Effects of Marine Toxins on the Reproduction and Early Stages Development of Aquatic Organisms



Abstract
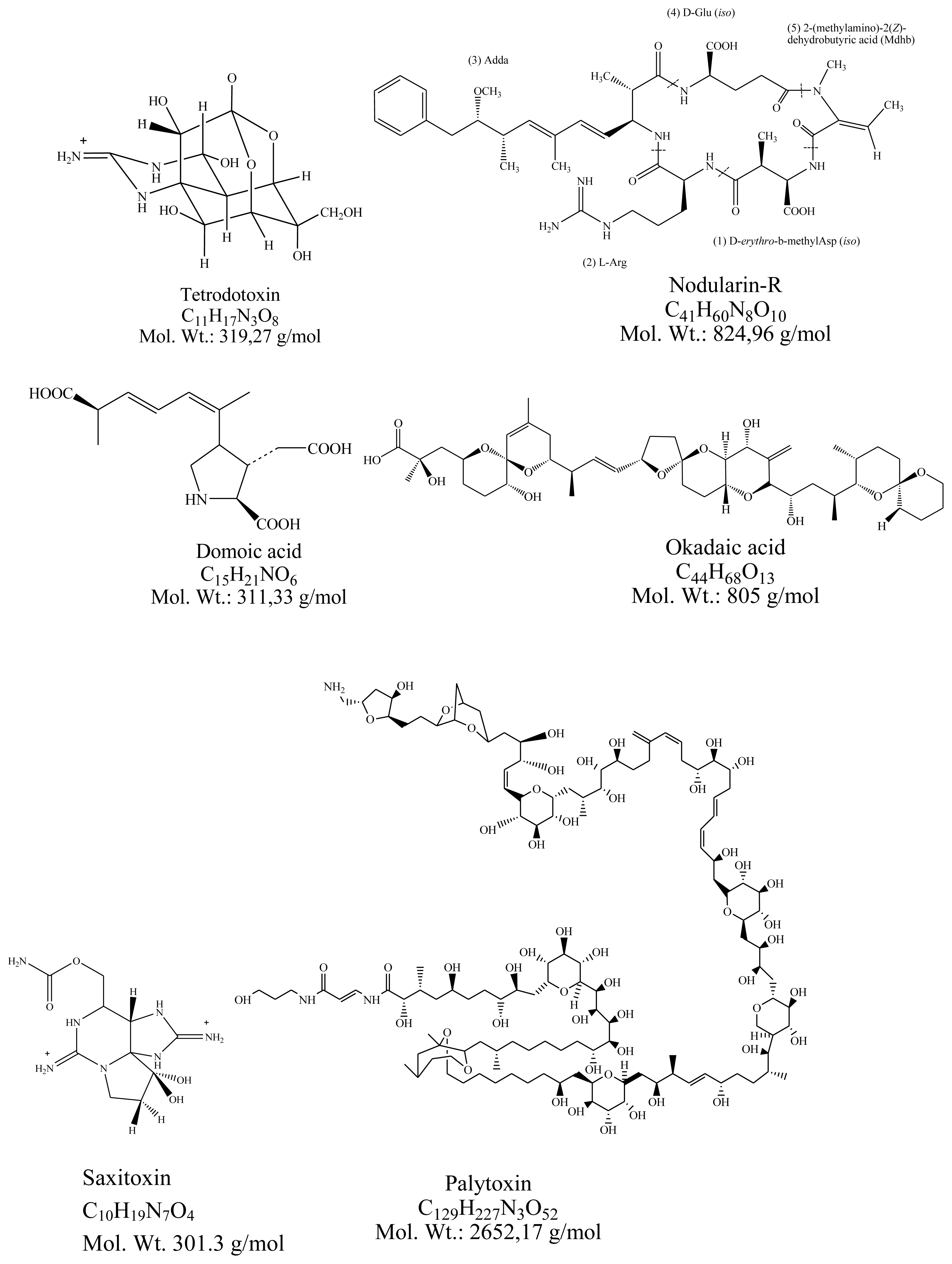
:1. Main Marine Toxins Produced by Bacteria and Phytoplankton
2. Effects of Marine Toxins in Aquatic Invertebrate Larval Development
2.1. Tetrodotoxin
2.2. Nodularin
2.3. Domoic acid
2.4. Okadaic acid
2.5. Saxitoxins
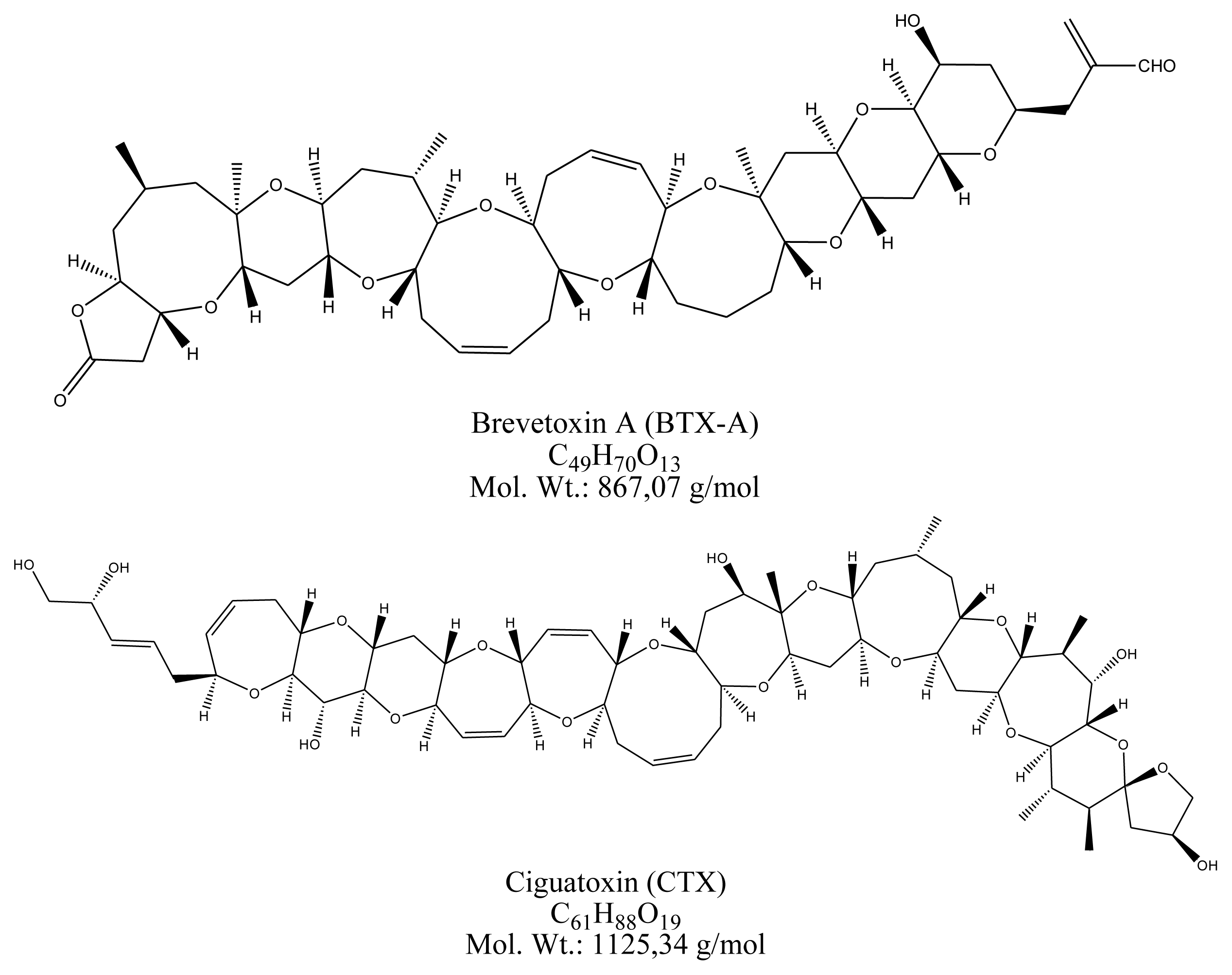
2.6. Ciguatoxin
3. Effects of Marine Toxins in Aquatic Vertebrate Early Development
3.1. Domoic acid
3.2. Okadaic acid
3.3. Saxitoxins
3.4. Palitoxin
3.5. Brevetoxin
3.6. Ciguatoxin
4. Conclusions
Acknowledgements
References
- Kao, CY. Structure-activity relations of tetrodotoxin, saxitoxin and analogues. Ann N Y Acad Sci 1986, 479, 52–67. [Google Scholar]
- Scheib, H; McLay, I; Guex, N; Clare, J; Blaney, F; Dale, T; Tate, S; Robertson, G. Modeling the pore structure of voltage-gated sodium channels in closed, open, and fast-inactivated conformation reveals details of site 1 toxin and local anesthetic binding. J Mol Model 2006, 12, 813–822. [Google Scholar]
- Lee, MJ; Jeong, D-Y; Kim, W-S; Kim, H-D; Kim, C-H; Park, W-W; Kim, K-S; Kim, H-M; Kim, D-S. A tetrodotoxin-producing Vibrio strain LM-1, from the Puffer fish Fugu vermicularis radiates. Appl Environ Microbiol 2000, 66, 1698–1701. [Google Scholar]
- Yu, C-F; Yu, PH-F; Chan, P-L; Yan, Q; Wong, P-K. Two novel species of tetrodotoxin-produciong bacteria isolated from toxic marine puffer fishes. Toxicon 2004, 44, 641–647. [Google Scholar]
- Yan, Q; Yu, PHF; Li, H-Z. Detection of tetrodotoxin and bacterial production by Serratia marcescens. World J Microbiol Biotechnol 2005, 21, 1255–1258. [Google Scholar]
- Maruyama, J; Noguchi, T; Narita, H; Jeon, JK; Otsuka, M; Hashimoto, K. Occurrence of tetrodotoxin in a starfish, Astropecten scoparius. Agric Biol Chem 1985, 49, 3069–3070. [Google Scholar]
- Noguchi, T; Uzu, A; Koyama, K; Hashimoto, K. Occurrence of tetrodotoxin as the major toxinin xanthid crab Atergatis floridus. Bull Jpn Soc Sci Fish 1983, 49, 1887–1892. [Google Scholar]
- Hanifin, CT; Brodie, ED, 3rd; Brodie, ED, Jr. Tetrodotoxin levels of the rough-skin newt, Taricha granulosa, increase in long-term captivity. Toxicon 2002, 40, 1149–1153. [Google Scholar]
- Honkanen, RE; Dukelow, M; Zwiller, J; Moore, RE; Khatra, BS; Boynton, AL. Cyanobacterial nodularin is a potent inhibitor of type 1 and type 2A protein phosphatases. Mol Pharmacol 1991, 40, 577–583. [Google Scholar]
- Tiedeken, JA; Ramsdell, JS; Ramsdell, AF. Developmental toxicity of domoic acid in zebrafish (Danio rerio). Neurotoxicol Teratol 2005, 27, 711–717. [Google Scholar]
- Bates, SS; Gaudet, J; Kaczmarska, I; Ehrman, JM. Interaction between bacteria and the domoic-acid-producing diatom Pseudo-nitzschia multiseries (Hasle) Hasle; can bacteria produce domoic acid autonomously? Harmful Algae 2004, 3, 11–20. [Google Scholar]
- Tanti, JF; Grémeaux, T; van Obberghen, E; Le Marchand-Brustel, Y. Effects of okadaic acid, an inhibitor of protein phosphatases-1 and -2A, on glucose transport and metabolism in skeletal muscle. J Biol Chem 1991, 266, 2099–2103. [Google Scholar]
- Boczar, BA; Beitler, MK; Liston, J; Sullivan, J; Cattolico, RA. Paralytic shellfish toxins in Protogonyaulax tamarensis and P. catenella in axenic culture. Plant Physiol 1988, 88, 1285–1290. [Google Scholar]
- Kan, Y; Uemura, D; Hirata, Y; Ishiguro, M; Iwashita, T. Complete NMR signal assignment of palytoxin and N-acetylpalytoxin. Tetrahedron Lett 2001, 42, 3197–3202. [Google Scholar]
- Katikou, P. Botana, LM, Ed.; Chemistry of Palytoxins and Ostreocins. In Phycotoxins, Chemistry and Biochemistry; Blackwell Publishing: Ames, IA, USA, 2007; pp. 75–93. [Google Scholar]
- Satoh, E; Ishii, T; Nishimura, M. Palytoxin-induced increase in cytosolic-free Ca2+ in mouse spleen cells. Eur J Pharmacol 2003, 465, 9–13. [Google Scholar]
- Valverde, I; Lago, J; Vieites, JM; Cabado, AG. In vitro approaches to evaluate palytoxin-induced toxicity and cell death in intestinal cells. J Appl Toxicol 2008, 28, 294–302. [Google Scholar]
- Colman, JR; Ramsdell, JS. The type B Brevetoxin (PbTx-3) adversely affects development, cardiovascular function, and survival in Medaka (Oryzias latipes) embryos. Environ Health Perspect 2003, 111, 1920–1925. [Google Scholar]
- Bagnis, R; Chanteau, S; Chungue, E; Hurtel, JM; Yasumoto, T; Inoue, A. Origins of ciguatera fish poisoning: a new dinoflagellate, Gambierdiscus toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 18, 199–208. [Google Scholar]
- Labrusse, H; Matile, L. Toxicological biotest on diptera larvae to detect ciaguatoxins and various other toxic substances. Toxicon 1996, 34, 881–891. [Google Scholar]
- McMahon, BR; Tanaka, K; Doyle, JE; Chu, KH. A change of heart: cardiovascular development in the shrimp Metapenaeus ensis. Comp Biochem Physiol, Part A: Mol Integr Physiol 2002, 133, 577–587. [Google Scholar]
- Lopes, V; Antunes, A; Martins, R; Welker, M; Vasconcelos, V. Morphological, toxicological and molecular characterization of a benthic Nodularia strain isolated from a Portuguese estuary. Res Microbiol 2009. [Google Scholar] [CrossRef]
- Korpinen, S; Karjalainen, M; Viitasalo, M. Effects of cyanobacteria on survival and reproduction of the littoral crustacean Gammarus zaddachi (Amphipoda). Hydrobiologia 2006, 559, 285–295. [Google Scholar]
- Koski, M; Engstrom, J; Viitasalo, M. Reproduction and survival of the calanoid copepod Eurytemora affinis fed with toxic and non-toxic cyanobacteria. Mar Ecol Prog Ser 1999, 186, 187–197. [Google Scholar]
- Koski, M; Schmidt, K; Engstrom.Ost, J; Viitasalo, M; Jónasdóttir, S; Repka, S; Sivonen, K. Calanoid copepods fed and produce eggs in the presence of toxic cyanobacteria Nodularia spumigena. Limnol Oceanogr 2002, 47, 878–885. [Google Scholar]
- Ojaveer, E; Simm, M; Balode, M; Purina, I; Suursaar, U. Effect of Microcystis aeruginosa and Nodularia spumigena on survival of Eurytemora affinis and the embryonic and larval development of the Baltic herring Clupea harengus membras. Environ Toxicol 2003, 18, 236–242. [Google Scholar]
- Kozlowsky-Suzuki, B; Koski, M; Hallberg, E; Wallén, R; Carlsson, P. Glutathione transferase activity and oocyte development in copepods exposed to toxic phytoplankton. Harmful Algae 2009, 8, 395–406. [Google Scholar]
- Beattie, KA; Ressler, J; Wiegand, C; Kraiuse, E; Codd, GA; Steinberg, CEW; Pflugmacher, S. Comparative effects and metabolism of two microcystins and nodularin in the brine shrimp Artemia salina. Aquat Toxicol 2003, 62, 219–226. [Google Scholar]
- Maneiro, I; Iglesias, P; Guisande, C; Riveiro, I; Barreiro, A; Zervoudaki, S; Granéli, E. Fate of domoic acid ingested by the copepod Acartia clausi. Mar Biol 2005, 148, 123–130. [Google Scholar]
- Liu, H; Kelly, MS; Campbell, DA; Dong, SL; Zhu, JX; Wang, SF. Exposure to domoic acid affects larval development of king scallop Pecten maximus (Linnaeus, 1758). Aquat Toxicol 2007, 81, 152–158. [Google Scholar]
- Liu, H; Kelly, MS; Campbell, DA; Fang, J; Zhu, J. Accumulation of domoic acid and its effects on juvenile king scallop Pecten maximus (Linnaeus, 1758). Aquaculture 2008, 284, 224–230. [Google Scholar]
- Perez, MF; Sulkin, SD. Palatability of autotrophic dinoflagellates to newly hatched larval crabs. Mar Biol 2005, 46, 771–780. [Google Scholar]
- Ajuzie, CC. Palatability and fatality of the dinoflagellate Prorocentrum lima to Artemia salina. J Appl Phycol 2007, 19, 513–519. [Google Scholar]
- Bag⊘ien, E; Miranda, A; Reguera, B; Franco, JM. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Effect of the toxic dinoflagellate Alexandrium minutum on the copepod Euterpina acutifrons. In Harmful and Toxic Algal Blooms; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 385–388. [Google Scholar]
- Zhenxing, W; Yinglin, Z; Mingyuan, Z; Zongling, W; Dan, W. Effects of toxic Alexandrium species on the surbvival and feeding rates of brine shrimp Artemia salina. Acta Ecol Sin 2006, 26, 3942–3947. [Google Scholar]
- Robineau, B; Gagné, JA; Fortier, L; Cembella, AD. Potential impact of a toxic dinoflagellate (Alexandrium excavatum) bloom on survival of fish and crustacean larvae. Mar Biol 1991, 108, 293–301. [Google Scholar]
- Hinz, S; Sulkin, S; Strom, S; Testermann, J. Discrimination in ingestion of protistan prey by larval crabs. Mar Ecol Prog Ser 2001, 222, 155–162. [Google Scholar]
- Sulkin, S; Hinz, S; Rodriguez, M. Effects of the exposure to toxic and non-toxic dinoflagellates on oxygen consumption and locomotion in stage 1 larvae of the crabs Cancer oregonensis and C. magister. Mar Biol 2003, 142, 205–211. [Google Scholar]
- Kelly, AM; Kohler, CC; Tindall, DR. Are crustaceans linked to the ciguatera food chain? Environ Biol Fishes 1992, 33, 275–286. [Google Scholar]
- Bejarano, AC; Gulland, FM; Goldstein, T; Leger, J; Hunter, M; Schwacke, LH; VanDolah, FM; Rowles, TK. Demographics and spatio-temporal signature of the biotoxin domoic acid in California sea lion (Zalophus californianus) stranding records. Mar Mamm Sci 2008, 24, 899–912. [Google Scholar]
- Oberemm, A; Becker, J; Codd, GA; Steinber, C. Effects of cyanobacteria toxins and aqueous crude extracts of cyanobacteria on the development of fish and amphibians. Environ Toxicol 1999, 14, 77–88. [Google Scholar]
- Edmunds, JSG; McCarthy, RA; Ramsdell, JS. Ciguatoxin reduces larval survivability in finfish. Toxicon 1999, 37, 1827–1832. [Google Scholar]
- Escoffier, N; Gaudin, J; Mezhoud, K; Huet, H; Chateau-Joubert, S; Turquet, J; Crespeau, F; Edery, M. Toxicity to medaka fish embryo development of okadaic acid and crude extracts of Prorocentrum dinoflagellates. Toxicon 2007, 49, 1182–1192. [Google Scholar]
- Casarini, L; Franchini, A; Malagoli, D; Ottaviani, E. Evaluation of the effects of the marine toxin okadaic acid by using FETAX assay. Toxicol Lett 2007, 169, 145–151. [Google Scholar]
- Terao, K; Ito, E; Yanagi, T; Yasumoto, T. Histopathological studies on experimental marine toxin poisoning. I. Ultrastructural changes in the small intestine and liver of suckling mice induced by dinophysistoxin-1 and pectenotoxin-1. Toxicon 1986, 24, 1141–1151. [Google Scholar]
- Franchini, A; Casarini, L; Malagoli, D; Ottaviani, E. Expression of the genes siamois, engrailed-2, bmp4 and myf5 during Xenopus development in the presence of the marine toxins okadaic acid and palytoxin. Chemosphere 2009, 77, 308–312. [Google Scholar]
- Gosselin, S; Fortier, L; Gagné, JA. Vulnerability of marine fish larvae to the toxic dinoflagellate Protogonyaulax tamarensis. Mar Ecol Prog Ser 1989, 57, 1–10. [Google Scholar]
- Lefebvre, KA; Trainer, VL; Scholz, NL. Morphological abnormalities and sensorimotor deficits in larval fish exposed to dissolved saxitoxin. Aquat Toxicol 2004, 66, 159–170. [Google Scholar]
- Lefebvre, KA; Elder, NE; Hershberger, PK; Trainer, VL; Stehr, CM; Scholz, NL. Dissolved saxitoxin causes transient inhibition of sensorimotor function in larval Pacific herring (Clupea harengus pallasi). Mar Biol 2005, 147, 1393–1402. [Google Scholar]
- Franchini, A; Casarini, L; Ottavianio, E. Toxicological effects of marine palytoxin evaluated by FETAX assay. Chemosphere 2008, 73, 267–271. [Google Scholar]
- Nusse, R. Wnt signaling in disease and in development. Cell Res 2005, 15, 28–32. [Google Scholar]
- Poli, MA; Lewis, RJ; Dickey, RW; Musser, SM; Buckner, CA; Carpenter, LG. Identification of Caribbean ciguatoxins as the cause of an outbreak of fish poisoning among US soldiers in Haiti. Toxicon 1997, 35, 733–741. [Google Scholar]
- Colman, JR; Dechraoui, M-YB; Dickey, RW; Ramsdell, JS. Characterization of the developmental toxicity of Caribbean ciguatoxins in finfish embryos. Toxicon 2004, 44, 59–66. [Google Scholar]



{kind=link}
{kind=link}
| Toxin | Example of producing organism | Chemical type | Target/action |
|---|---|---|---|
| Tetrodotoxin | Bacteria Vibrio sp. Serratia marcescens | Alkaloid | Na+ channels/blockage |
| Nodularin | Cyanobacteria Nodularia spumigena | Cyclic peptide | Protein Phosphatases/inhibition |
| Domoic acid | Diatoms Pseudo-nitzschia spp. | Amino acid | Glutamate receptor/agonist |
| Okadaic acid | Dinoflagellates Dynophysis spp. | Polyether | Protein Phosphatases/inhibition |
| Saxitoxins | Alexandrium spp. | Alkaloid | Na+ channels/blockage |
| Palytoxin | Ostreopsis ovata | Aliphatic backbone with polyethers | Na+/K+ pumps/pore forming |
| Brevetoxins | Karenia brevis | Polyethers | Na+ channels/ |
| Ciguatoxin | Gambierdiscus toxicus | Polycyclic ethers | Na+ channels/ |
| Toxin | Invertebrate species | Concentrations tested or cell or filaments density | Toxicity | Reference |
|---|---|---|---|---|
| NOD | Eurytemora affinis | 0.58 μg NOD mg−1 1.85–22.2 × 103 cells mL−1 | 44–72% mortality | [24] |
| 0.11 μg NOD mg−1 0.5 × 103 fil mL−1 | TL50–2.5 days | [26] | ||
| E. affinis Acartia bifilosa | 9.2–13.0 μg NOD L−1 | n.s.l. | [25] | |
| A. tonsa Temora longicornis | 0.35 pg NOD cell−1 300 μg C L−1 | n.s.l. | [27] | |
| Gammarus zaddachi | 12–101 μg NOD L−1 | TL50–1 day (41 μg NOD L−1) | [23] | |
| DA | A. clausi | 0.31 pg DA cell−1 1.9 × 104 cell mL−1 | n.s.l. | [29] |
| Pecten maximus | 50 ng DA mL−1 | 18.3% survivorship not significantly different from control | [30] | |
| OA | Cancer magister, C. oregonensis, C. productus, C. gracilis, Hemigrapsus nudus, H. oregonensis | 103 cells mL−1 | Cells not ingested n.s.l. | [32] |
| Artemia salina | n.a. (cell free media) | LT50–1.7 h | [33] | |
| STX | A. salina | 2,000 cells mL−1 | LT50–12 h | [35] |
| Homarus americanus | n.a. via contaminated zooplankton | Maximum lethality–43% | [36] | |
| C. magister, C. oregonensis, H. oregonensis, Rhinolithodes wosnessenskii | 1,000 cells mL−1 | n.s.l. | [37] | |
| C. oregonensis, C. magister | 5 × 102 cells mL−1 (filtrate) | Reduction of oxygen consumption | [38] | |
| CTX | Artemia spp. | 2–1,000 cells Artemia−1 | LD50–2.9 to 42.6 cells | [39] |
| Toxin | Fish species | Concentrations tested | Toxicity | Reference |
|---|---|---|---|---|
| DA | Danio rerio | 0.16–23.8 ng DA egg−1 * | EC50 0.56–1.68 ng DA egg−1 * | [10] |
| STX | 10–500 μg STX L−1 | 21 d LC50 500 μg STX L−1 | [41] | |
| Clupea harengus | 50–1600 μg STX L−1 | Not significant | [49] | |
| BTX | Oryzias latipes | 1–8.5 ng PbTX-3 egg−1 | LD50 4 ng PbTX-3 egg−1 | [18] |
| CTX | 0.1–20 pg CTX egg−1 | LD50 1–9 pg CTX egg−1 | [42] | |
| OA | 0.1–1 μg OA mL−1 | LD50 0.52 μg OA mL−1 | [43] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vasconcelos, V.; Azevedo, J.; Silva, M.; Ramos, V. Effects of Marine Toxins on the Reproduction and Early Stages Development of Aquatic Organisms. Mar. Drugs 2010, 8, 59-79. https://doi.org/10.3390/md8010059
Vasconcelos V, Azevedo J, Silva M, Ramos V. Effects of Marine Toxins on the Reproduction and Early Stages Development of Aquatic Organisms. Marine Drugs. 2010; 8(1):59-79. https://doi.org/10.3390/md8010059
Chicago/Turabian StyleVasconcelos, Vítor, Joana Azevedo, Marisa Silva, and Vítor Ramos. 2010. "Effects of Marine Toxins on the Reproduction and Early Stages Development of Aquatic Organisms" Marine Drugs 8, no. 1: 59-79. https://doi.org/10.3390/md8010059
APA StyleVasconcelos, V., Azevedo, J., Silva, M., & Ramos, V. (2010). Effects of Marine Toxins on the Reproduction and Early Stages Development of Aquatic Organisms. Marine Drugs, 8(1), 59-79. https://doi.org/10.3390/md8010059