Novel Anti-Infective Compounds from Marine Bacteria


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Anti-methicillin-resistant Staphylococcus aureus (MRSA) and Anti-vancomycin-resistant Enterococci (VRE) Compounds Derived from Marine Bacteria
3. Conclusions
References
- Michael, A; Fischbach, MA; Walsh, CT. Antibiotics for emerging pathogens. Science 2009, 325, 1089–1093. [Google Scholar]
- Nathan, C. Antibiotics at the crossroads. Nature 2004, 431, 899–902. [Google Scholar]
- Nussbaum, FV; Brands, M; Hinzen, B; Weigand, S; Habich, D. Antibacterial natural products in medicinal chemistry – exodus or revival. Angew Chem Int Ed 2006, 45, 5072–5129. [Google Scholar]
- Newman, DJ; Cragg, GM. Natural products as sources of new drugs over the last 25 years. J Nat Prod 2007, 70, 461–477. [Google Scholar]
- Rahman, H. Unusual Sesquiterpenes: Gorgonenes and Further Bioactive Secondary Metabolites Derived from Marine and Terrestrial Bacteria. In PhD Thesis; Universität Göttingen: Germany, 2008; p. 158. [Google Scholar]
- Laatsch, H. AntiBase, A Data Base for Rapid Structural Determination of Microbial Natural Products, and annual updates; Wiley VCH: Weinheim, Germany, 2008. [Google Scholar]
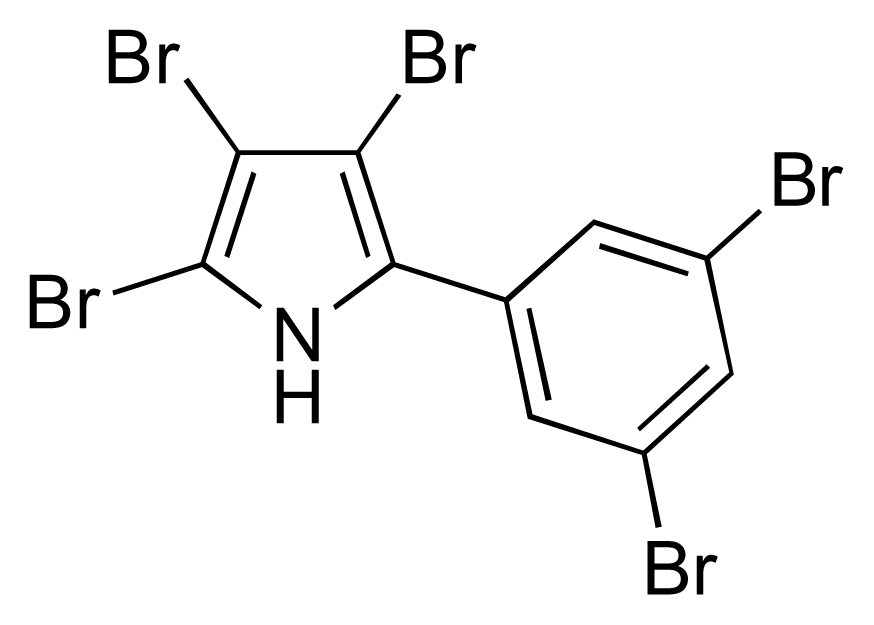
- Burkholder, PR; Pfister, RM; Leitz, FH. Production of a pyrrole antibiotic by a marine bacterium. Appl Microbiol 1966, 14, 649–653. [Google Scholar]
- Riedlinger, J; Reicke, A; Zähner, H; Krismer, B; Bull, AT; Maldonado, LA; Ward, AC; Goodfellow, M; Bister, B; Bischoff, D; Süssmuth, RD; Fiedler, HP. Abyssomicins, inhibitors of the para-aminobenzoic acid pathway produced by the marine Verrucosispora strain AB-18-032. J Antibiot 2004, 57, 271–279. [Google Scholar]
- Donia, M; Hamann, MT. Marine natural products and their potential applications as anti-infective agents. Lancet Infect Dis 2003, 3, 338–348. [Google Scholar]
- Newman, DJ; Cragg, GM; Snader, KM. The influence of natural products upon drug discovery. Nat Prod Rep 2000, 17, 215–234. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat Prod Rep 2002, 19, 1–48. [Google Scholar]
- El Sayed, KA; Bartyzel, P; Shen, X; Perry, TL; Zjawiony, JK; Hamann, MT. Marine natural products as antituberculosis agents. Tetrahedron 2000, 56, 949–953. [Google Scholar]
- El Sayed, KA. Natural products as antiviral agents. Stud Nat Prod Chem 2000, 24, 473–572. [Google Scholar]
- Kayser, O; Kiderlen, AF; Croft, SL. Natural product as potential antiparasitic drugs. Stud Nat Prod Chem 2002, 26, 779–848. [Google Scholar]
- Ghisalberti, EL. Secondary metabolites with antinematodal activity. Stud Nat Prod Chem 2002, 26, 425–506. [Google Scholar]
- Li, H-Y; Matsunaga, S; Fusetani, N. Antifungal metabolites from marine sponges. Curr Org Chem 1998, 2, 649–682. [Google Scholar]
- Klevens, RM; Morrison, MA; Nadle, J; Petit, S; Gershman, K; Ray, S; Harrison, LH; Lynfield, R; Dumyati, G; Townes, JM; Craig, AS; Zell, ER; Fosheim, GE; McDougal, LK; Carey, RB; Fridkin, SK. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar]
- Huang, H; Flynn, NM; King, JH; Monchaud, C; Morita, M; Cohen, SH. Comparisons of community-associated methicillin-resistant Staphylococcus aureus (MRSA) and hospital-associated MRSA infections in Sacramento, California. J Clin Microbiol 2006, 44, 2423–2427. [Google Scholar]
- Hiramatsu, K; Okuma, K; Hori, S; Kapi, M. New trends in Staphylococcus aureus infections: glycopeptide resistance in hospital and methicillin resistance in the community. Curr Opin Infect Dis 2002, 15, 407–413. [Google Scholar]
- Witte, W. Antibiotic resistance in Gram-positive bacteria: epidemiological aspects. J Antimicrob Chemother 1999, 44, 1–9. [Google Scholar]
- Madani, TA. Epidemiology and clinical features of methicillin-resistant Staphylococcus aureus in the university hospital, Jeddah, Saudi Arabia. Can J Infect Dis 2002, 13, 245–250. [Google Scholar]
- Hanaki, H; Labischinski, H; Inaba, Y; Kondo, N; Murakami, H; Hiramatsu, K. Increase in glutamine-non-amidated muropeptides in the peptidoglycan of vancomycin-resistant Staphylococcus aureus strain Mu50. J Antimicrob Chemother 1998, 42, 315–320. [Google Scholar]
- Patersen, DL. Reduced susceptibility of Staphylococcus aureus to vancomycin––a review of current knowledge. Commun Dis Intell 1999, 24, 69–73. [Google Scholar]
- Mainardi, JL; Shlaes, DM; Goering, RV; Shlaes, JH; Acar, JF; Goldstein, FW. Decreased teicoplanin susceptibility of methicillin-resistant strains of Staphylococcus aureus. J Infect Dis 1995, 171, 1646–1650. [Google Scholar]
- Vaudaux, P; Francois, P; Berger-Bachi, B; Lew, DP. In vivo emergence of subpopulations expressing teicoplanin or vancomycin resistance phenotypes in a glycopeptide-susceptible, methicillin-resistant strain of Staphylococcus aureus. J Antimicrob Chemother 2001, 47, 163–170. [Google Scholar]
- Leclerq, R; Derlot, E; Duval, J; Courvalin, P. Plasmid-mediated resistance to vancomycin and teicoplanin in Enterococcus faecium. N Engl J Med 1988, 319, 157–161. [Google Scholar]
- Weigel, LM; Clewell, DB; Gill, SR; Clark, NC; McDougal, IM; Flannagan, SE; Kolonay, JF; Shetty, J; Killgore, GE; Tenover, FC. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar]
- Edmond, MB; Ober, JF; Weinbaum, DL; Pfaller, MA; Hwang, T; Sanford, MD; Wenzel, RP. Vancomycin-resistant Enterococcus faecium bacteremia: risk factors for infection. Clin Infect Dis 1995, 20 , 1126–1133. [Google Scholar]
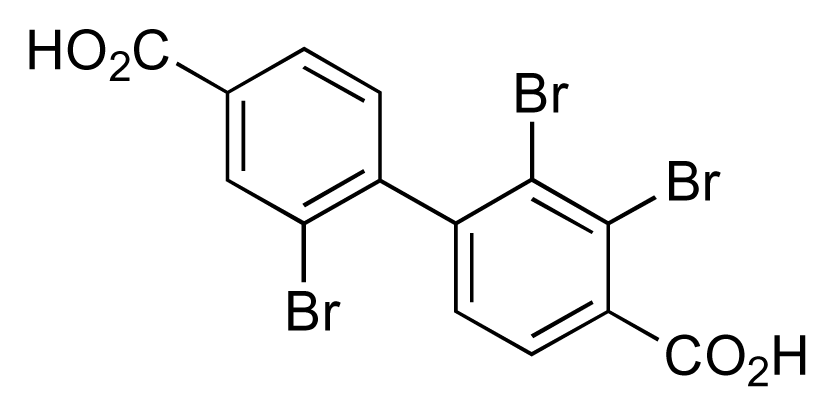
- Hughes, CC; Prieto-Davo, A; Jensen, PR; Fenical, W. The marinopyrroles, antibiotics of an unprecedented structure class from a marine Streptomyces sp. Org Lett 2008, 10, 629–631. [Google Scholar]
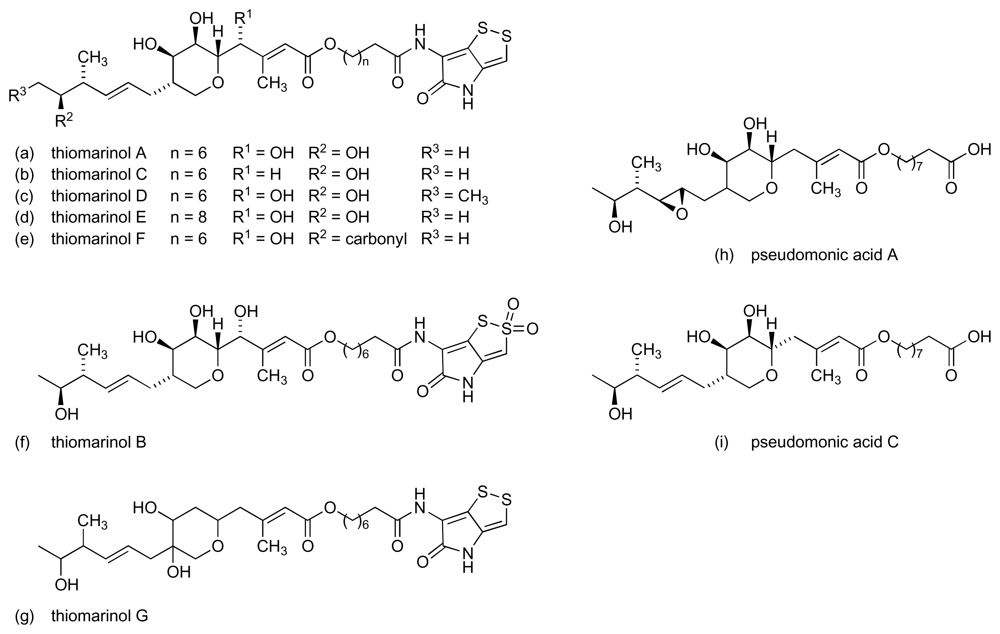
- Shiozawa, H; Shimada, A; Takahashi, S. Thiomarinols D, E, F and G, new hybrid antimicrobial antibiotics produced by a marine bacterium; isolation, structure, and antimicrobial activity. J Antibiot 1997, 50, 449–452. [Google Scholar]
- Shiozawa, H; Kagasaki, T; Kinoshita, T; Haruyama, H; Domon, H; Utsui, Y; Kodama, K; Takahashi, S. Thiomarinol, a hybrid antimicrobial antibiotic produced by a marine bacterium fermentation, isolation, structure and antimicrobial activity. J Antibiot 1993, 46, 1834–1842. [Google Scholar]
- Shiozawa, H; Takahashi, S. Configurational studied on thiomarinol. J Antibiot 1994, 47, 851–853. [Google Scholar]
- Shiozawa, H; Kagasaki, T; Torikata, A; Tanaka, N; Fujimoto, K; Hata, T; Furukawa, Y; Takahashi, S. Thiomarinol B, thiomarinol C, new antimicrobial antibiotics produced by a marine bacterium. J Antibiot 1995, 48, 907–909. [Google Scholar]
- Fuller, AT; Mellows, G; Woolford, M; Banks, GT; Barrow, KD; Chain, EB. Pseudomonic acid – antibiotic produced by Pseudomonas fluorescens. Nature 1971, 234, 416. [Google Scholar]
- Hughes, J; Mellows, G. Interaction of pseudomonic acid A with Escherichia coli B-isoleucyl transfer RNA synthetase. Biochem J 1980, 191, 209–219. [Google Scholar]
- Isnansetyo, A; Kamei, Y. Anti-methicillin-resistant Staphylococcus aureus (MRSA) activity of MC21-B, an antibacterial compound produced by the marine bacterium Pseudoalteromonas phenolica O-BC30T. Int J Antimicrob Agents 2009, 34, 131–135. [Google Scholar]
- Harada, H; Kamei, Y. Selective cytotoxicity of marine algae extract to several human leukemic cell lines. Cytotechnology 1997, 25, 213–219. [Google Scholar]
- Isnansetyo, A; Kamei, Y. MC21-A, a bactericidal antibiotic against methicillin-resistant Staphylococcus aureus. Antimicrob Ag Chemother 2003, 47, 480–488. [Google Scholar]
- Yoongho, L; Jun-Hwan, C; Jong-Hoon, K; Jung-Woo, S; Jae-Kyung, J; Chul-Hoon, L. Structure elucidation of a potent anti-MRSA antibiotic AM3, produced by Streptomyces sp. Han’guk Nonghwa Hakhoechi 1995, 38, 516–521. [Google Scholar]
- Keller, S; Nicholson, G; Drahl, C; Sorensen, E; Fiedler, HP; Süssmuth, RD. Abyssomicins G and H and atrop-abyssomicin C from the marine Verrucosispora strain AB-18-032. J Antibiot 2007, 60, 391–394. [Google Scholar]
- Bister, B; Bischoff, D; Strobele, M; Riedlinger, J; Reicke, A; Wolter, F; Bull, AT; Zahner, H; Fiedler, HP; Süssmuth, RD. Abyssomicin C–a polycyclic antibiotic from a marine Verrucosispora strain as an inhibitor of the p-aminobenzoic acid/tetrahydrofolate biosynthesis pathway. Angew Chem Int Edit 2004, 43, 2574–2576. [Google Scholar]
- Nicolaou, KC; Harrison, ST. Total synthesis of abyssomicin C and atrop-abyssomicin C. Angew Chem Int Ed 2006, 45, 3256–3260. [Google Scholar]
- Zapf, CW; Harrison, BA; Drahl, C; Sorensen, EJ. A Diels-Alder macrocyclization enables an efficient asymmetric synthesis of the antibacterial natural product abyssomicin C. Angew Chem Int Ed 2005, 44, 6533–6537. [Google Scholar]
- Peters, R; Fischer, DF. Total syntheses of the antibacterial natural product abyssomicin C. Angew Chem Int Ed 2006, 45, 5736–5739. [Google Scholar]
- Niu, XM; Li, SH; Gorls, H; Schollmeyer, D; Hilliger, M; Grabley, S; Sattler, I. Abyssomicin E, a highly functionalized polycyclic metabolite from Streptomyces species. Org Lett 2007, 9, 2437–2440. [Google Scholar]
- Nicolaou, KC; Harrison, ST. Total synthesis of abyssomicin C, atrop-abyssomicin C, and abyssomicin D: implications for natural origins of atrop-abyssomicin C. J Am Chem Soc 2007, 129, 429–440. [Google Scholar]
- Keller, S; Schadt, HS; Ortel, I; Süssmuth, RD. Action of atrop-abyssomicin C as an inhibitor of 4-amino-4-deoxychorismate synthase PabB. Angew Chem Int Ed 2007, 46, 8284–8286. [Google Scholar]
- Bartlett, PA; Johnson, CR. An inhibitor of chorismate mutase resembling the transition state conformation. J Am Chem Soc 1985, 107, 7792–7793. [Google Scholar]
- Furumai, T; Eto, K; Sasaki, T; Higuchi, H; Onaka, H; Saito, N; Fujita, T; Naoki, H; Igarashi, Y. TPU-0037-A, B, C and D, novel lydicamycin congeners with anti-MRSA activity from Streptomyces platensis TP-A0598. J Antibiot 2002, 55, 873–880. [Google Scholar]
- Hayakawa, Y; Kanamaru, N; Shimazu, A; Seto, H. Lydicamycin, a new antibiotic of a novel skeletal type. I. Taxonomy, fermentation, isolation and biological activity. J Antibiot 1991, 44, 282–287. [Google Scholar]
- Hayakawa, Y; Kanamaru, N; Morisaki, N; Furihata, K; Seto, H. Lydicamycin, a new antibiotic of a novel skeletal type. II. Physico-chemical properties and structure elucidation. J Antibiot 1991, 44, 288–292. [Google Scholar]
- Kushida, H; Nakajima, S; Koyama, T; Suzuki, H; Ojiri, K; Suda, H. Antitumoric BE-43472 manufacture with streptomyces. 1996. JP 08143569. [Google Scholar]
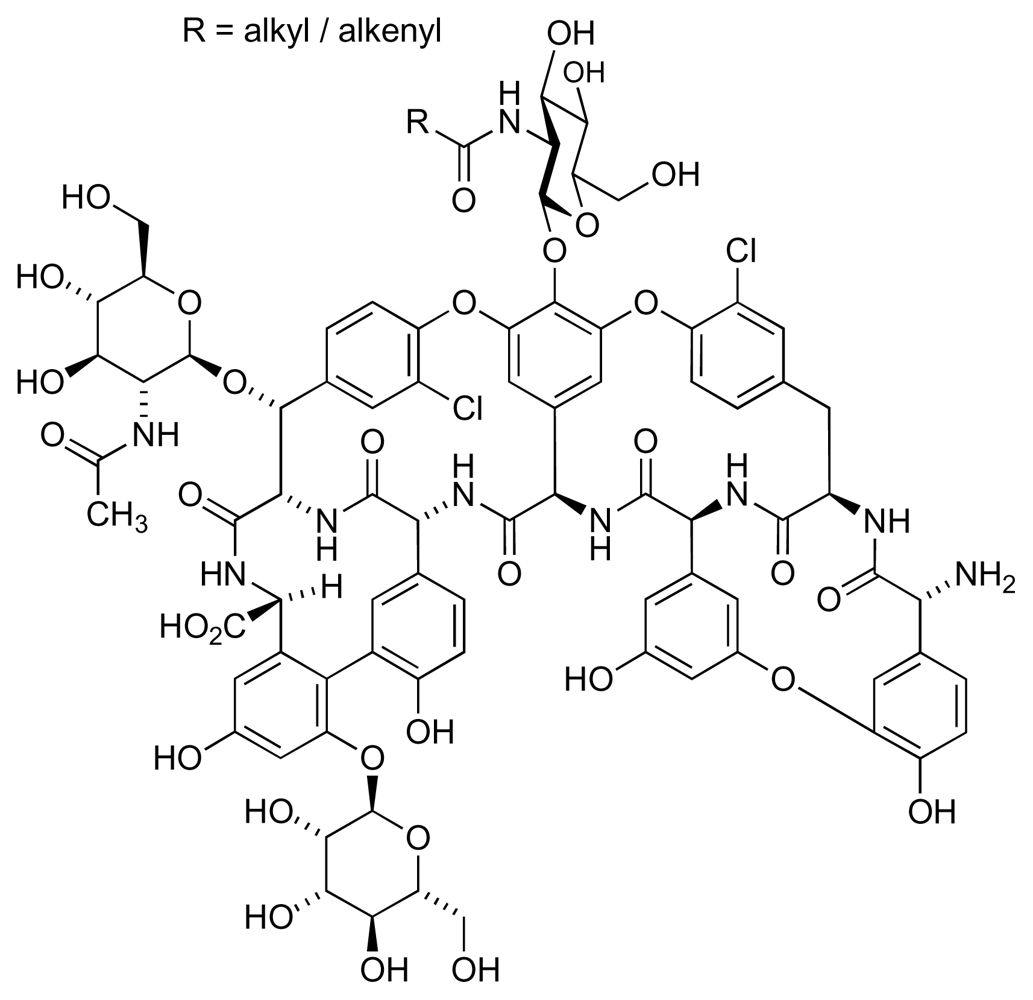
- Socha, AM; Garcia, D; Sheffer, R; Rowley, DC. Antibiotic bisanthraquinones produced by a streptomycete isolated from a cyanobacterium associated with Ecteinascidia turbinata. J Nat Prod 2006, 69, 1070–1073. [Google Scholar]
- Socha, AM; LaPlante, KL; Rowley, DC. New bisanthraquinone antibiotics and semi-synthetic derivatives with potent activity against clinical Staphylococcus aureus and Enterococcus faecium isolates. Bioorg Med Chem 2006, 14, 8446–8454. [Google Scholar]
- Nicolaou, KC; Lim, YH; Becker, J. Total synthesis and absolute configuration of the bisanthraquinone antibiotic BE-43472B. Angew Chem Int Ed 2009, 48, 3444–3448. [Google Scholar]
- Nicolaou, KC; Becker, J; Lim, YH; Lemire, A; Neubauer, T; Montero, A. Total synthesis and biological evaluation of (+)- and (−)-bisanthraquinone antibiotic BE-43472B and related compounds. J Am Chem Soc 2009, 131, 14812–14826. [Google Scholar]
- Michelle, S; Teisan, S; Tsueng, G; Venkat, M; Kin, L. Seawater requirement for the production of lipoxazolidinones by marine actinomycete strain NPS8920. J Ind Microb Biotech 2008, 35, 761–765. [Google Scholar]
- Macherla, VR; Liu, JN; Sunga, M; White, DJ; Grodberg, J; Teisan, S; Lam, KS; Potts, BCM. Lipoxazolidinones A, B, and C: antibacterial 4-oxazolidinones from a marine actinomycete isolates from a Guam marine sediment. J Nat Prod 2007, 70, 1454–1457. [Google Scholar]
- Maya, SP; Kong, F; Jeffrey, JE; Daniel, AA; Paola, SA; Valerie, BS; Peter, PJ; Weiss, WJ; Carter, G; Greenstein, M. Novel alpha-pyrones produced by a marine Pseudomonas sp. F92S91: taxonomy and biological activities. J Antibiot 2003, 56, 1033–1044. [Google Scholar]
- Kamei, Y; Isnansetyo, A. Lysis of methicillin-resistant Staphylococcus aureus by 2,4-diacetylphloroglucinol produced by Pseudomonas sp. AMSN isolated from a marine alga. Int J Antimicrob Agents 2003, 21, 71–74. [Google Scholar]
- Isnansetyo, A; Longzhu, C; Keiichi, H; Kamei, Y. Antibacterial activity of 2,4-diacetylphloroglucinol produced by Pseudomonas sp. AMSN isolated from a marine algae, against vancomycin-resistant Staphylococcus aureus. Int J Antimicrob Agents 2003, 22, 545–547. [Google Scholar]
- Isnansetyo, A; Horikawa, M; Kamei, Y. In vitro anti-methicillin resistant Staphylococcus aureus activity of 2,4-diacetylphloroglucinol produced by a Pseudomonas sp AMSN isolated from a marine alga. J Antimicrob Chemother 2001, 47, 724–725. [Google Scholar]
- Wang, J; Soisson, SM; Young, K; Shoop, W; Kodali, S; Galgoci, A; Painter, R; Parthasarathy, G; Tang, YS; Cummings, R; Ha, S; Dorso, K; Motyl, M; Jayasuriya, H; Ondeyka, J; Herath, K; Zhang, C; Hernandez, L; Allocco, J; Basilio, Á; Tormo, JR; Genilloud, O; Vicente, F; Pelaez, F; Colwell, L; Lee, SH; Michael, B; Felcetto, T; Gill, C; Silver, LL; Hermes, JD; Bartizal, K; Barrett, J; Schmatz, D; Becker, JW; Cully, D; Singh, SB. Platensimycin is a selective FabF inhibitor with potent antibiotic properties. Nature 2006, 441, 358–361. [Google Scholar]
- Abdelfattah, MS. New Secondary Metabolites from Bacteria: Seitomycin with high Anti- Helicobacter pylori Activity, Exfoliazone B, new Steffimycinones, Espicufolin B, Flavomarine A and B, and BS-46 with a Novel Carbon Skeleton. In PhD Thesis; Universität Göttingen, 2005. [Google Scholar]
- Brinster, S; Lamberet, G; Staels, B; Trieu-Cuot, P; Gruss, A; Poyart, C. Type II fatty acid synthesis is not a suitable antibiotic target for Gram-positive pathogens. Nature 2009, 458, 83–85. [Google Scholar]
- Barsby, T; Kelly, MT; Gagne, SM; Andersen, RJ. Bogorol A, produced in culture by a marine Bacillus sp. reveals a novel template for cationic peptide antibiotics. Org Lett 2001, 3, 437–440. [Google Scholar]
- Gerard, JM; Haden, P; Kelly, MT; Andersen, RJ. Loloatins A-D, cyclic decapeptide antibiotics produced in culture by a tropical marine bacterium. J Nat Prod 1999, 62, 80–85. [Google Scholar]
- Gerard, J; Haden, P; Kelly, MT; Andersen, RJ. Loloatin B, a cyclic decapeptide antibiotic produced in culture by a tropical marine bacterium. Tetrahedron Lett 1996, 37, 7201–7204. [Google Scholar]
- Krachkovskii, SA; Sobol, AG; Ovchinnikova, TV; Tagaev, AA; Yakimenko, ZA; Azizbekyan, RR; Kuznetsova, NI; Shamshina, TN; Arseniev, AS. The isolation, biological properties, and spatial structure of antibiotic loloatin A. Russ J Bioorg Chem 2002, 28, 269–273. [Google Scholar]
- Chen, H; Haynes, RK; Scherkenbeck, J. Synthesis of cyclic hexapeptides based on the antibiotic cyclice decapeptide loloatin C by an in situ indirect cyclization method. Eur J Org Chem 2004, 1, 38–47. [Google Scholar]
- Chen, H; Haynes, RK; Scherkenbeck, J; Sze, KH; Zhu, G. Elucidation of the solution conformations of loloatin C by NMR spectroscopy and molecular simulation. Eur J Org Chem 2004, 31–37. [Google Scholar]
- Chen, HR; Guo, XK. Study of the conformation of cyclodecapeptide loloatin C with obvious antibiotic activity. Chinese J Struct Chem 2005, 24, 273–278. [Google Scholar]
- Chen, HR; Guo, XK; Zhong, XB. Synthesis of aladan and [Ald(6)]loloactin C and study of their fluorescent properties. Chin J Chem 2006, 24, 1411–1417. [Google Scholar]
- Tuin, AW; Grotenbreg, GM; Spalburg, E; de Neeling, AJ; Mars-Groenendijk, RH; van der Marel, GA; Noort, D; Overkleeft, HS; Overhand, M. Structural and biological evaluation of some loloatin C analogues. Bioorg Med Chem 2009, 17, 6233–6240. [Google Scholar]
- Kwon, HC; Kauffman, CA; Jensen, PR; Fenical, W. Marinomycins A-D, antitumor-antibiotics of a new structure class from a marine actinomycete of the recently discovered genus “Marinispora”. J Am Chem Soc 2006, 128, 1622–1632. [Google Scholar]
- Needham, J; Kelly, MT; Ishige, M; Andersen, RJ. Andrimid and moiramides A-C, metabolites produced in culture by a marine isolate of the bacterium Pseudomonas fluorescens: structure elucidation and biosynthesis. J Org Chem 1994, 59, 2058–2063. [Google Scholar]
- Fredenhagen, A; Tamura, SY; Kenny, PTM; Komura, H; Naya, Y; Nakanishi, K; Nishiyama, K; Sugiura, M; Kita, H. Andrimid, a new peptide antibiotic produced by an intracellular bacterial symbiont isolated from a brown planthopper. J Am Chem Soc 1987, 109, 4409–4411. [Google Scholar]
- McWhorter, W; Fredenhagen, A; Nakanishi, K; Komura, H. Stereocontrolled synthesis of andrimid and a structural requirement for the activity. J Chem Soc, Chem Commun 1989, 5 , 299–301. [Google Scholar]
- Carlson, JC; Li, S; Burr, DA; Sherman, DH. Isolation and characterization of tirandamycins from a marine-derived Streptomyces sp. J Nat Prod 2009, 72, 2076–2079. [Google Scholar]
- Hagenmaier, H; Jaschke, KH; Santo, L; Scheer, M; Zahner, H. Metabolic products of microorganisms. 158. Tiransamycin B. Arch Microbiol 1976, 109, 65–74. [Google Scholar]
- Meyer, CE. Tirandamycin, a new antibiotic isolation and characterization. J Antibiot 1971, 24, 558–560. [Google Scholar]
- McArthur, KA; Mitchell, SS; Tsueng, G; Rheingold, A; White, DJ; Grodberg, J; Lam, KS; Potts, BCM. Lynamicins A–E, chlorinated bisindole pyrrole antibiotics from a novel marine actinomycete. J Nat Prod 2008, 71, 1732–1737. [Google Scholar]
- Omura, S; Iwai, Y; Hirano, A; Nakagawa, A; Awaya, J; Tsuchiya, H; Takahashi, Y; Masuma, R. New alkaloid AM-2282 of Streptomyces origin taxonomy, fermentation, isolation and preliminary characterization. J Antibiot 1977, 30, 275–282. [Google Scholar]
- Bush, JA; Long, BH; Catino, JJ; Bradner, WT. Production and biological activity of rebeccamycin, a novel antitumor agent. J Antibiot 1987, 40, 668–678. [Google Scholar]
- Hoshino, T; Kojima, Y; Hayashi, T; Uchiyama, T; Kaneko, K. Studies on the biosynthesis of violacein. 4. A new metabolite of tryptophan, chromopyrrolic acid, produced by Chromobacterium violaceum. Biosci Biotech Biochem 1993, 57, 775–781. [Google Scholar]
- Frode, R; Hinze, C; Josten, I; Schmidt, B; Steffan, B; Steglich, W. Isolation and synthesis of 3,4-bis(indol-3-yl)pyrroles-2,5-dicarboxylic acid derivatives from the slime mold Lycogala epidendrum. Tetrahedron Lett 1994, 35, 1689–1690. [Google Scholar]
- Hashimoto, T; Yasuda, A; Akazawa, K; Takaoka, S; Tori, M; Asakawa, Y. 3 novel dimethyl pyrroledicarboxylate, lycogarubins A-C, from the myxomycetes Lycogala epidendrum. Tetrahedron Lett 1994, 35, 2559–2560. [Google Scholar]
- Cueto, M; Jensen, PR; Kauffman, C; Fenical, W; Lobkovsky, E; Clardy, J. Pestalone, a new antibiotic produced by a marine fungus in response to bacterial challenge. J Nat Prod 2001, 64 , 1444–1446. [Google Scholar]
- Yan, LM; Boyd, KG; Adams, DR; Burgess, JG. Biofilm specific cross-species induction of antimicrobial compounds in bacilli. Appl Environ Microbiol 2003, 69, 3719–3727. [Google Scholar]
- Berdy, J. Bioactive microbial metabolites – a personal view. J Antibiot 2005, 58, 1–26. [Google Scholar]
- Boyd, KG; Adams, DR; Burgess, JG. Antibacterial and repellent activities of marine bacteria associated with algal surfaces. Biofouling 1999, 14, 227–236. [Google Scholar]
- Mearns-Spragg, A; Bregu, M; Boyd, KG; Burgess, JG. Cross-species induction and enhancement of antibiotic production by epiphytic bacteria from marine algae and invertebrates, after exposure to terrestrial bacteria. Lett Appl Microbiol 1998, 27, 142–146. [Google Scholar]
- Induction of microbial secondary metabolites. SeaRch® technology platform. Priority Application GB06008445





















© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rahman, H.; Austin, B.; Mitchell, W.J.; Morris, P.C.; Jamieson, D.J.; Adams, D.R.; Spragg, A.M.; Schweizer, M. Novel Anti-Infective Compounds from Marine Bacteria. Mar. Drugs 2010, 8, 498-518. https://doi.org/10.3390/md8030498
Rahman H, Austin B, Mitchell WJ, Morris PC, Jamieson DJ, Adams DR, Spragg AM, Schweizer M. Novel Anti-Infective Compounds from Marine Bacteria. Marine Drugs. 2010; 8(3):498-518. https://doi.org/10.3390/md8030498
Chicago/Turabian StyleRahman, Hafizur, Brian Austin, Wilfrid J. Mitchell, Peter C. Morris, Derek J. Jamieson, David R. Adams, Andrew Mearns Spragg, and Michael Schweizer. 2010. "Novel Anti-Infective Compounds from Marine Bacteria" Marine Drugs 8, no. 3: 498-518. https://doi.org/10.3390/md8030498
APA StyleRahman, H., Austin, B., Mitchell, W. J., Morris, P. C., Jamieson, D. J., Adams, D. R., Spragg, A. M., & Schweizer, M. (2010). Novel Anti-Infective Compounds from Marine Bacteria. Marine Drugs, 8(3), 498-518. https://doi.org/10.3390/md8030498