Antioxidant Activity, Inhibition of Intestinal Cancer Cell Growth and Polyphenolic Compounds of the Seagrass Posidonia oceanica’s Extracts from Living Plants and Beach Casts

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Phenological Parameters of Posidonia oceanica Plants, and Its Meadow and ‘Banquette’ Characteristics
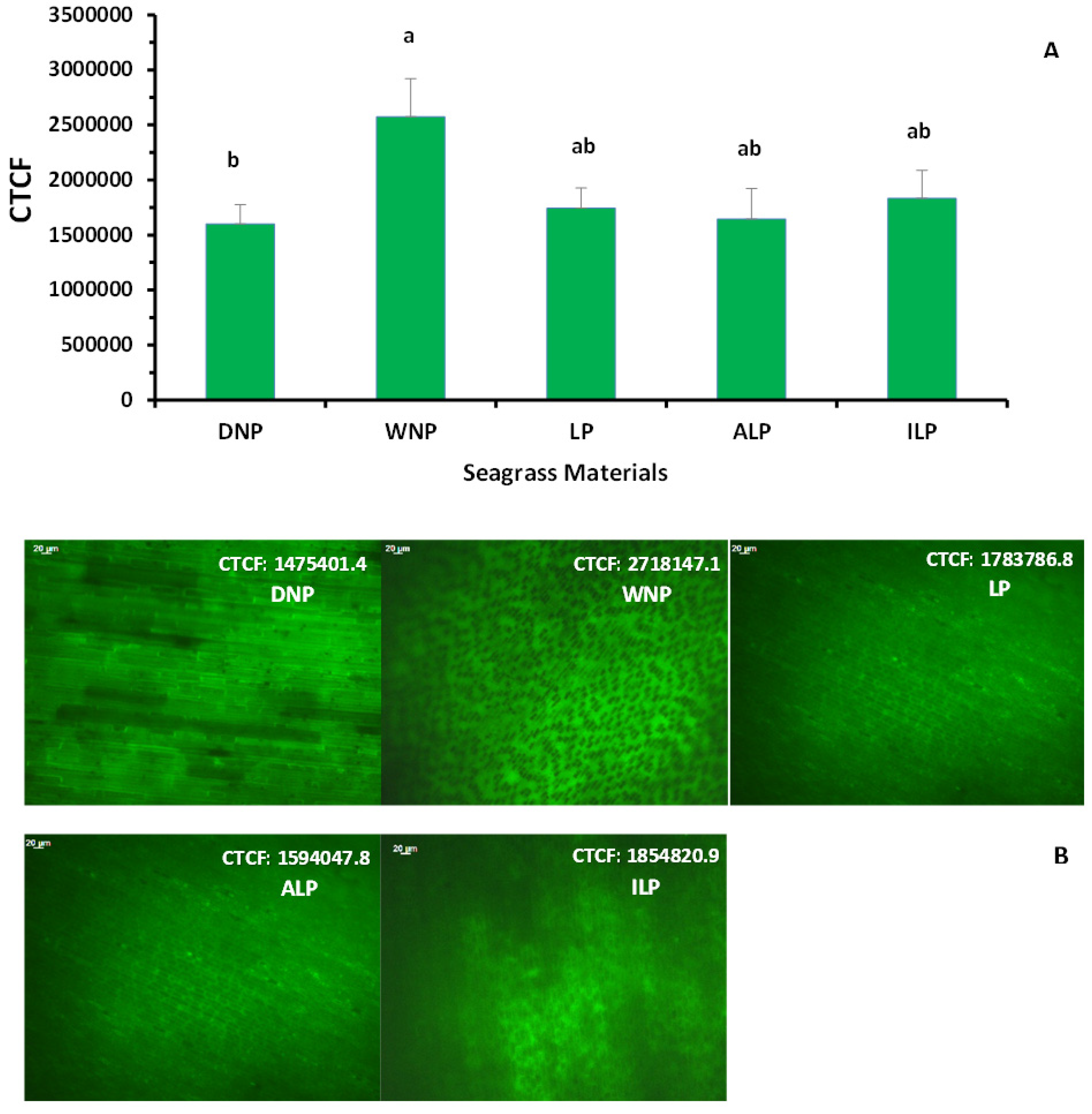
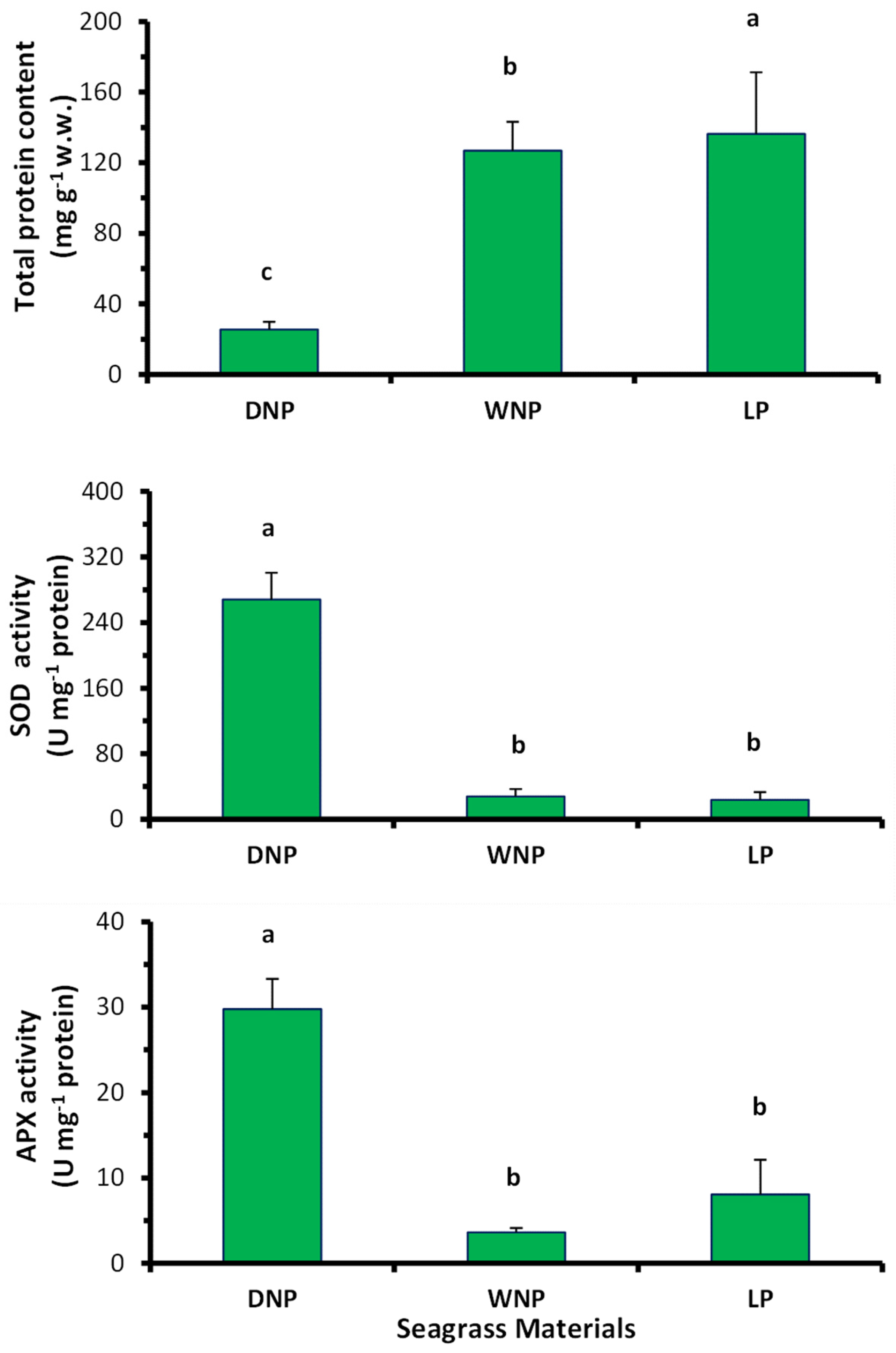
2.2. Oxidative Stress—Antioxidant Enzyme Production and Protein Content of Posidonia oceanica Leaves
2.3. Posidonia oceanica’s Extraction Yield
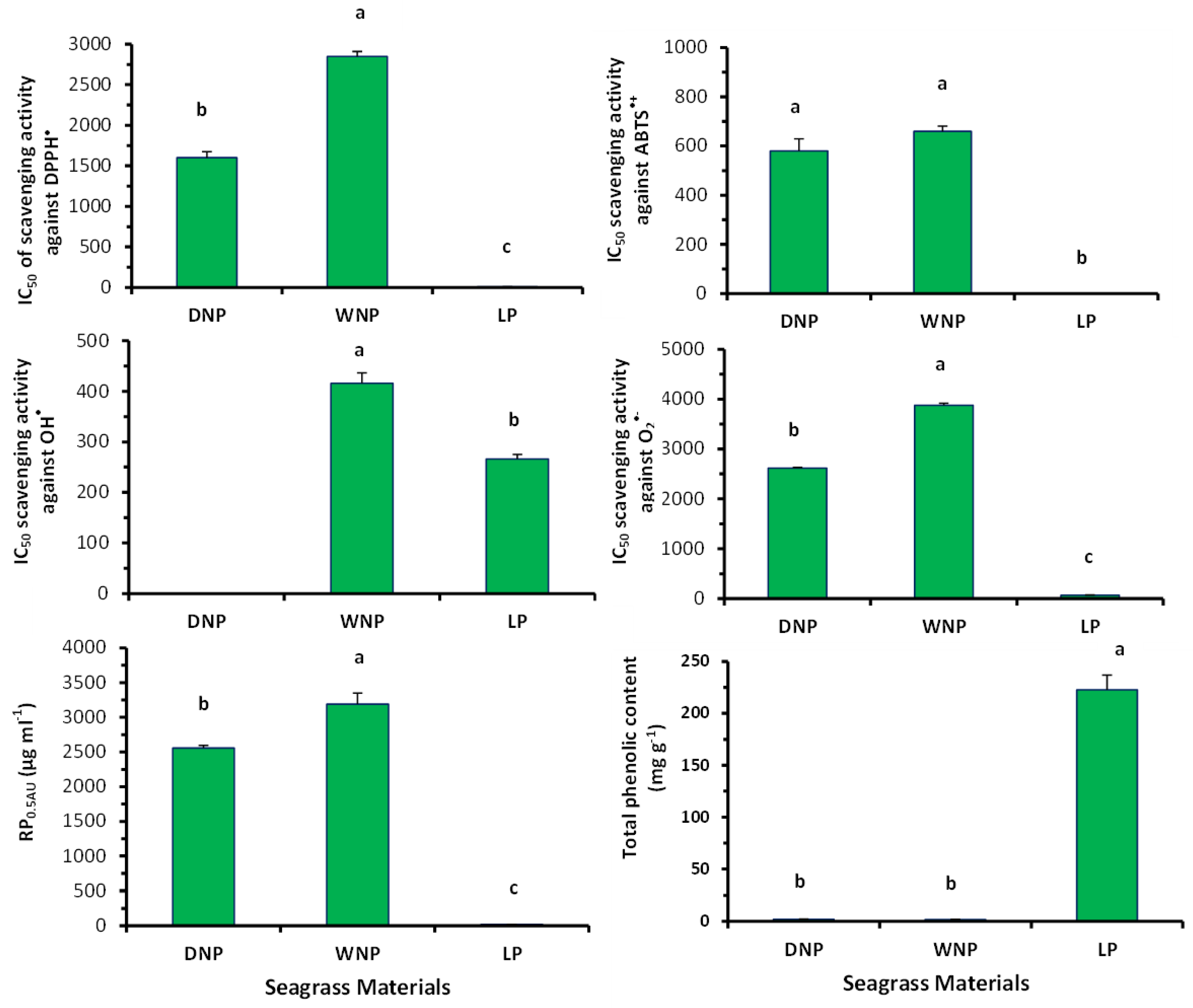
2.4. Free Radical Scavenging Activity of Posidonia oceanica’s Extracts
2.5. Phenolic Content of the Posidonia oceanica’s Extracts
2.6. Relationship between the Investigated Parameters
2.7. Antiproliferative Activity of Posidonia oceanica Extracts against Cancer Cells
2.8. Quantification of the Phenolic Compounds Produced by Posidonia oceanica Dry ‘Necromass’ in the Wider Study Area—A Case Study
3. Discussion
4. Materials and Methods
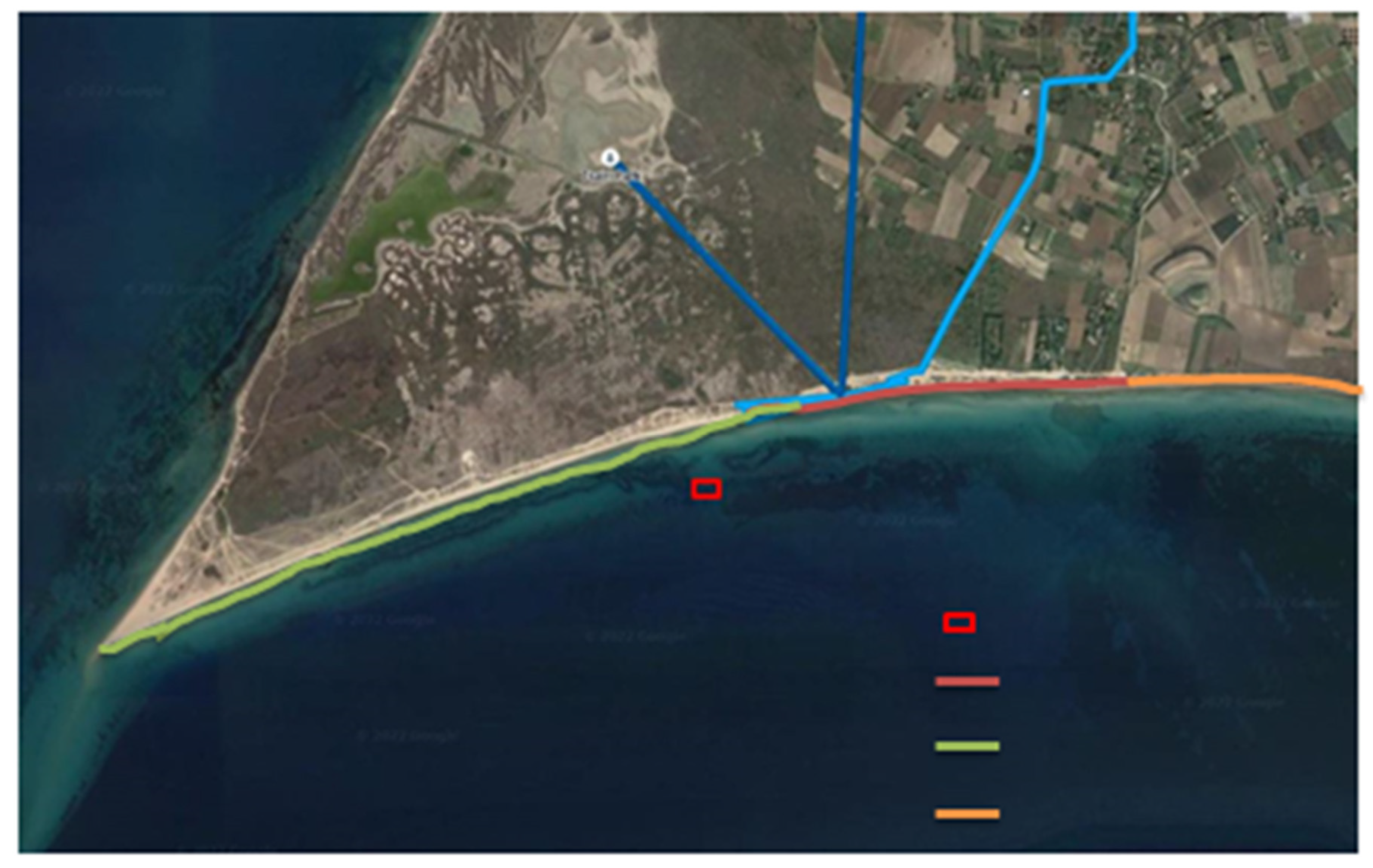
4.1. Collection of Posidonia oceanica Samples
4.2. Imaging of Hydrogen Peroxide Production in Living Leaves (LP) of Posidonia oceanica
4.3. Assessment of Antioxidant Enzymes in LP, WNP and DNP Samples and Protein Content
4.4. Preparation of Posidonia oceanica Extracts
4.5. Phenolic Content
4.6. 2.2-Diphenyl-Picrylhydrazyl (DPPH•) Radical Scavenging Assay
4.7. 2.2-Azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS•+) Radical Scavenging Assay
4.8. Hydroxyl Radical (•OH) Scavenging Assay
4.9. Superoxide Anion Radical (O2•−) Scavenging Assay
4.10. Reducing Power (RP) Assay
4.11. Cell Culture Conditions
4.12. XTT Assay for Inhibition of Cell Proliferation
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Larkum, A.W.D.; Orth, R.J.; Duarte, C.M. Seagrasses: Biology, Ecology and Conservation; Springer: Dordrect, The Netherlands, 2006; p. 691. [Google Scholar]
- Malea, P.; Mylona, Z.; Kevrekidis, T. Trace elements in the seagrass Posidonia oceanica: Compartmentation and relationships with seawater and sediment concentrations. Sci. Total Environ. 2019, 686, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Mylona, Z.; Kevrekidis, T. Improving the utility of the seagrass Posidonia oceanica as a biological indicator of past trace element contamination. Ecol. Indic. 2019, 107, 105596. [Google Scholar] [CrossRef]
- de los Santos, C.B.; Scott, A.; Arias-Ortiz, A.; Jones, B.; Kennedy, H.; Mazarrasa, I.; McKenzie, L.; Nordlund, L.M.; de la Torre-Castro, M.; Unsworth, R.K.F.; et al. Seagrass ecosystem services: Assessment and scale of benefits. In UNEP. Out of the Blue: The Value of Seagrasses to the Environment and to People; UNEP: Nairobi, Kenya, 2020; pp. 48–55. [Google Scholar]
- de los Santos, C.B.; Olivea, I.; Moreira, M.; Silva, A.; Freitas, C.; Luna, R.A.; Quental-Ferreira, H.; Martins, M.; Costa, M.M.; Silva, J.; et al. Seagrass meadows improve inflowing water quality in aquaculture ponds. Aquaculture 2020, 528, 735502. [Google Scholar] [CrossRef]
- United Nations Environment Programme-World Conservation Monitoring Centre, UNEP-WCMC. Available online: https://unep-wcmc.org/en/news/new-research-shows-which-mediterranean-coastal-communities-must-plan-now-for-seagrass-loss (accessed on 30 November 2023).
- Green, A.E.; Unsworth, R.K.F.; Chadwick, M.A.; Jones, P.J.S. Historical analysis exposes catastrophic seagrass loss for the United Kingdom. Front. Plant Sci. 2021, 12, 629962. [Google Scholar] [CrossRef] [PubMed]
- International Union for Conservation of Nature. POSBEMED 2. Governance and Management of Posidonia Beach Dune Systems across the Mediterranean; Action 3.3: Proposals for the implementation of systems management and intervention measure Posidonia in the study area; Interreg Mediterranean: Thessaloniki, Greece, 2021; p. 108. [Google Scholar]
- Rotini, A.; Chiesa, S.; Manfra, L.; Borrello, P.; Piermarini, R.; Silvestri, C.; Cappucci, S.; Parlagreco, L.; Devoti, S.; Pisapia, M. Efectiveness of the “Ecological Beach” Model: Beneficial Management of Posidonia Beach Casts and Banquette. Water 2020, 12, 3238. [Google Scholar] [CrossRef]
- Zamani, N.P.; Rahman, L.; Rosada, R.L.; Tirtama, W. Overview of bioactivity studies on marine natural products. IOP Conf. Ser. Earth Environ. Sci. 2021, 944, 012029. [Google Scholar] [CrossRef]
- Kim, J.H.; Jenrow, K.A.; Brown, S.L. Mechanisms of radiation-induced normal tissue toxicity and implications for future clinical trials. Radiat. Oncol. J. 2014, 32, 103–115. [Google Scholar] [CrossRef]
- Zulkifli, L.; Muksin, Y.D.; Hartanto, P.; Desimarlina, Y.; Idrus, A.A.; Syukus, A. Phytochemical propiles and rthnomedicine preliminary studies on seagrass species in the Southern Coast of Lombok island Indonisia. IOP Conf. Ser. Earth Environ. Sci. 2021, 913, 012076. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Sadeer, N.B.; Zengin, G.; Mahomoodally, M.F.; Cziaky, Z.; Jeko, J.; Diuzheva, A.; Abdallah, H.H.; Kim, D.H. Biopharmaceutical potential, chemical profile and in silico study of the seagrass Syringodium isoetifolium (Asch.) Dandy. S. Afr. J. Bot. 2019, 127, 167–175. [Google Scholar]
- Cocozza, C.; Parente, A.; Zaccone, C.; Mininni, C.; Santamaria, P.; Miano, T. Chemical, physical and spectroscopic characterization of Posidonia oceanica (L.) Del. residues and their possible recycle. Biomass Bioenergy 2011, 35, 799–807. [Google Scholar] [CrossRef]
- Cocozza, C.; Parente, A.; Zaccone, C.; Mininni, C.; Santamaria, P.; Miano, T. Comparative mangement of offshore Posidonia residues: Composting vs. energy recovery. Waste Manag. 2011, 31, 78–84. [Google Scholar] [CrossRef]
- Benito-González, I.; López-Rubio, A.; Martínez-Abad, A.; Ballester, A.-R.; Falcó, I.; González-Candelas, L.; Sánchez, G.; Lozano-Sánchez, J.; Borrás-Linares, I.; Segura-Carretero, A.; et al. In-Depth Characterization of Bioactive Extracts from Posidonia oceanica Waste Biomass. Mar. Drugs 2019, 17, 409. [Google Scholar] [CrossRef]
- Messina, C.M.; Arena, R.; Manuguerra, S.; Pericot, Y.; Curcuraci, E.; Kerninon, F.; Renda, G.; Hellio, C.; Santulli, A. Antioxidant Bioactivity of Extracts from Beach Cast Leaves of Posidonia oceanica (L.) Delile. Mar. Drugs 2021, 19, 560. [Google Scholar] [CrossRef] [PubMed]
- Mininni, C.; Parente, A.; Santamaria, P. Indagine sulla diffusione in Puglia dei residui spiaggiati di Posidonia. In II Caso dei Residui Spiaggiati di Posidonia oceanica: Da rifiuto a Risorsa; Parente, A., Santamaria, P., Eds.; Levante Editori: Bari, Italy, 2009; pp. 43–78. [Google Scholar]
- Rubio-Portillo, E.; Martin-Cuadrado, A.-B.; Ramos-Esplá, A.Á.; Antón, J. Metagenomics unveils Posidonia oceanica ‘banquettes’ as apotential source of novel bioactive compounds and carbohydrate active enzymes (CAZymes). mSystems 2021, 6, e00866-21. [Google Scholar] [CrossRef] [PubMed]
- Hemminga, M.A.; Duarte, C.M. Seagrass Ecology; Cambridge University Press: Cambridge, UK, 2000; p. 298. [Google Scholar]
- Malea, P.; Boubonari, T.; Kevrekidis, T. Iron, zinc, copper, lead and cadmium contents in Ruppia maritima from a Mediterranean coastal lagoon: Monthly variation and distribution in different plant fractions. Bot. Mar. 2008, 51, 320–330. [Google Scholar] [CrossRef]
- Malea, P.; Kevrekidis, T.; Potouroglou, Μ. Seasonal variation of trace metal (Mn, Zn, Cu, Pb, Co, Cd) concentrations in compartments of the seagrass Cymodocea Nodosa. Bot. Mar. 2013, 56, 169–184. [Google Scholar] [CrossRef]
- Malea, P.; Kokkinidi, D.; Kevrekidou, A.; Adamakis, I.D.S. Environmentally relevant bisphenol A concentrations effects on the seagrass Cymodocea nodosa different parts elongation: Perceptive assessors of toxicity. Environ. Sci. Pollut. Res. 2020, 27, 7267–7279. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Charitonidou, Κ.; Sperdouli, I.; Mylona, Z.; Moustakas, M. Zinc uptake, photosynthetic efficiency and oxidative stress in the seagrass Cymodocea nodosa exposed to ZnO nanoparticles. Materials 2019, 12, 2101. [Google Scholar] [CrossRef]
- Malea, P.; Mylona, Z.; Panteris, E.; Kevrekidis, D.F.; Kevrekidis, T. Nickel uptake kinetics and its structural and physiological impacts in the seagrass Halophila stipulacea. Ecotoxicol. Environ. Saf. 2021, 208, 111386. [Google Scholar] [CrossRef]
- Malea, P.; Mylona, Z.; Panteris, E.; Kevrekidis, T. Structural and physiological effects of chromium uptake in the seagrass Halophila stipulacea. Ecol. Indic. 2021, 122, 107224. [Google Scholar] [CrossRef]
- Malea, P.; Emmanouilidis, A.; Kevrekidis, D.P.; Moustakas, M. Copper uptake kinetics and toxicological effects of ionic Cu and CuO nanoparticles on the seaweed Ulva rigida. Environ. Sci. Pollut. Res. 2022, 29, 57523–57542. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Ferreira, I.C.F.R. A review on antioxidants, prooxidants and related controversy: Natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Kokkinidi, D.; Kevrekidou, A.; Adamakis, I.-D.S. The Enzymatic and Non-Enzymatic Antioxidant System Response of the Seagrass Cymodocea nodosa to Bisphenol-A Toxicity. Int. J. Mol. Sci. 2022, 23, 1348. [Google Scholar] [CrossRef] [PubMed]
- Adamakis, I.D.; Malea, P.; Sperdouli, I.; Panteris, E.; Kokkinidi, D.; Moustakas, M. Evaluation of the spatiotemporal effects of bisphenol A on the leaves of the seagrass Cymodocea nodosa. J. Hazard. Mater. 2021, 404, 124001. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.R.R.; Arumugam, R.; Anantharaman, P. Antibacterial potential of three seagrasses against human pathogens. Asian Pac. J. Trop. Med. 2010, 3, 890–893. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Arumugam, R.; Micheline, G.-D.; Anatharaman, P. Antioxidant activity of seagrasses of the Mandapan coast, India. Pharm. Biol. 2012, 50, 182–187. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Arumugam, R.; Thangaradjou, T.; Anatharaman, P. Phytochemical constituents, antioxidant properties and p-coumaric acid analysis in some seagrasses. Food Res. Int. 2013, 54, 1229–1236. [Google Scholar] [CrossRef]
- Nuissier, G.; Rezzonico, B.; Grignon-Dubois, M. Chicoric acid from Syringodium filiforme. Food Chem. 2010, 120, 783–788. [Google Scholar] [CrossRef]
- Kestaouli, O.; Marzouki, M.N.; Maugard, T.; Limam, F. In vitro evaluation of antioxidant activities of free and bound phenolic compounds from Posidonia oceanica (I.) Delile leaves. Afr. J. Biotechnol. 2011, 10, 3176–3185. [Google Scholar]
- Velika, B.; Kron, I. Antioxidant Properties of Benzoic Acid Derivatives against Superoxide Radical. Free. Radic. Antioxid. 2012, 2, 62–67. [Google Scholar] [CrossRef]
- Hamdy, A.-H.A.; Mettwally, W.S.A.; El Fotouh, M.A.; Rodriguez, B.; El-Dewany, A.I.; El-Toumy, S.A.A.; Hussein, A.A. Bioactive Phenolic Compounds from the Egyptian Red Sea Seagrass Thalass. Ciliatum. Z. Naturforschung C J. Biosci. 2015, 67, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Custódio, L.; Laukaityte, S.; Engelen, A.H.; João Rodrigues, M.; Pereira, H.; Vizetto-Duarte, C.; Barreira, L.; Rodríguez, H.; Alberício, F. Comparative Evaluation of Biological Activities and Bioactive Compounds of the Seagrasses Zostera marina and Zostera noltei from Southern Portugal. Nat. Prod. Res. 2016, 30, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Hu, J.; Hu, D.; Yang, X. A Role of Gallic Acid in Oxidative Damage Diseases: A Comprehensive Review. Nat. Prod. Commun. 2019, 14, 1934578X19874174. [Google Scholar] [CrossRef]
- Kim, D.H.; Mahomoodally, M.F.; Sadeer, N.B.; Seok, P.G.; Zengin, G.; Palaniveloo, K.; Khalil, A.A.; Rauf, A.; Rengasamy, K.R.R. Nutritional and bioactive potential of seagrasses: A review. S. Afr. J. Bot. 2021, 137, 216–227. [Google Scholar] [CrossRef]
- Grignon-Dubois, M.; Rezzonico, B. The economic potential of the beach-cast seagrass—Cymodocea nodosa: A pomising renewable source of chicoric acid. Bot. Mar. 2013, 56, 303–311. [Google Scholar] [CrossRef]
- Cuny, P.; Serve, L.; Jupin, H.; Boudouresque, C.-F. Water Soluble Phenolic Compounds of the Marine Phanerogam Posidonia oceanica in a Mediterranean Area Colonised by the Introduced Chlorophyte Caulerpa taxifolia. Aquat. Bot. 1995, 52, 237–242. [Google Scholar] [CrossRef]
- Agostini, S.; Desjobert, J.M.; Pergent, G. Distribution of phenolic compounds in the seagrass Posidonia oceanica. Phytochemistry 1998, 48, 611–617. [Google Scholar] [CrossRef]
- Haznedaroglu, M.Z.; Zeybeky, U. HPLC Determination of Chicoric Acid in Leaves of Posidonia oceanica. Pharm. Biol. 2008, 45, 745–748. [Google Scholar] [CrossRef]
- Subhashini, P.; Dilipan, E.; Thangaradjou, T.; Papenbrock, J. Bioactive Natural Products from Marine Angiosperms: Abundance and Functions. Nat. Prod. Bioprospecting 2013, 3, 129–136. [Google Scholar] [CrossRef]
- Cornara, L.; Pastorino, G.; Borghesi, B.; Salis, A.; Clericuzio, M.; Marchetti, C.; Damonte, G.; Burlando, B. Modulates Cell Activities with Skin Health Applications. Mar. Drugs 2018, 16, 21. [Google Scholar] [CrossRef]
- Heglmeier, A.; Zidorn, C. Secondary metabolites of Posidonia oceanica (Posidoniaceae). Biochem. Syst. Ecol. 2010, 38, 964–970. [Google Scholar] [CrossRef]
- Piazzini, V.; Vasarri, M.; Degl’Innocenti, D.; Guastini, A.; Barletta, E.; Salvatici, M.C.; Bergonzi, M.C. Comparison of Chitosan Nanoparticles and Soluplus Micelles to optimize the Bioactivity of Posidonia oceanica Extract on Human Neuroblastoma Cell Migration. Pharmaceutics 2019, 11, 655. [Google Scholar] [CrossRef] [PubMed]
- Selvimli-Gur, C.; Yesil-Celiktas, O. Cytotoxicity screening of supercritical fluid extracted seaweeds and phenylpropanoids. Mol. Biol. Rep. 2019, 46, 3691–3699. [Google Scholar] [CrossRef] [PubMed]
- Kolsi, R.A.; Fakhfakh, J.; Krichen, F.; Jribi, I.; Chiarore, A.; Patti, F.P.; Blecker, C.; Allouche, N.; Belghith, H.; Belghith, K. Structural characterization and functional properties of antihypertensive Cymodocea nodosa sulfated polysaccharide. Carbohydr. Polym. 2016, 151, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kolsi, R.A.; Salah, H.B.; Saidi, S.A.; Allouche, N.; Belghith, H.; Belghith, K. Evaluation of nutritional value, characteristics, functional properties of Cymodocea nodosa and its benefits on health diseases. Lipids Health Dis. 2017, 16, 238. [Google Scholar] [CrossRef] [PubMed]
- Kolsi, R.B.A.; Gargouri, B.; Sassi, S.; Frikha, D.; Lassoued, S.; Belghith, K. In vitro biological properties and health benefits of the novel sulfated polysaccharide isolated from Cymodocea nodosa. Lipids Health Dis. 2017, 16, 252. [Google Scholar] [CrossRef] [PubMed]
- Smadi, A.; Ciavatta, M.L.; Bitam, F.; Carbone, M.; Villani, G.; Gavagnin, M. Prenylated Flavonoids and Phenolic Compounds from the Rhizomes of Marine Phanerogam Cymodocea Nodosa. Planta Medica 2018, 84, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Milović, S.; Stanković, I.; Nikolić, D.; Radović, J.; Kolundžić, M.; Nikolić, V.; Stanojković, T.; Petović, S. Chemical Analysis of Selected Seaweeds and Seagrass from the Adriatic Coast of Montenegro. Chem. Biodivers. 2019, 16, e1900327. [Google Scholar] [CrossRef]
- Migliore, L.; Rotini, A.; Randazzo, D.; Albanese, N.N.; Giallongo, A. Phenols content and 2-D electrophoresis protein pattern: A promising tool to monitor Posidonia meadows health state. BMC Ecol. 2007, 7, 6. [Google Scholar] [CrossRef]
- Malea, P.; Kevrekidis, T. Trace element (Al, As, B, Ba, Cr, Mo, Ni, Se, Sr, Tl, U and V) distribution and seasonality in compartments of the seagrass Cymodocea nodosa. Sci. Total Environ. 2013, 463–464, 611–623. [Google Scholar] [CrossRef]
- Tutar, O.; Marín-Guirao, L.; Ruizc, J.M.; Procaccini, G. Antioxidant response to heat stress in seagrasses. A gene expression study. Mar. Environ. Res. 2017, 132, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Papathanasiou, V.; Kariofillidou, G.; Malea, P.; Orfanidis, S. Effects of air exposure on desiccation and photosynthetic performance of Cymodocea nodosa with and without epiphytes and Ulva rigida in comparison, under laboratory conditions. Mar. Environ. Res. 2020, 158, 104948. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.; Saiz, L.; Sanmartí, N.; Llagostera, I.; Romero, J. Responses to the simulated herbivory of the seagrass Cymodocea nodosa. In Proceedings of the 10th International Seagrass Biology Workshop ‘12, Rio de Janeiro, Brazil, 25–30 November 2012. [Google Scholar]
- Sureda, A.; Box, A.; Tejada, S. Assessment and physiological state of the Posidonia oceanica meadows in Posto Cristo (Manacor, Spain). Cintenental Shelf Res. 2015, 111, 327–332. [Google Scholar] [CrossRef]
- Pergent-Martini, C.; Pergent, G.; Fernandez, G.; Ferrat, C. Value and use of Posidonia oceanica as a biological indicator. In Proceedings MEDCOAST 99-EMECS 99 Joint Conference “Land-Ocean Interactions: Managing Coastal Ecosystems”, MEDCOAST; Middle East Technical Universiy Publications: Ankara, Turkey, 1999; pp. 73–90. [Google Scholar]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Green, E.P.; Short, F.T. World Atlas of Seagrasses; University of California Press: London, UK, 2003; p. 298. [Google Scholar]
- Gobert, S.; Cambridge, M.L.; Velimirov, B.; Pergrent, G.; Lepoit, G.; Bouquegneau, J.-M.; Daubi, P.; Pergent-Martini, C.; Walker, D.I. Biology of Posidonia. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrect, The Netherlands, 2006; pp. 386–408. [Google Scholar]
- Liu, J.; Tang, X.; Wang, Y.; Zang, Y.; Zhou, B.A. Zostera marina manganese superoxide dismutase gene involved in the responses to temperature stress. Gene 2016, 575, 718–724. [Google Scholar] [CrossRef]
- Trifan, A.; Vasincu, A.; Luca, S.V.; Neophytou, C.; Wolfram, E.; Opitz, S.E.W.; Sava, D.; Bucur, L.; Cioroiu, B.I.; Miron, A.; et al. Unravelling the potential of seaweeds from the Black Sea coast of Romania as bioactive compounds sources. Part I: Cystoseira barbata (Stackhouse) C. Agardh. Food Chem. Toxicol. 2019, 134, 110820. [Google Scholar] [CrossRef]
- De La Fuente, G.; Fontana, M.; Asnaghi, V.; Chiantore, M.; Mirata, S.; Salis, A.; Damonte, G.; Scarfì, S. The Remarkable Antioxidant and Anti-Inflammatory Potential of the Extracts of the Brown Alga Cystoseira amentacea var. stricta. Mar. Drugs 2020, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Sapatinha, M.; Oliveira, A.; Costa, S.; Pedro, S.; Gonçalves, A.; Mendes, R.; Bandarra, N.M.; Pires, C. Red and brown seaweeds extracts: A source of biologically active compounds. Food Chem. 2022, 393, 133453. [Google Scholar] [CrossRef]
- Goutzourelas, N.; Kevrekidis, D.P.; Malea, P.; Barda, S.; Giakountis, A.; Trachana, V.; Savvidi, S.; Kevrekidou, A.; Assimopoulou, A.; Goutas, A.; et al. Free radical scavenging, protection from ROS-induced DNA damage, total polyphenolic content and cytotoxicity against cancer cells of extracts from seaweed species of the Aegean Sea. Foods 2023, 12, 1310. [Google Scholar] [CrossRef]
- Bharathi, N.P.; Jayalakshmi, M.; Amudha, P.; Vanitha, V. Phytochemical screening and in vitro antioxidant activity of the seagrass Cymodocea Serrulata. Indian J. Geo Mar. Sci. 2019, 48, 1216–1221. [Google Scholar]
- Santoso, J.; Anwariyah, S.; Rumiantin, R.O.; Putri, A.P.; Ukhty, N.; Yoshie-Stark, Y. Phenol content, antioxidant activity and fibers profile of four tropical Seagrasses from Indonesia. J. Coast. Dev. 2012, 15, 189–196. [Google Scholar]
- Wisespongpand, P.; Khantavong, A.; Phothong, P.; Wanghom, W. Antimicrobial, Antioxidant, and Antifouling Activity from Extracts of Aboveground and Belowground Parts of Seagrasses Cymodocea rotundata and Cymodocea serrulata. J. Fish. Environ. 2022, 46, 37–53. [Google Scholar]
- Mezghani, S.; Bourguiba, I.; Hfaiedh, I.; Amri, M. Antioxidant potential of Ulva rigida extracts: Protection of HeLa cells against H2O2 cytotoxicity. Biol. Bull. 2013, 225, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological potential of marine macroalgae of the genus Cystoseira. Acta Biol. Hung. 2015, 66, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Güner, A.; Köksal, Ç.; Erel, Ş.B.; Kayalar, H.; Nalbantsoy, A.; Sukatar, A.; Karabay Yavaşoğlu, N.Ü. Antimicrobial and antioxidant activities with acute toxicity, cytotoxicity and mutagenicity of Cystoseira compressa (Esper) Gerloff & Nizamuddin from the coast of Urla (Izmir, Turkey). Cytotechnology 2015, 67, 135–143. [Google Scholar]
- Al-Enazi, N.M.; Awaad, A.S.; Zain, M.E.; Alqasoumi, S.I. Antimicrobial, antioxidant and anticancer activities of Laurencia catarinensis, Laurencia majuscula and Padina pavonica extracts. Saudi Pharm. J. 2018, 26, 44–52. [Google Scholar] [CrossRef]
- Chouh, A.; Nouadri, T.; Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phlorotannins of the Brown Algae Sargassum vulgare from the Mediterranean Sea Coast. Antioxidants 2022, 11, 1055. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in marine algae and their bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef]
- Mateos, R.; Pérez-Correa, J.R.; Domínguez, H. Bioactive Properties of Marine Phenolics. Mar. Drugs 2020, 18, 501. [Google Scholar] [CrossRef]
- Hsu, C.Y. Antioxidant activity of extract from Polygonum aviculare L. Biol. Res. 2006, 39, 281–288. [Google Scholar] [CrossRef]
- Sieg, R.D.; Kubanek, J. Chemical ecology of marine angiosperms: Opportunities at the interface of marine and terrestrial systems. J. Chem. Ecol. 2013, 39, 687–711. [Google Scholar] [CrossRef]
- Murat, K.; Ilkay, O.; Mahmud, A.-A.; Şenol, F.S.; Tahir, A.; Bilge, Ş. Antioxidant and anticholinesterase assets and liquid chromatography-mass spectrometry preface of various fresh-water and marine macroalgae. Pharmacogn. Mag. 2009, 5, 291–297. [Google Scholar]
- Dumay, O.; Costa, J.; Desjobert, J.; Pergent, G. Variations in the concentration of phenolic compounds in the seagrass Posidonia oceanica under conditions of competition. Phytochemistry 2004, 65, 3211–3220. [Google Scholar] [PubMed]
- Rotini, A.; Anello, L.; Di Bernardo, M.; Giallongo, A.; Valiante, L. Comparative analysis of bed density, total phenol content and protein expression pattern in Posidonia oceanica (L.) Delile). Open J. Ecol. 2013, 3, 438–444. [Google Scholar] [CrossRef]
- Rotini, A.; Belmonte, A.; Barrote, I.; Micheli, C.; Peirano, A.; Santos, R.O.; Silva, J.; Migliore, L. Effectiveness and consistency of a suite of descriptors for assessing the ecological status of seagrass meadows (Posidonia oceanica L. Delile). Estuar. Coast. Shelf Sci. 2013, 130, 252–259. [Google Scholar] [CrossRef]
- Boumaza, S.; Boudefoua, N.; Boumasa, R.; Semroud, R. Effects of urban effluents on spatial structure. Morphology and total phenols of Posidonia oceanica: Comparison with a reference site. J. Exp. Mar. Biol. Ecol. 2014, 457, 113–119. [Google Scholar] [CrossRef]
- Levin, D.A. Plant Phenolics: An Ecological Perspective. Am. Nat. 1971, 105, 157–181. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef]
- Francavilla, M.; Franchi, M.; Monteleone, M.; Caroppo, C. The red seaweed Gracilaria gracilis as a multi products source. Mar. Drugs 2013, 11, 3754–3776. [Google Scholar] [CrossRef]
- Caf, F.; Yilmaz, Ö.; Durucan, F.; Şen Özdemir, N. Biochemical components of three marine macroalgae (Padina pavonica, Ulva lactuca and Taonia atomaria) from the Levantine Sea Coast of Antalya, Turkey. J. Bus. Econ. Stat. 2015, 6, 401–411. [Google Scholar]
- Neto, R.T.; Marçal, C.; Queirós, A.S.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef]
- Wekre, M.E.; Kåsin, K.; Underhaug, J.; Holmelid, B.; Jordheim, M. Quantification of Polyphenols in Seaweeds: A Case Study of Ulva intestinalis. Antioxidants 2019, 8, 612. [Google Scholar] [CrossRef] [PubMed]
- Ferrat, L.; Pergent-Martini, C.; Roméo, M. Assessment of the use of biomarkers in aquatic plants for the evaluation of environmental quality: Application to seagrasses. Aquat. Toxicol. 2003, 65, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Bauer, R.; Reminger, P.; Jurcic, K.; Wagner, H. Influence of Echinacea extracts on phagocytic activity. Phytother. Res. 1998, 10, 43–48. [Google Scholar]
- Liu, C.Z.; Abassi, B.H.; Gao, M.; Murch, S.J.; Saxena, P.K. Caffeic acid derivatives production by hairy root cultures of Echinacea purpurea. J. Agric. Food Chem. 2006, 54, 8456–8460. [Google Scholar] [CrossRef] [PubMed]
- Binns, S.E.; Hudson, J.; Merali, S.; Arnason, J.T. Antiviral activity of characterized extracts from Echinacea spp. (Heliantheae: Asteraceae) against Herpes simplex virus (HSV-I). Planta Medica 2002, 68, 780–783. [Google Scholar] [CrossRef]
- Dalby-Brown, L.; Barsett, H.; Landbo, A.K.; Meyer, A.S.; Molgaard, P. Synergistic antioxidative effects of alkamides, caffeic acid derivatives and polysaccharides fractions from Echinacea purpurea on in vitro oxidation of human low density lipoproteins. J. Agric. Food Chem. 2005, 53, 9413–9423. [Google Scholar] [CrossRef] [PubMed]
- Grignon-Dubois, M.; Rezzonico, B. Phenolic fingerprint of the seagrass Posidonia oceanica from four locations in the Mediterranean Sea: First evidence for the large predominance of chicoric acid. Bot. Mar. 2015, 58, 379–391. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chuang, Y.C.; Ku, Y.H. Quantification of bioactive compounds in citrus fruits cultivated in Taiwan. Food Chem. 2006, 10, 1163–1171. [Google Scholar]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2011, 38, 167–197. [Google Scholar] [CrossRef]
- Leri, M.; Ramazzotti, M.; Vasarri, M.; Peri, S.; Barletta, E.; Pretti, C.; Degl’Innocenti, D. Bioactive compounds from Posidonia oceanica (L.) Delile Impair Malignant Cell Migration through Autophagy Modulation. Mar. Drugs 2018, 16, 137. [Google Scholar] [CrossRef]
- Kontiza, I.; Vagias, C.; Jakupovic, J.; Moreau, D.; Roussakis, C.; Roussis, V. Cymodienol and Cymodiene: New Cytotoxic Diarylheptanoids from the Seagrass Cymodocea nodosa. Tetrahedron Lett. 2005, 46, 2845–2847. [Google Scholar] [CrossRef]
- Abdelhameed, R.; Habib, E.; Goda, M.; Fahim, J.R.; Hassanean, H.; Eltamany, E.; Al-Warhi, T.; Bringmann, G.; Ahmed, S.A.; Abdelmohsen, U.R. Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum. Mar. Drugs 2020, 18, 354. [Google Scholar] [CrossRef]
- Hernández-Balmaseda, I.; Guerra, I.R.; Declerck, K.; Herrera Isidrón, J.A.; Pérez-Novo, C.; Van Camp, G.; DeWever, O.; González, K.; Labrada, M.; Carr, A.; et al. Marine Seagrass Extract of Thalassia testudinum Suppresses Colorectal Tumor Growth, Motility and Angiogenesis by Autophagic Stress and Immunogenic Cell Death Pathways. Mar. Drugs 2021, 19, 52. [Google Scholar] [CrossRef]
- Vasarri, M.; Barletta, E.; Degl’Innocenti, D. Posidonia oceanica (L.) Delile Extract Reduces Lipid Accumulation through Autophagy Activation in HepG2 Cells. Pharmaceuticals 2021, 14, 969. [Google Scholar] [CrossRef]
- Lazli, W.; Hellal, A. Biosorption of phenol by modified dead leaves of Posidonia oceanica immobilized in calcium alginate beads: Optimal experimental parameters using central composite design. Water SA 2020, 46, 322–329. [Google Scholar]
- Harrison, P.G. Detrital processing in seagrass systems: A review of factors affecting decay rates, remineralization and detritivory. Aquat. Bot. 1989, 23, 263–288. [Google Scholar] [CrossRef]
- Achamlale, S.; Rezzonico, B.; Grignon-Dubois, M. Evaluation of Zostera detritus as a potential new source of zosteric acid. J. Appl. Phycol. 2009, 21, 347–352. [Google Scholar] [CrossRef]
- Cariello, L.; Zanetti, L.; De Stefano, S. Posidonia ecosystem-V: Phenolic compounds from marine phanerogames, Cymodocea nodosa and Posidonia oceanica. Comp. Biochem. Physiol. 1979, 62, 159–161. [Google Scholar] [CrossRef]
- ACCOLAGOONS and NATURA 2000. Accolaggons-Actions for the Conservation of Coastal Habitats and Significant Avifauna Species in Natura 2000; Action A2: Network Sites of Epanomi and Aggelochori Lagoons, Greece. 09 NAT/GR/000343; Life: Thessaloniki, Greece, 2012; p. 60. [Google Scholar]
- Boudouresque, C.F.; Bernard, G.; Bonhomme, P.; Charbonnel, E.; Diviacco, G.; Meinesz, A.; Pergent, G.; Pergent-Martini, C.; Ruitto, N.S.T. Protection and Conservation of Posidonia oceanica Meadows; RAMOGE, RAC/SPA and GIS Posidonie Publications: Marseilles, Framce, 2012; p. 202. [Google Scholar]
- Mylona, Z.; Panteris, E.; Moustakas, M.; Kevrekidis, T.; Malea, P. Physiological, structural and ultrastructural impacts of silver nanoparticles on the seagrass Cymodocea nodosa. Chemosphere 2020, 248, 126066. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and sensitive method for the quantitation microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.F.J.R.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Apostolou, A.; Stagos, D.; Galitsiou, E.; Spyrou, A.; Haroutounian, S.; Portesis, N.; Trizoglou, I.; Wallace Hayes, A.; Tsatsakis, A.M.; Kouretas, D. Assessment of polyphenolic content, antioxidant activity, protection against ROS-induced DNA damage and anticancer activity of Vitis vinifera stem extracts. Food Chem. Toxicoogy 2013, 61, 60–68. [Google Scholar] [CrossRef]
- Kerasioti, E.; Stagos, D.; Priftis, A.; Aivazidis, S.; Tsatsakis, A.M.; Hayes, A.W.; Kouretas, D. Antioxidant effects of whey protein on muscle C2C12 cells. Food Chem. 2014, 155, 271–278. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Living Leaves (LP) (5000 mg L−1) | ||||
|---|---|---|---|---|
| Phenolic Compounds | Phenolic Compounds (mg) | Percentage (%) of Phenolic Compounds (mg)/Polyphenols (mg) | Percentage (%) of Phenolic Compounds (mg)/Sample (mg) | Elution Time (min) |
| Caftaric acid | 0.014 | 1.176 | 0.162 | 4.66 |
| Chicoric acid | 0.625 | 51.229 | 7.059 | 12.50 |
| Quercetin | - | - | - | - |
| Caffeic acid | - | - | - | - |
| (-)-Epigallocatechin gallate | 0.348 | 28.542 | 3.933 | 10.46 |
| p-Coumaric acid | 0.170 | 13.903 | 1.916 | 11.87 |
| trans-Ferulic acid | 0.008 * | 0.666 * | 0.092 * | 13.07 |
| Sinapinic acid | 0.017 | 1.371 | 0.189 | 13.59 |
| Rutin hydrate | 0.035 | 2.836 | 0.391 | 16.98 |
| trans-Cinnamic acid | 0.003 * | 0.277 * | 0.038 * | 20.77 |
| Hesperidin | - | - | - | - |
| 4′,5,7-Trihydroxyflavone | - | - | - | - |
| Total phenolic content (mg) | 1.221 | 13.781 | ||
| Amount of samples (mg) | 8.86 | |||
| Dry ‘Necromass’ (DNP) (7000 mg L−1) | ||||
| Caftaric acid | - | - | - | - |
| Chicoric acid | - | - | - | - |
| Quercetin | - | - | - | - |
| Caffeic acid | T | T | T | 9.55 |
| (-)-Epigallocatechin gallate | 0.010 * | 59.377 * | 0.097 * | 10.44 |
| p-Coumaric acid | - | - | - | - |
| trans-Ferulic acid | - | - | - | - |
| Sinapinic acid | - | - | - | - |
| Rutin hydrate | - | - | - | |
| trans-Cinnamic acid | 0.003 * | 15.335 * | 0.025 * | 20.78 |
| Hesperidin | - | - | - | - |
| 4′,5,7-Trihydroxyflavone | - | - | - | - |
| Total phenolic content (mg) | 0.018 | 0.168 | ||
| Amount of samples (mg) | 10.74 | |||
| r | n | |
|---|---|---|
| APX–SOD activity | 0.895 *** | 12 |
| APX–protein content | −0.930 *** | 12 |
| SOD–protein content | −0.951 *** | 12 |
| APX–CTCF value | −0.710 * | 12 |
| SOD–CTCF value | −0.621 * | 12 |
| protein content–CTCF value | 0.680 * | 12 |
| DPPH•–ABTS•+ | 0.783 * | 9 |
| DPPH•–OH• | 0.943 ** | 6 |
| DPPH•–O2•− | 0.917 ** | 9 |
| DPPH•–RP0.5AU | 0.870 ** | 9 |
| ABTS•+–OH• | 0.600 ns | 6 |
| ABTS•+–O2•− | 0.850 ** | 9 |
| ABTS•+–RP0.5AU | 0.812 ** | 9 |
| OH•–O2•− | 0.657 ns | 6 |
| OH•–RP0.5AU | 0.638 ns | 6 |
| O2•−–RP0.5AU | 0.879 ** | 9 |
| DPPH•–phenolic content | −0.867 ** | 9 |
| ABTS•+–phenolic content | −0.600 ns | 9 |
| OH•–phenolic content | −1.000 *** | 6 |
| O2•c–phenolic content | −0.683 * | 9 |
| RP–phenolic content | −0.812 ** | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kevrekidou, A.; Assimopoulou, A.N.; Trachana, V.; Stagos, D.; Malea, P. Antioxidant Activity, Inhibition of Intestinal Cancer Cell Growth and Polyphenolic Compounds of the Seagrass Posidonia oceanica’s Extracts from Living Plants and Beach Casts. Mar. Drugs 2024, 22, 130. https://doi.org/10.3390/md22030130
Kevrekidou A, Assimopoulou AN, Trachana V, Stagos D, Malea P. Antioxidant Activity, Inhibition of Intestinal Cancer Cell Growth and Polyphenolic Compounds of the Seagrass Posidonia oceanica’s Extracts from Living Plants and Beach Casts. Marine Drugs. 2024; 22(3):130. https://doi.org/10.3390/md22030130
Chicago/Turabian StyleKevrekidou, Alkistis, Andreana N. Assimopoulou, Varvara Trachana, Dimitrios Stagos, and Paraskevi Malea. 2024. "Antioxidant Activity, Inhibition of Intestinal Cancer Cell Growth and Polyphenolic Compounds of the Seagrass Posidonia oceanica’s Extracts from Living Plants and Beach Casts" Marine Drugs 22, no. 3: 130. https://doi.org/10.3390/md22030130
APA StyleKevrekidou, A., Assimopoulou, A. N., Trachana, V., Stagos, D., & Malea, P. (2024). Antioxidant Activity, Inhibition of Intestinal Cancer Cell Growth and Polyphenolic Compounds of the Seagrass Posidonia oceanica’s Extracts from Living Plants and Beach Casts. Marine Drugs, 22(3), 130. https://doi.org/10.3390/md22030130