

Effects of Polymannuronic Acid on the Intestinal Microbiota in Mice after Long-Term Intragastric Administration

Abstract
:1. Introduction
2. Results
2.1. Effects of PM Intragastric Administration on Body Weight and Fecal Water Content in Mice
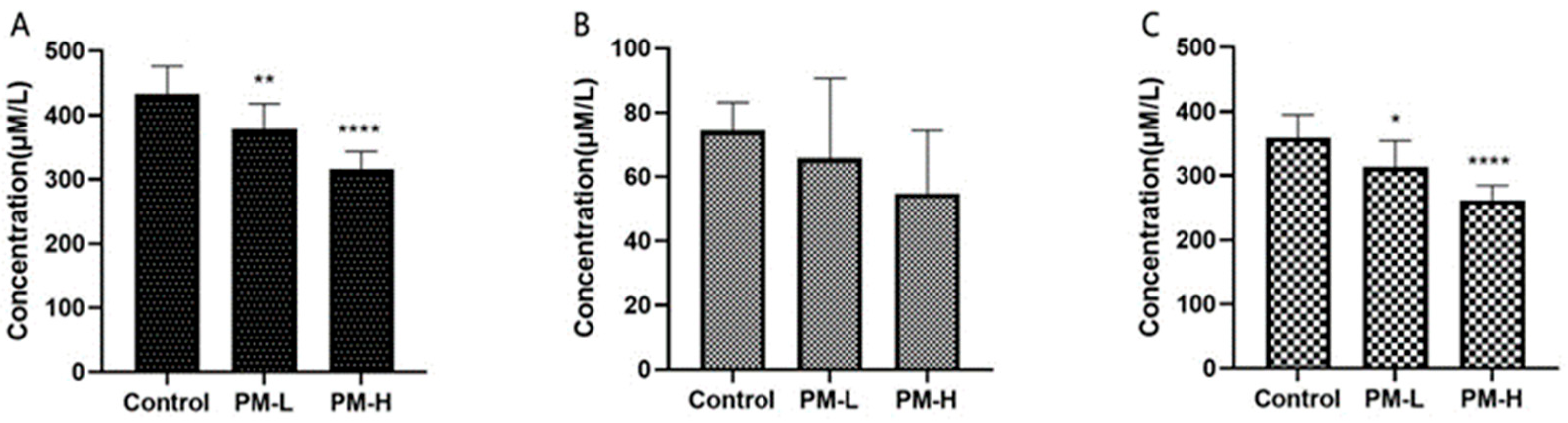
2.2. Effects of PM Intragastric Administration on the Organ Indices of Mice
2.3. 16S rRNA Gene Sequencing
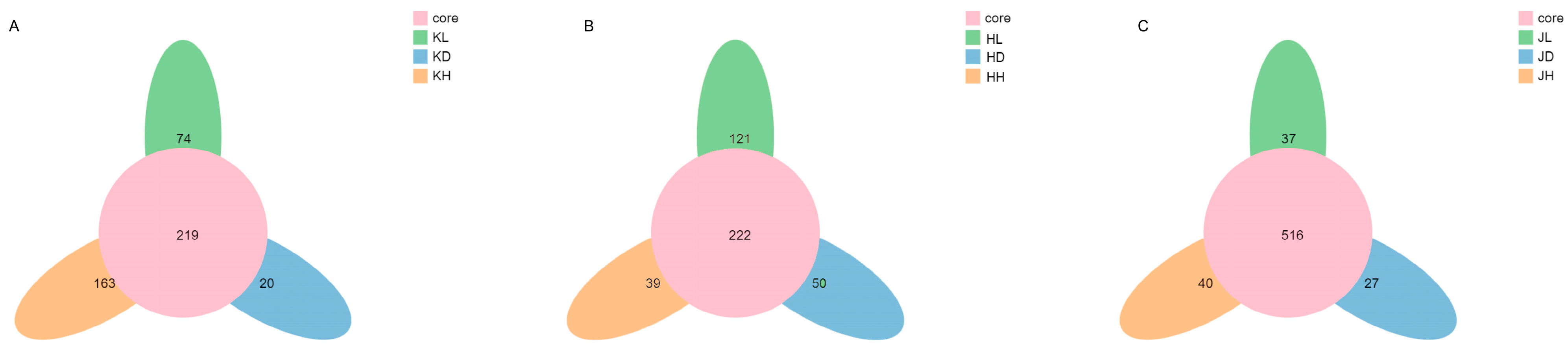
2.3.1. Analysis of Mouse Intestinal Flora with Operation Taxonomic Unit (OTU) Core-Pan Map
2.3.2. Effect of PM on the Alpha Diversity of Intestinal Flora in Mice
2.3.3. Effect of PM on the Beta Diversity of Intestinal Flora in Mice
2.3.4. Analysis of Species Composition of the Microflora in Each Intestinal Segment after the Intragastric Administration of PM
- Bacteroidetes→Bacteroidia→Bacteroidales→Prevotellaceae→Prevotella;
- Firmicutes→Negativicutes→Selenomonadales→Acidaminococcaceae→Phascolarctobacterium;
- Firmicutes→Bacilli→Lactobacillales→Lactobacillaceae→Lactobacillus.
2.3.5. Analysis of the Differences in the Composition of Dominant Species at the Portal Level of Each Intestinal Segment after the Intragastric Administration of PM
2.3.6. Analysis of the Horizontal Dominant Species Composition in Each Intestinal Segment after the Intragastric Administration of PM
2.3.7. Correlation Analysis of Dominant Species in Each Intestinal Segment after Intragastric Administration of PM
2.3.8. Analysis of the Kyoto Encyclopedia of Genes and Genomes (KEGG) Function Difference in Each Intestinal Segment after Intragastric Administration of PM
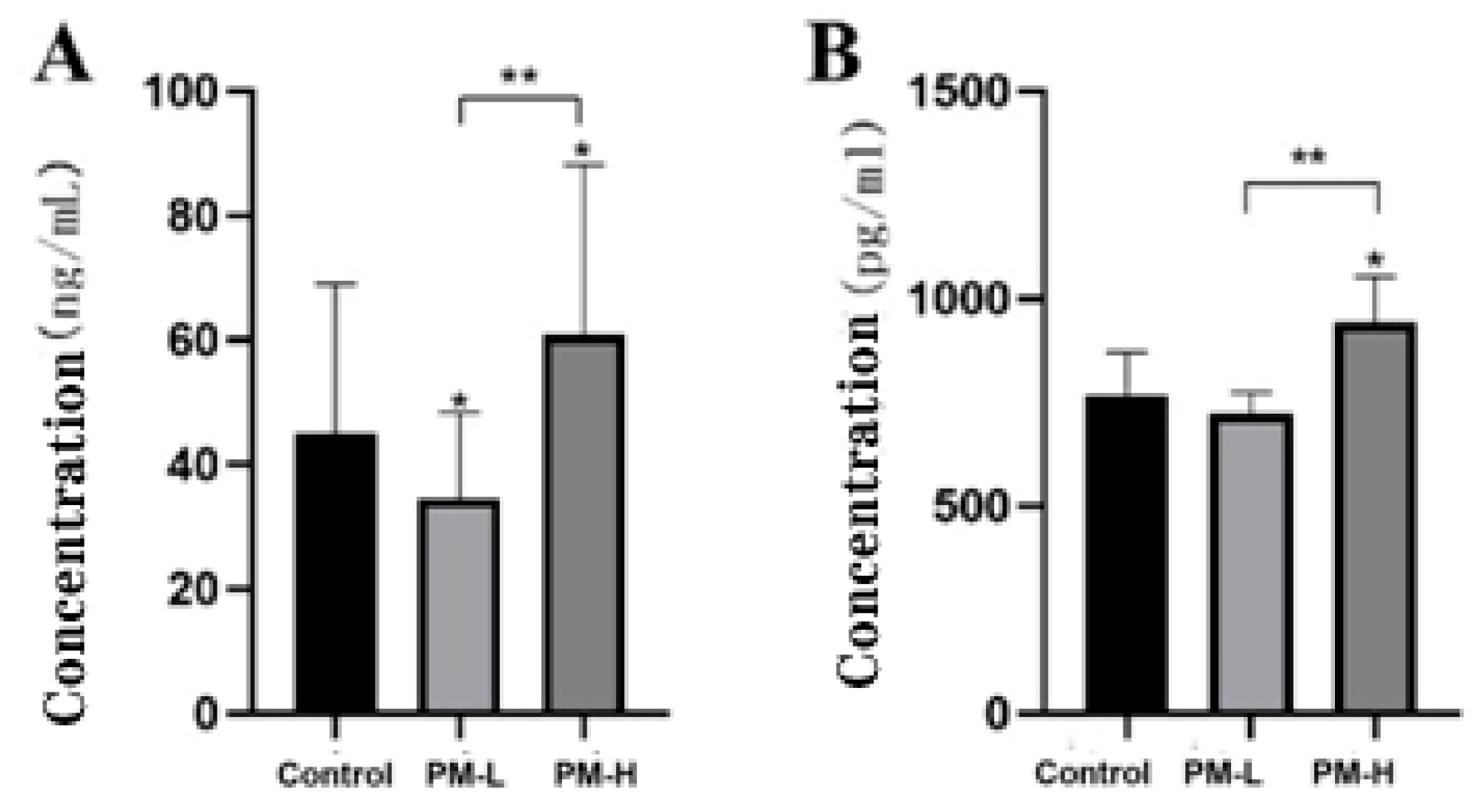
2.4. Validation of 16S rRNA Gene Analysis of Intestinal Flora in Mice
3. Discussion
4. Materials and Methods
4.1. Main Experimental Instruments and Reagents
4.2. Grouping of Experimental Animals and PM Administration
4.3. Monitoring the Changes in Body Weight and Fecal Water Content in Mice
4.4. Sampling of the Blood, Intestinal Segments, and Organs of Mice
4.5. Sample Collection and 16S rRNA Gene Sequencing to Determine Intestinal Flora
4.6. Verification of Findings from 16S rRNA Analysis of Mouse Intestinal Flora
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wee, S.; Gombotz, W.R. Protein release from alginate matrices. Adv. Drug Deliv. Rev. 1998, 31, 267–285. [Google Scholar]
- Remya, R.R.; Samrot, A.V.; Kumar, S.S.; Mohanavel, V.; Karthick, A.; Chinnaiyan, V.K.; Umapathy, D.; Muhibbullah, M. Bioactive Potential of Brown Algae. Adsorpt. Sci. Technol. 2022, 2022, 9104835. [Google Scholar] [CrossRef]
- Xing, M.C.; Cao, Q.; Wang, Y.; Xiao, H.; Zhao, J.R.; Zhang, Q.; Ji, A.G.; Song, S.L. Advances in Research on the Bioactivity of Alginate Oligosaccharides. Mar. Drugs 2020, 18, 144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, M.; Rauf, A.; Khalil, A.A.; Shan, Z.; Chen, C.; Rengasamy, K.R.R.; Wan, C. Process and applications of alginate oligosaccharides with emphasis on health beneficial perspectives. Crit. Rev. Food Sci. Nutr. 2023, 63, 303–329. [Google Scholar] [CrossRef] [PubMed]
- Mrudulakumari Vasudevan, U.; Lee, O.K.; Lee, E.Y. Alginate derived functional oligosaccharides: Recent developments, barriers, and future outlooks. Carbohydr. Polym. 2021, 267, 118158. [Google Scholar] [CrossRef]
- Song, S.; Wei, Q.; Wang, K.; Yang, Q.; Wang, Y.; Ji, A.; Chen, G. Fluorescent Labeling of Polymannuronic Acid and Its Distribution in Mice by Tail Vein Injection. Mar. Drugs 2022, 20, 289. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Xiu, C.-K.; Yang, J.; Wang, X.; Hu, Y.-H.; Fang, J.-Y.; Lei, Y. Effect of Ginseng Radix et Rhizoma, Notoginseng Radix et Rhizoma and Chuanxiong Rhizoma extracts on intestinal flora of vascular aging mice induced by high glucose and high lipid. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2020, 45, 2938–2946. [Google Scholar]
- Wei, S.; Bahl, M.I.; Baunwall, S.M.D.; Hvas, C.L.; Licht, T.R. Determining Gut Microbial Dysbiosis: A Review of Applied Indexes for Assessment of Intestinal Microbiota Imbalances. Appl. Environ. Microbiol. 2021, 87, e00395-21. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Weiss, J. Bactericidal/permeability-increasing protein (BPI) and lipopolysaccharide-binding protein (LBP): Structure, function and regulation in host defence against Gram-negative bacteria. Biochem. Soc. Trans. 2003, 31, 785–790. [Google Scholar] [CrossRef]
- Schumann, R.R. Function of lipopolysaccharide (LPS)-binding protein (LBP) and CD14, the receptor for LPS/LBP complexes: A short review. Res. Immunol. 1992, 143, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Sakura, T.; Morioka, T.; Shioi, A.; Kakutani, Y.; Miki, Y.; Yamazaki, Y.; Motoyama, K.; Mori, K.; Fukumoto, S.; Shoji, T.; et al. Lipopolysaccharide-binding protein is associated with arterial stiffness in patients with type 2 diabetes: A cross-sectional study. Cardiovasc. Diabetol. 2017, 16, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Biologic basis for interleukin-1 in disease. Blood 1996, 87, 2095–2147. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor. Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Asadpoor, M.; Ithakisiou, G.-N.; Henricks, P.A.J.; Pieters, R.; Folkerts, G.; Braber, S. Non-Digestible Oligosaccharides and Short Chain Fatty Acids as Therapeutic Targets against Enterotoxin-Producing Bacteria and Their Toxins. Toxins 2021, 13, 175. [Google Scholar] [CrossRef] [PubMed]
- Nie, Q.X.; Chen, H.H.; Hu, J.L.; Tan, H.Z.; Nie, S.P.; Xie, M.Y. Effects of Nondigestible Oligosaccharides on Obesity. In Annual Review of Food Science and Technology; Doyle, M.P., McClements, D.J., Eds.; Annual Reviews: Palo Alto, CA, USA, 2020; Volume 11, pp. 205–233. [Google Scholar]
- Liu, Y.; Chen, J.; Tan, Q.; Deng, X.; Tsai, P.J.; Chen, P.H.; Ye, M.; Guo, J.; Su, Z. Nondigestible Oligosaccharides with Anti-Obesity Effects. J. Agric. Food Chem. 2020, 68, 4–16. [Google Scholar] [CrossRef]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Fernandes, R.; do Rosario, V.A.; Mocellin, M.C.; Kuntz, M.G.F.; Trindade, E.B.S.M. Effects of inulin-type fructans, galacto-oligosaccharides and related synbiotics on inflammatory markers in adult patients with overweight or obesity: A systematic review. Clin. Nutr. 2017, 36, 1197–1206. [Google Scholar] [CrossRef]
- Nazli, A.; Yang, P.C.; Jury, J.; Howe, K.; Watson, J.L.; Soderholm, J.D.; Sherman, P.M.; Perdue, M.H.; McKay, D.M. Epithelia under metabolic stress perceive commensal bacteria as a threat. Am. J. Pathol. 2004, 164, 947–957. [Google Scholar] [CrossRef]
- Boulange, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.-E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Zoetendal, E.G.; Raes, J.; van den Bogert, B.; Arumugam, M.; Booijink, C.C.G.M.; Troost, F.J.; Bork, P.; Wels, M.; de Vos, W.M.; Kleerebezem, M. The human small intestinal microbiota is driven by rapid uptake and conversion of simple carbohydrates. ISME J. 2012, 6, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Galvez-Ontiveros, Y.; Paez, S.; Monteagudo, C.; Rivas, A. Endocrine Disruptors in Food: Impact on Gut Microbiota and Metabolic Diseases. Nutrients 2020, 12, 1158. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Lombard, V.; Henrissat, B. Complex Carbohydrate Utilization by the Healthy Human Microbiome. PLoS ONE 2012, 7, e28742. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Pereira Indiani, C.M.; Rizzardi, K.F.; Castelo, P.M.; Caldas Ferraz, L.F.; Darrieux, M.; Parisotto, T.M. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef]
- Riva, A.; Borgo, F.; Lassandro, C.; Verduci, E.; Morace, G.; Borghi, E.; Berry, D. Pediatric obesity is associated with an altered gut microbiota and discordant shifts in Firmicutes populations. Environ. Microbiol. 2017, 19, 95–105. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology—Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. Fems Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Metwaly, A.; Reitmeier, S.; Haller, D. Microbiome risk profiles as biomarkers for inflammatory and metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent Advances in Understanding Enteric Pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.; Pasolli, E.; Masetti, G.; Ercolini, D.; Segata, N. Prevotella diversity, niches and interactions with the human host. Nat. Rev. Microbiol. 2021, 19, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.J.; Nguyen, L.H.; Song, M.Y.; Wang, D.D.; Franzosa, E.A.; Cao, Y.; Joshi, A.; Drew, D.A.; Mehta, R.; Ivey, K.L.; et al. Dietary fiber intake, the gut microbiome, and chronic systemic inflammation in a cohort of adult men. Genome Med. 2021, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef]
- Pan, L.; Han, P.; Ma, S.; Peng, R.; Wang, C.; Kong, W.; Cong, L.; Fu, J.; Zhang, Z.; Yu, H.; et al. Abnormal metabolism of gut microbiota reveals the possible molecular mechanism of nephropathy induced by hyperuricemia. Acta Pharm. Sin. B 2020, 10, 249–261. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Gasbarrini, A.; Mele, M.C. Food Additives, Gut Microbiota, and Irritable Bowel Syndrome: A Hidden Track. Int. J. Environ. Res. Public Health 2020, 17, 8816. [Google Scholar] [CrossRef]
- Vitetta, L.; Llewellyn, H.; Oldfield, D. Gut Dysbiosis and the Intestinal Microbiome: Streptococcus thermophilus a Key Probiotic for Reducing Uremia. Microorganisms 2019, 7, 228. [Google Scholar] [CrossRef]
- Heeney, D.D.; Gareau, M.G.; Marco, M.L. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr. Opin. Biotech. 2018, 49, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Magruder, M.; Edusei, E.; Zhang, L.; Albakry, S.; Satlin, M.J.; Westblade, L.F.; Malha, L.; Sze, C.; Lubetzky, M.; Dadhania, D.M.; et al. Gut commensal microbiota and decreased risk for Enterobacteriaceae bacteriuria and urinary tract infection. Gut Microbes 2020, 12, 1805281. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cui, Y.; Xu, B.; Wang, Y.; Lv, F.; Li, Z.; Li, H.; Chen, X.; Peng, X.; Chen, Y.; et al. Main active components of Jiawei Gegen Qinlian decoction protects against ulcerative colitis under different dietary environments in a gut microbiota-dependent manner. Pharmacol. Res. 2021, 170, 105694. [Google Scholar] [CrossRef]
- Su, J.; Su, L.; Li, D.; Shuai, O.; Zhang, Y.; Liang, H.; Jiao, C.; Xu, Z.; Lai, Y.; Xie, Y. Antitumor Activity of Extract From the Sporoderm-Breaking Spore of Ganoderma lucidum: Restoration on Exhausted Cytotoxic T Cell With Gut Microbiota Remodeling. Front. Immunol. 2018, 9, 1765. [Google Scholar] [CrossRef] [PubMed]
- Bhandarkar, N.S.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Brown, L.; Panchal, S.K. Coffee Pulp, a By-Product of Coffee Production, Modulates Gut Microbiota and Improves Metabolic Syndrome in High-Carbohydrate, High-Fat Diet-Fed Rats. Pathogens 2021, 10, 1369. [Google Scholar] [CrossRef]
- Liu, X.M.; Mao, B.Y.; Gu, J.Y.; Wu, J.Y.; Cui, S.M.; Wang, G.; Zhao, J.X.; Zhang, H.; Chen, W. Blautia-a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Xing, M.; Wang, K.; Yang, Q.; Zhao, J.; Wang, Y.; Li, X.; Ji, K.; Song, S. Fucoidan Is Not Completely Dependent on Degradation to Fucose to Relieve Ulcerative Colitis. Pharmaceuticals 2022, 15, 430. [Google Scholar] [CrossRef]
- Yi, R.; Tan, F.; Zhou, X.; Mu, J.; Li, L.; Du, X.; Yang, Z.; Zhao, X. Effects of Lactobacillus fermentum CQPC04 on Lipid Reduction in C57BL/6J Mice. Front. Microbiol. 2020, 11, 573586. [Google Scholar] [CrossRef]
- Wang, Y.; Gunewardena, S.; Li, F.; Matye, D.J.; Chen, C.; Chao, X.; Jung, T.; Zhang, Y.; Czerwiński, M.; Ni, H.M.; et al. An FGF15/19-TFEB regulatory loop controls hepatic cholesterol and bile acid homeostasis. Nat. Commun. 2020, 11, 3612. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Chao | Ace | Shannon | Simpson | Coverage |
|---|---|---|---|---|---|
| control | 153.83 ± 21.08 | 172.28 ± 17.90 | 1.69 ± 0.21 | 0.40 ± 0.16 | 0.9995 ± 0.0002 |
| L-group | 271.22 ± 51.09 ** | 275.77 ± 49.98 ** | 2.05 ± 0.47 * | 0.39 ± 0.10 | 0.9992 ± 0.0003 |
| H-group | 306.19 ± 60.01 ** | 311.22 ± 49.91 ** | 1.55 ± 0.37 | 0.43 ± 0.10 | 0.9992 ± 0.0003 |
| Group | Chao | Ace | Shannon | Simpson | Coverage |
|---|---|---|---|---|---|
| control | 196.17 ± 59.35 | 165.14 ± 13.36 | 1.64 ± 0.31 | 0.36 ± 0.10 | 0.9993 ± 0.0002 |
| L-group | 211.21 ± 45.06 | 204.14 ± 40.38 * | 1.68 ± 0.43 | 0.30 ± 0.13 | 0.9991 ± 0.0002 |
| H-group | 248.17 ± 51.95 * | 237.57 ± 37.20 ** | 1.62 ± 0.43 | 0.40 ± 0.07 | 0.9990 ± 0.0003 |
| Group | Chao | Ace | Shannon | Simpson | Coverage |
|---|---|---|---|---|---|
| control | 373.19 ± 36.04 | 370.27 ± 40.11 | 3.54 ± 0.10 | 0.11 ± 0.04 | 0.9989 ± 0.0002 |
| L-group | 427.84 ± 38.70 * | 430.34 ± 39.68 * | 3.52 ± 0.16 | 0.11 ± 0.02 | 0.9998 ± 0.0002 |
| H-group | 455.38 ± 37.00 ** | 450.81 ± 40.94 ** | 3.42 ± 0.17 | 0.12 ± 0.02 | 0.9998 ± 0.0002 |
| Group | Jejunum | Ileum | Colon |
|---|---|---|---|
| control | 0.0040 | 0.0034 | 0.3561 |
| L-group | 0.0370 ** | 0.0110 ** | 0.3981 * |
| H-group | 0.1330 ** | 0.1095 ** | 0.9001 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, E.; Wei, Q.; Li, X.; Song, S. Effects of Polymannuronic Acid on the Intestinal Microbiota in Mice after Long-Term Intragastric Administration. Mar. Drugs 2024, 22, 125. https://doi.org/10.3390/md22030125
Zhang E, Wei Q, Li X, Song S. Effects of Polymannuronic Acid on the Intestinal Microbiota in Mice after Long-Term Intragastric Administration. Marine Drugs. 2024; 22(3):125. https://doi.org/10.3390/md22030125
Chicago/Turabian StyleZhang, E, Qiang Wei, Xia Li, and Shuliang Song. 2024. "Effects of Polymannuronic Acid on the Intestinal Microbiota in Mice after Long-Term Intragastric Administration" Marine Drugs 22, no. 3: 125. https://doi.org/10.3390/md22030125
APA StyleZhang, E., Wei, Q., Li, X., & Song, S. (2024). Effects of Polymannuronic Acid on the Intestinal Microbiota in Mice after Long-Term Intragastric Administration. Marine Drugs, 22(3), 125. https://doi.org/10.3390/md22030125