Primary Screening of the Bioactivity of Brackishwater Cyanobacteria: Toxicity of Crude Extracts to Artemia salina Larvae and Paracentrotus lividus Embryos




Abstract
:1. Introduction
2. Results and Discussion
3. Experimental Section
3.1. Cyanobacteria sampling
3.2. Taxonomic characterization
3.3. Preparation of extracts
3.4. Preparation of the bioassays
3.5. A. salina lethality assay
3.6. P. lividus embryo-larval toxicity assay
3.7. Statistical analyses
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References and Notes
- Singh, S; Kate, BN; Banerjee, UC. Bioactive compounds from cyanobacteria and microalgae: an overview. Crit Rev Biotechnol 2005, 25, 73–95. [Google Scholar]
- Jaiswal, P; Singh, PK; Prasanna, R. Cyanobacterial bioactive molecules--an overview of their toxic properties. Can J Microbiol 2008, 54, 701–717. [Google Scholar]
- Sánches-Fortún, S; Sanz, S; Barahona, MV. Acute toxicity of several organophosphorous insecticides and protection by cholinergic antagonist and 2-PAM on Artemia salina larvae. Arch Environ Contam Toxicol 1996, 31, 391–398. [Google Scholar]
- Caldwell, GS; Bentley, MG; Olive, PJW. The use of a brine shrimp (Artemia salina) bioassay to assess the toxicity of diatom extracts and short chain aldehydes. Toxicon 2003, 42, 301–306. [Google Scholar]
- Vanhaecke, P; Persoone, G; Claus, C; Sorgeloos, P. Proposal for a short-term toxicity test with Artemia nauplii. Ecotoxicol Environ Saf 1981, 5, 382–387. [Google Scholar]
- Sleet, RB; Brendel, K. Brine shrimp, Artemia salina: a potential screening organism for initial teratology screening. Proc West Pharmacol Soc 1983, 26, 169–170. [Google Scholar]
- Carballo, JL; Hernandez-Inda, ZL; Perez, P; Garcia-Gravalos, MD. A comparison between two brine shrimp assays to detect in vitro cytotoxicity in marine natural products. BMC Biotechnol 2002, 2, 17. [Google Scholar]
- Nunes, BS; Carvalho, FD; Guilhermino, LM; Van Stappen, G. Use of the genus Artemia in ecotoxicity testing. Environ Pollut 2006, 14, 453–462. [Google Scholar]
- Krueger, RJ. Book Review of Bioactive Natural Products. Detection, Isolation, and Structural Determination. Second Edition. J Med Chem 2008, 51, 3659–3659. [Google Scholar]
- Boudouresque, CF; Verlaque, M. Ecology of Paracentrotus lividus. In Edible Sea Urchins: Biology and Ecology; Lawrence, JM, Ed.; Elsevier publ.: Amsterdam, The Netherlands, 2007; pp. 243–285. [Google Scholar]
- Martins, R; Fernandez, N; Beiras, R; Vasconcelos, V. Toxicity assessment of crude and partially purified extracts of marine Synechocystis and Synechococcus cyanobacterial strains in marine invertebrates. Toxicon 2007, 50, 791–799. [Google Scholar]
- Ehrenreich, IM; Waterbury, JB; Webb, EA. Distribution and diversity of natural product genes in marine and freshwater cyanobacterial cultures and genomes. Appl Environ Microbiol 2005, 71, 7401–7413. [Google Scholar]
- Alexandra, AR; Janine, NC; Mohamed, AM; Brett, AN. The Synechocystis sp. PCC6803 Sfp-Type Phosphopantetheinyl Transferase Does Not Possess Characteristic Broad-Range Activity. ChemBioChem 2009, 10, 1869–1877. [Google Scholar]
- Sugni, M; Mozzi, D; Barbaglio, A; Bonasoro, F; Candia Carnevali, MD. Endocrine disrupting compounds and echinoderms: New ecotoxicological sentinels for the marine ecosystem. Ecotoxicology 2007, 16, 95–108. [Google Scholar]
- Da Rocha, AB; Lopes, R; Schwartsmann, G. Natural products in anticancer therapy. Curr Opin Pharmacol 2001, 364–369. [Google Scholar]
- Wiegand, C; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites: A short review. Toxicol Appl Pharmacol 2005, 203, 201–218. [Google Scholar]
- Mundt, S; Kreitlow, S; Nowotny, A; Effmert, U. Biochemical and pharmacological investigations of selected cyanobacteria. Int J Hyg Environ Health 2001, 203, 327–334. [Google Scholar]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae; Norwegian Institute for Water Research: Blindern, Oslo, Norway, 1972; p. 5. [Google Scholar]
- Ripka, R. Isolation and purification of cyanobacteria. Methods Enzymol 1988, 167, 3–28. [Google Scholar]
- Komárek, J; Anagnostidis, K. Cyanoprokaryota, Part 1: Chroococcales, Süsswasserflora von Mitteleuropa, Bd 19/1; Gustav Fischer Verlag: Stuttgart, Germany, 1998; p. 548. [Google Scholar]
- Komárek, J; Anagnostidis, K. Cyanoprokaryota, Part 2: Oscillatoriales, Süsswasserflora von Mitteleuropa, Bd19/2; Elsevier/Spektrum Akademischer Verlag: Heidelberg, Germany, 2005; p. 759. [Google Scholar]
- Boone, DR; Castenholz, RW. Bergey’s Manual of Systematic Bacteriology; Garrity, GM, Ed.; Springer-Verlag: New York, NY, USA, 2001; Volume 1, p. 600. [Google Scholar]
- Jungblut, A.-D; Neilan, BA. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch Microbiol 2006, 185, 107–114. [Google Scholar]
- Neilan, BA; Jacobs, D; Del Dot, T; Blackall, LL; Hawkins, PR; Cox, PT; Goodman, AE. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus. Microcystis Int J Syst Bacteriol 1997, 47, 693–697. [Google Scholar]
- Fernández, N; Beiras, R. Combined toxicity of dissolved mercury with copper, lead and cadmium on embryogenesis and early larval growth of the Paracentrotus lividus sea-urchin. Ecotoxicology 2001, 10, 263–271. [Google Scholar]





{kind=link}
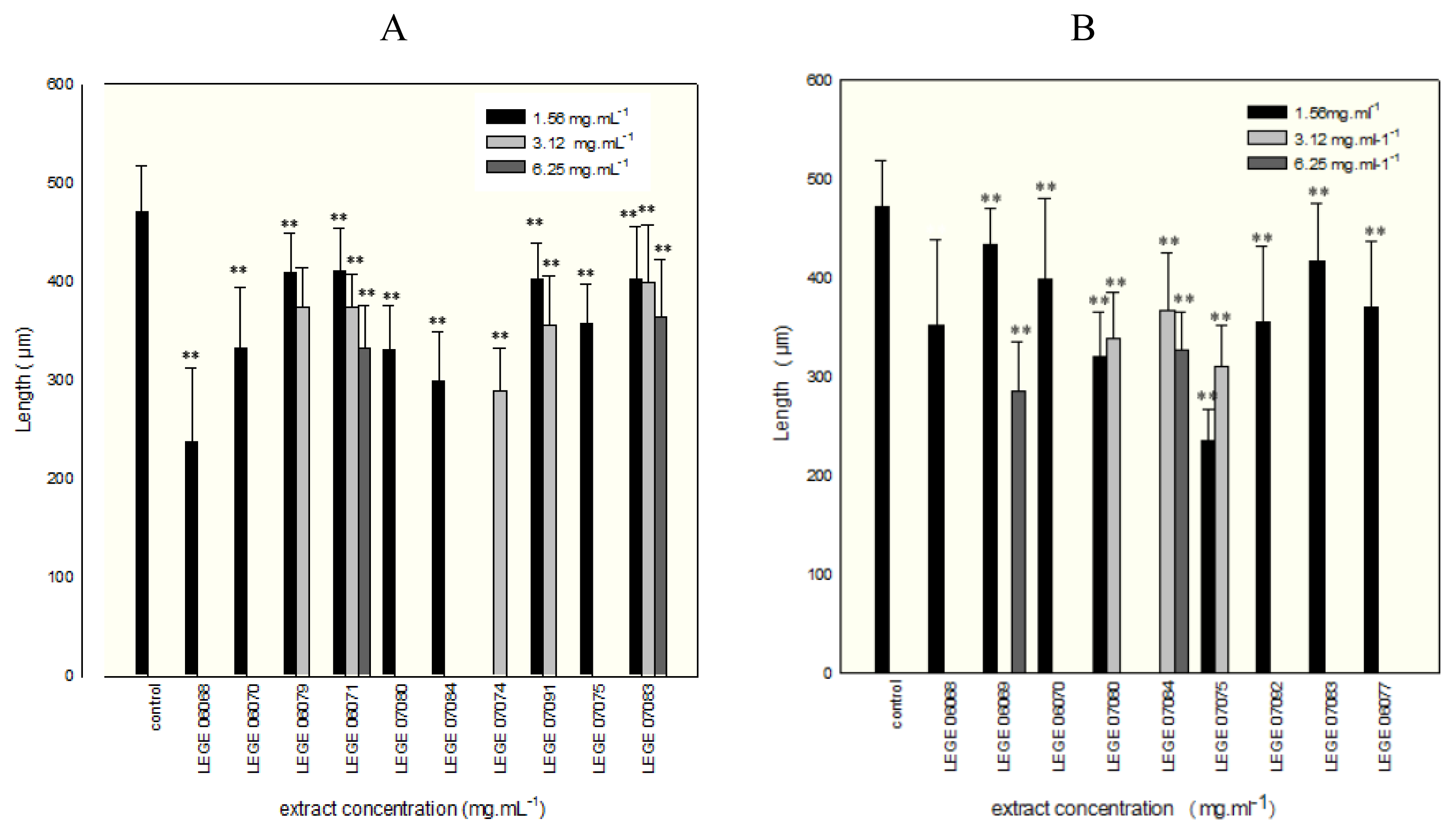{kind=link}
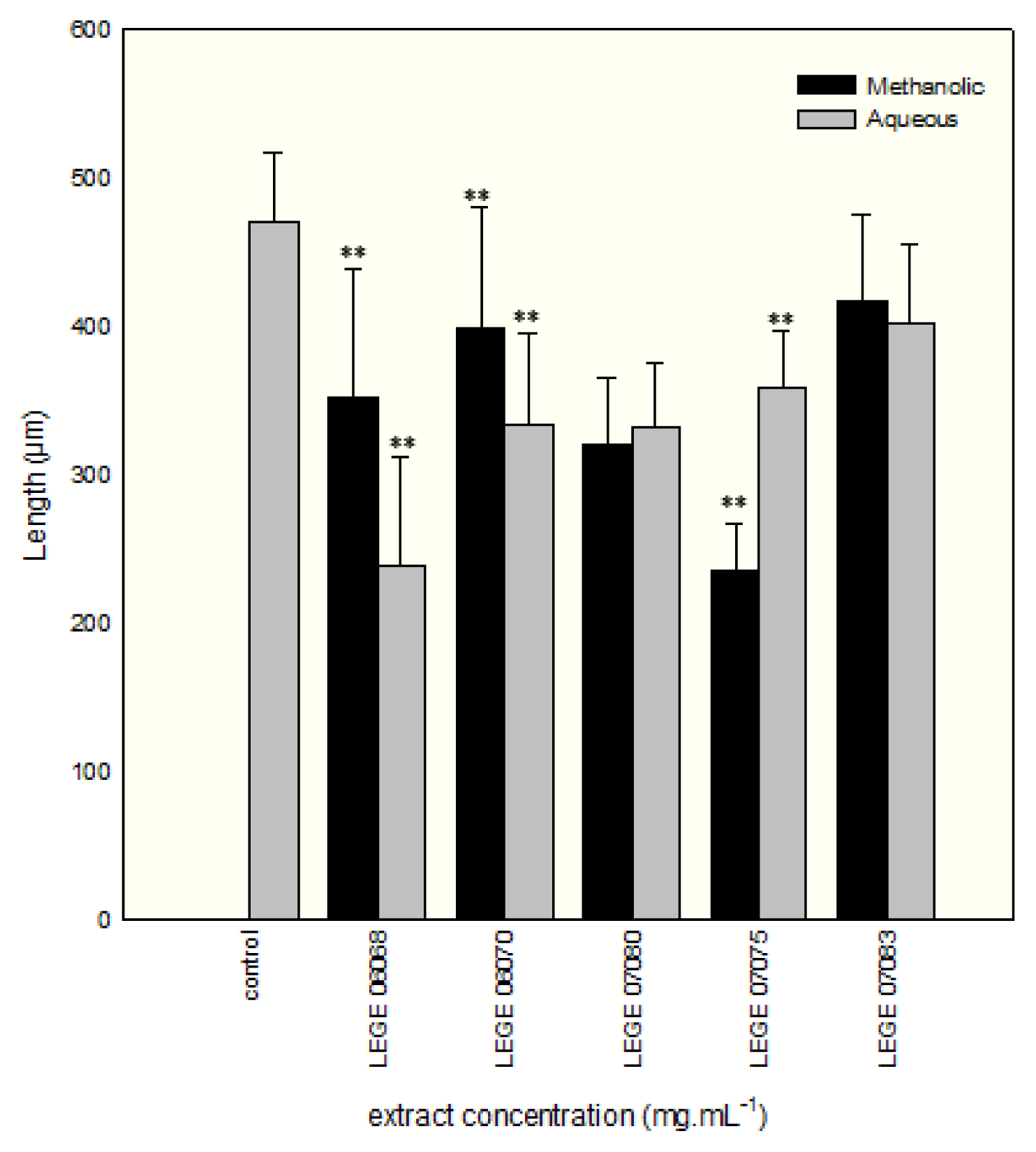{kind=link}
{kind=link}
| Identification | Coordinates | ||
|---|---|---|---|
| Lab code | Taxon | Latitude | Longitude |
| LEGE 06068 | Cyanobium sp. | 41° 8′50.77″N | 8°39′12.89″W |
| LEGE 06069 | Leptolyngbya sp. | 41° 8′50.45″N | 8°38′2.13″W |
| LEGE 06070 | Leptolyngbya sp. | 41° 8′50.45″N | 8°38′2.13″W |
| LEGE 06071 | Nodularia sp. | 40°38′32.87″N | 8°39′47.85″W |
| LEGE 06072 | Phormidium sp. | 40°40′16.42″N | 8°43′24.36″W |
| LEGE 06078 | Phormidium sp. | 41° 8′12.20″N | 8°39′54.65″W |
| LEGE 06079 | Synechocystis sp. | 41° 8′12.20″N | 8°39′54.65″W |
| LEGE 07073 | Synechocystis sp. | 40°40′16.42″N | 8°43′24.36″W |
| LEGE 07074 | Leptolyngbya sp. | 41° 8′48.17″N | 8°39′38.79″W |
| LEGE 07075 | Leptolyngbya sp. | 41° 8′50.45″N | 8°38′2.13″W |
| LEGE 07076 | Microcoleus sp. | 41°54′5.00″N | 8°48′51.88″W |
| LEGE 07077 | Nostoc sp. | 41°52′40.13″N | 8°50′6.33″W |
| LEGE 07080 | Leptolyngbya sp. | 41°52′2.50″N | 8°51′35.90″W |
| LEGE 07083 | Synechocystis sp. | 41° 8′48.17″N | 8°39′38.79″W |
| LEGE 07084 | Leptolyngbya sp. | 41°52′16.76″N | 8°50′39.66″W |
| LEGE 07085 | Leptolyngbya sp. | 41° 8′50.77″N | 8°39′12.89″W |
| LEGE 07091 | Leptolyngbya sp. | 40°40′16.42″N | 8°43′24.36″W |
| LEGE 07092 | Microcoleus sp. | 40°40′16.42″N | 8°43′24.36″W |
| LC50 (95% confidence limits ) mg mL−1 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Methanolic extracts | Aqueous extracts | |||||||
| Code | 24 h | 48 h | 24 h | 48 h | ||||
| LEGE 06068 | - | - | - | - | - | - | 79.92 | (36.28–653.08) |
| LEGE 06069 | - | - | - | - | - | - | 32.62 | (27.22–39.10) |
| LEGE 06070 | - | - | - | - | - | - | 15.42 | (5.00–27.56) |
| LEGE 06071 | - | - | 17.81 | (11.63–27.30) | - | - | 53.20 | (37.97–65.94) |
| LEGE 06072 | - | - | - | - | - | - | 20.13 | (12.18–30.36) |
| LEGE 06077 | - | - | - | - | 72.11 | (54.96–94.60) | 72.01 | (57.88–86.61) |
| LEGE 06078 | - | - | - | - | - | - | 28.60 | (8.93–40.58) |
| LEGE 06079 | - | - | - | - | 22.63 | NR | - | - |
| LEGE 07073 | - | - | - | - | - | - | - | - |
| LEGE 07074 | - | - | - | - | - | - | 13.78 | (11.46–16.55) |
| LEGE 07075 | - | - | - | - | - | - | 8.95 | (4.98–15.00) |
| LEGE 07076 | - | - | - | - | - | - | 12.10 | (9.95–14.64) |
| LEGE 07080 | - | - | - | - | - | - | - | - |
| LEGE 07083 | - | - | - | - | - | - | - | - |
| LEGE 07084 | - | - | - | - | 32.62 | (28.00–38.00) | 32.62 | (28.00–38.00) |
| LEGE 07085 | - | - | - | - | 94.9 | (73.14–133.15) | 64.61 | (52.23–79.92) |
| LEGE 07091 | - | - | - | - | 26.15 | (19.85–34.4) | 26.15 | (19.85–34.4) |
| LEGE 07092 | - | - | - | - | 20.68 | (15.17–28.18) | 18.08 | (14.15–23.10) |
| Aqueous | Methanolic | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Code | 25.00 | 12.50 | 6.25 | 3.12 | 1.56 | 25.00 | 12.50 | 6.25 | 3.12 | 1.56 |
| LEGE 06068 | G | G | G | P | P | M | M | M/G | M/P | G/P |
| LEGE 06069 | M | M | M | M | M | M | M | P/L | G | M/P/L |
| LEGE 06070 | B | B | B | G/P | G/P/L | M | M | M | M/G | M/G/L |
| LEGE 06078 | B | B | B | B | G/P | G | G/P | G/P | G/P | G/P |
| LEGE 06079 | M | G | P | L | L | M | M | M | M | M/L |
| LEGE 06071 | G | P | L | L | L | B | B | B | B | B |
| LEGE 06072 | G | G | G/P | G/P | P | M | M/B | G | G | P |
| LEGE 07080 | M | M | M | M/G/L | M/G/L | B | G | G/P | L | L |
| LEGE 07084 | B | B | B | B | G/P/L | G | G | P/L | L | L |
| LEGE 07074 | B | B | B | G | G/P/L | M | M | M | P/L | M/G/L |
| LEGE 07085 | B | B | B | B | G | B | B | G | P | P |
| LEGE 07075 | G | G | G | P | L | M | B | P | G | P/L |
| LEGE 07091 | M | M | M | M | M | M | M | G | P/L | L |
| LEGE 07076 | B | B | B/G | B/G | G | M | M | M | G | M/G/L |
| LEGE 07092 | G | G | G | G | G | M | M | M/G | G/P | G/L |
| LEGE 06077 | M | M | M | M | G/P | M | M | M/P | P | P/L |
| LEGE 07073 | B | B | B | G | G | M | M | M | P | M/P/L |
| LEGE 07083 | M | G/P | P/L | L | L | M | B | B | B/G/P | L |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lopes, V.R.; Fernández, N.; Martins, R.F.; Vasconcelos, V. Primary Screening of the Bioactivity of Brackishwater Cyanobacteria: Toxicity of Crude Extracts to Artemia salina Larvae and Paracentrotus lividus Embryos. Mar. Drugs 2010, 8, 471-482. https://doi.org/10.3390/md8030471
Lopes VR, Fernández N, Martins RF, Vasconcelos V. Primary Screening of the Bioactivity of Brackishwater Cyanobacteria: Toxicity of Crude Extracts to Artemia salina Larvae and Paracentrotus lividus Embryos. Marine Drugs. 2010; 8(3):471-482. https://doi.org/10.3390/md8030471
Chicago/Turabian StyleLopes, Viviana R., Nuria Fernández, Rosário F. Martins, and Vitor Vasconcelos. 2010. "Primary Screening of the Bioactivity of Brackishwater Cyanobacteria: Toxicity of Crude Extracts to Artemia salina Larvae and Paracentrotus lividus Embryos" Marine Drugs 8, no. 3: 471-482. https://doi.org/10.3390/md8030471
APA StyleLopes, V. R., Fernández, N., Martins, R. F., & Vasconcelos, V. (2010). Primary Screening of the Bioactivity of Brackishwater Cyanobacteria: Toxicity of Crude Extracts to Artemia salina Larvae and Paracentrotus lividus Embryos. Marine Drugs, 8(3), 471-482. https://doi.org/10.3390/md8030471