
Marine-Derived Metabolites Act as Promising Antifungal Agents


Abstract
:1. Introduction
2. Disrupting the Cell Wall
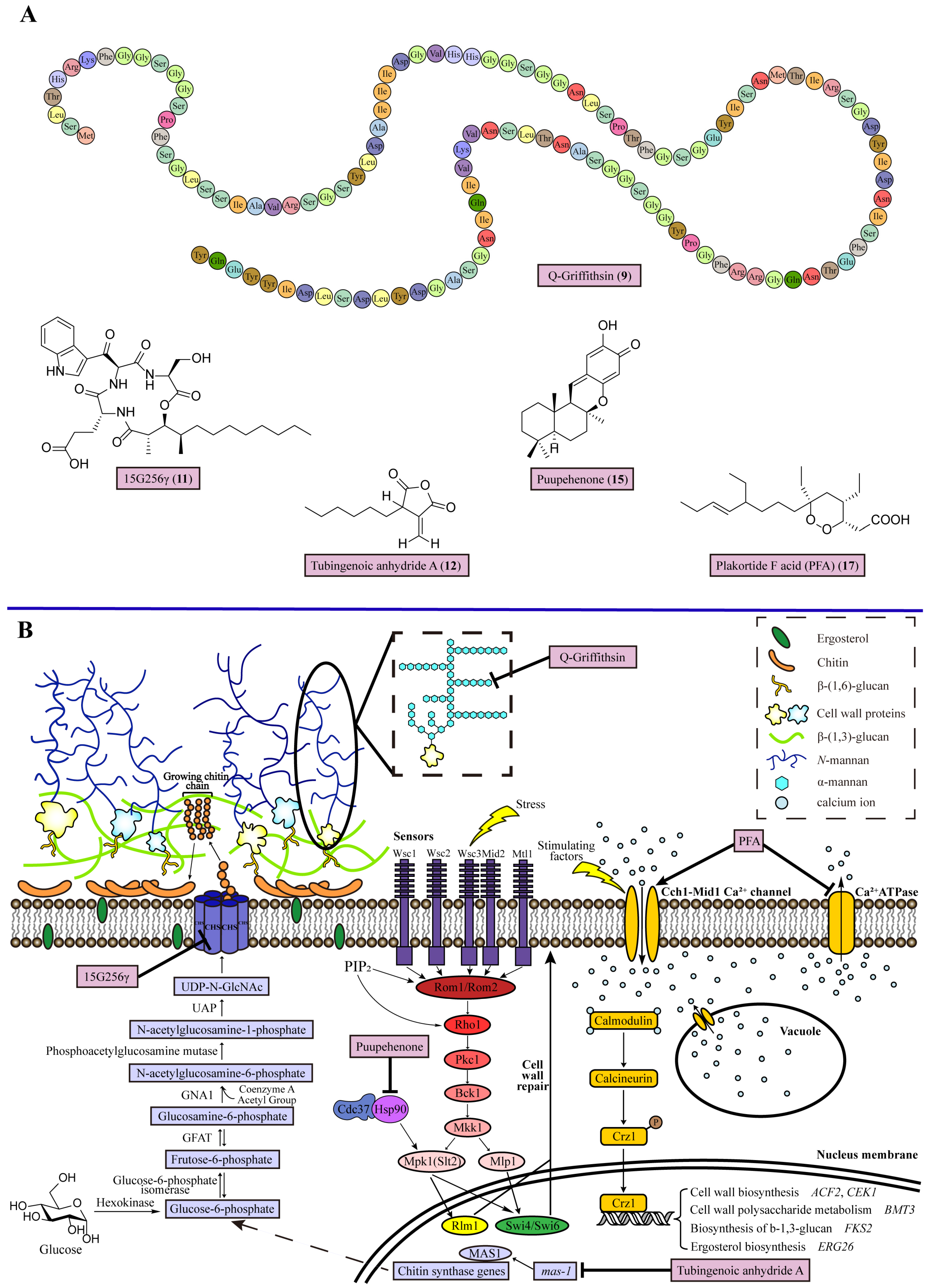
2.1. Inhibiting Mannan Biosynthesis
2.2. Inhibiting Chitin Biosynthesis
2.3. Inhibiting the Cell Wall Integrity (CWI) Pathway
2.4. Disrupting Ca2+ Homeostasis
3. Disrupting the Cell Membrane
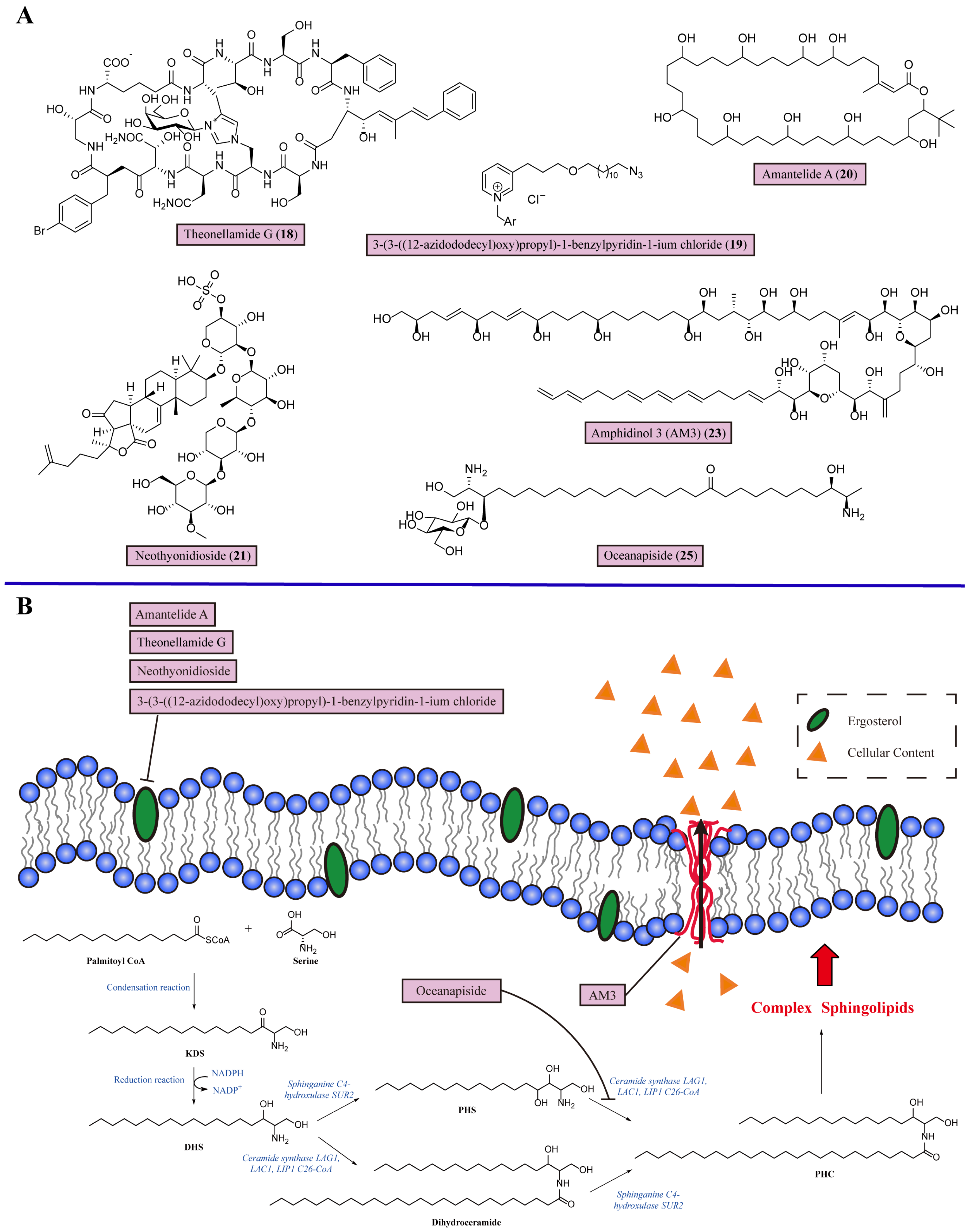
3.1. Targeting Ergosterol in the Cell Membrane
3.2. Inhibiting Sphingolipid Biosynthesis
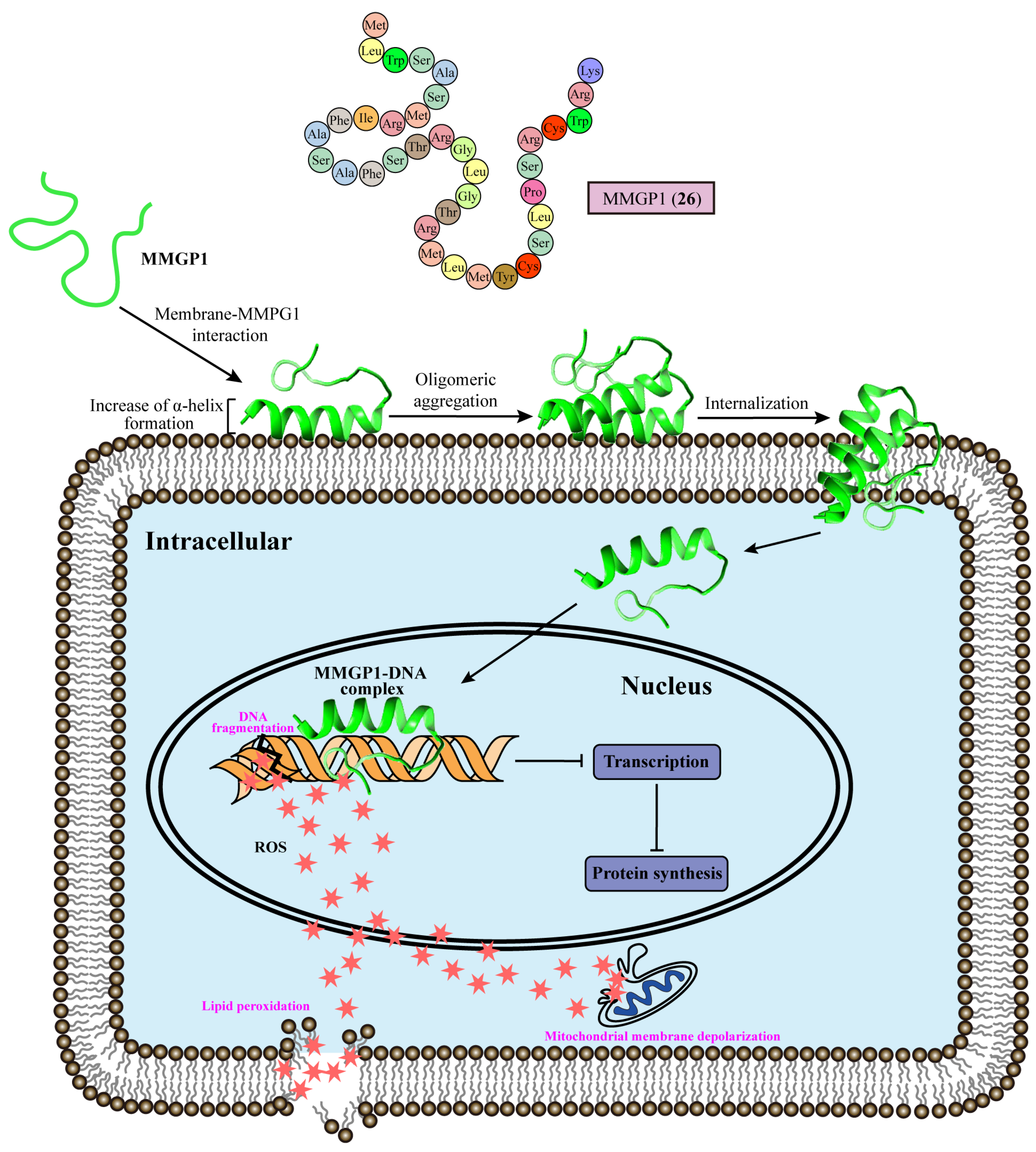
4. Effects on Fungal Chromosomes
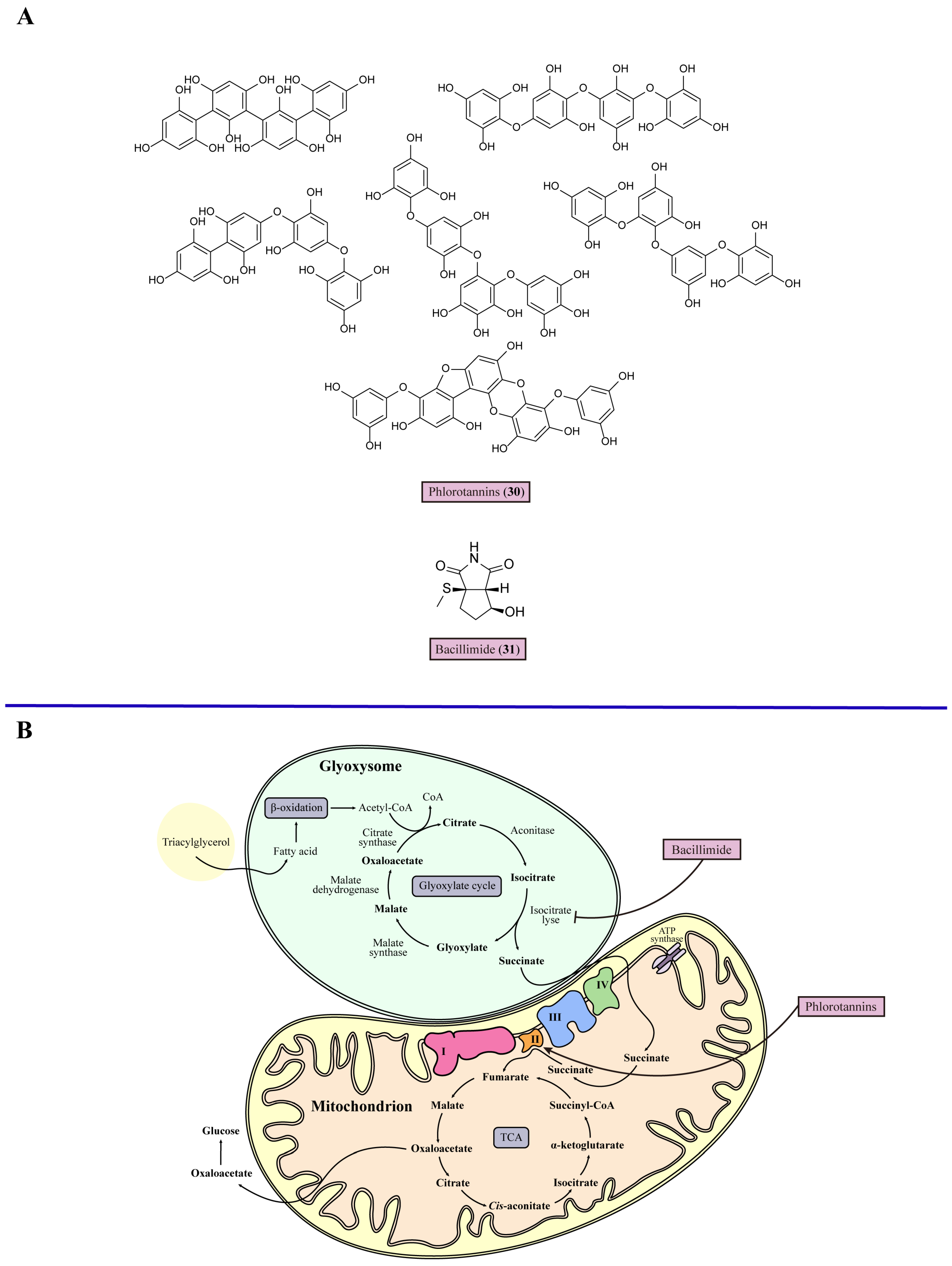
5. Mitochondrial Dysfunction
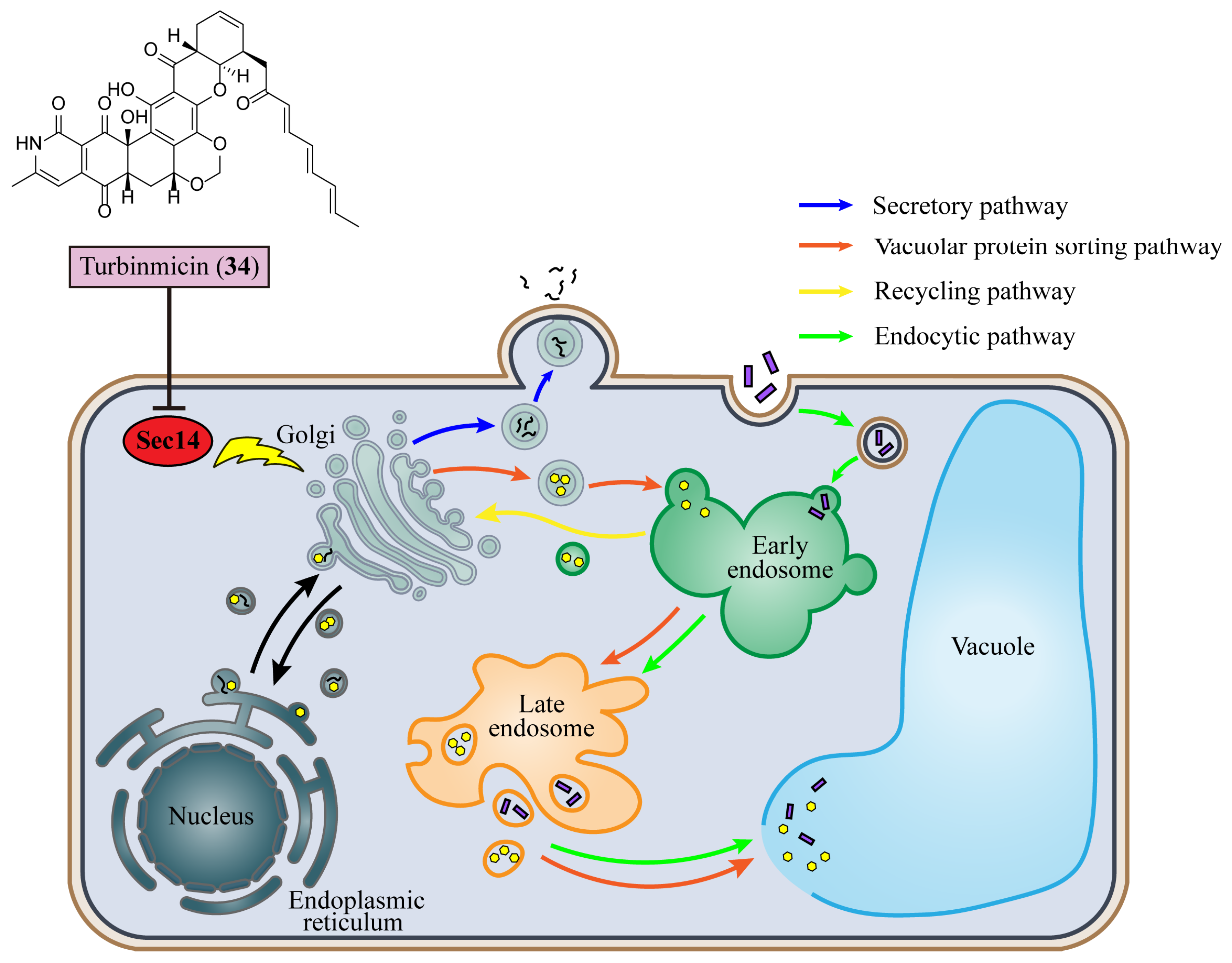
6. Inhibition of Vesicular Trafficking
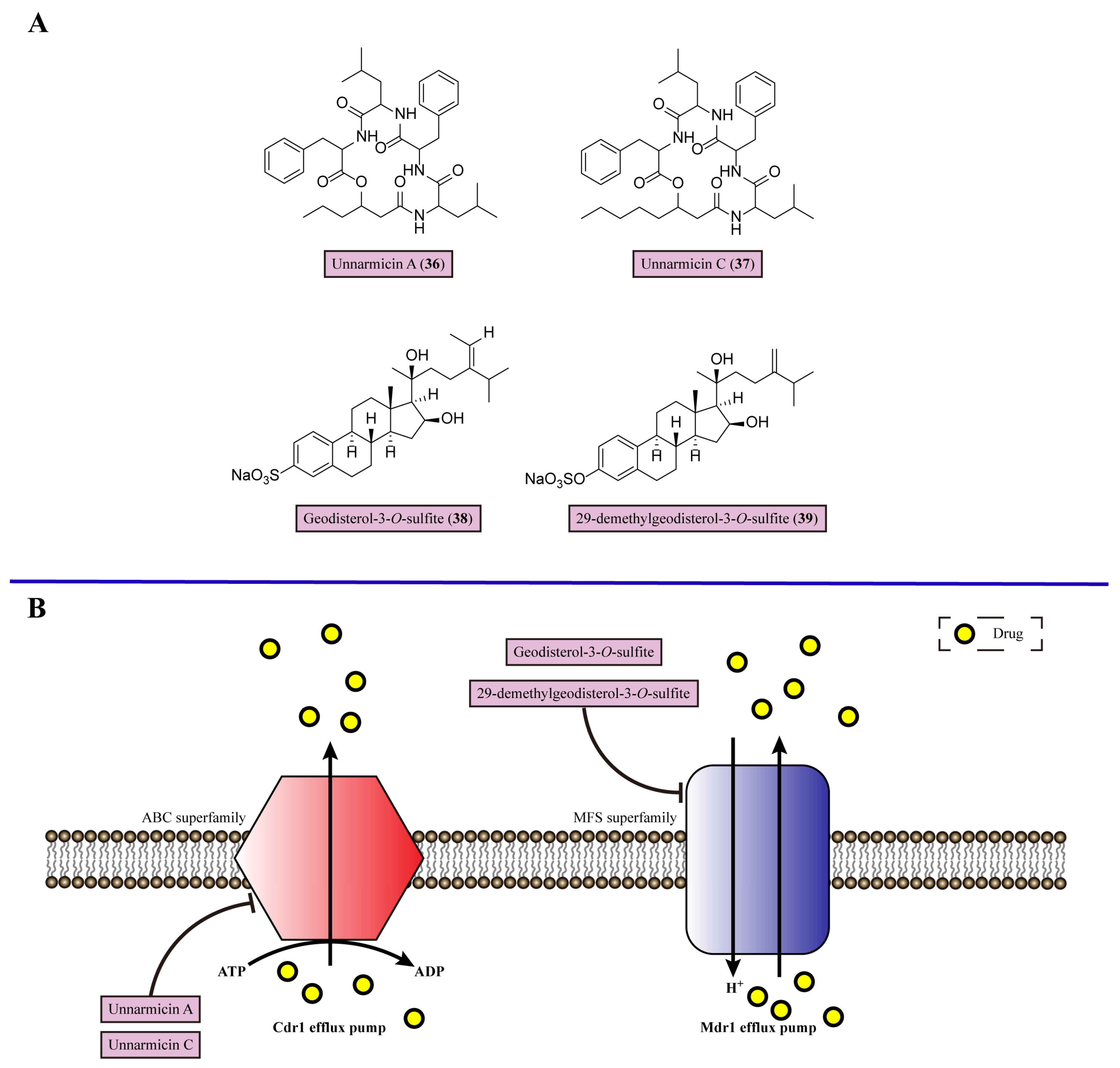
7. Inhibiting Efflux Pumps
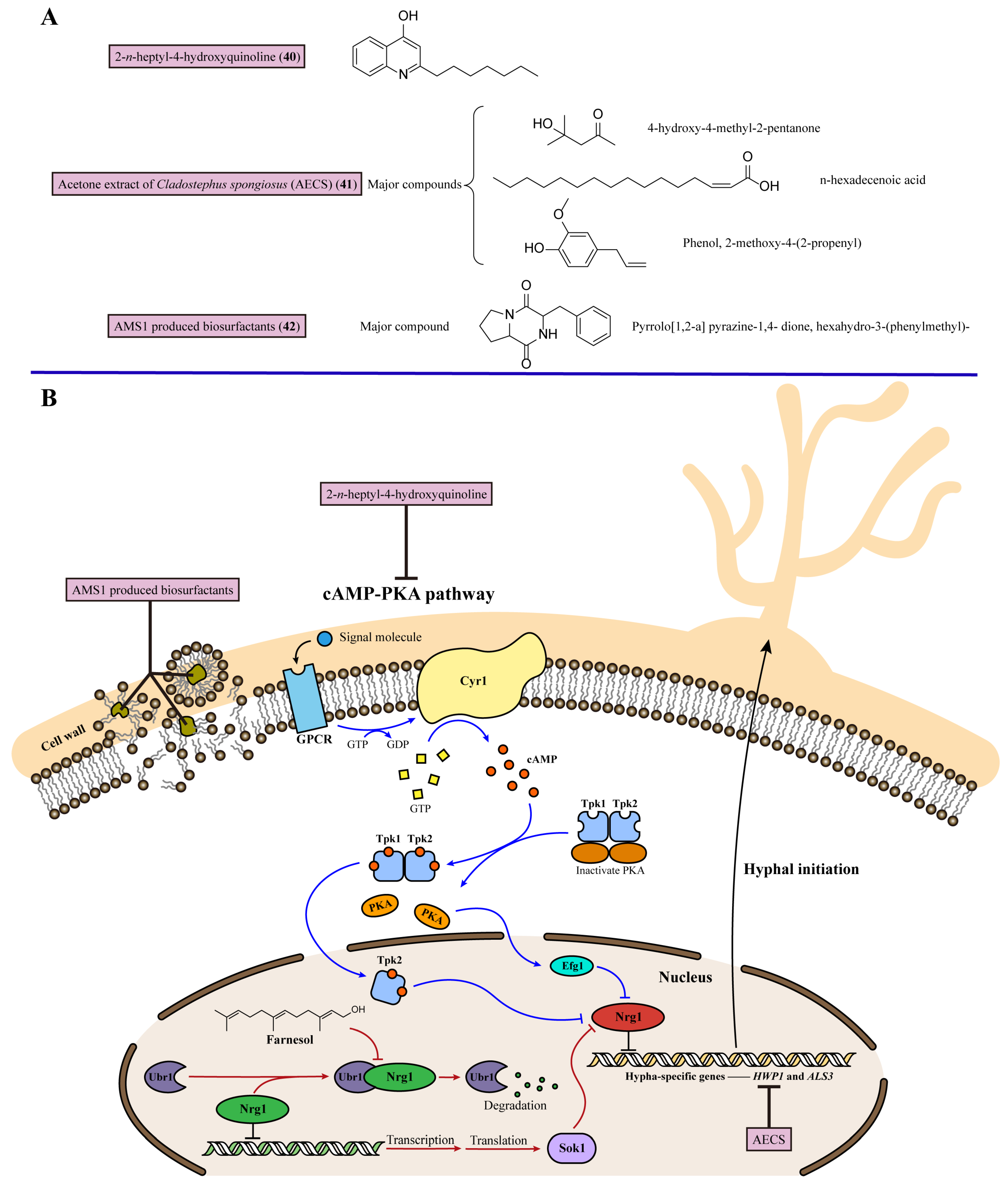
8. Inhibiting Fungal Hyphal Growth
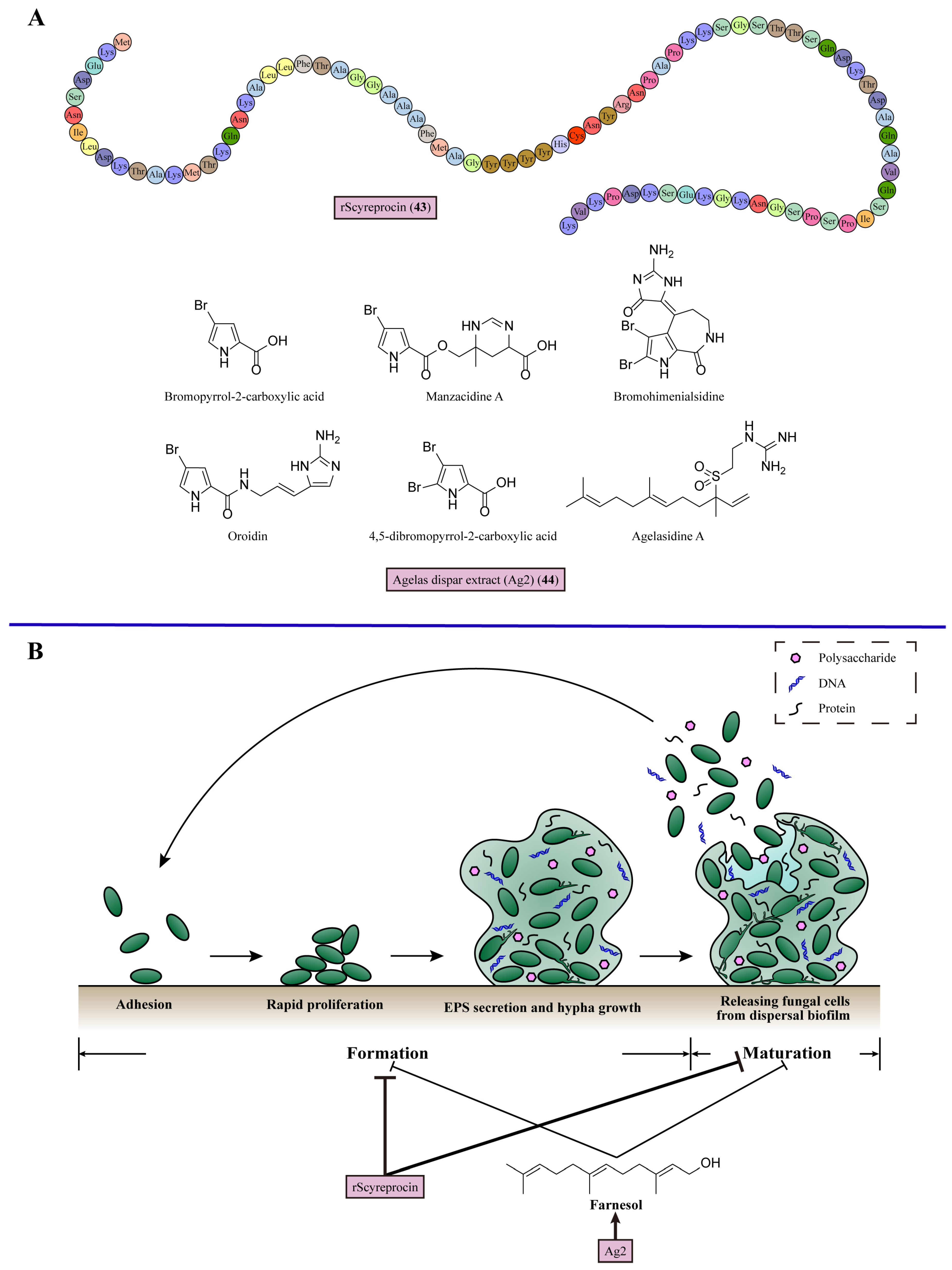
9. Inhibiting Biofilm Formation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolites | Mechanism | Source | Activity | Refs. |
|---|---|---|---|---|---|
| 1 | Q-Griffithsin (9) | Binds with α-mannan to break the outer layer of the fungal cell wall | Red alga Griffithsia species with glutamine substitution of Met78 | Candida albicans, Candida glabrata, Candida parapsilosis, Candida krusei, and Candida auris (MIC = 6, 95, 24, 95, and 48 mg/mL) | [32] |
| 2 | 15G256γ (11) | Inhibits the activity of chitin synthase to weaken fungal cell wall | Marine fungus Hypoxylon oceanicum | Trichophyton rubrum, T. mentagrophytes, Epidermophyton floccosum, Microsporum audoinii, C. albicans, C. parapsiliosis, and C. glabrata (MIC = 2–16 μg/mL) | [44] |
| 3 | Tubingenoic anhydride A (12) | Suppresses the expression of mas-1 mediating part of chitin synthase expression | Fungus Aspergillus tubingensis OY907 from sponge Ircinia variabilis | Neurospora crassa (MIC = 330 μM) | [45] |
| 4 | Puupehenone (15) | Acts as an Hsp90 inhibitor to block cell wall integrity pathway | Marine sponge Hyrtios species | Cryptococcus neoformans, C. glabrata, and C. albicans (puupehenone + caspofungin FICI = 0.38, 0.48, and 0.39) | [51] |
| 5 | Plakortide F acid (PFA) (17) | Disrupts fungal intracellular calcium ion homeostasis | Marine sponge Plakortis halichondrioides | C. albicans, C. neoformans, and Aspergillus fumigatus (MIC = 0.08, 2.5, and 5.00 μg/mL) | [57] |
| 6 | Theonellamide G (18) | Binds with the 3β-OH group in ergosterol | Marine sponge Theonella species | Wide-type C. albicans and amphotericin B-resistant C. albicans (IC50 = 4.49 and 2.0 μM) | [63] |
| 7 | 3-(3-((12-azidododecyl)oxy)propyl)-1-benzylpyridin-1-ium chloride (19) | Binds with membrane ergosterol | Marine sponges of the Haplosclerida order | C. albicans, C. glabrata, C. krusei, and Candida tropicalis (MIC = 3.9–7.8 μg/mL) | [64] |
| 8 | Amantelide A (20) | Recognizes ergosterol to bind with membrane | Marine gray cyanobacteria Oscilliatoriales | Saccharmomyces cerevisiae and Schizosaccharomyces pombe (MIC = 50 and 12.5 μM) | [65] |
| 9 | Neothyonidioside (21) | Forms a large complex with ergosterol to reduce the ability of the fungal membrane to bend and form multivesicular body vesicles | Sea cucumber Australostichopus mollis | S. cerevisiae (MIC = 1 μM) | [66] |
| 10 | Amphidinol 3 (AM3) (23) | Recognizes 3β-OH group in ergosterol through hydrogen bonding to permeabilize the fungal cell membrane | Dinoflagellate Amphidinium klebsii | Aspergillus Niger (MEC = 9.0 μg/disk) | [68] |
| 11 | Oceanapiside (25) | Blocks sphingolipid biosynthesis in fungi | Marine sponge Oceanapia phillipensis | C. glabrata (MIC = 10 μg/mL) | [75] |
| 12 | MMGP1 (26) | Internalized into the cytosol to form MMPG1–DNA complex, interfering with transcription | Marine metagenome | C. albicans (MIC = 0.57 μM) | [80] |
| 13 | Phlorotannins (30) | Stimulates the activity of electron transport chain Complex II and regulates mitochondrial membrane potential to induce mitochondrial dysfunction | Brown seaweeds Cytoseira nodicaulis, Cystoseira usneoides, and Fucus spiralis | C. albicans (MIC = 15.6 mg/mL) | [89] |
| 14 | Bacillimide (31) | Inhibits the transcription of isocitrate lyase to suppress the glyoxylate cycle | Marine actinomycete Streptomyces bacillaris | Candida albicans (IC50 = 44.24 μM) | [92] |
| 15 | Turbinmicin (34) | Binds with Sec14 to interfere with the secretion during vesicular trafficking in the Golgi and decreases the production of fungal extracellular vesicles | Sea squirt Micromonospora species | C. auris, C. albicans, C. tropicalis, C. glabrata, A. fumigatus, Fusarium species, and Scedosporium species (MIC = 0.03–0.5 μg/mL) | [96] |
| 16 | Unnarmicin A (36) | Targets Cdr1 efflux pump in C. albicans | Marine γ-proteobacterium | Azole-resistant C. albicans (unnarmicin A + fluconazole MIC = 10 μg/mL) | [103] |
| 17 | Unnarmicin C (37) | Targets Cdr1 efflux pump in C. albicans | Marine γ-proteobacterium | Azole-resistant C. albicans (unnarmicin C + fluconazole MIC = 10 μg/mL) | [103] |
| 18 | Geodisterol-3-O-sulfite (38) | Targets Mdr1 efflux pump in C. albicans | Marine sponge Topsentia species | Fluconazole-resistant C. albicans (Geodisterol-3-O-sulfite + fluconazole FICI = 0.2) | [104] |
| 19 | 29-demethylgeodisterol-3-O-sulfite (39) | Targets Mdr1 efflux pump in C. albicans | Marine sponge Topsentia species | Fluconazole-resistant C. albicans (29-demethylgeodisterol-3-O-sulfite + fluconazole FICI = 0.2) | [104] |
| 20 | 2-n-heptyl-4-hydroxyquinoline (40) | Regulates cAMP-Efg1 pathway to decrease expression of the hypha-specific genes HWP1 and ALS3 | Marine Streptomyces species MBTG13 | Hyphal form of C. albicans (IC50 = 11.4 μg/mL) | [110] |
| 21 | Acetone extract of Cladostephus spongiosus (AECS) (41) | Downregulates the expression of the hypha-specific genes HWP1, ALS1, and SAP4 | Marine mactoalgal Cladostephus spongiosus | C. krusei, C. glabrata, C. parapsilosis, and C.albicans (MIC = 80, 90, 100, and 90 μg/mL) | [111] |
| 22 | AMS1 produced biosurfactants (42) | Acts as a biosurfactant to inhibit the transition from yeast to hyphae | Marine bacterium AMS1 | C. albicans (MIC = 160 μg/mL) | [112] |
| 23 | rScyreprocin (43) | Suppresses biofilm formation and eradicates mature biofilms | Mud crab Scylla paramamosain | C. neoformans, C. albicans, C. krusei, C. parapsilosis, C. tropicalis, N. crassa, Fusarium species and Aspergillus species (MIC = 1–32 μM) | [117] |
| 24 | Agelas dispar extract (Ag2) (44) | Produces farnesol to prevent biofilm formation | Marine sponge Agelas dispar | C. albicans, C. tropicalis, C. krusei, C. glabrata, and C. parapsilosis (MIC = 0.15625–2.5 mg/mL) | [118] |
10. Discussion
11. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Casadevall, A. Immunity to Invasive Fungal Diseases. Annu. Rev. Immunol. 2022, 40, 121–141. [Google Scholar] [CrossRef]
- Lu, H.; Hong, T.; Jiang, Y.; Whiteway, M.; Zhang, S. Candidiasis: From cutaneous to systemic, new perspectives of potential targets and therapeutic strategies. Adv. Drug Deliv. Rev. 2023, 199, 114960. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Azoulay, E.; Kullberg, B.J.; Ruhnke, M.; Shoham, S.; Vazquez, J.; Giacobbe, D.R.; Calandra, T. EORTC/MSGERC Definitions of Invasive Fungal Diseases: Summary of Activities of the Intensive Care Unit Working Group. Clin. Infect. Dis. 2021, 72, S121–S127. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W. Global incidence and mortality of severe fungal disease. Lancet Infect. 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Lu, H.; Whiteway, M.; Jiang, Y. Understanding fluconazole tolerance in Candida albicans: Implications for effective treatment of candidiasis and combating invasive fungal infections. J. Glob. Antimicrob. Resist. 2023, 35, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Shrivastava, M.; Whiteway, M.; Jiang, Y. Candida albicans targets that potentially synergize with fluconazole. Crit. Rev. Microbiol. 2021, 47, 323–337. [Google Scholar] [CrossRef]
- Lu, H.; Li, W.; Whiteway, M.; Wang, H.; Zhu, S.; Ji, Z.; Feng, Y.; Yan, L.; Fang, T.; Li, L.; et al. A Small Molecule Inhibitor of Erg251 Makes Fluconazole Fungicidal by Inhibiting the Synthesis of the 14alpha-Methylsterols. mBio 2023, 14, e0263922. [Google Scholar] [CrossRef] [PubMed]
- Zhen, C.; Lu, H.; Jiang, Y. Novel Promising Antifungal Target Proteins for Conquering Invasive Fungal Infections. Front. Microbiol. 2022, 13, 911322. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lu, H.; Jiang, Y. Natural Polyketides Act as Promising Antifungal Agents. Biomolecules 2023, 13, 1572. [Google Scholar] [CrossRef]
- Jenks, J.D.; Cornely, O.A.; Chen, S.C.-A.; Thompson, G.R., III; Hoenigl, M. Breakthrough invasive fungal infections: Who is at risk? Mycoses 2020, 63, 1021–1032. [Google Scholar] [CrossRef]
- Feng, Z.; Lu, H.; Jiang, Y. Promising immunotherapeutic targets for treating candidiasis. Front. Cell Infect. Microbiol. 2024, 14, 1339501. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Wang, L.; Feng, Z.; Hang, S.; Yu, J.; Feng, Y.; Lu, H.; Jiang, Y. Halofantrine Hydrochloride Acts as an Antioxidant Ability Inhibitor That Enhances Oxidative Stress Damage to Candida albicans. Antioxidants 2024, 13, 223. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Xiong, J.; Wang, L.; Feng, Z.; Hang, S.; Yu, J.; Li, W.; Feng, Y.; Lu, H.; Jiang, Y. Unexpected Inhibitory Effect of Octenidine Dihydrochloride on Candida albicans Filamentation by Impairing Ergosterol Biosynthesis and Disrupting Cell Membrane Integrity. Antibiotics 2023, 12, 1675. [Google Scholar] [CrossRef] [PubMed]
- Fakhim, H.; Badali, H.; Dannaoui, E.; Nasirian, M.; Jahangiri, F.; Raei, M.; Vaseghi, N.; Ahmadikia, K.; Vaezi, A. Trends in the Prevalence of Amphotericin B-Resistance (AmBR) among Clinical Isolates of Aspergillus Species. J. Med. Mycol. 2022, 32, 101310. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, D.; Neofytos, D. Invasive Aspergillosis and the Impact of Azole-resistance. Curr. Fungal Infect. Rep. 2023, 17, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Habbu, P.; Warad, V.; Shastri, R.; Madagundi, S.; Kulkarni, V.H. Antimicrobial metabolites from marine microorganisms. Chin. J. Nat. Med. 2016, 14, 101–116. [Google Scholar] [CrossRef]
- Cardoso, J.; Nakayama, D.G.; Sousa, E.; Pinto, E. Marine-Derived Compounds and Prospects for Their Antifungal Application. Molecules 2020, 25, 5856. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and Antifungal Compounds from Marine Fungi. Mar. Drugs 2015, 13, 3479–3513. [Google Scholar] [CrossRef] [PubMed]
- Touati, I.; Ruiz, N.; Thomas, O.; Druzhinina, I.S.; Atanasova, L.; Tabbene, O.; Elkahoui, S.; Benzekri, R.; Bouslama, L.; Pouchus, Y.F.; et al. Hyporientalin A, an anti-Candida peptaibol from a marine Trichoderma orientale. World J. Microbiol. Biotechnol. 2018, 34, 98. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Pierce, M.L.; Howe, K.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine pharmacology in 2018: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Pharmacol. Res. 2022, 183, 106391. [Google Scholar] [CrossRef]
- Costa Vasconcelos Alves, A.M.; Cruz-Martins, N.; Rodrigues, C.F. Marine Compounds with Anti-Candida sp. Activity: A Promised “Land” for New Antifungals. J. Fungi 2022, 8, 669. [Google Scholar] [CrossRef] [PubMed]
- Ganeshkumar, A.; Goncale, J.C.; Rajaram, R.; Junqueira, J.C. Anti-Candidal Marine Natural Products: A Review. J. Fungi 2023, 9, 800. [Google Scholar] [CrossRef] [PubMed]
- Thawabteh, A.M.; Swaileh, Z.; Ammar, M.; Jaghama, W.; Yousef, M.; Karaman, R.; Bufo, S.A.; Scrano, L. Antifungal and Antibacterial Activities of Isolated Marine Compounds. Toxins 2023, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Cortes, J.C.G.; Curto, M.A.; Carvalho, V.S.D.; Perez, P.; Carlos Ribas, J. The fungal cell wall as a target for the development of new antifungal therapies. Biotechnol. Adv. 2019, 37, 107352. [Google Scholar] [CrossRef]
- Lima, S.L.; Colombo, A.L.; de Almeida Junior, J.N. Fungal Cell Wall: Emerging Antifungals and Drug Resistance. Front. Microbiol. 2019, 10, 2573. [Google Scholar] [CrossRef] [PubMed]
- Cow, N.A.R.; Lenardon, M.D. Architecture of the dynamic fungal cell wall. Nat. Rev. Microbiol. 2023, 21, 248–259. [Google Scholar] [CrossRef]
- Cutler, J.E. N-glycosylation of yeast, with emphasis on Candida albicans. Med. Mycol. 2001, 39, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A.; Bates, S.; Buurman, E.T.; Hughes, H.B.; MacCallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.J.; Bain, J.M.; Brand, A.; et al. Mnt1p and Mnt2p of Candida albicans are partially redundant α-1,2-mannosyltransferases that participate in O-linked mannosylation and are required for adhesion and virulence. J. Biol. Chem. 2005, 280, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Diaz-Jimenez, D.F.; Lopez-Romero, E.; Zinker, S.; Ponce-Norla, P.; Kullberg, B.J.; Brown, A.J.P.; Odds, F.C.; et al. Endoplasmic reticulum α-glycosidases of Candida albicans are required for N glycosylation, cell wall integrity, and normal host-fungus interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef]
- Kakushima, M.; Masuyoshi, S.; Hirano, M.; Shinoda, M.; Ohta, A.; Kamei, H.; Oki, T. In vitro and in vivo antifungal activities of BMY-28864, a water-soluble pradimicin derivative. Antimicrob. Agents Chemother. 1991, 35, 2185–2190. [Google Scholar] [CrossRef]
- Hiramoto, F.; Nomura, N.; Furumai, T.; Oki, T.; Igarashi, Y. Apoptosis-like cell death of Saccharomyces cerevisiae induced by a mannose-binding antifungal antibiotic, pradimicin. J. Antibiot. 2003, 56, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Nabeta, H.W.; Kouokam, J.C.; Lasnik, A.B.; Fuqua, J.L.; Palmer, K.E. Novel Antifungal Activity of Q-Griffithsin, a Broad-Spectrum Antiviral Lectin. Microbiol. Spectrum 2021, 9, e00957-21. [Google Scholar] [CrossRef] [PubMed]
- Nabeta, H.W.; Lasnik, A.B.; Fuqua, J.L.; Wang, L.; Rohan, L.C.; Palmer, K.E. Antiviral lectin Q-Griffithsin suppresses fungal infection in murine models of vaginal candidiasis. Front. Cell. Infect. Microbiol. 2022, 12, 976033. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.M.; Tupe, S.G.; Deshpande, M.V. Chitin Synthase Inhibitors as Antifungal Agents. Mini-Rev. Med. Chem. 2013, 13, 222–236. [Google Scholar]
- Brauer, V.S.; Pessoni, A.M.; Freitas, M.S.; Cavalcanti-Neto, M.P.; Ries, L.N.A.; Almeida, F. Chitin Biosynthesis in Aspergillus Species. J. Fungi 2023, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Jiang, C.; Wang, Q.; Zhao, Z.; Jin, Q.; Xu, J.-R.; Liu, H. Evolution and Functional Insights of Different Ancestral Orthologous Clades of Chitin Synthase Genes in the Fungal Tree of Life. Front. Plant Sci. 2016, 7, 168614. [Google Scholar] [CrossRef] [PubMed]
- Mandel, M.A.; Galgiani, J.N.; Kroken, S.; Orbach, M.J. Coccidioides posadasii contains single chitin synthase genes corresponding to classes I to VII. Fungal Genet. Biol. 2006, 43, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A. Chapter Four—Chitin and Glucan, the Yin and Yang of the Fungal Cell Wall, Implications for Antifungal Drug Discovery and Therapy. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 83, pp. 145–172. [Google Scholar]
- Free, S.J. Fungal Cell Wall Organization and Biosynthesis. In Advances in Genetics; Friedmann, T., Dunlap, J.C., Goodwin, S.F., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; Volume 81, pp. 33–82. [Google Scholar]
- Gong, Z.; Zhang, S.; Liu, J. Recent Advances in Chitin Biosynthesis Associated with the Morphology and Secondary Metabolite Synthesis of Filamentous Fungi in Submerged Fermentation. J. Fungi 2023, 9, 205. [Google Scholar] [CrossRef]
- Rivera-Molina, F.E.; González-Crespo, S.; Cruz, Y.M.-D.l.; Ortiz-Betancourt, J.M.; Rodríguez-Medina, J.R. 2,3-Butanedione monoxime increases sensitivity to Nikkomycin Z in the budding yeast Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2006, 22, 255–260. [Google Scholar] [CrossRef]
- Li, R.K.; Rinaldi, M.G. In Vitro Antifungal Activity of Nikkomycin Z in Combination with Fluconazole or Itraconazole. Antimicrob. Agents Chemother. 1999, 43, 1401–1405. [Google Scholar] [CrossRef]
- Larwood, D.J. Nikkomycin Z—Ready to Meet the Promise? J. Fungi 2020, 6, 261. [Google Scholar] [CrossRef] [PubMed]
- Abbanat, D.; Leighton, M.; Maiese, W.; Jones, E.B.G.; Pearce, C.; Greestein, M. Cell wall active antifungal compounds produced by the marine fungus Hypoxylon oceanicum LL-15G256. J. Antibiot. 1998, 51, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Koch, L.; Lodin, A.; Herold, I.; Ilan, M.; Carmeli, S.; Yarden, O. Sensitivity of Neurospora crassa to a Marine-Derived Aspergillus tubingensis Anhydride Exhibiting Antifungal Activity That Is Mediated by the MAS1 Protein. Mar. Drugs 2014, 12, 4713–4731. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Regulation of Cell Wall Biogenesis in Saccharomyces cerevisiae: The Cell Wall Integrity Signaling Pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Juvvadi, P.R.; Steinbach, W.J. Heat shock protein 90 (Hsp90): A novel antifungal target against Aspergillus fumigatus. Crit. Rev. Microbiol. 2016, 42, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.C.; Santos, C.A.; Malavazi, I. The Regulatory Function of the Molecular Chaperone Hsp90 in the Cell Wall Integrity of Pathogenic Fungi. Curr. Proteom. 2019, 16, 44–53. [Google Scholar] [CrossRef]
- Hawle, P.; Horst, D.; Bebelman, J.P.; Yang, X.X.; Siderius, M.; van der Vies, S.M. Cdc37p is required for stress-induced high-osmolarity glycerol and protein kinase C mitogen-activated protein kinase pathway functionality by interaction with Hog1p and Slt2p (Mpk1p)v. Eukaryot. Cell 2007, 6, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Sellers-Moya, Á.; Nuévalos, M.; Molina, M.; Martín, H. Clotrimazole-Induced Oxidative Stress Triggers Novel Yeast Pkc1-Independent Cell Wall Integrity MAPK Pathway Circuitry. J. Fungi 2021, 7, 647. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.K.; Feng, Q.; Liu, L.; Levin, D.E.; Roy, K.K.; Doerksen, R.J.; Baerson, S.R.; Shi, X.; Pan, X.; Xu, W.-H.; et al. Puupehenone, a Marine-Sponge-Derived Sesquiterpene Quinone, Potentiates the Antifungal Drug Caspofungin by Disrupting Hsp90 Activity and the Cell Wall Integrity Pathway. mSphere 2020, 5, 10–1128. [Google Scholar] [CrossRef]
- Zhang, C.; Ren, Y.; Gu, H.; Gao, L.; Zhang, Y.; Lu, L. Calcineurin-mediated intracellular organelle calcium homeostasis is required for the survival of fungal pathogens upon extracellular calcium stimuli. Virulence 2021, 12, 1091–1110. [Google Scholar] [CrossRef]
- Sanglard, D. Aequorin as a Useful Calcium-Sensing Reporter in Candida albicans. J. Fungi 2021, 7, 319. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Shrivastava, M.; Lu, H.; Jiang, Y.Y. Calcium-calcineurin signaling pathway in Candida albicans: A potential drug target. Microbiol. Res. 2021, 249, 126786. [Google Scholar] [CrossRef] [PubMed]
- William, E.C. Characterization of a Novel, Broad-Based Fungicidal Activity for the Antiarrhythmic Drug Amiodarone. J. Pharmacol. Exp. Ther. 2002, 300, 195. [Google Scholar] [CrossRef]
- Sen Gupta, S.; Ton, V.K.; Beaudry, V.; Rulli, S.; Cunningham, K.; Rao, R. Antifungal activity of amiodarone is mediated by disruption of calcium homeostasis. J. Biol. Chem. 2003, 278, 28831–28839. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Feng, Q.; Jacob, M.R.; Avula, B.; Mask, M.M.; Baerson, S.R.; Tripathi, S.K.; Mohammed, R.; Hamann, M.T.; Khan, I.A.; et al. The Marine Sponge-Derived Polyketide Endoperoxide Plakortide F Acid Mediates Its Antifungal Activity by Interfering with Calcium Homeostasis. Antimicrob. Agents Chemother. 2011, 55, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Maresova, L.; Muend, S.; Zhang, Y.-Q.; Sychrova, H.; Rao, R. Membrane Hyperpolarization Drives Cation Influx and Fungicidal Activity of Amiodarone. J. Biol. Chem. 2009, 284, 2795–2802. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chen, B. Role of cell membrane homeostasis in the pathogenicity of pathogenic filamentous fungi. Virulence 2024, 15, 2299183. [Google Scholar] [CrossRef]
- Sant, D.G.; Tupe, S.G.; Ramana, C.V.; Deshpande, M.V. Fungal cell membrane-promising drug target for antifungal therapy. J. Appl. Microbiol. 2016, 121, 1498–1510. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef]
- Espiritu, R.A.; Matsumori, N.; Murata, M.; Nishimura, S.; Kakeya, H.; Matsunaga, S.; Yoshida, M. Interaction between the marine sponge cyclic peptide theonellamide A and sterols in lipid bilayers as viewed by surface plasmon resonance and solid-state (2)H nuclear magnetic resonance. Biochemistry 2013, 52, 2410–2418. [Google Scholar] [CrossRef]
- Youssef, D.T.A.; Shaala, L.A.; Mohamed, G.A.; Badr, J.M.; Bamanie, F.H.; Ibrahim, S.R.M. Theonellamide G, a Potent Antifungal and Cytotoxic Bicyclic Glycopeptide from the Red Sea Marine Sponge Theonella swinhoei. Mar. Drugs 2014, 12, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.T.; Lima, W.G.; Sousa, J.F.; Saldanha, A.A.; De Sa, N.P.; Morais, F.B.; Prates Silva, M.K.; Ribeiro Viana, G.H.; Johann, S.; Soares, A.C.; et al. Design, synthesis, and biodistribution studies of new analogues of marine alkaloids: Potent in vitro and in vivo fungicidal agents against Candida spp. Eur. J. Med. Chem. 2021, 210, 113048. [Google Scholar] [CrossRef] [PubMed]
- Elsadek, L.A.; Matthews, J.H.; Nishimura, S.; Nakatani, T.; Ito, A.; Gu, T.; Luo, D.; Salvador-Reyes, L.A.; Paul, V.J.; Kakeya, H.; et al. Genomic and Targeted Approaches Unveil the Cell Membrane as a Major Target of the Antifungal Cytotoxin Amantelide A. ChemBioChem 2021, 22, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Yibmantasiri, P.; Leahy, D.C.; Busby, B.P.; Angermayr, S.A.; Sorgo, A.G.; Boeger, K.; Heathcott, R.; Barber, J.M.; Moraes, G.; Matthews, J.H.; et al. Molecular basis for fungicidal action of neothyonidioside, a triterpene glycoside from the sea cucumber, Australostichopus mollis. Mol. Biosyst. 2012, 8, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Houdai, T.; Matsuoka, S.; Morsy, N.; Matsumori, N.; Satake, M.; Murata, M. Hairpin conformation of amphidinols possibly accounting for potent membrane permeabilizing activities. Tetrahedron 2005, 61, 2795–2802. [Google Scholar] [CrossRef]
- Houdai, T.; Matsuoka, S.; Matsumori, N.; Murata, M. Membrane-permeabilizing activities of amphidinol 3, polyene-polyhydroxy antifungal from a marine dinoflagellate. BBA-Biomembranes 2004, 1667, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Espiritu, R.A.; Matsumori, N.; Tsuda, M.; Murata, M. Direct and Stereospecific Interaction of Amphidinol 3 with Sterol in Lipid Bilayers. Biochemistry 2014, 53, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Montefusco, D.J.; Matmati, N.; Hannun, Y.A. The yeast sphingolipid signaling landscape. Chem. Phys. Lipids 2014, 177, 26–40. [Google Scholar] [CrossRef]
- Jadhav, S.; Greenberg, M.L. Harnessing the power of yeast to elucidate the role of sphingolipids in metabolic and signaling processes pertinent to psychiatric disorders. Clin. Lipidol. 2014, 9, 533–551. [Google Scholar] [CrossRef]
- Mor, V.; Rella, A.; Farnoud Amir, M.; Singh, A.; Munshi, M.; Bryan, A.; Naseem, S.; Konopka James, B.; Ojima, I.; Bullesbach, E.; et al. Identification of a New Class of Antifungals Targeting the Synthesis of Fungal Sphingolipids. mBio 2015, 6, e00647-15. [Google Scholar] [CrossRef]
- Perfect, J.R. The antifungal pipeline: A reality check. Nat. Rev. Drug Discov. 2017, 16, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, G.M.; Hong, T.W.; Molinski, T.F.; Lerch, M.L.; Cancilla, M.T.; Lebrilla, C.B. Oceanapiside, an Antifungal Bis-α,ω-amino Alcohol Glycoside from the Marine Sponge Oceanapia phillipensis. J. Nat. Prod. 1999, 62, 1678–1681. [Google Scholar] [CrossRef] [PubMed]
- Dalisay, D.S.; Rogers, E.W.; Molinski, T.E. Oceanapiside, a Marine Natural Product, Targets the Sphingolipid Pathway of Fluconazole-Resistant Candida glabrata. Mar. Drugs 2021, 19, 126. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Chen, C.P.; Jou, M.L.; Lee, A.Y.L.; Lin, Y.C.; Yu, Y.P.; Huang, W.T.; Wu, S.H. Structural and DNA-binding studies on the bovine antimicrobial peptide, indolicidin: Evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Res. 2005, 33, 4053–4064. [Google Scholar] [CrossRef] [PubMed]
- Edlind, T.D.; Katiyar, S.K. Mutational Analysis of Flucytosine Resistance in Candida glabrata. Antimicrob. Agents Chemother. 2010, 54, 4733–4738. [Google Scholar] [CrossRef] [PubMed]
- Pushpanathan, M.; Rajendhran, J.; Jayashree, S.; Sundarakrishnan, B.; Jayachandran, S.; Gunasekaran, P. Direct cell penetration of the antifungal peptide, MMGP1, in Candida albicans. J. Pept. Sci. 2012, 18, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Pushpanathan, M.; Pooja, S.; Gunasekaran, P.; Rajendhran, J. Critical Evaluation and Compilation of Physicochemical Determinants and Membrane Interactions of MMGP1 Antifungal Peptide. Mol. Pharm. 2016, 13, 1656–1667. [Google Scholar] [CrossRef]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Mechanisms of the Antifungal Action of Marine Metagenome-Derived Peptide, MMGP1, against Candida albicans. PLoS ONE 2013, 8, e69316. [Google Scholar] [CrossRef]
- Morita, S.-y.; Ikeda, Y. Regulation of membrane phospholipid biosynthesis in mammalian cells. Biochem. Pharmacol. 2022, 206, 115296. [Google Scholar] [CrossRef]
- Murante, D.; Hogan, D.A. New Mitochondrial Targets in Fungal Pathogens. mBio 2019, 10, e02258-19. [Google Scholar] [CrossRef]
- Shentu, L.; Mu, Y. Research progress in mitochondrial dysfunction and its related diseases. Chin. Bull. Life Sci. 2018, 30, 87–93. [Google Scholar]
- Duvenage, L.; Munro, C.A.; Gourlay, C.W. The potential of respiration inhibition as a new approach to combat human fungal pathogens. Curr. Genet. 2019, 65, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, F.; Kontoyiannis, D.P. Mitochondrial Respiratory Pathways Inhibition in Rhizopus oryzae Potentiates Activity of Posaconazole and Itraconazole via Apoptosis. PLoS ONE 2013, 8, e63393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Thornton, C.R.; Kershaw, M.J.; Li, D.B.; Talbot, N.J. The glyoxylate cycle is required for temporal regulation of virulence by the plant pathogenic fungus Magnaporthe grisea. Mol. Microbiol. 2003, 47, 1601–1612. [Google Scholar] [CrossRef]
- Lorenz, M.C.; Fink, G.R. The glyoxylate cycle is required for fungal virulence. Nature 2001, 412, 83–86. [Google Scholar] [CrossRef]
- Yamashita, K.; Miyazaki, T.; Fukuda, Y.; Mitsuyama, J.; Saijo, T.; Shimamura, S.; Yamamoto, K.; Imamura, Y.; Izumikawa, K.; Yanagihara, K.; et al. The Novel Arylamidine T-2307 Selectively Disrupts Yeast Mitochondrial Function by Inhibiting Respiratory Chain Complexes. Antimicrob. Agents Chemother. 2019, 63, e00374-19. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Pinto, E.; Andrade, P.B.; Valentao, P. Antifungal Activity of Phlorotannins against Dermatophytes and Yeasts: Approaches to the Mechanism of Action and Influence on Candida albicans Virulence Factor. PLoS ONE 2013, 8, e72203. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Thompson, C.B. Bcl-2 proteins: Regulators of apoptosis or of mitochondrial homeostasis? Nat. Cell Biol. 1999, 1, E209–E216. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Liu, Y.; Jia, L.; Zhou, H.-M.; Kong, Y.; Yang, G.; Jiang, L.-P.; Li, Q.-J.; Zhong, L.-F. Curcurnin induces apoptosis through mitochondrial hyperpolarization and mtDNA damage in human hepatoma G2 cells. Free Radic. Biol. Med. 2007, 43, 968–975. [Google Scholar] [CrossRef]
- Chung, B.; Hwang, J.-Y.; Park, S.C.; Kwon, O.-S.; Cho, E.; Lee, J.; Lee, H.-S.; Oh, D.-C.; Shin, J.; Oh, K.-B. Inhibitory Effects of Nitrogenous Metabolites from a Marine-Derived Streptomyces bacillaris on Isocitrate Lyase of Candida albicans. Mar. Drugs 2022, 20, 138. [Google Scholar] [CrossRef]
- Feyder, S.; De Craene, J.-O.; Baer, S.; Bertazzi, D.L.; Friant, S. Membrane Trafficking in the Yeast Saccharomyces cerevisiae Model. Int. J. Mol. Sci. 2015, 16, 1509–1525. [Google Scholar] [CrossRef] [PubMed]
- Curwin, A.J.; Fairn, G.D.; McMaster, C.R. Phospholipid Transfer Protein Sec14 Is Required for Trafficking from Endosomes and Regulates Distinct trans-Golgi Export Pathways. J. Biol. Chem. 2009, 284, 7364–7375. [Google Scholar] [CrossRef] [PubMed]
- Demuyser, L.; Van Dyck, K.; Timmermans, B.; Van Dijck, P. Inhibition of Vesicular Transport Influences Fungal Susceptibility to Fluconazole. Antimicrob. Agents Chemother. 2019, 63, e01998-18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, M.; Braun, D.R.; Ericksen, S.S.; Piotrowski, J.S.; Nelson, J.; Peng, J.; Ananiev, G.E.; Chanana, S.; Barns, K.; et al. A marine microbiome antifungal targets urgent-threat drug-resistant fungi. Science 2020, 370, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, F.; Zarnowski, R.; Barns, K.; Jones, R.; Fossen, J.; Sanchez, H.; Rajski, S.R.; Audhya, A.; Bugni, T.S.; et al. Turbinmicin inhibits Candida biofilm growth by disrupting fungal vesicle-mediated trafficking. J. Clin. Investig. 2021, 131, e145123. [Google Scholar] [CrossRef]
- Mitchell, K.F.; Zarnowski, R.; Sanchez, H.; Edward, J.A.; Reinicke, E.L.; Nett, J.E.; Mitchell, A.P.; Andes, D.R. Community participation in biofilm matrix assembly and function. Proc. Natl. Acad. Sci. USA 2015, 112, 4092–4097. [Google Scholar] [CrossRef] [PubMed]
- Zarnowski, R.; Sanchez, H.; Covelli, A.S.; Dominguez, E.; Jaromin, A.; Bernhardt, J.; Mitchell, K.F.; Heiss, C.; Azadi, P.; Mitchell, A.; et al. Candida albicans biofilm-induced vesicles confer drug resistance through matrix biogenesis. PLoS Biol. 2018, 16, e2006872. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Lata Panwar, S.; Smriti. Drug resistance in yeasts—An emerging scenario. In Advances in Microbial Physiology; Academic Press: Cambridge, MA, USA, 2002; Volume 46, pp. 155–201. [Google Scholar]
- Diwischek, F.; Morschhäuser, J.; Holzgrabe, U. Cerulenin Analogues as Inhibitors of Efflux Pumps in Drug-resistant Candida albicans. Arch. Pharm. 2009, 342, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cui, Z.-F. Pleiotropic drug resistance ABC transporters in fungi. Yi chuan = Hereditas 2011, 33, 1048–1056. [Google Scholar] [CrossRef]
- Tanabe, K.; Lamping, E.; Adachi, K.; Takano, Y.; Kawabata, K.; Shizuri, Y.; Niimi, M.; Uehara, Y. Inhibition of fungal ABC transporters by unnarmicin A and unnarmicin C, novel cyclic peptides from marine bacterium. Biochem. Biophys. Res. Commun. 2007, 364, 990–995. [Google Scholar] [CrossRef]
- DiGirolamo, J.A.; Li, X.-C.; Jacob, M.R.; Clark, A.M.; Ferreira, D. Reversal of Fluconazole Resistance by Sulfated Sterols from the Marine Sponge Topsentia sp. J. Nat. Prod. 2009, 72, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhou, X.; Ren, B.; Cheng, L. The regulation of hyphae growth in Candida albicans. Virulence 2020, 11, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Dalle, F.; Waechtler, B.; L’Ollivier, C.; Holland, G.; Bannert, N.; Wilson, D.; Labruere, C.; Bonnin, A.; Hube, B. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell. Microbiol. 2010, 12, 248–271. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.S.; Brunke, S.; Albrecht, A.; Thewes, S.; Laue, M.; Edwards, J.E., Jr.; Filler, S.G.; Hube, B. The Hyphal-Associated Adhesin and Invasin Als3 of Candida albicans Mediates Iron Acquisition from Host Ferritin. PLoS Pathog. 2008, 4, e1000217. [Google Scholar] [CrossRef] [PubMed]
- Staab, J.F.; Bradway, S.D.; Fidel, P.L.; Sundstrom, P. Adhesive and mammalian transglutaminase substrate properties of Candida albicans Hwp1. Science 1999, 283, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Su, C.; Liu, H. Candida albicans hyphal initiation and elongation. Trends Microbiol. 2014, 22, 707–714. [Google Scholar] [CrossRef]
- Kim, H.; Hwang, J.-Y.; Chung, B.; Cho, E.; Bae, S.; Shin, J.; Oh, K.-B. 2-Alkyl-4-hydroxyquinolines from a Marine-Derived Streptomyces sp. Inhibit Hyphal Growth Induction in Candida albicans. Mar. Drugs 2019, 17, 133. [Google Scholar] [CrossRef] [PubMed]
- El Zawawy, N.A.; El-Shenody, R.A.; Ali, S.S.; El-Shetehy, M. A novel study on the inhibitory effect of marine macroalgal extracts on hyphal growth and biofilm formation of candidemia isolates. Sci. Rep. 2020, 10, 9339. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, R.; Banu, S.F.; Sowndarya, J.; Parveen, J.H.S.; Rubini, D.; Wilson, A.; Nithyanand, P. Biosurfactant synergized with marine bacterial DNase disrupts polymicrobial biofilms. Folia Microbiol. 2021, 66, 831–842. [Google Scholar] [CrossRef]
- Faria Luiz, R.L.; Machado Vila, T.V.; Palazzo de Mello, J.C.; Nakamura, C.V.; Rozental, S.; Ishida, K. Proanthocyanidins polymeric tannin from Stryphnodendron adstringens are active against Candida albicans biofilms. BMC Complement. Altern. Med. 2015, 15, 68. [Google Scholar] [CrossRef]
- Li, R.; Wang, Z.; Du, Y. Recent progress in Candida albicans biofilm. Acta Microbiol. Sin. 2017, 57, 1206–1218. [Google Scholar]
- Deveau, A.; Hogan, D.A. Linking Quorum Sensing Regulation and Biofilm Formation by Candida albicans. In Quorum Sensing: Methods and Protocols; Rumbaugh, K.P., Ed.; Humana: Totowa, NJ, USA, 2011; Volume 692, pp. 219–233. [Google Scholar]
- Kovács, R.; Bozó, A.; Gesztelyi, R.; Domán, M.; Kardos, G.; Nagy, F.; Tóth, Z.; Majoros, L. Effect of caspofungin and micafungin in combination with farnesol against Candida parapsilosis biofilms. Int. J. Antimicrob. Agents 2016, 47, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, F.; Chen, H.-Y.; Peng, H.; Hao, H.; Wang, K.-J. A Novel Antimicrobial Peptide Scyreprocin From Mud Crab Scylla paramamosain Showing Potent Antifungal and Anti-biofilm Activity. Front. Microbiol. 2020, 11, 536261. [Google Scholar] [CrossRef] [PubMed]
- Junior, A.C.V.; de Castro Nogueira Diniz Pontes, M.; Barbosa, J.P.; Hofling, J.F.; Araujo, R.M.; Boniek, D.; de Resende Stoianoff, M.A.; Andrade, V.S. Antibiofilm and Anti-Candidal Activities of the Extract of the Marine Sponge Agelas dispar. Mycopathologia 2021, 186, 819–832. [Google Scholar] [CrossRef]
- Wang, Y.-T.; Xue, Y.-R.; Liu, C.-H. A Brief Review of Bioactive Metabolites Derived from Deep-Sea Fungi. Mar. Drugs 2015, 13, 4594–4616. [Google Scholar] [CrossRef] [PubMed]
- El-Hossary, E.M.; Cheng, C.; Hamed, M.M.; El-Sayed Hamed, A.N.; Ohlsen, K.; Hentschel, U.; Abdelmohsen, U.R. Antifungal potential of marine natural products. Eur. J. Med. Chem. 2017, 126, 631–651. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical Research on the Bioactivity of New Marine Natural Products Discovered during the 28 Years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hang, S.; Lu, H.; Jiang, Y. Marine-Derived Metabolites Act as Promising Antifungal Agents. Mar. Drugs 2024, 22, 180. https://doi.org/10.3390/md22040180
Hang S, Lu H, Jiang Y. Marine-Derived Metabolites Act as Promising Antifungal Agents. Marine Drugs. 2024; 22(4):180. https://doi.org/10.3390/md22040180
Chicago/Turabian StyleHang, Sijin, Hui Lu, and Yuanying Jiang. 2024. "Marine-Derived Metabolites Act as Promising Antifungal Agents" Marine Drugs 22, no. 4: 180. https://doi.org/10.3390/md22040180
APA StyleHang, S., Lu, H., & Jiang, Y. (2024). Marine-Derived Metabolites Act as Promising Antifungal Agents. Marine Drugs, 22(4), 180. https://doi.org/10.3390/md22040180