of kaempferol and
p-coumaric acid (1:2) (2.2 µmol/cm
2 to exert the same activity), concluding that compound
62 presented higher activity than both kaempferol (
63) and
p-coumaric acid, thus suggesting that the combination of the kaempferol and
p-coumaric acid moieties is associated with an improvement in antifouling activity [
13].
Several flavonol aglycones, including kaempferol (
63), quercetin (
64), myricetin (
65), and glycosides, such as kaempferol 3-
O-rhamnoside (
66), kaempferol 3-
O-glucoside (
67), quercitrin (
68), neoisorutin (
69), as well as kaempferol 3-
O-(2″,6″-di-
O-(
E)-
p-coumaroyl-β-
d-glucopyranoside (
62), previously reported by Yamashita et al. [
13], were tested by Singh and collaborators for their antifouling potential against the settlement of blue mussel
M. edulis, and their activity was compared with the standard antifouling compound copper sulfate [
23]. Although these flavonols were obtained commercially for this study, these compounds are found in nature, namely in terrestrial and marine sources, and are associated with a wide range of biological activities [
9,
24,
25,
26,
27]. Flavonoid
62 presented the highest activity in this study, with a more than twofold higher activity than the positive control copper sulfate. On the other hand, the other flavonoids presented only moderate activity, with 5% to 20% of the activity of copper sulfate. By comparing the activity of the tested flavonols, it appeared that flavonoid glycosides with a rhamnosyl moiety were more active than analogues with glycosyl moiety, and the presence of two hydroxy groups on the B ring allied to the two hydroxy groups on the A ring appeared to enhance the anti-settlement activity [
23].
The antifouling activity of the hydroxylated flavonols kaempferol (
63) and neoisorutin (
69) was also reported by Zhou et al. [
28]. Both compounds showed a promising effect against the larval settlement of the barnacle
Amphibalanus amphitrite, displaying EC
50 values of 33.54 μM (9.6 µg/mL) and 65.68 μM (40.1 µg/mL), respectively. These results reinforce the potential of these natural flavonols in the prevention of the settlement of macrofouling species. In addition to these flavonoids, calycopterin (
70), casticin (
71), 5,4′-dihydroxy-3,6,7-trimethoxyflavone (
72), 3,5-dihydroxy-7,4′-dimethoxyflavone (
73), and icariin (
74) also behave as inhibitors of the larval settlement of the same barnacle [
28] (EC
50 values ranging from 7.26 μM to 57.17 μM, 2.5 µg/mL–35.3 µg/mL), with casticin (
71, EC
50: 8.01 μM, 3.0 µg/mL; LC
50/EC
50: >16.7) and 5,4′-dihydroxy-3,6,7-trimethoxyflavone (
72, EC
50: 7.26 μM, 2.5 µg/mL; LC
50/EC
50: >20.0) being the most active flavonols. All flavonols,
63,
69, and
70–
74, were obtained from terrestrial plants. The analysis of these results suggests that the presence of a hydroxy group at C5 is important for antifouling activity, whereas the presence of a bulky group such as rhamnosyl at C3 may reduce the activity [
28].
In a study performed by Kong et al., the flavonols kaempferol (
63) and quercetin (
64), isolated from the methanol extract of the leaves of the marine halophyte
Apocynum venetum, were tested for their antifouling activity against the marine fouling bacteria
Bacillus thuringiensis,
Pseudoalteromonas elyakovii and
Pseudomonas aeruginosa [
29]. At a concentration of 100 µg/disc, quercetin (
64) (inhibition zone: 9.3–9.5 mm) and kaempferol (
63) (inhibition zone: 9.5–17.4 mm) displayed moderate antibacterial activities against the three tested bacteria, compared to the positive control chloramphenicol at the same concentration (inhibition zone 29.2–33.7 mm) [
29].
The effect of quercetin (
64) on microfouling organisms was also studied by Gopikrishnan et al. [
30]. This natural flavonol, isolated from the estuarine sediment actinobacterium
Streptomyces fradiae PE7, was able to inhibit the growth of 12 bacterial strains isolated from marine fouling samples (
Staphylococcus sp. M1,
Micrococcus sp. M50,
Lactobacillus sp. M6,
Bacillus sp. N16,
Aeromonas sp. N8,
Alcaligenes sp. P2,
Alcaligenes sp. N22,
Alcaligenes sp. E4,
Vibrio sp. M25,
Pseudomonas sp. P1,
Pseudomonas sp. N9, and
Kurthia sp. P3), with MIC values ranging from 5.36 to 82.72 μM (1.62–25.0 µg/mL). This compound was also shown to inhibit the cyanobacterial
Anabaena sp. and
Nostoc sp. spore germination at 100 μg/mL and the adherence of the
Perna indica mussel foot at 306 ± 19.6 μg/mL [
30]. Interestingly, the compound quercetin (
64) was tested in different studies against different marine organisms, namely a mussel species, marine bacteria, and algae, showing its potential to act against different levels of fouling community.
Our group also reported the synthesis and antifouling evaluation of a series of nature-inspired sulfated compounds, including flavonoids [
31]. Among them, rutin persulfate (
75) showed moderate anti-settlement activity towards plantigrades of the mussel
Mytilus galloprovincialis, with an EC
50 value of 22.59 μM and a LC
50/EC
50 ratio >22.13. Moreover, this flavonoid also displayed significant inhibitory activity against the growth of the marine bacteria
Vibrio harveyi (EC
50 value of 7.69 µM) and was shown to be non-toxic in
Artemia salina and Luminescent
Vibrio fischeri ecotoxicity assays. In this study, another sulfated flavonoid glycoside (compound
76) was shown to moderately inhibit the growth of the marine bacteria
Halomonas aquamarina, with an EC
50 value of 42.3 µM [
31].
Haider and colleagues tested the antifouling activity of a series of flavonoids against diatoms
Chaetoceros socialis,
Navicula leavissima, and
Navicula parva, including seven flavonols, namely casticin (
71) and flavonols
77–
82 [
32]. Flavonols
77–
79 were isolated from the marine-derived fungus
Aspergillus candidus, whereas the natural flavonoids,
80,
81 (morin) and
82, tested in this study were commercially obtained. All tested flavonols displayed antidiatom activity against
C. socialis (EC
50 11.62–44.73 μM, 4.0–13.52 µg/mL). Regarding their activity against
N. leavissima, compounds
71,
80, and
81 displayed activities with EC
50 values ranging between 20.32 and 54.33 μM (6.14–16.42 µg/mL). Only flavonols
80–
81 showed activity against
N. parva (EC
50 18.76–20.38 μM, 5.67–6.16 µg/mL), whereas the commercial biocide Sea-Nine 211 showed activity against all strains of diatoms, with EC
50 values ranging between 3.79 and 5.21 μM (1.07–1.47 µg/mL). Interestingly, these two flavonols with hydroxy groups as substituents on the B ring displayed activity against the three strains of diatoms tested, being the most promising compounds in the study, whereas an unsubstituted B ring leads to compound
82, which has lower antidiatom activity [
32].
2.3. Flavones
Thirteen flavones were also found to exert some influence in the prevention of marine biofouling, being the structures of compounds presented in
Figure 5.
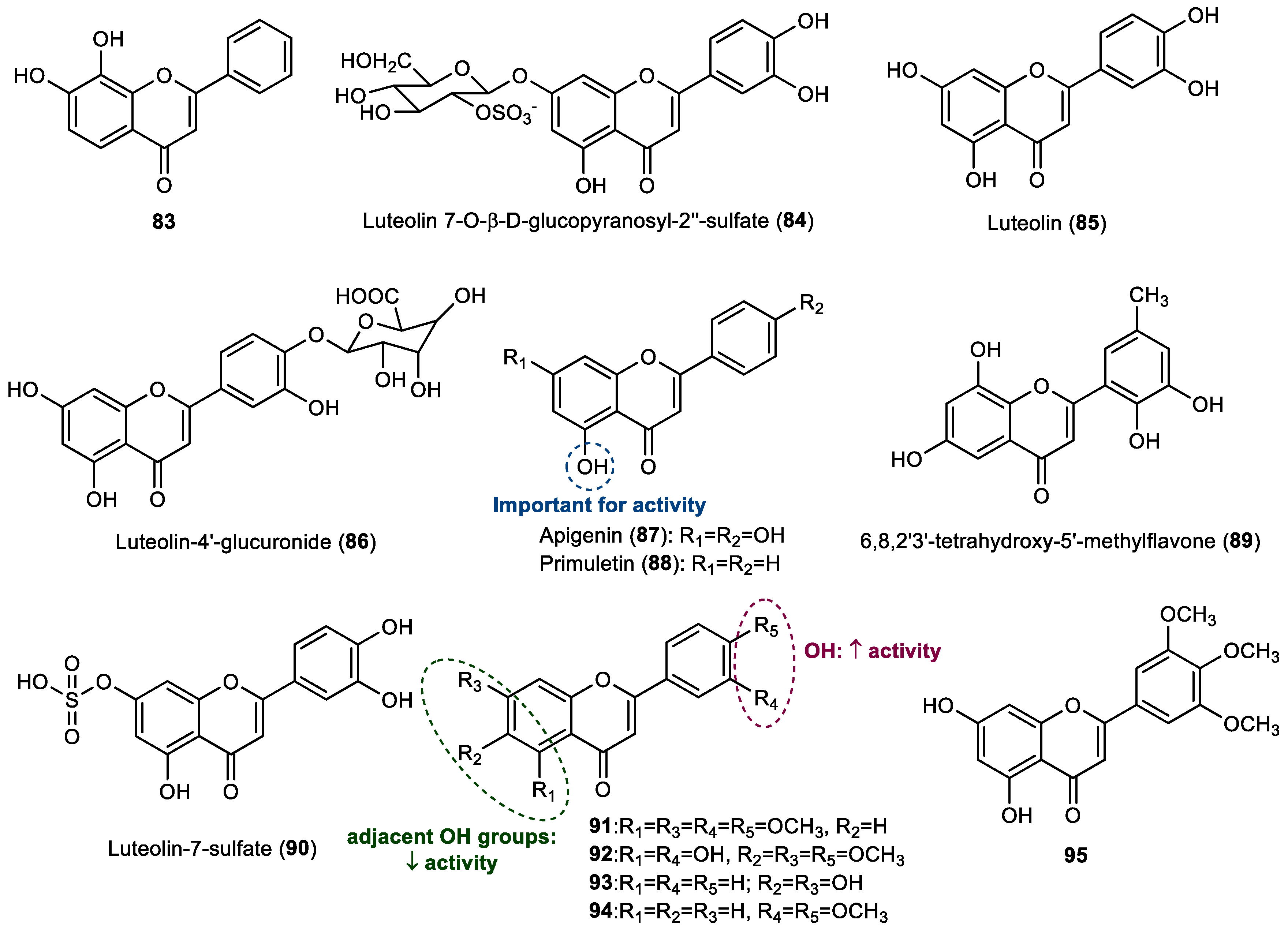
Besides the evaluation of a series of flavonols, already reported in
Section 2.2, Singh et al. also tested the activity of 7,8-dihydroxyflavone (
83), known as tropoflavin, against the settlement of the blue mussel
M. edulis. This compound showed 25% of the activity of the positive control copper sulfate [
23]. Although this compound was commercially obtained for this study, tropoflavin occurs in nature, namely in terrestrial plants, and has been proven to exert therapeutic potential [
33].
Jensen et al. observed that a lower number of thraustochytrid protists were observed to be associated with healthy leaf tissue in the marine angiosperm
Thalassia testudinum than were found in sterilized samples, suggesting that
T. testudinum may biosynthesize some metabolites which could be responsible for the low attachment of the thraustochytrid
Schizochytrium aggregatum [
34]. Therefore, the antifouling potential of
T. testudinum extracts was evaluated and studies were performed aiming to identify the possible natural products responsible for the anti-settlement activity. It was demonstrated that extracts of
T. testudinum leaf tissues inhibited the growth of the co-occurring thraustochytrid
S. aggregatum and deterred the attachment of
S. aggregatum motile zoospores to an extract-impregnated substrate. Bioassay-guided chemical fractionation of this extract allowed us to identify the flavone glycoside luteolin 7-
O-
β-
d-glucopyranosyl-2″-sulfate (
84), which was able to reduce the attachment of
S. aggregatum zoospores, with an IC
50 of 511.9 μM (270 µg/mL) [
34]. These results support the hypothesis that luteolin 7-
O-
β-
d-glucopyranosyl-2″-sulfate (
84) could be responsible for the anti-settlement activity observed in the
Thalassia testudinum extracts.
Qi et al. reported the extraction and isolation of flavonoids from the South China Sea seagrass
Enhalus acoroides [
35]. Among them, the flavones luteolin (
85) and luteolin-4′-glucuronide (
86) showed some antibacterial activity against several strains of marine bacteria, with luteolin being active against
Loktanella hongkongensis,
Pseudoalteromonas piscida,
Rhodovulum sp., and
Vibrio alginolyticus, whereas luteolin-4′-glucuronide presented some activity against
L. hongkongensis,
V. alginolyticus,
Vibrio furnissii, and
Vibrio halioticoli. Moreover, luteolin 4′-glucuronide highly inhibited the larval settlement of
Bugula neritina at a nontoxic concentration with an EC
50 value of 1.12 μM (0.52 µg/mL) [
35].
Later, luteolin (
85), obtained from the terrestrial plant
Arachis hypogata, as well as apigenin (
87), obtained from terrestrial plant
Apium graveolens, and the commercially obtained primuletin (
88) were evaluated against the larval settlement of the barnacle
A. amphitrite by Zhou et al., along with the flavonols reported in
Section 2.2. [
28]. Although primuletin was commercially obtained for this study, this flavone is found in nature, namely in terrestrial plants [
36]. These compounds were shown to have a high activity, with EC
50 values ranging from 11.47 to 13.28 μM (2.8–3.8 µg/mL). Considering that these compounds presented with similar activity, it seems that the presence of the hydroxy group at C5 is important for antifouling activity against barnacle larval settlement, whereas the presence of the other substituents is not essential for this activity [
28].
The 6,8,2′3′-tetrahydroxy-5′-methylflavone (
89), isolated from a broth of gorgonian coral-associated fungus
Penicillium sp. SCSGAF 0023 by Bao et al., showed significant antifouling activity against the larval settlement of
A. amphitrite, with an EC
50 value of 22.35 μM (6.71 µg/mL) and low toxicity (LC
50/EC
50 ratio > 14.9) [
37].
Considering the potential of the surface of the eelgrass Zostera marina to inhibit the colonization of microfouler organisms, Guan et al. performed an extraction of the
Z. marina leaves in propanol and tested the extracted fractions against marine bacterial strains [
38]. The authors identified the presence of the flavones luteolin (
85) and luteolin-7-sulfate (
90) in the extract which might be responsible for the activity. Furthermore, both flavones, luteolin (
85) and luteolin-7-sulfate (
90) were tested against the marine bacteria
Vibrio cyclitrophicus and
Marivita litorea. At concentration of 14.5 µg/mL, both luteolin (
85) and luteolin-7-sulfate (
90) exhibited comparable activity against both marine bacteria [
38].
Besides the evaluation of flavonols
71 and
77–
82 against diatoms
C. socialis,
N. leavissima, and
N. parva, Haider et al. also studied the activity of the flavones luteolin (
85) and flavones
91–
94 against these biofouling organisms [
32]. Luteolin (
85) was the most promising flavone in this study, being active against all strains of diatoms tested (
N. leavissima: EC
50 = 30.39 μM, 8.70 µg/mL;
C. socialis: EC
50 = 21.59 μM, 6.18 µg/mL; and
N. parva: EC
50 = 27.15 μM, 7.77 µg/mL), whereas flavones
91–
92 were active against
N. leavissima (
91: EC
50 of 26.52 µM, 9.08 µg/mL;
92: 58.32 µM, 20.08 µg/mL) and
C. socialis (
91: EC
50 of 10.75 µM, 3.68 µg/mL;
92: 25.15 µM, 8.66 µg/mL). Compound
93 was only active for
C. socialis (EC
50 57.15 µM, 14.53 µg/mL), while compound
94 only showed activity against
N. leavissima (EC
50 of 55.19 µM, 15.58 µg/mL) [
32]. The presence of hydroxy groups on the B ring seems to be favorable for antidiatom activity, as well as the substituents on the A ring, but the presence of adjacent hydroxy groups on the A ring is not beneficial.
Flavone tricetin 3′,4′,5′-trimethyl ether (
95), synthesized by us, showed promising activity against the settlement of the mussel
M. galloprovincialis larvae, with an EC
50 value of 8.34 µM (2.87 µg/mL), and with low toxicity against the non-target marine organism
A. salina [
18]. In an attempt to study the mechanism of action of antifouling compounds, and based on the role of tyrosinase enzyme on the production of byssal threads by fouling organisms, an evaluation of the inhibition of this enzyme was performed, with flavone
95 able being to decrease tyrosinase activity, reaching 23.5% of inhibition at 100 µM [
18].
2.4. Flavanones
As far as we know, only four flavanones with antifouling activity have been reported so far (
Figure 6).
Yoshioka et al. showed that the flavanone naringenin (
96), isolated from the methanol extracts of the bark of
Prunus jamasakura showed some repellent activity against the blue mussel
M. edulis at a concentration of 1.2 mg/cm
2, although the activity of this flavonoid is lower than the positive control, copper sulfate, at a concentration of 0.08 mg/cm
2 [
39].
Naringenin (
96), as well as the flavanones naringin (
97) (obtained commercially) and pinocembrin (
98) (isolated from
Eucalyptus signata), were evaluated against the settlement of blue mussel
M. edulis by Singh et al. and the activity was compared with the standard antifouling copper sulfate [
23]. In comparison with positive control, compounds
96,
97 and
98 presented lower activity, having 11%, 24%, and 25% of the activity of the positive control copper sulfate, respectively. Naringin
97 showed to be more active than naringenin
96, inferring the importance of glycosyl linkage for this activity [
23].
The flavanone 2′-methoxykurarinone (
99), isolated from the ethyl acetate extract of the Chinese herb
Sophora flavescens by Feng and coworkers, was shown to strongly inhibit the settlement of cyprid larvae of
Balanus albicostatus (EC
50 of 4.46 µM, 2.02 µg/mL) by a non-toxic mechanism, since the LC
50 value was higher than 25 µg/mL [
40].
2.5. Isoflavones
A total of three isoflavones with antifouling activity were described (
Figure 7).
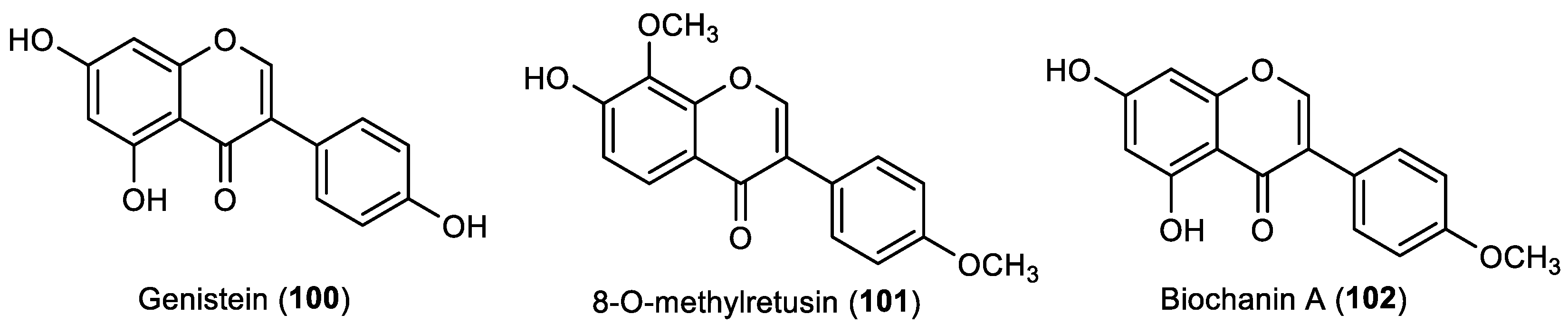
Besides the activity evaluation of the flavanone naringenin (
96), Yoshioka et al. showed that the isoflavone genistein (
100), also isolated from the methanolic extracts of the bark of
Prunus jamasakura, exhibited some anti-settlement activity against the blue mussel
M. edulis. Nevertheless, the activity of this isoflavone (9% of mussel settlement inhibition at 1.6 mg/cm
2 concentration) was substantially lower when compared with the positive control copper sulfate (100% of mussel settlement inhibition at 0.08 mg/cm
2 concentration) and also lower than the flavanone naringenin [
39].
In addition to the evaluation of seven flavonols (
Section 2.2), one flavone (
Section 2.3) and three flavanones (
Section 2.4), Singh and collaborators also tested the activity of isoflavone genistein (
100) against the settlement of blue mussel
M. edulis. This compound was found to display activity in the inhibition of the settlement of mussels, with 9% of the activity of the positive control copper sulfate, having a similar activity compared to the other flavonoids tested in the same study [
23].
The antifouling activity of genistein (
100), as well as the isoflavone 8-
O-methylretusin (
101), was also assessed by Zhou et al. The authors studied the effect of some flavonoids, including isoflavones genistein and 8-
O-methylretusin, which were obtained by the purification of extracts of terrestrial plants
Genista tinctoria and
Pueraria alopecuroides, respectively, against the larval settlement of the barnacle
A. amphitrite [
28]. Genistein (
100) presented strong activity (EC
50: 11.10 µM, 3.0 µg/mL; LC
50/EC
50: >16.7), while isoflavone 8-
O-methylretusin (
101) presented with an EC
50 value of 44.59 µM (13.3 µg/mL). Complementary assays in the field with the incorporation of the genistein in a resin-based paint showed that, after one month of the test panels being submerged, the density of barnacles was significantly lower when compared to the control panel tests. Moreover, some panels were coated with a mixture of genistein and a copper sulfate formulation, which showed lower densities of barnacles than the panels coated with the single compound after 3 months of submergence. Considering that the fate of the environment is a concern for the study of potential antifoulants, to evaluate the genistein behavior in seawater, hydrolysis assays were performed. Genistein seemed to be readily hydrolyzed in the marine environment, with the degradation speed being significantly affected by pH, namely in an alkaline environment, which should counteract the accumulation of high levels of this compound in marine environments [
28].
In other study, it was reported that
O-methylretusin (
101), isolated from the methanolic extract of the leaves of the marine halophyte
Apocynum venetum, presented activity against the marine fouling bacteria
B. thuringiensis (inhibition zone 9.4 mm) and
P. aeruginosa (inhibition zone 10.5 mm) at a concentration of 100 µg/disc, whereas the positive control chloramphenicol presented an inhibition zone of 33.7 and 30.3 mm at the same concentration [
29]. These results suggest the potential of
O-methylretusin (
101) to prevent biofouling promoted by macro and microfouling agents.
In addition to the evaluation of flavonols
71 and
77–
82, and flavones
81 and
91–
94 described in
Section 2.2 and
Section 2.3, respectively, Haider and collaborators also tested the antifouling activity of the isoflavone biochanin A (
102) against diatoms
C. socialis,
N. leavissima, and
N. parva [
32]. Although a commercial stock was used, this compound is found in nature, particularly in terrestrial plants, and display a wide range of biological activities [
41]. This compound was able to inhibit the growth of diatom
C. socialis, with an EC
50 value of 28.60 µM (8.13 µg/mL), whereas the positive control Sea-Nine 211
® presented an EC
50 value of 3.93 µM (1.11 µg/mL). Moreover, compared to the other flavonoids tested in the same study, biochanin A (
102) was only active against one strain of diatom, as well as flavonoids
77–
79,
82, and
93–
94, whereas the remaining flavonoids were active against two or three strains of diatoms [
32].
2.6. Other Flavonoids
Other flavonoids, including two flavans, one flavanol, and one flavanonol (
Figure 8), also showed some potential in marine biofouling prevention and are reported below.
Sideroxylonal A (
103), a flavan isolated from the leaves of
Eucalyptus grandis by Singh et al., showed high repellent activity against blue mussel
M. edulis at a concentration of 0.032 µmol/cm
2 [
42]. Another flavan, grandinal (
104), isolated from the methanolic extract of the leaves of
E. grandis by the same research group, also showed anti-settlement activity against the blue mussel
M. edulis, comparable with the standard antifouling copper sulfate at the same concentrations tested [
43]. Interestingly, the weaker activity of grandinal (
104) compared with sideroxylonal A (
103) may be due to the presence of tautomeric forms of grandinal. Grandinal was also found to exert some antibacterial activity against
Staphylococcus aureus and
B. subtilis, with an MIC of 50 and 100 µg/mL, respectively, which leads us to infer the dual activity of this molecule against both macrofouling and microfouling species [
43].
In addition to the evaluation of flavonols
63 and
64, and isoflavone
101 as referred to in
Section 2.2 and
Section 2.5, respectively, Kong et al. also studied the activity of the flavanol epicatechin (
105), isolated from the methanolic extract of the leaves of the marine halophyte
Apocynum venetum, against the marine fouling bacteria
B. thuringiensis,
P. elyakovii, and
P. aeruginosa. At a concentration of 100 μg/disc, this compound showed some antibacterial activity against all the tested bacteria, with an inhibition zone diameter of 9.2–10.9 mm, whereas the positive control chloramphenicol presented an inhibition zone diameter of 29.2–33.7 mm at the same concentration. Moreover, compound
105 was shown to display a similar activity to flavonols
63 and
64, whereas isoflavone
101 was only active against two strains of marine bacteria [
29].
Gopikrishnan et al. reported the production and isolation of the flavanonol taxifolin (
106) from the mangrove-derived actinobacterium
Streptomyces sampsonii (PM33) [
44]. After the isolation and characterization of this flavonoid, antifouling assays were performed. It was found that the taxifolin (
106) is active against several biofouling bacterial strains, namely
Staphylococcus sp. M1,
Micrococcus sp. M50,
Lactobacillus sp. M6,
Bacillus sp. N16,
Aeromonas sp. N8,
Alcaligenes sp. P2,
Alcaligenes sp. N22,
Alcaligenes sp. E4,
Vibrio sp. M25,
Pseudomonas sp. P1,
Pseudomonas sp. N9, and
Kurthia sp. P3, with MIC values ranging from 5.32 to 82.17 µM; 1.62–25 μg/mL. Moreover, at a concentration of 100 μg/mL, this compound was shown to reduce the
Anabaena sp. and
Nostoc sp. spore germination by more than 70%. Taxifolin (
106) was also shown to reduce the adherence of the mussel
Perna indica. Furthermore, field experiments revealed the antifouling activity of taxifolin when tested on wooden surface and PVC panels. A toxicity assay using zebra fish model showed that in a concentration of 0.5 μg/mL and 1 μg/mL, taxifolin (
106) caused pericardial edema in 0.6% of zebrafish embryos, while with an increase in the concentration to 1.5 μg/mL and 2.0 μg/mL, 60% of zebrafish embryos had pericardial edema. The cytotoxicity of taxifolin (
106) was also assayed in vitro against the normal lung Bronchial Epithelial BEAS – 2B cell line, with LC
50 values ranging between 150 μg/mL and 200 μg/mL [
44].


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







