
Towards the Exploration and Evolution of Insulin-like Venoms in Actiniaria (Sea anemones)


Abstract
:1. Introduction
2. Results
2.1. Identification
2.2. Phylogenomics
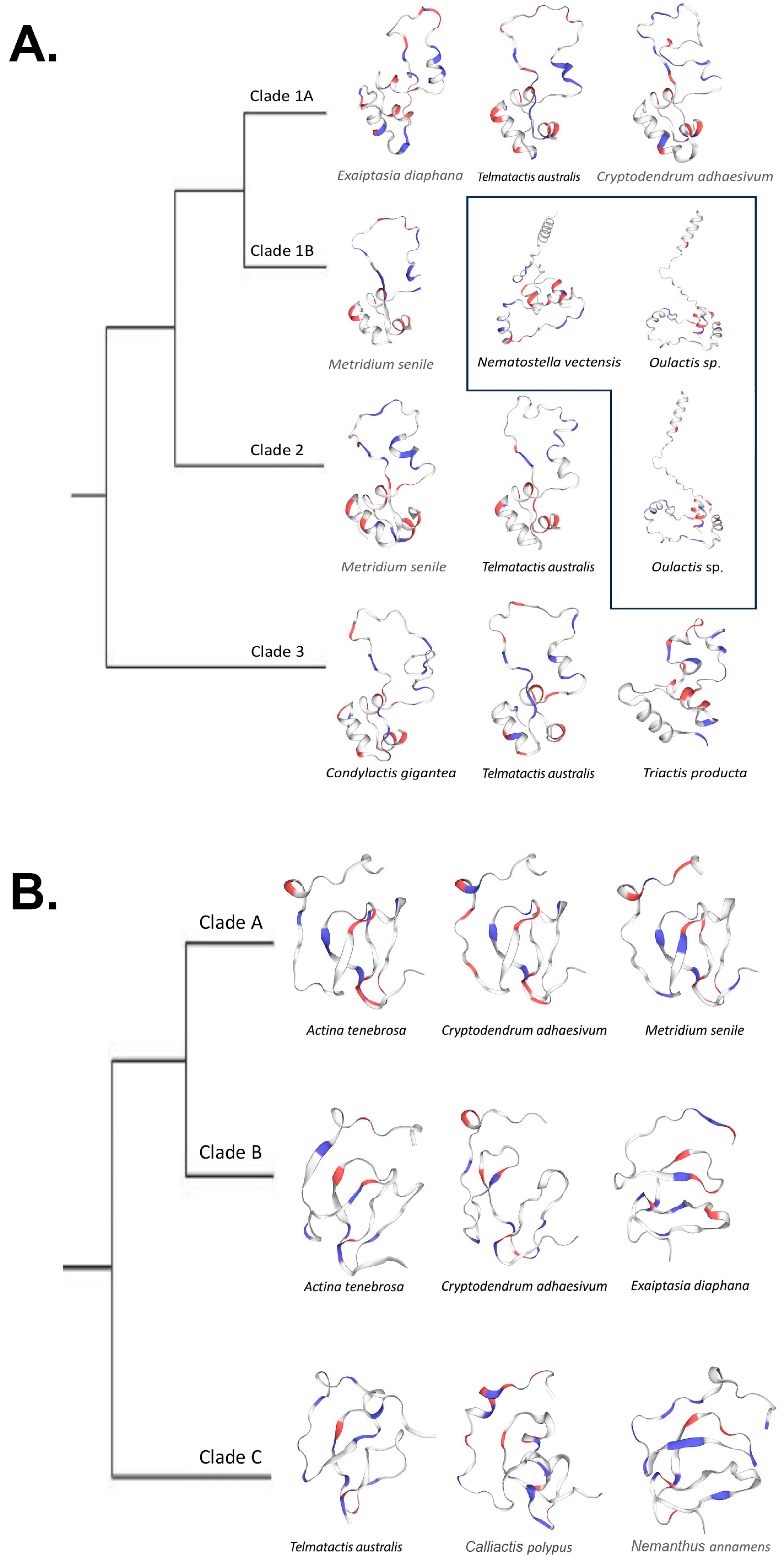
2.3. Predicted Protein Structure
2.4. Sites under Selection
3. Discussion
3.1. Insulin-like Venoms in Sea anemones
3.2. Phylogenetics
3.3. Alternative Splicing
3.4. Structural Predictions
3.5. Selection Analysis
3.6. Sites under Selection—A Scaling Issue?
4. Materials and Methods
4.1. Annotation/Identification
4.2. Phylogenomic Analyses
4.3. Structural Predictions
4.4. Mode of Selecetion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Muscatine, L. Coelenterate Biology: Reviews and New Perspectives; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Chintiroglou, C.C.; Doumenc, D.; Zamponi, M. Commented list of the Mediterranean Actiniaria and Corallimorpharia (Anthozoa). Acta Adriat. 1997, 38, 65–70. [Google Scholar]
- Daly, M. Functional and genetic diversity of toxins in sea anemones. In Evolution of Venomous Animals and Their Toxins; Malhotra, A., Gopalakrishnakone, P., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 87–104. [Google Scholar]
- Madio, B.; Undheim, E.A.B.; King, G.F. Revisiting venom of the sea anemone Stichodactyla haddoni: Omics techniques reveal the complete toxin arsenal of a well-studied sea anemone genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef]
- Macrander, J.; Broe, M.; Daly, M. Tissue-specific venom composition and differential gene expression in sea anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef]
- Ashwood, L.M.; Norton, R.S.; Undheim, E.A.B.; Hurwood, D.A.; Prentis, P.J. Characterising functional venom profiles of anthozoans and medusozoans within their ecological context. Mar. Drugs 2020, 18, 202. [Google Scholar] [CrossRef]
- Ashwood, L.M.; Undheim, E.A.; Madio, B.; Hamilton, B.R.; Daly, M.; Hurwood, D.A.; King, G.F.; Prentis, P.J. Venoms for all occasions: The functional toxin profiles of different anatomical regions in sea anemones are related to their ecological function. Mol. Ecol. 2022, 31, 866–883. [Google Scholar] [CrossRef]
- Delgado, A.; Benedict, C.; Macrander, J.; Daly, M. Never, Ever make an enemy… out of an anemone: Transcriptomic comparison of clownfish hosting sea anemone venoms. Mar. Drugs 2022, 20, 730. [Google Scholar] [CrossRef]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea anemone (Cnidaria, Anthozoa, Actiniaria) toxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef]
- Menezes, C.; Thakur, N.L. Sea anemone venom: Ecological interactions and bioactive potential. Toxicon 2022, 208, 31–46. [Google Scholar] [CrossRef]
- Smith, E.G.; Surm, J.M.; Macrander, J.; Simhi, A.; Amir, G.; Sachkova, M.Y.; Lewandowska, M.; Reitzel, A.M.; Moran, Y. Micro and macroevolution of sea anemone venom phenotype. Nat. Commun. 2023, 14, 249. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.C.; Konaté, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or normalized counts? A comparative study of quantification measures for the analysis of rna-seq data from the nci patient-derived models repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef]
- Sunagar, K.; Moran, Y. The rise and fall of an evolutionary innovation: Contrasting strategies of venom evolution in ancient and young animals. PLoS Genet. 2015, 11, e1005596. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Sunagar, K.; Federman Gross, A.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an ancient venom: Recognition of a novel family of cnidarian toxins and the common evolutionary origin of sodium and potassium neurotoxins in sea anemones. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef]
- Gibbs, H.L.; Rossiter, W. Rapid Evolution by positive selection and gene gain and loss: PLA2 venom genes in closely related Sistrurus rattlesnakes with divergent diets. J. Mol. Evol. 2008, 66, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Modica, M.V.; Gorson, J.; Fedosov, A.E.; Malcolm, G.; Terryn, Y.; Puillandre, N.; Holford, M. Macroevolutionary analyses suggest that environmental factors, not venom apparatus, play key role in terebridae marine snail diversification. Syst. Biol. 2020, 69, 413–430. [Google Scholar] [CrossRef]
- Binford, G.; Wells, M. The phylogenetic distribution of sphingomyelinase D activity in venoms of Haplogyne spiders. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 135, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, A.M.; Rodríguez, E.; Faircloth, B.C.; Cowman, P.F.; Brugler, M.R.; Farfan, G.A.; Hellberg, M.E.; Kitahara, M.V.; Morrison, C.L.; Paz-García, D.A.; et al. Palaeoclimate ocean conditions shaped the evolution of corals and their skeletons through deep time. Nat. Ecol. Evol. 2020, 4, 1531–1538. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Macrander, J.; Surm, J.M.; Aharoni, R.; Menard-Harvey, S.S.; Klock, A.; Leach, W.B.; Reitzel, A.M.; Moran, Y. Some like it hot: Population-specific adaptations in venom production to abiotic stressors in a widely distributed cnidarian. BMC Biol. 2020, 18, 121. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Singer, S.A.; Macrander, J.; Reitzel, A.M.; Peigneur, S.; Tytgat, J.; Moran, Y. The birth and death of toxins with distinct functions: A case study in the sea anemone Nematostella. Mol. Biol. Evol. 2019, 36, 2001–2012. [Google Scholar] [CrossRef]
- Prentis, J.; Pavasovic, A.P.; Norton, R.S. Sea anemones: Quiet achievers in the field of peptide toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef]
- Escoubas, P.; King, G.F. Venomics as a drug discovery platform. Expert Rev. Proteom. 2009, 6, 221–224. [Google Scholar] [CrossRef]
- Beeton, C.; Pennington, M.W.; Norton, R.S. Analogs of the sea anemone potassium channel blocker ShK for the treatment of autoimmune diseases. Inflamm. Allergy Drug Targets 2011, 10, 313–321. [Google Scholar] [CrossRef]
- Pennington, M.W.; Lanigan, M.D.; Kalman, K.; Mahnir, V.M.; Rauer, H.; McVaugh, C.T.; Behm, D.; Donaldson, D.; Chandy, K.G.; Kem, W.R.; et al. Role of disulfide bonds in the structure and potassium channel blocking activity of ShK toxin. Biochemistry 1999, 38, 14549–14558. [Google Scholar] [CrossRef]
- Tysoe, C.; Withers, S.G. Structural dissection of Helianthamide reveals the basis of its potent inhibition of human pancreatic α-Amylase. Biochemistry 2018, 57, 5384–5387. [Google Scholar] [CrossRef]
- Sintsova, O.; Gladkikh, I.; Kalinovskii, A.; Zelepuga, E.; Monastyrnaya, M.; Kim, N.; Shevchenko, L.; Peigneur, S.; Tytgat, J.; Kozlovskaya, E.; et al. Magnificamide, a β-Defensin-like peptide from the mucus of the sea anemone Heteractis magnifica, is a strong inhibitor of mammalian α-Amylases. Mar. Drugs 2019, 17, 542. [Google Scholar] [CrossRef]
- D’Ambra, I.; Lauritano, C. A review of toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.M. Evolution of the mammalian insulin (Ins) gene; changes in proteolytic processing. Peptides 2021, 135, 170435. [Google Scholar] [CrossRef]
- Rotwein, P. Diversification of the insulin-like growth factor 1 gene in mammals. PLoS ONE 2017, 12, e0189642. [Google Scholar] [CrossRef] [PubMed]
- Chowański, S.; Walkowiak-Nowicka, K.; Winkiel, M.; Marciniak, P.; Urbański, A.; Pacholska-Bogalska, J. Insulin-like peptides and cross-talk with other factors in the regulation of insect metabolism. Front. Physiol. 2021, 12, 701203. [Google Scholar] [CrossRef] [PubMed]
- Semaniuk, U.; Piskovatska, V.; Strilbytska, O.; Strutynska, T.; Burdyliuk, N.; Vaiserman, A.; Bubalo, V.; Storey, K.B.; Lushchak, O. Drosophila insulin-like peptides: From expression to functions—A review. Entomol. Exp. Appl. 2021, 169, 195–208. [Google Scholar] [CrossRef]
- Wu, Q.; Brown, M.R. Signaling and function of insulin-like peptides in insects. Annu. Rev. Entomol. 2006, 51, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Chiu, H.; Boudreau, J.; Papanicolaou, T.; Bendena, W.; Chin-Sang, I. A functional study of all 40 Caenorhabditis elegans insulin-like peptides. J. Biol. Chem. 2018, 293, 16912–16922. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.B.; Van Kesteren, R.E.; Li, K.W.; Van Minnen, J.; Spijker, S.; Van Heerikhuizen, H.; Geraerts, W.P.M. Towards understanding the role of insulin in the brain: Lessons from insulin-related signaling systems in the invertebrate brain. Prog. Neurobiol. 1998, 54, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.K.; El-kotby, D.A.; Tawfik, M.M.; Badr, R.E.; Bahgat, I.M. Antidiabetic effect of the Egyptian honey bee (Apis mellifera) venom in alloxan-induced diabetic rats. J. Basic Appl. Zool. 2019, 80, 58. [Google Scholar] [CrossRef]
- Wong, E.S.; Morgenstern, D.; Mofiz, E.; Gombert, S.; Morris, K.M.; Temple-Smith, P.; Renfree, M.B.; Whittington, C.M.; King, G.F.; Warren, W.C.; et al. Proteomics and deep sequencing comparison of seasonally active venom glands in the platypus reveals novel venom peptides and distinct expression profiles. Mol. Cell. Proteom. 2012, 11, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.S.W.; Nicol, S.; Warren, W.C.; Belov, K. Echidna venom gland transcriptome provides insights into the evolution of monotreme venom. PLoS ONE 2013, 8, e79092. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Attoub, S.; Musale, V.; Leprince, J.; Casewell, N.R.; Sanz, L.; Calvete, J.J. Isolation and characterization of cytotoxic and insulin-releasing components from the venom of the black-necked spitting cobra Naja nigricollis (Elapidae). Toxicon X 2020, 6, 100030. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, F.H.R.; Aparicio, R.; dos Santos, M.L.; Diz Filho, E.B.S.; Oliveira, S.C.B.; Toyama, D.O.; Toyama, M.H. A Catalytically Inactive Lys49 PLA2 Isoform from Bothrops jararacussu venom that stimulates insulin secretion in pancreatic beta cells. Protein Pept. Lett. 2011, 18, 1133–1139. [Google Scholar] [CrossRef]
- Furman, B.L. The development of Byetta (exenatide) from the venom of the Gila monster as an anti-diabetic agent. Toxicon 2012, 59, 464–471. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Gajewiak, J.; Karanth, S.; Robinson, S.D.; Ueberheide, B.; Douglass, A.D.; Schlegel, A.; Imperial, J.S.; Watkins, M.; Bandyopadhyay, P.K.; et al. Specialized insulin is used for chemical warfare by fish-hunting cone snails. Proc. Natl. Acad. Sci. USA 2015, 112, 1743–1748. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Hu, H.; Gorasia, D.G.; Bandyopadhyay, P.K.; Veith, P.D.; Young, N.D.; Reynolds, E.C.; Yandell, M.; Olivera, B.M.; Purcell, A.W. Combined proteomic and transcriptomic interrogation of the venom gland of Conus geographus uncovers novel components and functional compartmentalization. Mol. Cell. Proteom. 2014, 13, 938–953. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Safavi-Hemami, H. Insulin as a weapon. Toxicon 2016, 123, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.L.; Hossain, M.A.; Lin, F.; Pinheiro-Junior, E.L.; Peigneur, S.; Wai, D.C.; Delaine, C.; Blyth, A.J.; Forbes, B.E.; Tytgat, J.; et al. Identification, synthesis, conformation and activity of an insulin-like peptide from a sea anemone. Biomolecules 2021, 11, 1785. [Google Scholar] [CrossRef]
- Fu, J.; He, Y.; Peng, C.; Tang, T.; Jin, A.; Liao, Y.; Shi, Q.; Gao, B. Transcriptome Sequencing of the pale anemone (Exaiptasia diaphana) revealed functional peptide gene resources of sea anemone. Front. Mar. Sci. 2022, 9, 856501. [Google Scholar] [CrossRef]
- Darling, J.A.; Reitzel, A.R.; Burton, P.M.; Mazza, M.E.; Ryan, J.F.; Sullivan, J.C.; Finnerty, J.R. Rising starlet: The starlet sea anemone, Nematostella vectensis. BioEssays 2005, 27, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Ruiming, Z.; Yibao, M.; Yawen, H.; Zhiyong, D.; Yingliang, W.; Zhijian, C.; Wenxin, L. Comparative venom gland transcriptome analysis of the scorpion Lychas mucronatus reveals intraspecific toxic gene diversity and new venomous components. BMC Genom. 2010, 11, 452. [Google Scholar] [CrossRef]
- Klompen, A.M.L.; Macrander, J.; Reitzel, A.M.; Stampar, S.N. transcriptomic analysis of four cerianthid (Cnidaria, Ceriantharia) venoms. Mar. Drugs 2020, 18, 413. [Google Scholar] [CrossRef]
- Ja, C. The dynamic genome of Hydra. Nature 2010, 464, 592–596. [Google Scholar]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef]
- Norton, R.S.; Olivera, B.M. Conotoxins down under. Toxicon 2006, 48, 780–798. [Google Scholar] [CrossRef]
- Ogawa, T.; Oda-Ueda, N.; Hisata, K.; Nakamura, H.; Chijiwa, T.; Hattori, S.; Isomoto, A.; Yugeta, H.; Yamasaki, S.; Fukumaki, Y.; et al. Alternative mRNA Splicing in three venom families underlying a possible production of divergent venom proteins of the habu snake, Protobothrops flavoviridis. Toxins 2019, 11, 581. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; He, C.; Yang, Y.; Sun, Y.H.; Xiong, S.; Chan, K.C.; Si, Y.; Xiao, S.; Zhao, X.; Lin, H.; et al. Comprehensive isoform-level analysis reveals the contribution of alternative isoforms to venom evolution and repertoire diversity. Genome Res. 2023, 33, 1554–1567. [Google Scholar] [CrossRef] [PubMed]
- Haney, R.A.; Matte, T.; Forsyth, F.S.; Garb, J.E. Alternative transcription at venom genes and its role as a complementary mechanism for the generation of venom complexity in the common house spider. Front. Ecol. Evol. 2019, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, I.W.; McArdle, A.J.; Coulson, R.M.; Wagner, M.A.; Schmid, R.; Hirai, H.; Hoffmann, K.F. Developmentally regulated expression, alternative splicing and distinct sub-groupings in members of the Schistosoma mansoni venom allergen-like (SmVAL) gene family. BMC Genom. 2008, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Zhijian, C.; Feng, L.; Yingliang, W.; Xin, M.; Wenxin, L. Genetic mechanisms of scorpion venom peptide diversification. Toxicon 2006, 47, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Viala, V.L.; Hildebrand, D.; Trusch, M.; Fucase, T.M.; Sciani, J.M.; Pimenta, D.C.; Arni, R.K.; Schlüter, H.; Betzel, C.; Mirtschin, P. Venomics of the Australian eastern brown snake (Pseudonaja textilis): Detection of new venom proteins and splicing variants. Toxicon 2015, 107, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, L.M.; Elnahriry, K.A.; Stewart, Z.K.; Shafee, T.; Naseem, M.U.; Szanto, T.G.; van der Burg, C.A.; Smith, H.L.; Surm, J.M.; Undheim, E.A.; et al. Genomic, functional and structural analyses elucidate evolutionary innovation within the sea anemone 8 toxin family. BMC Biol. 2023, 21, 121. [Google Scholar] [CrossRef] [PubMed]
- Heldenbrand, J.; Ren, Y.; Asmann, Y.; Mainzer, L.S. Step-by-step guide for downloading very large datasets to a supercomputer using the SRA Toolkit. 2017, pp. 1–14. Available online: https://www.protocols.io/view/step-by-step-guide-for-downloading-very-large-data-kb6csre.html (accessed on 17 November 2023).
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Mahram, A.; Herbordt, M.C. NCBI BLASTP on High-Performance Reconfigurable Computing Systems. ACM Trans. Reconfigurable Technol. Syst. 2015, 7, 33. [Google Scholar] [CrossRef]
- Reeks, T.A.; Fry, B.G.; Alewood, P.F. Privileged frameworks from snake venom. Cell. Mol. Life Sci. 2015, 72, 1939–1958. [Google Scholar] [CrossRef] [PubMed]
- Tamaoki, H.; Miura, R.; Kusunoki, M.; Kyogoku, Y.; Kobayashi, Y.; Moroder, L. Folding motifs induced and stabilized by distinct cystine frameworks. Protein Eng. Des. Sel. 1998, 11, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Cao, Z.; Li, W.; Wu, Y. Cloning and characterization of a novel Kunitz-type inhibitor from scorpion with unique cysteine framework. Toxicon 2013, 72, 5–10. [Google Scholar] [CrossRef]
- Fainzilber, M.; Nakamura, T.; Gaathon, A.; Lodder, J.C.; Kits, K.S.; Burlingame, A.L.; Zlotkin, E. A new cysteine framework in sodium channel blocking conotoxins. Biochemistry 1995, 34, 8649–8656. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega. Curr. Protoc. Bioinform. 2014, 48, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Baumgarten, S.; Simakov, O.; Esherick, L.Y.; Liew, Y.J.; Lehnert, E.M.; Michell, C.T.; Li, Y.; Hambleton, E.A.; Guse, A.; Oates, M.E.; et al. The genome of Aiptasia, a sea anemone model for coral symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 11893–11898. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Kiefer, F.; Arnold, K.; Künzli, M.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository and associated resources. Nucleic Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Bienert, S.; Waterhouse, A.; De Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository—New features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting individual sites subject to episodic diversifying selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A Fast, Unconstrained Bayesian AppRoximation for Inferring Selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef]
- Weaver, S.; Shank, S.D.; Spielman, S.J.; Li, M.; Muse, S.V.; Kosakovsky Pond, S.L. Datamonkey 2.0: A modern web application for characterizing selective and other evolutionary processes. Mol. Biol. Evol. 2018, 35, 773–777. [Google Scholar] [CrossRef]
- Delport, W.; Poon, A.F.Y.; Frost, S.D.W.; Kosakovsky Pond, S.L. Datamonkey 2010: A suite of phylogenetic analysis tools for evolutionary biology. Bioinformatics 2010, 26, 2455–2457. [Google Scholar] [CrossRef]
- Kosakovsky Pond, S.L.; Frost, S.D.W. Not so different after all: A comparison of methods for detecting amino acid sites under selection. Mol. Biol. Evol. 2005, 22, 1208–1222. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cnidoinsulin | VP302 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Clades | Clades | ||||||||
| Superfamily | Family | Species | 1a | 1b | 2 | 3 | A | B | C |
| Actinioidea | Actiniidae | Actinia equina | |||||||
| Actinia tenebrosa | |||||||||
| Anthopleura buddemeieri | |||||||||
| Aulactinia veratra | |||||||||
| Cnidopus japonicus | |||||||||
| Condylactis gigantea | |||||||||
| Dofleinia armata | |||||||||
| Entacmaea quadricolor | |||||||||
| Epiactis prolifera | |||||||||
| Oulactis sp. | |||||||||
| Macrodactyla doreensis | |||||||||
| Actinodendridae | Actinodendron plumosum | ||||||||
| Heteractidae | Heteractis crispa | ||||||||
| Stichodactylidae | Stichodactyla haddoni | ||||||||
| Thalassianthidae | Cryptodendrum adhaesivum | ||||||||
| Heterodactyla hemprichi | |||||||||
| Actinostoloidea | Actinostolidae | Stomphia coccinea | |||||||
| Edwardsioidea | Edwardsiidae | Edwardsiella carnea | |||||||
| Nematostella vectensis | |||||||||
| Metridioidea (Cuticulate) | Andvakiidae | Telmatactis australis | |||||||
| Hormathiidae | Caliactis polypus | ||||||||
| Nemanthidae | Nemanthus annamensis | ||||||||
| Metridioidea (Acuticulate) | Aiptasiidae | Aiptasiogeton eruptaurantia | |||||||
| Bartholomea annulata | |||||||||
| Bellactis lux | |||||||||
| Exaiptasia diaphana | |||||||||
| Neoaiptasia morbilla | |||||||||
| Aliciidae | Lebrunia danae | ||||||||
| Triactis producta | |||||||||
| Boloceroididae | Bunodeopsis globulifera | ||||||||
| Diadumenidae | Diadumene leucolena | ||||||||
| Diadumene lineata | |||||||||
| Metridiidae | Metridium senile | ||||||||
| Sagartiidae | Sagartia elegans | ||||||||
| Clade | Genes Recovered within Isoforms | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Superfamily | Family | Species | A | B | C | VP302 | Kazal | Antistasin | KunitzBPTI |
| Edwardsioidea | Edwardsiidae | Nematostella vectensis | x | ||||||
| Actinioidea | Actiniidae | Actinia tenebrosa | x | x | x | x | |||
| Cnidopus japonicus | x | ||||||||
| Condylactis gigantea | x | ||||||||
| Dofleinia armata | x | ||||||||
| Entacmaea quadricolor | x | x | |||||||
| Thalassianthidae | Cryptodendrum adhaesivum | x | x | x | |||||
| Stichodactylidae | Stichodactyla haddoni | x | |||||||
| Metridioidea | Aiptasiidae | Aiptasiogeton eruptaurantia | x | x | x | x | |||
| Bartholomea annulata | x | x | x | x | |||||
| Bellactis lux | x | x | |||||||
| Exaiptasia diaphana | x | x | x | x | |||||
| Neoaiptasia morbilla | x | x | x | x | |||||
| Aliciidae | Triactis producta | x | x | x | x | ||||
| Boloceroididae | Bunodeopsis globulifera | x | x | x | x | ||||
| Diadumenidae | Diadumene leucolena | x | x | x | x | ||||
| Diadumene lineata | x | x | x | ||||||
| Metridiidae | Metridium senile | x | x | x | x | ||||
| Sagartiidae | Sagartia elegans | x | x | x | x | ||||
| Andvakiidae | Telmatactis australis | x | |||||||
| Hormathiidae | Calliactis polypus | x | |||||||
| Nemanthidae | Nemanthus annamensis | x | |||||||
| Venom | CLADE | Species | GMQE | Template | Seq Identity % | Coverage % |
|---|---|---|---|---|---|---|
| Cnidoinsulin | Clade 1a | Cryptodendrum adhaesivum | 0.3 | 2kqp.1.A Insulin | 19 | 61 |
| Exaiptasia diaphana | 0.37 | 2kqp.1.A Insulin | 28 | 71 | ||
| Telmatactis sp. | 0.25 | 3kr3.1.A Insulin-like growth factor II | 35.7 | 47 | ||
| Clade 1b | Metridium senile | 0.48 | 2kqp.1.A Insulin | 27 | 64 | |
| Nematostella vectensis | 0.66 | AlphaFold DB model of A7S6C3_NEMVE | 100 | 62 | ||
| Oulactis sp. | 0.63 | AlphaFold DB model of A0A6P8J5R7_ACTTE | 83.19 | 98 | ||
| Clade 2 | Metridium senile | 0.32 | 5l3m.1.A Insulin-like growth factor II | 26.67 | 61 | |
| Oulactis sp. | 0.63 | AlphaFold DB model of A0A6P8J5R7_ACTTE | 84.96 | 98 | ||
| Telmatactis sp. | 0.27 | 2kqp.1.A Insulin | 23 | 32 | ||
| Clade 3 | Condylactis gigantea | 0.32 | 5l3m.1.A Insulin-like growth factor II | 22.03 | 66 | |
| Telmatactis sp. | 0.26 | 7u23.1.C single-chain LCDV-1 viral insulin-like peptide | 22.41 | 51 | ||
| Triactis producta | 0.31 | 1h02.1.A INSULIN-LIKE GROWTH FACTOR I | 23.33 | 57 | ||
| VP302 | Clade A | Actina tenebrosa | 0.65 | 3tjq.1.A Serine protease HTRA1 | 45.28 | 91 |
| Cryptodendrum adhaesivum | 0.61 | 3tjq.1.A Serine protease HTRA1 | 44.4 | 92 | ||
| Metridium senile | 0.65 | 3tjq.1.A Serine protease HTRA1 | 45.28 | 91 | ||
| Clade B | Actinia tenebrosa | 0.54 | 3tjq.1.A Serine protease HTRA1 | 38.64 | 0.43 | |
| Exaiptasia diaphana | 0.57 | 3tjq.1.A Serine protease HTRA1 | 43.75 | 0.84 | ||
| Cryptodendrum adhaesivum | 0.83 | A0A6P8IHV9.1.A Four-domain proteases inhibitor-like | 75.86 | 0.98 | ||
| Clade C | Calliactis polypus | 0.52 | 3tjq.1.A Serine protease HTRA1 | 38.18 | 0.87 | |
| Nemanthus annamensis | 0.51 | 3tjq.1.A Serine protease HTRA1 | 35.19 | 0.87 | ||
| Telmatactis sp. | 0.43 | 1wqj.1.A Insulin-like growth factor binding protein 4 | 32.65 | 0.91 |
| Venom | Rate Distributions (MEME) | FUBAR | ||
|---|---|---|---|---|
| Clade | dN/dS | Sites under (+) Selection | Sites (+) | Sites (−) |
| Cnidoinsulin | ||||
| Clade 1a | 0.0125 | 0 | 0 | 42 |
| Clade 1b | 0.0075 | 1 | 1 | 32 |
| Clade 1 (1a+1b) | 0.0247 | 3 | 0 | 38 |
| Clade 2 | 0.0557 | 3 | 0 | 26 |
| Clade 3 | 0.156 | 4 | 0 | 11 |
| All ((1+2)+3)) | 0.0211 | 0 | 0 | 36 |
| Chain A | 0.185 | 2 | / | / |
| Chain B | 0.212 | 0 | / | / |
| VP302 | ||||
| Clade A | 0.256 | 0 | 0 | 30 |
| Clade B | 0.0979 | 5 | 0 | 43 |
| Clade (A+B) | 0.119 | 7 | 0 | 45 |
| Clade C | 0.178 | 2 | 0 | 26 |
| All (A+B)+C) | 0.172 | 10 | 0 | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, A.; Sozanski, K.S.; Daly, M. Towards the Exploration and Evolution of Insulin-like Venoms in Actiniaria (Sea anemones). Mar. Drugs 2024, 22, 136. https://doi.org/10.3390/md22030136
Delgado A, Sozanski KS, Daly M. Towards the Exploration and Evolution of Insulin-like Venoms in Actiniaria (Sea anemones). Marine Drugs. 2024; 22(3):136. https://doi.org/10.3390/md22030136
Chicago/Turabian StyleDelgado, Alonso, Kyle S. Sozanski, and Marymegan Daly. 2024. "Towards the Exploration and Evolution of Insulin-like Venoms in Actiniaria (Sea anemones)" Marine Drugs 22, no. 3: 136. https://doi.org/10.3390/md22030136
APA StyleDelgado, A., Sozanski, K. S., & Daly, M. (2024). Towards the Exploration and Evolution of Insulin-like Venoms in Actiniaria (Sea anemones). Marine Drugs, 22(3), 136. https://doi.org/10.3390/md22030136