Complementary UV-Absorption of Mycosporine-like Amino Acids and Scytonemin is Responsible for the UV-Insensitivity of Photosynthesis in Nostoc flagelliforme

Abstract
:
1. Introduction
2. Results and Discussion
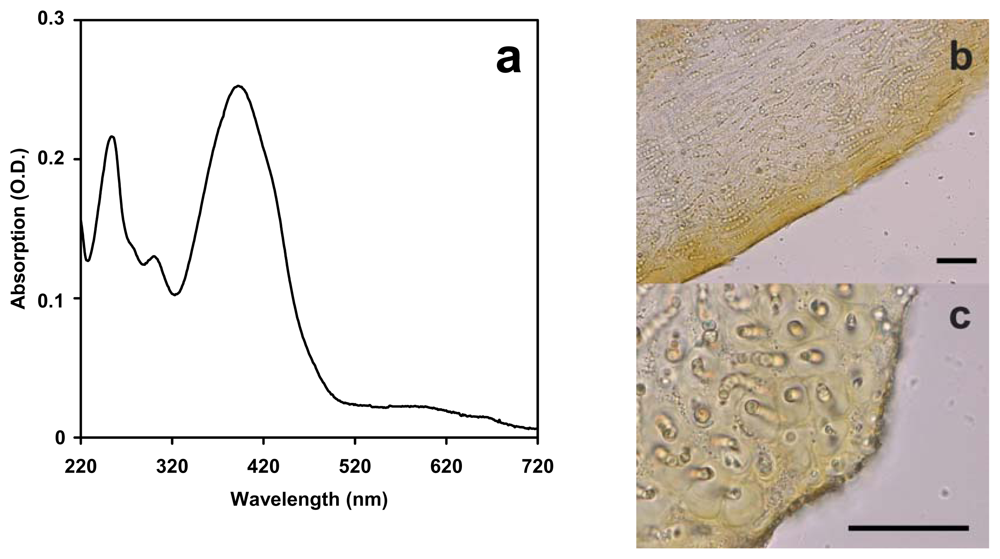
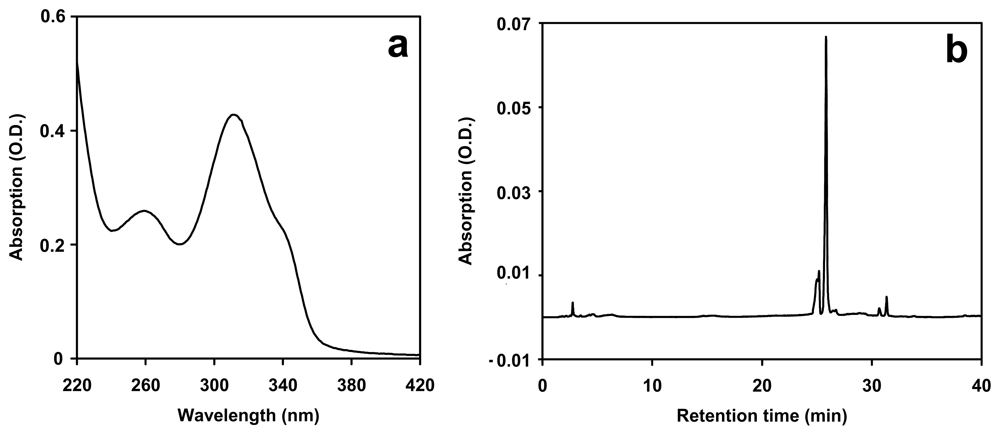
2.1. Complement of UV-absorbing compounds in Nostoc flagelliforme
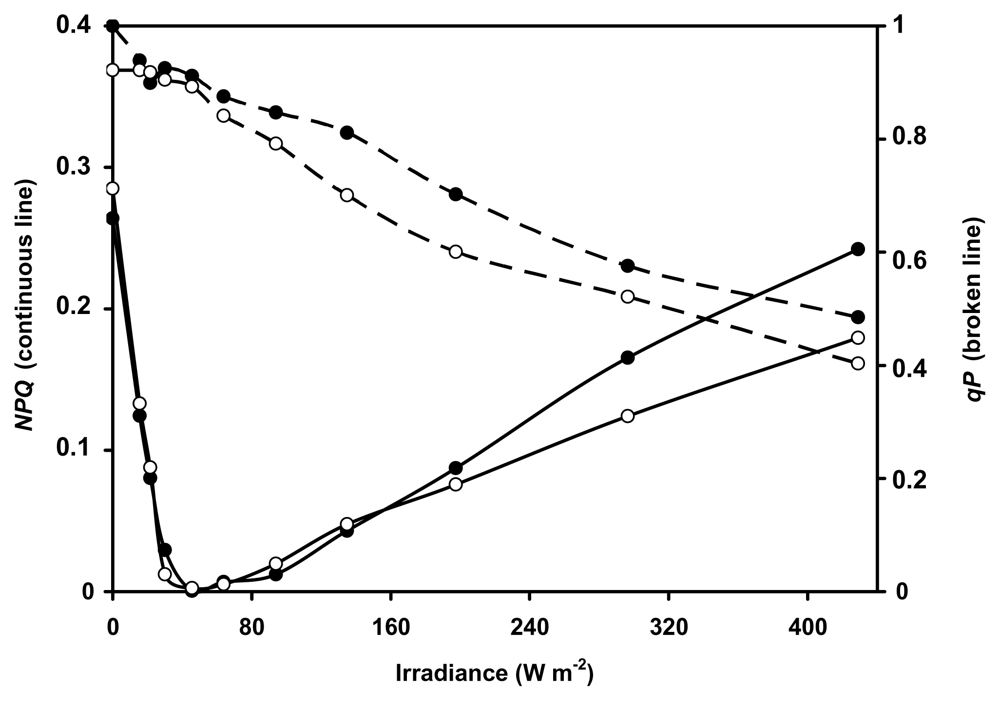
2.2. Fluorescence quenching analysis
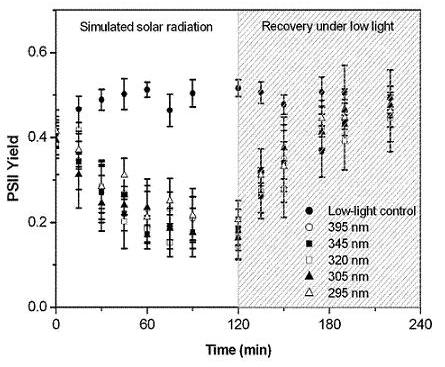
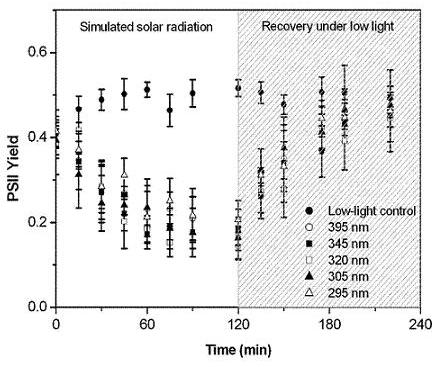
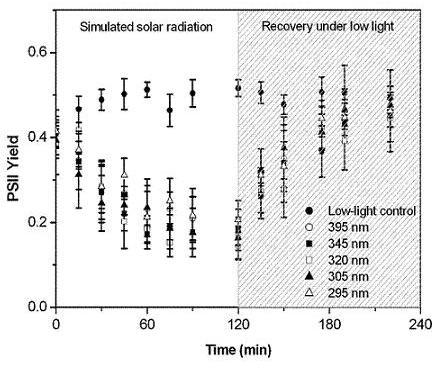
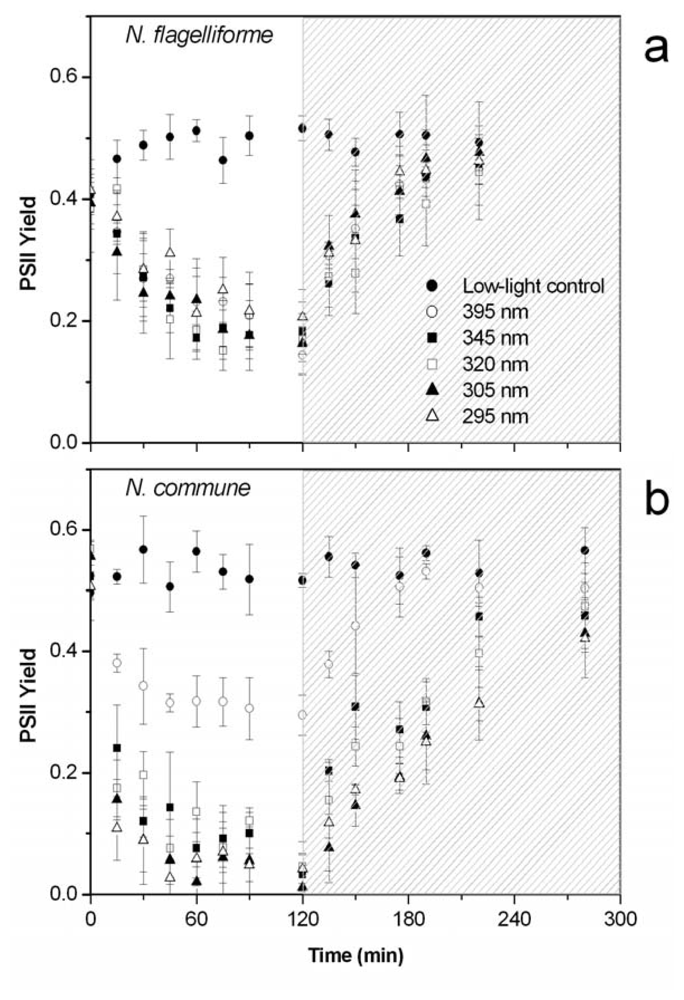
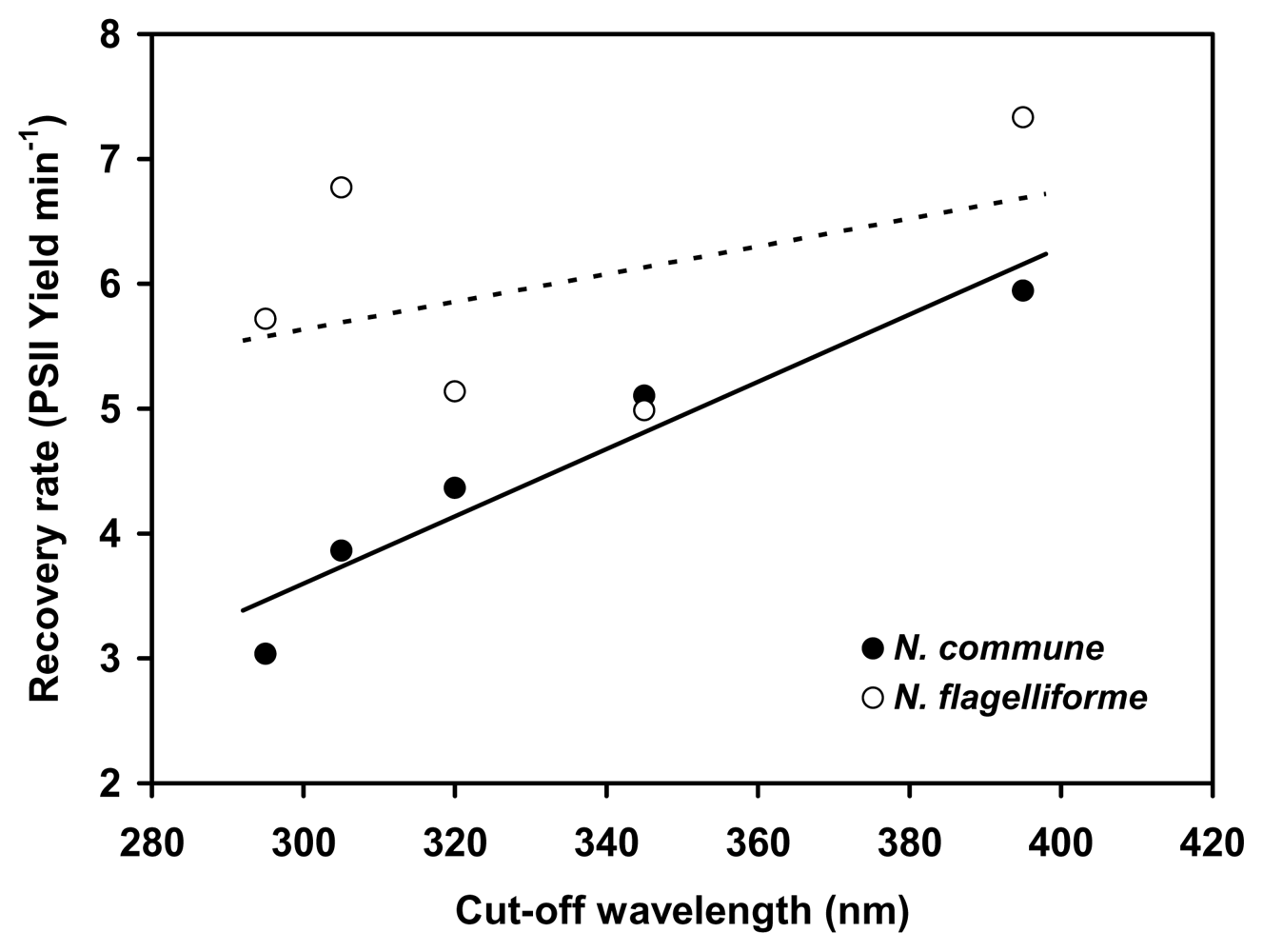
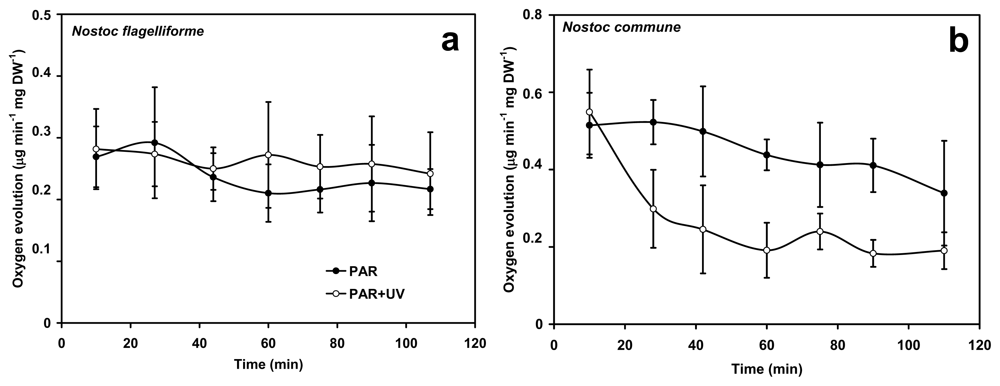
2.3. Protection of PSII activity in Nostoc flagelliforme in the UV-radiation range
3. Conclusions
4. Experimental Section
4.1. Experimental organisms
4.2. Extraction and analysis of UV-absorbing compounds
4.3. Microscopy
4.4. Modulated chlorophyll fluorescence: Preparation of samples and equipment
4.5. Irradiance-response curves of chlorophyll fluorescence quenching
4.6. Effect of UV irradiation on the PSII quantum yield
4.7. Oxygen evolution measurements
4.8. Statistics
Acknowledgements
- Samples Availability: Available from the authors.
References
- Cockell, CS. Ultraviolet radiation, evolution and the π-electron system. Biol J Linn Soc Lond 1998, 63, 449–457. [Google Scholar]
- Cockell, CS; Knowland, J. Ultraviolet radiation screening compounds. Biol Rev Camb Philos Soc 1999, 74, 311–345. [Google Scholar]
- Dunlap, WC; Shick, JM. Ultraviolet radiation-absorbing mycosporine-like amino acids in coral reef organisms: a biochemical and environmental perspective. J Phycol 1998, 34, 418–430. [Google Scholar]
- Sinha, RP; Klisch, M; Gröniger, A; Häder, D-P. Ultraviolet-absorbing/screening substances in cyanobacteria, phytoplankton and macroalgae. J Photochem Photobiol B Biol 1998, 47, 83–94. [Google Scholar]
- Nakamura, H; Kobayashi, J; Hirata, Y. Separation of mycosporine-like amino acids in marine organisms using reverse-phase high performance liquid chromatography. J Chromatogr 1982, 250, 113–186. [Google Scholar]
- Klisch, M; Häder, D-P. Mycosporine-like amino acids and marine toxins—The common and the different. Mar Drugs 2008, 6, 147–163. [Google Scholar]
- Hill, DR; Hladun, SL; Scherer, S; Potts, M. Water stress proteins of Nostoc commune (Cyanobacteria) are secreted with UV-A/B-absorbing pigments and associate with 1,4-β-d-xylanxylanohydrolase activity. J Biol Chem 1994, 269, 7726–7734. [Google Scholar]
- Böhm, GA; Pfleiderer, W; Böger, P; Scherer, S. Structure of a novel oligosaccharide-mycosporine-amino acid ultraviolet A/B sunscreen pigment from the terrestrial cyanobacterium Nostoc commune. J Biol Chem 1995, 270, 8536–8539. [Google Scholar]
- Proteau, PJ; Gerwick, WH; Garcia-Pichel, F; Castenholz, R. The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Experientia 1993, 49, 825–829. [Google Scholar]
- Stevenson, CS; Capper, EA; Roshak, AK; Marquez, B; Eichman, C; Jackson, JR; Mattern, M; Gerwick, WH; Jacobs, RS; Marshall, LA. The identification and characterization of the marine natural product scytonemin as a novel antiproliferative pharmacophore. J Pharmacol Experim Therapeut 2002, 303, 858–866. [Google Scholar]
- Klisch, M; Sinha, RP; Helbling, EW; Häder, D-P. Induction of thymine dimers by solar radiation in natural freshwater phytoplankton assemblages in Patagonia, Argentina. Aquat Sci 2005, 67, 72–78. [Google Scholar]
- He, Y-Y; Klisch, M; Häder, D-P. Adaptation of cyanobacteria to UV-B stress correlated with oxidative stress and oxidative damage. Photochem Photobiol 2002, 76, 188–196. [Google Scholar]
- Tirlapur, U; Scheuerlein, R; Häder, D-P. Motility and orientation of a dinoflagellate, Gymnodinium, impaired by solar and ultraviolet radiation. FEMS Microbiol Ecol 1993, 102, 167–174. [Google Scholar]
- Barbato, R; Bergo, E; Szabo, I; Dalla Vecchia, F; Giacometti, GM. Ultraviolet B exposure of whole leaves of barley affects structure and functional organization of photosystem II. J Biol Chem 2000, 275, 10976–10982. [Google Scholar]
- Sinha, RP; Klisch, M; Vaishampayan, A; Häder, D-P. Biochemical and spectroscopic characterization of the cyanobacterium Lyngbya sp. inhabiting mango (Mangifera indica) trees: presence of an ultraviolet-absorbing pigment, scytonemin. Acta Protozool 1999, 38, 291–298. [Google Scholar]
- Gao, K; Ye, C. Photosynthetic insensitivity of the terrestrial cyanobacterium Nostoc flagelliforme to solar UV radiation while rehydrated or desiccated. J Phycol 2007, 43, 628–635. [Google Scholar]
- Gao, K. Chinese studies on the edible blue-green alga, Nostoc flagelliforme: a review. J Appl Phycol 1998, 10, 37–49. [Google Scholar]
- Qiu, B; Gao, K. Dried field populations of Nostoc flagelliforme (Cyanophyceae) require exogenous nutrients for their photosynthetic recovery. J Appl Phycol 1999, 11, 535–541. [Google Scholar]
- Gao, K; Qiu, B; Xia, J; Yu, A. Light dependency of the photosynthetic recovery of Nostoc flagelliforme. J Appl Phycol 1998, 10, 51–53. [Google Scholar]
- Qiu, B; Gao, K. Photosynthetic characteristics of the terrestrial blue-green alga, Nostoc flagelliforme. Eur J Phycol 2001, 36, 147–156. [Google Scholar]
- Qiu, B; Zhang, A; Zhou, W; Wei, J; Dong, H; Liu, Z. Effects of potassium on the photosynthetic recovery of the terrestrial cyanobacterium, Nostoc flagelliforme (Cyanophyceae) during rehydration. J Phycol 2004, 40, 323–332. [Google Scholar]
- Zhao, XM; Bi, YH; Chen, L; Hu, S; Hu, ZY. Responses of photosynthetic activity in the drought-tolerant cyanobacterium, Nostoc flagelliforme to rehydration at different temperature. J Arid Environ 2008, 72, 370–377. [Google Scholar]
- Sheffner, AL. The reduction in vitro in viscosity of mucoprotein solutions by a new mucolytic agent, N-acetyl-L-cysteine. Ann N Y Acad Sci 1963, 106, 298–310. [Google Scholar]
- Neale, PJ; Banaszak, AT; Jarriel, CR. Ultraviolet sunscreens in Gymnodinium sanguineum (Dinophyceae): mycosporine-like amino acids protect against inhibition of photosynthesis. J Phycol 1998, 34, 928–938. [Google Scholar]
- Garcia-Pichel, F; Wingard, CE; Castenholz, RW. Evidence regarding the UV sunscreen role of a mycosporine-like compound in the cyanobacterium Gloeocapsa sp. Appl Environ Microbiol 1993, 59, 170–176. [Google Scholar]
- Klisch, M; Sinha, RP; Richter, PR; Häder, D-P. Mycosporine-like amino acids (MAAs) protect against UV-B-induced damage in Gyrodinium dorsum Kofoid. J Plant Physiol 2001, 158, 1449–1454. [Google Scholar]
- Garcia-Pichel, F; Castenholz, RW. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl Environ Microbiol 1993, 59, 163–169. [Google Scholar]
- Karapetyan, NV. Protective dissipation of excess absorbed energy by photosynthetic apparatus of cyanobacteria: role of antenna terminal emitters. Photosyn Res 2008, 97, 195–204. [Google Scholar]
- Campbell, D; Hurry, V; Clarke, AK; Gustafsson, P; Öquist, G. Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol Mol Biol Rev 1998, 62, 667–683. [Google Scholar]
- Kirilovsky, D. Photoprotection in cyanobacteria: the orange carotenoid protein (OCP)-related non-photochemical-quenching mechanism. Photosyn Res 2007, 93, 7–16. [Google Scholar]
- Mullineaux, CW; Allen, JF. The state 2 transition in the cyanobacterium Synechococcus 6301 can be driven by respiratory electron flow into the plastoquinone pool. FEBS Lett 1986, 205, 155–160. [Google Scholar]
- Schubert, H; Matthijs, H; Mur, L. In vivo assay of P700 redox changes in the cyanobacterium Fremyella diplosiphon and the role of cytochrome-c-oxidase in regulation of photosynthetic electron transfer. Photosynthetica 1995, 31, 517–527. [Google Scholar]
- Häder, D-P; Lebert, M; Helbling, EW. Effects of solar radiation on the Patagonian macroalga Enteromorpha linza (L.) J. Agardh - Chlorophyceae. J Photochem Photobiol B Biol 2001, 62, 43–54. [Google Scholar]
- Bergo, E; Segalla, A; Giacometti, GM; Tarantino, D; Soave, C; Andreucci, F; Barbato, R. Role of visible light in the recovery of photosystem II structure and function from ultraviolet-B stress in higher plants. J Exp Bot 2003, 54, 1665–1673. [Google Scholar]
- Bouchard, JN; Roy, S; Campbell, DA. UVB effects on the photosystem II-D1 protein of phytoplankton and natural phytoplankton communities. Photochem Photobiol 2006, 82, 936–951. [Google Scholar]
- Rippka, R; Deruelles, J; Wazterbury, JB; Herdmann, M; Stanier, RY. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J Gen Microbiol 1979, 111, 1–61. [Google Scholar]
- Bilger, W; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosyn Res 1990, 25, 173–185. [Google Scholar]
- Genty, B; Briantais, J-M; Baker, NR. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 1989, 990, 87–92. [Google Scholar]
- Weis, E; Berry, A. Quantum efficiency of photosystem II in relation to the energy dependent quenching of chlorophyll fluorescence. Biochim Biophys Acta 1987, 894, 198–208. [Google Scholar]
- Brown, AM. A new software for carrying out one-way ANOVA post hoc tests. Comput Methods Programs Biomed 2005, 79, 89–95. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition |
|---|---|
| F0 | Minimum fluorescence in the dark-adapted state |
| F0′ | Minimum fluorescence in the light-adapted state |
| FM dark | Maximum fluorescence in the dark-adapted state |
| FM′ | Maximum fluorescence in the light-adapted state |
| FM | Maximum fluorescence estimated as the highest FM′ in irradiance-response curves |
| Ft | Steady-state fluorescence |
| NPQ | Non-photochemical chlorophyll fluorescence quenching |
| qP | Photochemical quenching of variable chlorophyll fluorescence |
| Y | Actual quantum yield of photochemical energy conversion in PSII |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ferroni, L.; Klisch, M.; Pancaldi, S.; Häder, D.-P. Complementary UV-Absorption of Mycosporine-like Amino Acids and Scytonemin is Responsible for the UV-Insensitivity of Photosynthesis in Nostoc flagelliforme. Mar. Drugs 2010, 8, 106-121. https://doi.org/10.3390/md8010106
Ferroni L, Klisch M, Pancaldi S, Häder D-P. Complementary UV-Absorption of Mycosporine-like Amino Acids and Scytonemin is Responsible for the UV-Insensitivity of Photosynthesis in Nostoc flagelliforme. Marine Drugs. 2010; 8(1):106-121. https://doi.org/10.3390/md8010106
Chicago/Turabian StyleFerroni, Lorenzo, Manfred Klisch, Simonetta Pancaldi, and Donat-Peter Häder. 2010. "Complementary UV-Absorption of Mycosporine-like Amino Acids and Scytonemin is Responsible for the UV-Insensitivity of Photosynthesis in Nostoc flagelliforme" Marine Drugs 8, no. 1: 106-121. https://doi.org/10.3390/md8010106
APA StyleFerroni, L., Klisch, M., Pancaldi, S., & Häder, D. -P. (2010). Complementary UV-Absorption of Mycosporine-like Amino Acids and Scytonemin is Responsible for the UV-Insensitivity of Photosynthesis in Nostoc flagelliforme. Marine Drugs, 8(1), 106-121. https://doi.org/10.3390/md8010106