Brominated Compounds from Marine Sponges of the Genus Aplysina and a Compilation of Their 13C NMR Spectral Data

Abstract
:1. Introduction
The Genus Aplysina
2. Methodology
3. Discussion
3.1. Chemotaxonomy Importance of Aplysina Sponges
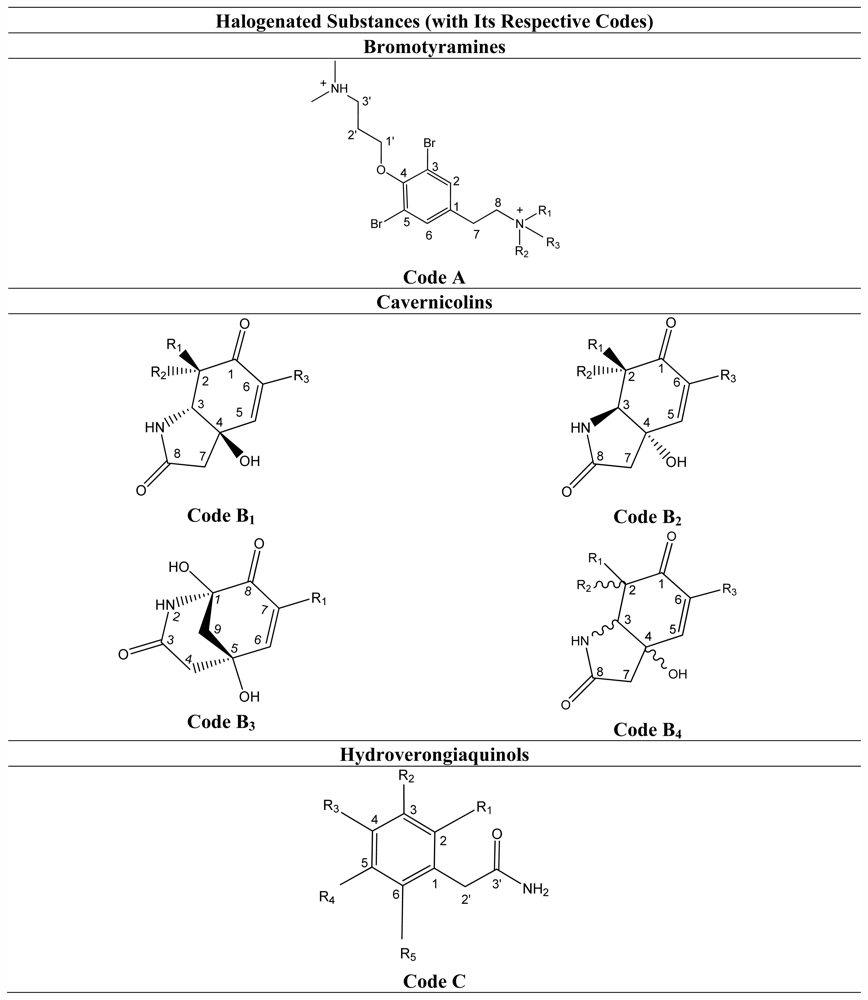
3.2. Bromotyramines
3.3. Cavernicolins
3.4. Hydroverongiaquinols
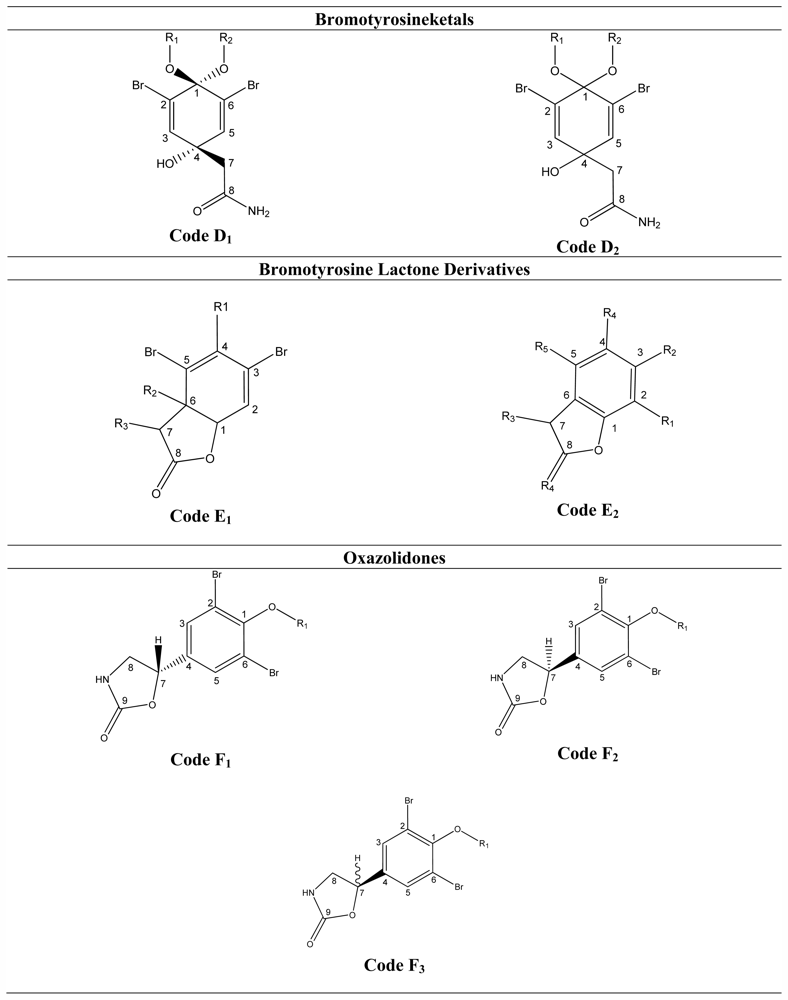
3.5. Bromotyrosineketals
3.6. Bromotyrosine Lactone Derivatives
3.7. Oxazolidones
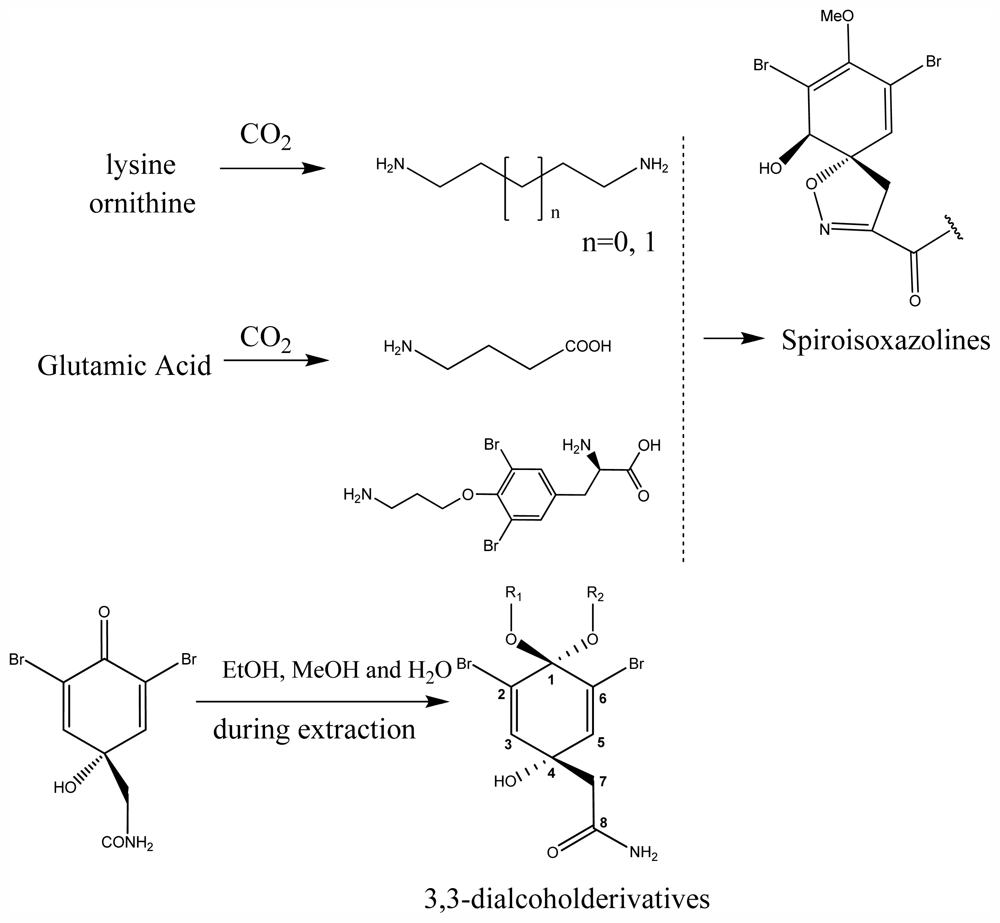
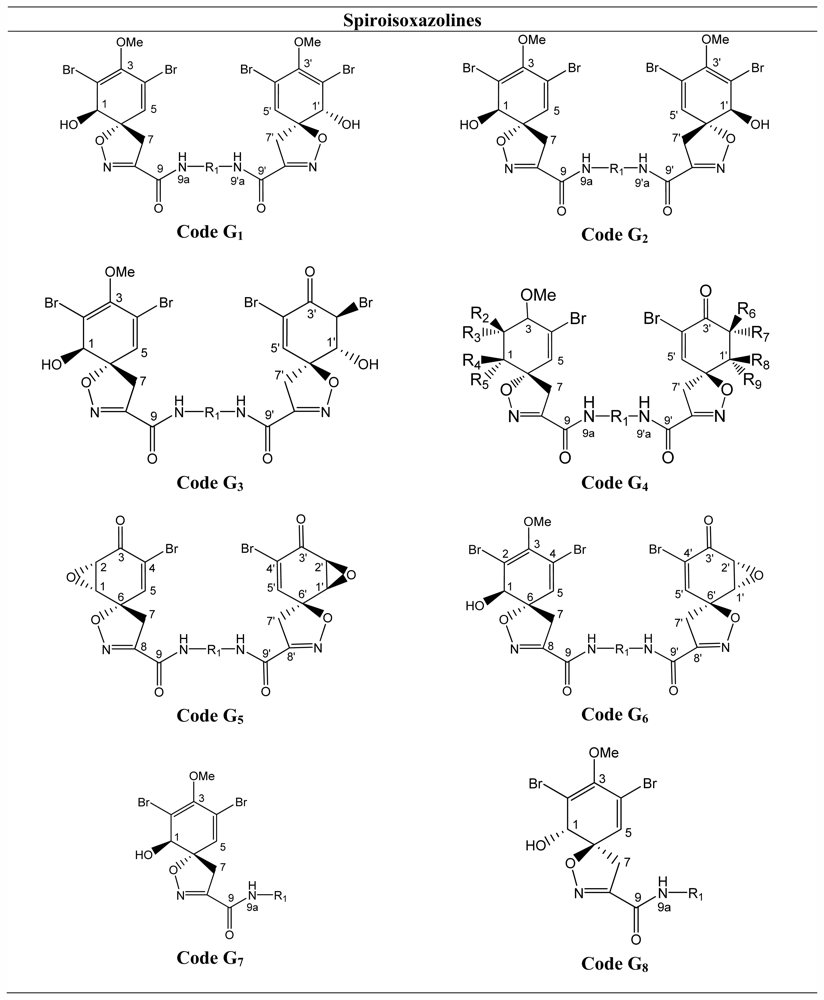
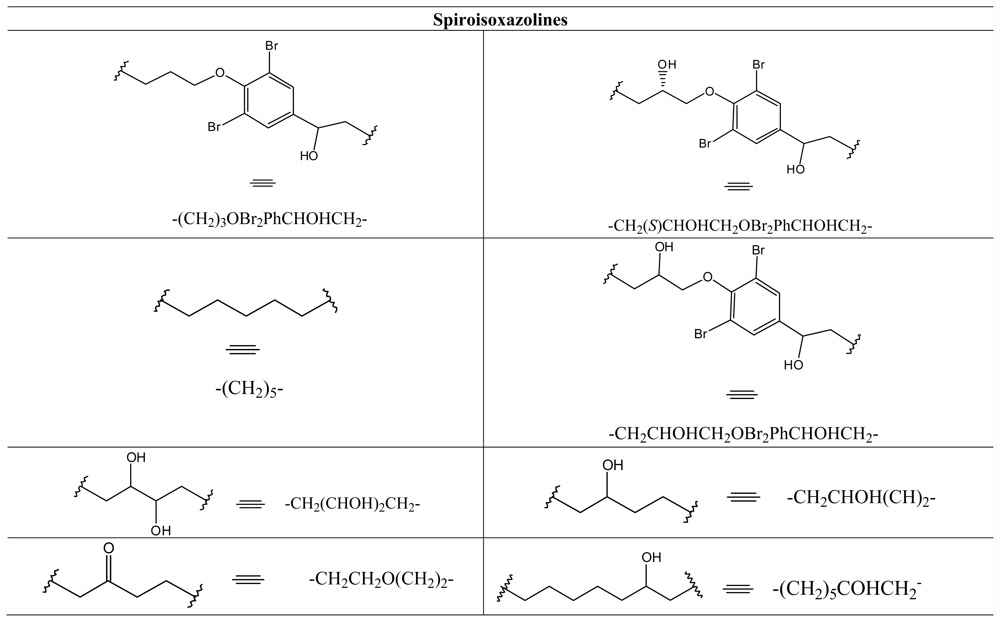
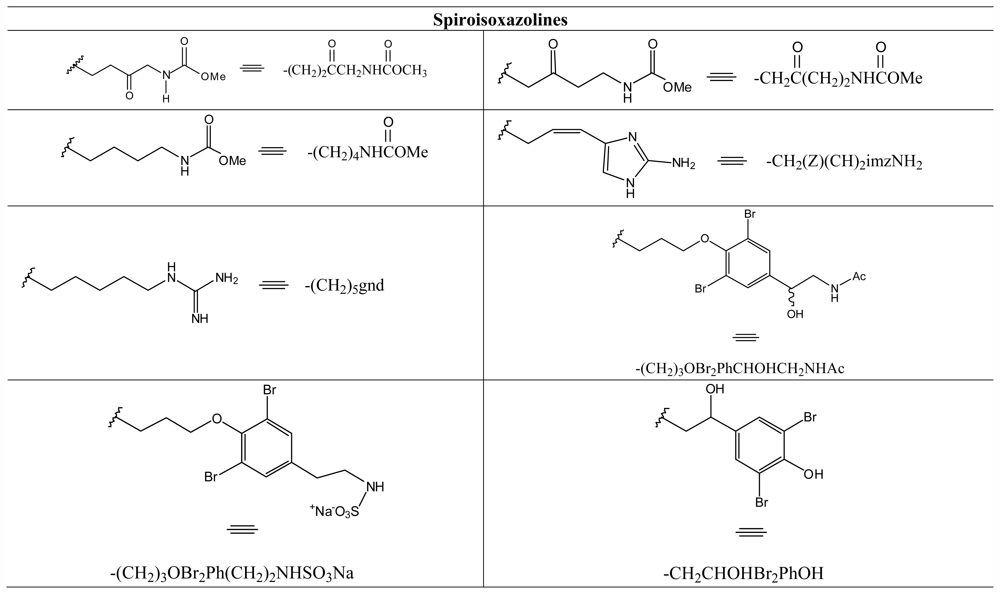
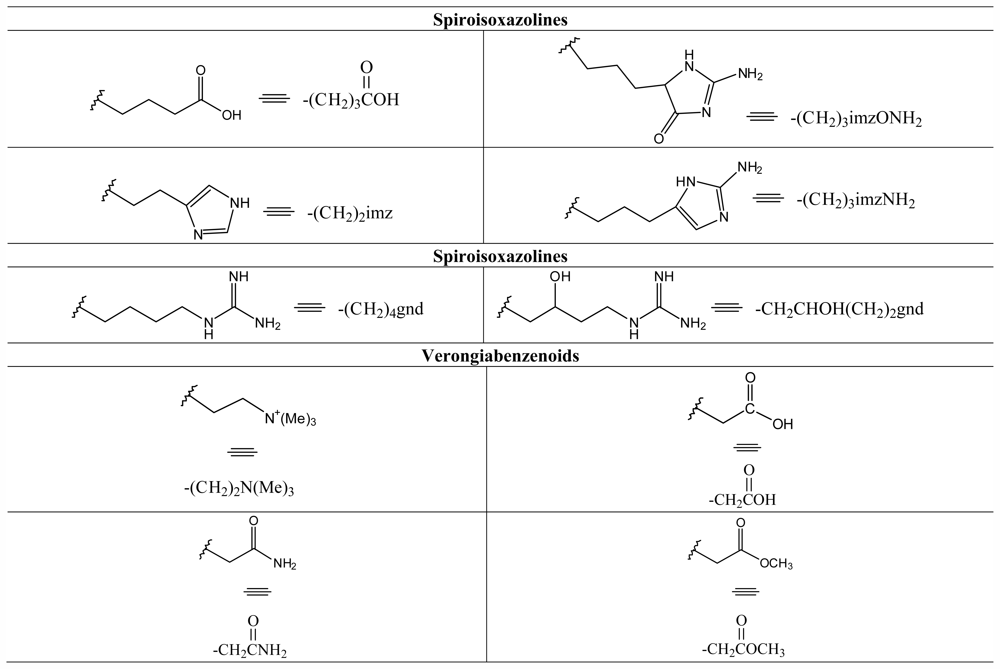
3.8. Spiroisoxazolines
3.9. Verongiabenzenoids
3.10. Verongiaquinols
3.11. Dibromocyclohexadienes
3.12. Structural Elucidation
4. Conclusions
Acknowledgements
References
- Sponge. Available online: http://www.britannica.com (accessed on 5 October 2010).
- Hentschel, U.; Hopke, J.; Horn, M.; Friedrich, A.B.; Wagner, M.; Hacker, J.; Moore, B.S. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol 2002, 68, 4431–4440. [Google Scholar]
- Radjasa, O.K.; Sabdono, A.; Junaidi, J.; Zocchi, E. Richness of secondary metabolite-producing marine bacteria associated with sponge Haliclona sp. Int. J. Pharmacol 2007, 3, 275–279. [Google Scholar]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association-a review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar]
- Ebada, S.S.; Lin, W.; Proksch, P. Bioactive sesterterpenes and triterpenes from marine sponges: Occurrence and pharmacological significance. Mar. Drugs 2010, 8, 313–346. [Google Scholar]
- Andavan, G.S.B.; Lemmens-Gruber, R. Cyclodepsipeptides from marine sponges: Natural agents for drug research. Mar. Drugs 2010, 8, 810–834. [Google Scholar]
- Balan, V.; Nangia-Makker, P.; Raz, A. Galectins as cancer biomarkers. Cancers 2010, 2, 592–610. [Google Scholar]
- Exposito, J.-Y.; Valcourt, U.; Cluzel, C.; Lethias, C. The fibrillar collagen family. Int. J. Mol. Sci 2010, 11, 407–426. [Google Scholar]
- Longeon, A.; Copp, B.R.; Quévrain, E.; Roué, M.; Kientz, B.; Cresteil, T.; Petek, S.; Debitus, C.; Bourguet-Kondracki, M.-L. Bioactive indole derivatives from the south pacific marine sponges Rhopaloeides odorabile and Hyrtios sp. Mar. Drugs 2011, 9, 879–888. [Google Scholar]
- Lee, Y.K.; Lee, J.-H.; Lee, H.K. Microbial symbiosis in marine sponges. J. Microbiol 2001, 39, 254–264. [Google Scholar]
- Belarbi, E.H.; Gómez, A.C.; Chisti, Y.; Camacho, F.G.; Grima, E.M. Producing drugs from marine sponges. Biotechnol. Adv 2003, 21, 585–598. [Google Scholar]
- Fieseler, L.; Horn, M.; Wagner, M.; Hentschel, U. Discovery of the novel candidate phylum “Poribacteria” in marine sponges. Appl. Environ. Microbiol 2004, 70, 3724–3732. [Google Scholar]
- Thoms, C.; Ebel, R.; Proksch, P. Activated chemical defense in Aplysina sponges revisited. J. Chem. Ecol 2006, 32, 97–123. [Google Scholar]
- Thakur, N.L.; Müller, W.E.G. Biotechnological potential of marine sponges. Curr. Sci. India 2004, 86, 1506–1512. [Google Scholar]
- Erpenbeck, D.; van Soest, R. Status and perspective of sponge chemosystematics. Mar. Biotechnol 2007, 9, 2–19. [Google Scholar]
- Fusetani, N.; Matsunaga, S. Bioactive sponge peptides. Chem. Rev 1993, 93, 1793–1806. [Google Scholar]
- Sagar, S.; Kaur, M.; Minneman, K.P. Antiviral lead compounds from marine sponges. Mar. Drugs 2010, 8, 2619–2638. [Google Scholar]
- Koopmans, M.; Martens, D.; Wijffels, R. Towards commercial production of sponge medicines. Mar. Drugs 2009, 7, 787–802. [Google Scholar]
- Sepčić, K.; Kauferstein, S.; Mebs, D.; Turk, T. Biological activities of aqueous and organic extracts from tropical marine sponges. Mar. Drugs 2010, 8, 1550–1566. [Google Scholar]
- Esteves, A.I.S.; Nicolai, M.; Humanes, M.; Goncalves, J. Sulfated polysaccharides in marine sponges: Extraction methods and anti-HIV activity. Mar. Drugs 2011, 9, 139–153. [Google Scholar]
- Alvarez, B.; Crisp, M.D.; Driver, F.; Hooper, J.N.A.; van Soest, R.W.M. Phylogenetic relationships of the family Axinellidae (Porifera: Demospongiae) using morphological and molecular data. Zool. Scr 2000, 29, 169–198. [Google Scholar]
- Proksch, P.; Edrada, R.; Ebel, R. Drugs from the seas—Current status and microbiological implications. Appl. Microbiol. Biot 2002, 59, 125–134. [Google Scholar]
- Sipkema, D.; Franssen, M.; Osinga, R.; Tramper, J.; Wijffels, R. Marine sponges as pharmacy. Mar. Biotechnol 2005, 7, 142–162. [Google Scholar]
- Tymiak, A.A.; Rinehart, K.L., Jr; Bakus, G.J. Constituents of morphologically similar sponges: Aplysina and Smenospongia species. Tetrahedron 1985, 41, 1039–1047. [Google Scholar]
- Rinehart, K.L., Jr; Shaw, P.D.; Shield, L.S.; Gloer, J.B.; Harbour, G.C.; Koker, M.E.S.; Samain, D.; Schwartz, R.E.; Tymiak, A.A.; Weller, D.L.; et al. Marine natural products as sources of antiviral, antimicrobial and antineoplastic agents. Pure Appl. Chem 1981, 53, 795–817. [Google Scholar]
- Goo, Y.M. Antimicrobial and antineoplastic tyrosine metabolites from a marine sponge, Aplysina fistularis. Arch. Pharm. Res 1985, 8, 21–30. [Google Scholar]
- Capon, R.; Macleod, J. Two epimeric dibromo nitriles from the Australian sponge Aplysina laevis. Aust. J. Chem 1987, 40, 341–346. [Google Scholar]
- Ajabnoor, M.A.M.; Tilmisany, A.K.; Taha, A.M.; Antonius, A. Isolation of antibiotic principles from some red sea sponges. Egypt. J. Pharm. Sci 1991, 32, 797–815. [Google Scholar]
- Gunasekera, S.P.; Cross, S.S. Fistularin 3 and 11-ketofistularin 3. feline leukemia virus active bromotyrosine metabolites from the marine sponge Aplysina archeri. J. Nat. Prod 1992, 55, 509–512. [Google Scholar]
- Rodríguez, A.D.; Piña, I.C. The structures of aplysinamisines I, II, and III: New bromotyrosine-derived alkaloids from the Caribbean sponge Aplysina cauliformis. J. Nat. Prod 1993, 56, 907–914. [Google Scholar]
- Gopichand, Y.; Schmitz, F.J. Marine natural products: Fistularin-1, -2 and -3 from the sponge Aplysina fistularis forma fulva. Tetrahedron Lett 1979, 20, 3921–3924. [Google Scholar]
- Ajabnoor, M.A.M.; Tilmisany, A.K.; Taha, A.M.; Antonius, A. Effect of red sea sponge extracts on blood glucose levels in normal mice. J. Ethnopharmacol 1991, 33, 103–106. [Google Scholar]
- Pinheiro, U.S.; Hajdu, E.; Custódio, M.R. Aplysina nardo (Porifera, Verongida, Aplysinidae) from the Brazilian coast with description of eight new species. Zootaxa 2007, 1609, 1–51. [Google Scholar]
- Ciminiello, P.; Costantino, V.; Fattorusso, E.; Magno, S.; Mangoni, A.; Pansini, M. Chemistry of Verongida sponges, II. Constituents of the Caribbean sponge Aplysina fistularis forma fulva. J. Nat. Prod 1994, 57, 705–712. [Google Scholar]
- Pauletti, P.M.; Cintra, L.S.; Braguine, C.G.; Filho, A.A.S.; Silva, M.L.A.; Cunha, W.R.; Januário, A.H. Halogenated indole alkaloids from marine invertebrates. Mar. Drugs 2010, 8, 1526–1549. [Google Scholar]
- Aiello, A.; Fattorusso, E.; Imperatore, C.; Menna, M.; Müller, W. Iodocionin, a cytotoxic iodinated metabolite from the Mediterranean Ascidian Ciona edwardsii. Mar. Drugs 2010, 8, 285–291. [Google Scholar]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar]
- Vairappan, C.S.; Ishii, T.; Lee, T.K.; Suzuki, M.; Zhaoqi, Z. Antibacterial activities of a new brominated diterpene from Borneon Laurencia spp. Mar. Drugs 2010, 8, 1743–1749. [Google Scholar]
- La Barre, S.; Potin, P.; Leblanc, C.; Delage, L. The Halogenated metabolism of brown algae (Phaeophyta), its biological importance and its environmental significance. Mar. Drugs 2010, 8, 988–1010. [Google Scholar]
- Güven, K.C.; Percot, A.; Sezik, E. Alkaloids in marine algae. Mar. Drugs 2010, 8, 269–284. [Google Scholar]
- Genovese, G.; Tedone, L.; Hamann, M.; Morabito, M. The Mediterranean red alga Asparagopsis: A source of compounds against Leishmania. Mar. Drugs 2009, 7, 361–366. [Google Scholar]
- Matta, C.B.B.; Souza, E.T.; Queiroz, A.C.; Lira, D.P.; Araújo, M.V.; Cavalcante-Silva, L.H.A.; Miranda, G.E.C.; Araújo-Júnior, J.X.; Barbosa-Filho, J.M.; Santos, B.V.O.; et al. Antinociceptive and anti-inflammatory activity from algae of the genus Caulerpa. Mar. Drugs 2011, 9, 307–318. [Google Scholar]
- Almeida, C.L.F.; Falcão, H.S.; Lima, G.R.M.; Montenegro, C.A.; Lira, N.S.; Athayde-Filho, P.F.; Rodrigues, L.C.; Souza, M.F.V.; Barbosa-Filho, J.M.; Batista, L.M. Bioactivities from marine algae of the genus Gracilaria. Int. J. Mol. Sci 2011, 12, 4550–4573. [Google Scholar]
- Souza, E.T.; Lira, D.P.; Queiroz, A.C.; Silva, D.J.C.; Aquino, A.B.; Mella, E.A.C.; Lorenzo, V.P.; Miranda, G.E.C.; Araújo-Júnior, J.X.; Chaves, M.C.O.; et al. The Antinociceptive and anti-inflammatory activities of caulerpin, a bisindole alkaloid isolated from seaweeds of the genus Caulerpa. Mar. Drugs 2009, 7, 689–704. [Google Scholar]
- Queiroz, T.M.; Machado, N.T.; Furtado, F.F.; Oliveira-Filho, A.A.; Alustau, M.C.; Figueiredo, C.S.; Miranda, G.E.C.; Barbosa-Filho, J.M.; Braga, V.A.; Medeiros, I.A. Vasorelaxation induced byDictyota pulchella(Dictyotaceae), a brown alga, is mediated via inhibition of calcium influx in rats. Mar Drugs 2011, 9, 2075–2088. [Google Scholar]
- Lira, N.S.; Monte-Neto, R.L.; Marchi, J.G.B.; Lins, A.C.S.; Pinheiro, U.S.; Tavares, J.F.; Silva, M.S.; Cunha, E.V.L.; Braz Filho, R.; Santos, C.F.; et al. Aplysfistularine: A novel dibromotyrosine derivative isolated fromAplysina fistularis. Mar. Drugs 2011. submitted. [Google Scholar]
- Lúcio, A.S.S.C.; Almeida, J.R.G.S.; Barbosa-Filho, J.M.; Pita, J.C.L.R.; Castello Branco, M.V.S.; Diniz, M.F.F.M.; Agra, M.F.; Cunha, E.V.L.; Silva, M.S.; Tavares, J.F. Azaphenathrene alkaloids with antitumoral activity from Anaxagorea dolichocarpa Sprague & Sandwith (Annonaceae). Molecules 2011, 16, 7125–7131. [Google Scholar]
- Bitencourt, M.A.O.; Dantas, G.R.; Lira, D.P.; Barbosa-Filho, J.M.; Miranda, G.E.C.; Santos, B.V.O.; Souto, J.T. Aqueous and methanolic extracts of Caulerpa mexicana suppress cell migration and ear edema induced by inflammatory agents. Mar. Drugs 2011, 9, 1332–1345. [Google Scholar]
- Alves, J.S.; Castro, J.C.M.; Freire, M.O.; da-Cunha, E.V.L.; Barbosa-Filho, J.M.; Silva, M.S. Complete assignment of the 1H and 13C NMR spectra of four triterpenes of the ursane, artane, lupane and friedelane groups. Magn. Reson. Chem 2000, 38, 201–206. [Google Scholar]
- Andrade, N.C.; Cunha, E.V.L.; Silva, M.S.; Agra, M.F.; Barbosa-Filho, J.M. Terpenoids of the Annonaceae: Distribution and Compilation of 13C NMR Data. In Recent Research Developments in Phytochemistry; Pandalai, S.G., Ed.; Research Signpost: Kerala, India, 2003; Volume 7, pp. 1–85. [Google Scholar]
- Sena-Filho, J.G.; Duringer, J.; Maia, G.L.A.; Tavares, J.F.; Xavier, H.S.; Silva, M.S.; da-Cunha, E.V.L.; Barbosa-Filho, J.M. Ecdysteroids from Vitex species: Distribution and compilation of their 13C-NMR spectral data. Chem. Biodivers 2008, 5, 707–713. [Google Scholar]
- Oliveira, S.L.; Silva, M.S.; Tavares, J.F.; Sena-Filho, J.G.; Lucena, H.F.S.; Romero, M.A.V.; Barbosa-Filho, J.M. Tropane alkaloids from Erythroxylum genus: Distribution and compilation of 13C-NMR spectral data. Chem. Biodivers 2010, 7, 302–326. [Google Scholar]
- Almeida, R.N.; Navarro, D.S.; Barbosa-Filho, J.M. Plants with central analgesic activity. Phytomedicine 2001, 8, 310–322. [Google Scholar]
- Silva, J.S.; Moura, M.D.; Oliveira, R.A.G.; Diniz, M.F.F.; Barbosa-Filho, J.M. Natural product inhibitors of ovarian neoplasia. Phytomedicine 2003, 10, 221–232. [Google Scholar]
- Falcão, H.S.; Mariath, I.R.; Diniz, M.F.F.M.; Batista, L.M.; Barbosa-Filho, J.M. Plants of the American continent with antiulcer activity. Phytomedicine 2008, 15, 132–146. [Google Scholar]
- Sousa, F.C.F.; Melo, C.T.V.; Citó, M.C.O.; Félix, F.H.C.; Vasconcelos, S.M.M.; Fonteles, M.M.F.; Barbosa Filho, J.M.; Viana, G.S.B. Plantas medicinais e seus constituintes bioativos: Uma revisão da bioatividade e potenciais benefícios nos distúrbios da ansiedade em modelos animais. Rev. Bras. Farmacogn 2008, 18, 642–654. [Google Scholar]
- Falcão, H.S.; Leite, J.A.; Barbosa-Filho, J.M.; Athayde-Filho, P.F.; Chaves, M.C.O.; Moura, M.D.; Ferreira, A.L.; Almeida, A.B.A.; Souza-Brito, A.R.M.; Diniz, M.F.F.M.; et al. Gastric and duodenal antiulcer activity of alkaloids: A review. Molecules 2008, 13, 3198–3223. [Google Scholar]
- Mota, K.S.L.; Dias, G.E.N.; Pinto, M.E.F.; Luiz-Ferreira, A.; Souza-Brito, A.R.S.; Hiruma-Lima, C.A.; Barbosa-Filho, J.M.; Batista, L.M. Flavonoids with gastroprotective activity. Molecules 2009, 14, 979–1012. [Google Scholar]
- Mariath, I.R.; Falcão, H.S.; Barbosa-Filho, J.M.; Sousa, L.C.F.; Tomaz, A.C.A.; Batista, L.M.; Diniz, M.F.F.M.; Athayde-Filho, P.F.; Tavares, J.F.; Silva, M.S.; et al. Plants of the American continent with antimalarial activity. Rev. Bras. Farmacogn 2009, 19, 158–191. [Google Scholar]
- Ribeiro Filho, J.; Falcao, H.S.; Batista, L.M.; Barbosa-Filho, J.M.; Piuvezam, M.R. Effects of Plant Extracts on HIV-1 Protease. Curr. HIV Res 2010, 8, 531–544. [Google Scholar]
- Lima, G.R.M.; Montenegro, C.A.; Almeida, C.L.F.; Athayde-Filho, P.F.; Barbosa-Filho, J.M.; Batista, L.M. Database survey of anti-inflammatory plants in South America: A review. Int. J. Mol. Sci 2011, 12, 2692–2749. [Google Scholar]
- Silva, F.L.; Fischer, D.C.H.; Tavares, J.F.; Silva, M.S.; Athayde-Filho, P.F.; Barbosa-Filho, J.M. Compilation of secondary metabolites from Bidens pilosa L. Molecules 2011, 16, 1070–1102. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S.; Pansini, M. Chemistry of verongida sponges VIII-bromocompounds from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. Tetrahedron 1997, 53, 6565–6572. [Google Scholar]
- Norte, M.; Rodriguez, M.L.; Fernández, J.J.; Eguren, L.; Estrada, D.M. Aplysinadiene and (R,R) 5 [3,5-dibromo-4-[(2-oxo-5-oxazolidinyl)] methoxyphenyl]-2-oxazolidinone, two novel metabolites from Aplysina aerophoba Synthes. Tetrahedron 1988, 44, 4973–4980. [Google Scholar]
- Borders, D.B.; Morton, G.O.; Wetzel, E.R. Structure of a novel bromine compound isolated from a sponge. Tetrahedron Lett 1974, 15, 2709–2712. [Google Scholar]
- Fattorusso, E.; Minale, L.; Sodano, G. Aeroplysinin-1, a new bromo-compound fromAplysina aerophoba. J. Chem. Soc. Chem. Comm 1970, 751–752. [Google Scholar]
- Cimino, G.; De Rosa, S.; De Stefano, S.; Self, R.; Sodano, G. The bromo-compounds of the true sponge Verongia aerophoba. Tetrahedron Lett 1983, 24, 3029–3032. [Google Scholar]
- Gunasekera, M.; Gunasekera, S.P. Dihydroxyaerothionin and aerophobin 1. Two brominated tyrosine metabolites from the deep water marine sponge Verongula rigida. J. Nat. Prod 1989, 52, 753–756. [Google Scholar]
- Kelecom, A.; Kannengiesser, G.J. Chemical constituents of Verongia sponges, II—Struture of two dibrominated compounds isolated from the Mediterranean sponge Verongia aerophoba. Ann. Acad. Braz. Sci 1979, 51, 639–641. [Google Scholar]
- Fattorusso, E.; Minale, L.; Sodano, G. Aeroplysinin-1, an antibacterial bromo-compound from the spongeVerongia aerophoba. J. Chem. Soc. Perk. Trans. 1 1972, 16–18. [Google Scholar]
- Makarieva, T.N.; Stonik, V.A.; Alcolado, P.; Elyakov, Y.B. Comparative study of the halogenated tyrosine derivatives from Demospongiae (Porifera). Comp. Biochem. Phys. B 1981, 68, 481–484. [Google Scholar]
- Mancini, I.; Guella, G.; Laboute, P.; Debitus, C.; Pietra, F. Hemifistularin 3: A degraded peptide or biogenetic precursor? Isolation from a sponge of the order Verongida from the coral sea or generation from base treatment of 11-oxofistularin 3. J. Chem. Soc. Perk. Trans. 1 1993, 3121–3125. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Magno, S. Archerine, a novel anti-histaminic bromotyrosine-derived compound from the Caribbean marine sponge Aplysina archeri. Eur. J. Org. Chem 2001, 2001, 55–60. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Magno, S.; Carrano, L.; Pansini, M. Chemistry of Verongida sponges. VII bromocompounds from the Caribbean sponge Aplysina archeri. Tetrahedron 1996, 52, 9863–9868. [Google Scholar]
- Saeki, B.M.; Granato, A.C.; Berlinck, R.G.S.; Magalhães, A.; Schefer, A.B.; Ferreira, A.G.; Pinheiro, U.S.; Hajdu, E. Two unprecedented dibromotyrosine-derived alkaloids from the Brazilian endemic marine sponge Aplysina caissara. J. Nat. Prod 2002, 65, 796–799. [Google Scholar]
- Lira, T.O.; Berlinck, R.G.S.; Nascimento, G.G.F.; Hajdu, E. Further dibromotyrosine-derived metabolites from the marine sponge Aplysina caissara. J. Braz. Chem. Soc 2006, 17, 1233–1240. [Google Scholar]
- Rodríguez, A.D.; Akee, R.K.; Scheuer, P.J. Two bromotyrosine-cysteine derived metabolites from a sponge. Tetrahedron Lett 1987, 28, 4989–4992. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Magno, S.; Pansini, M. Chemistry of Verongida sponges. 9. Secondary metabolite composition of the Caribbean sponge Aplysina cauliformis. J. Nat. Prod 1999, 62, 590–593. [Google Scholar]
- D’Ambrosio, M.; Guerriero, A.; Traldi, P.; Pietra, F. Cavernicolin-1 and cavernicolin-2, epimeric dibromolactams from the Mediterranean sponge Aplysina (Verongia) cavernicola. Tetrahedron Lett 1982, 23, 4403–4406. [Google Scholar]
- D’Ambrosio, M.; Guerriero, A.; de Clauser, R.; de Stanchina, G.; Pietra, F. Dichloroverongiaquinol, a new marine antibacterial compound from Aplysina cavernicola. Isolation and synthesis. Cell. Mol. Life Sci 1983, 39, 1091–1092. [Google Scholar]
- D’Ambrosio, M.; Guerriero, A.; Pietra, F. Novel, racemic or nearly-racemic antibacterial bromoand chloroquinols and γ-lactams of the verongiaquinol and the cavernicolin type from the marine sponge Aplysina (= Verongia) cavernicola. Helv. Chim. Acta 1984, 67, 1484–1492. [Google Scholar]
- D’Ambrosio, M.; Mealli, C.; Guerriero, A.; Pietra, F. 7-Bromocavernicolenone, a new α-bromoenone from the Mediterranean sponge Aplysina (=Verongia) cavernicola. Implied, unprecedented involvement of a halogenated dopa in the biogenesis of a natural product. Helv. Chim. Acta 1985, 68, 1453–1460. [Google Scholar]
- Pereira, F.R.; Marques, S.O.; Nascimento, A.M.; Gandolfi, R.C.; Hajdu, E.; Peixinho, S.; Custódio, M.R.; Berlinck, R.G.S. Metabólitos Derivados da Dibromotirosina das Esponjas Aplysina fulva, A. conulosa (sp. n.) and A. fistularis. In XXIX Reunião Anual da Sociedade Brasileira de Química; Sociedade Brasileira de Química: Águas de Lindóia, Brazil, 2006; p. 335. [Google Scholar]
- Xynas, R.; Capon, R.J. Two new bromotyrosine-derived metabolites from an Australian marine sponge, Aplysina sp. Aust. J. Chem 1989, 42, 1427–1433. [Google Scholar]
- Tymiak, A.A. Structure elucidation and biosynthesis of selected bioactive marine natural products. Dissert. Abs. Int. B 1985, 43, 3990–3991. [Google Scholar]
- Rinehart, K.L., Jr.; Johnson, R.D.; Paul, I.C.; McMillan, J.A.; Siuda, J.F.; Krejcarek, G.E. Identification of Compounds in Selected Marine Organisms by Gas Chromatography-Mass Spectrometry, Field Desorption Mass Spectrometry, and Other Physical Methods. In Food—Drugs from the Sea: Proceedings, 1974; Webber, H.H., Ruggieri, G.D., Eds.; Marine Technology Society: Columbia, MD, USA, 1976. [Google Scholar]
- Sharma, G.M.; Vig, B.; Burkholder, P.R. Antimicrobial substances of sponges. IV. Structure of a bromine-containing compound from a marine sponge. J. Org. Chem 1970, 35, 2823–2826. [Google Scholar]
- Gandolfi, R.C.; Medina, M.B.; Berlinck, R.G.S.; Lira, S.P.; Galetti, F.C.S.; Silva, C.L.; Veloso, K.; Ferreira, A.G.; Hajdu, E.; Peixinho, S. Metabólitos secundários das esponjas Aplysina fistularis e Dysidea sp. e atividade antituberculose da 11-cetofistularina-3. Quim. Nova 2010, 33, 1853–1858. [Google Scholar]
- Walker, R.P.; Thompson, J.E.; Faulkner, D.J. Exudation of biologically-active metabolites in the sponge Aplysina fistularis. II. Chemical evidence. Mar. Biol 1985, 88, 27–32. [Google Scholar]
- Kobayashi, J.I.; Tsuda, M.; Agemi, K.; Shigemori, H.; Ishibashi, M.; Sasaki, T.; Mikami, Y. Purealidins B and C, new bromotyrosine alkaloids from the Okinawan marine sponge Psammaplysilla purea. Tetrahedron 1991, 47, 6617–6622. [Google Scholar]
- Krejcarek, G.E.; White, R.H.; Hager, L.P.; McClure, W.O.; Johnson, R.D.; Rinehart, K.L.; McMillan, J.A.; Paul, I.C.; Shaw, P.D.; Brusca, R.C. A rearranged dibromotyrosine metabolite from Verongia aurea. Tetrahedron Lett 1975, 16, 507–510. [Google Scholar]
- Sharma, G.M.; Burkholder, P.R. Studies on antimicrobial substances of sponges. I. Isolation, purification, and properties of a new bromine-containing antibacterial substance. J. Antibiot 1967, 20, 200–203. [Google Scholar]
- Sharma, G.M.; Burkholder, P.R. Studies on the antimicrobial substances of sponges II. Structure and synthesis of a bromine-containing antibacterial, compound from a marine sponge. Tetrahedron Lett 1967, 8, 4147–4150. [Google Scholar]
- Minale, L. Natural product chemistry of the marine sponges. Pure Appl. Chem 1976, 48, 7–23. [Google Scholar]
- Minale, L.; Cimino, G.; de Stefano, S.; Sodano, G. Natural products from Porifera. Fortschr. Chem. Org. Naturst 1976, 33, 1–72. [Google Scholar]
- Carney, J.R.; Rinehart, K.L. Biosynthesis of brominated tyrosine metabolites by Aplysina fistularis. J. Nat. Prod 1995, 58, 971–985. [Google Scholar]
- Nuñez, C.V.; Almeida, E.V.R.; Granato, A.C.; Marques, S.O.; Santos, K.O.; Pereira, F.R.; Macedo, M.L.; Ferreira, A.G.; Hajdu, E.; Pinheiro, U.S.; et al. Chemical variability within the marine sponge Aplysina fulva. Biochem. Syst. Ecol 2008, 36, 283–296. [Google Scholar]
- Rogers, E.W.; Oliveira, M.F.; Berlinck, R.G.S.; König, G.M.; Molinski, T.F. Stereochemical heterogeneity in Verongid sponge metabolites. absolute stereochemistry of (+)-fistularin-3 and (+)-11-epi-fistularin-3 by microscale LCMS-Marfey’s analysis. J. Nat. Prod 2005, 68, 891–896. [Google Scholar]
- Rogers, E.W.; Molinski, T.F. Highly polar spiroisoxazolines from the sponge Aplysina fulva. J. Nat. Prod 2007, 70, 1191–1194. [Google Scholar]
- Encarnación, R.D.; Sandoval, E.; Malmstrøm, J.; Christophersen, C. Calafianin, a bromotyrosine derivative from the marine sponge Aplysina gerardogreeni. J. Nat. Prod 2000, 63, 874–875. [Google Scholar]
- Hernández-Guerrero, C.J.; Zubía, E.; Ortega, M.J.; Carballo, J.L. Cytotoxic dibromotyrosine-derived metabolites from the sponge Aplysina gerardogreeni. Bioorg. Med. Chem 2007, 15, 5275–5282. [Google Scholar]
- Fendert, T.; Wray, V.; van Soest, R.W.M.; Proksch, P. Bromoisoxazoline alkaloids from the Caribbean sponge Aplysina insularis. Z. Naturforsch 1999, 54c, 246–252. [Google Scholar]
- Compagnone, R.S.; Avila, R.; Suárez, A.I.; Abrams, O.V.; Rangel, H.R.; Arvelo, F.; Piña, I.C.; Merentes, E. 11-Deoxyfistularin-3, a new cytotoxic metabolite from the Caribbean sponge Aplysina fistularis insularis. J. Nat. Prod 1999, 62, 1443–1444. [Google Scholar]
- Acosta, A.L.; Rodríguez, A.D. 11-Oxoaerothionin: A cytotoxic antitumor bromotyrosine-derived alkaloid from the Caribbean marine sponge Aplysina lacunosa. J. Nat. Prod 1992, 55, 1007–1012. [Google Scholar]
- Evan, T.; Rudi, A.; Ilan, M.; Kashman, Y. Aplyzanzine A, a new dibromotyrosine derivative from a Verongida sponge. J. Nat. Prod 2001, 64, 226–227. [Google Scholar]
- Kossuga, M.H.; Lira, S.P.; Nascimento, A.M.; Gambardella, M.T.P.; Berlinck, R.G.S.; Torres, Y.R.; Nascimento, G.G.F.; Pimenta, E.F.; Silva, M.; Thiemann, O.H.; et al. Isolamento e atividades biológicas de produtos naturais das esponjas Monanchora arbuscula, Aplysina sp. Petromica ciocalyptoides e Topsentia ophiraphidites, da ascídia Didemnum ligulum e do octocoral Carijoa riisei. Quim. Nova 2007, 30, 1194–1202. [Google Scholar]
- Kashman, Y.; Groweiss, A.; Carmely, S.; Kinamoni, Z.; Czarkie, D.; Rotem, M. Recent research in marine natural products from the red sea. Pure Appl. Chem 1982, 54, 1995–2010. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Magno, S.; Pansini, M. Chemistry of Verongida sponges. 10. Secondary metabolite composition of the Caribbean Sponge Verongula gigantea. J. Nat. Prod 2000, 63, 263–266. [Google Scholar]
- Cruz, F.; Quijano, L.; Gómez-Garibay, F.; Rios, T. Brominated metabolites from the sponge Aplysina (Verongia) thiona. J. Nat. Prod 1990, 53, 543–548. [Google Scholar]
- Ayanoglu, E.; Djerassi, C.; Erdman, T.R.; Scheuer, P.J. Minor and trace sterols in marine invertebrates V. isolation, structure elucidation and synthesis of 3[beta]-hydroxy-26,27-bisnorcholest-5-en-24-one from the sponge Psammaplysilla purpurea. Steroids 1978, 31, 815–822. [Google Scholar]
- Carballeira, N.M.; Bou, C.R. The sterol composition of the marine sponge Aplysina (=Verongia) archeri: A comparative study of the Verongidae. Comp. Biochem. Phys. B 1989, 93, 175–179. [Google Scholar]
- Hertzberg, S.; Bergquist, P.; Liaaen-Jensen, S. Further occurrence of sulphated carotenoids in Ianthella species (Desmospongia). Biochem. Syst. Ecol 1989, 17, 51–53. [Google Scholar]
- Nichols, S.A. An evaluation of support for order-level monophyly and interrelationships within the class Demospongiae using partial data from the large subunit rDNA and cytochrome oxidase subunit I. Mol. Phylogenet. Evol 2005, 34, 81–96. [Google Scholar]
- Lawson, M.; Thompson, J.; Djerassi, C. Localization of long-chain fatty acids and unconventional sterols in spherulous cells of a marine sponge. Lipids 1988, 23, 1037–1048. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Pansini, M. Chemistry of Verongida sponges VI. Comparison of the secondary metabolic composition of Aplysina insularis and Aplysina fulva. Biochem. Syst. Ecol 1996, 24, 105–113. [Google Scholar]
- Hamann, M.T.; Scheuer, P.J.; Kelly-Borges, M. Biogenetically diverse, bioactive constituents of a sponge, order Verongida: Bromotyramines and sesquiterpene-shikimate derived metabolites. J. Org. Chem 1993, 58, 6565–6569. [Google Scholar]
- D’Ambrosio, M.; Guerriero, A.; Pietra, F. N-Carbamoylpyrrolidine and 7-chlorocavernicolenone, two new metabolites of the Mediterranean sponge Aplysina (=Verongia) cavernicola. Comp. Biochem. Phys. B 1986, 83, 309–312. [Google Scholar]
- Cavazza, M.; Guella, G.; Nucci, L.; Pergola, F.; Bicchierini, N.; Pietra, F. Anodic oxidation of 3,5-dihalogenotyrosines as a model reaction for the biogenesis of the cavernicolins, metabolites of the Verongid spongeAplysina cavernicola. J. Chem. Soc. Perk. Trans. 1 1993, 3117–3120. [Google Scholar]
- Calcul, L.; Inman, W.D.; Morris, A.A.; Tenney, K.; Ratnam, J.; McKerrow, J.H.; Valeriote, F.A.; Crews, P. Additional insights on the bastadins: Isolation of analogues from the sponge Ianthella cf. reticulata and exploration of the oxime configurations. J. Nat. Prod 2010, 73, 365–372. [Google Scholar]
- Andersen, R.J.; Faulkner, D.J. A novel antibiotic from a sponge of the genus Verongia. Tetrahedron Lett 1973, 14, 1175–1178. [Google Scholar]
- Kasperek, G.J.; Bruice, T.C.; Yagi, H.; Kaubsich, N.; Jerina, D.M. Solvolytic chemistry of 1,4-dimethylbenzene oxide. A new and novel mechanism for the NIH shift. J. Am. Chem. Soc 1972, 94, 7876–7882. [Google Scholar]
- Huang, X.-P.; Deng, Z.-W.; van Soest, R.W.M.; Lin, W.-H. Brominated derivatives from the Chinese sponge Pseudoceratina sp. J. Asian Nat. Prod. Res 2008, 10, 239–242. [Google Scholar]
- Kossuga, M.H.; MacMillan, J.B.; Rogers, E.W.; Molinski, T.F.; Nascimento, G.G.F.; Rocha, R.M.; Berlinck, R.G.S. (2S,3R)-2-Aminododecan-3-ol, a new antifungal agent from the Ascidian Clavelina oblonga. J. Nat. Prod 2004, 67, 1879–1881. [Google Scholar]
- Peng, J.; Li, J.; Hamann, M.T. The Marine Bromotyrosine Derivatives. In The Alkaloids: Chemistry and Biology; Cordell, G.A., Ed.; Academic Press: Gulf Breeze, FL, USA, 2005; Volume 61, p. 59. [Google Scholar]
- Aiello, A.; Fattorusso, E.; Menna, M.; Pansini, M. Chemistry of Verongida sponges—V. Brominated metabolites from the Caribbean sponge Pseudoceratina sp. Biochem. Syst. Ecol 1995, 23, 377–381. [Google Scholar]
- Moody, K.; Thomson, R.H.; Fattorusso, E.; Minale, L.; Sodano, G. Aerothionin and homoaerothionin: Two tetrabromo spirocyclohexadienylisoxazoles from Verongia sponges. J. Chem. Soc. Perk. Trans 1 1972, 1, 18–24. [Google Scholar]
- Bergquist, P.R. Sponges; University of California Press: Berkeley and Los Angeles, CA, USA, 1978. [Google Scholar]
- Teeyapant, R.; Proksch, P. Biotransformation of brominated compounds in the marine sponge Verongia aerophoba—Evidence for an induced chemical defense? Naturwissenschaften 1993, 80, 369–370. [Google Scholar]
- Ebel, R.; Brenzinger, M.; Kunze, A.; Gross, H.J.; Proksch, P. Wound Activation of protoxins in marine sponge Aplysina aerophoba. J. Chem. Ecol 1997, 23, 1451–1462. [Google Scholar]
- Puyana, M.; Fenical, W.; Pawlik, J.R. Are there activated chemical defenses in sponges of the genus Aplysina from the Caribbean? Mar. Ecol. Prog. Ser 2003, 246, 127–135. [Google Scholar]
- Weiss, B.; Ebel, R.; Elbrächter, M.; Kirchner, M.; Proksch, P. Defense metabolites from the marine sponge Verongia aerophoba. Biochem. Syst. Ecol 1996, 24, 1–7. [Google Scholar]
- Koulman, A.; Proksch, P.; Ebel, R.; Beekman, A.C.; van Uden, W.; Konings, A.W.T.; Pedersen, J.A.; Pras, N.; Woerdenbag, H.J. Cytoxicity and mode of action of aeroplysinin-1 and a related dienone from the sponge Aplysina aerophoba. J. Nat. Prod 1996, 59, 591–594. [Google Scholar]
- Nagaraja, K.V.; Shaw, P.D. Inhibition of wheat germ RNA polymerase II by 2,6-dibromobenzoquinone and related compounds from Aplysina fistularis. Arch. Biochem. Biophys 1982, 215, 544–550. [Google Scholar]
- Chang, C.W.J.; Weinheimer, A.J. 2-Hydroxy, 3,5-dibromo, 4-methoxyphenylacetamide. a dibromotyrosine metabolite from Psammopilla purpurea. Tetrahedron Lett 1977, 18, 4005–4007. [Google Scholar]
- Fulmor, W.; Van Lear, G.E.; Morton, G.O.; Mills, R.D. Isolation and absolute configuration of the aeroplysinin I enantiomorphic pair from Ianthella ardis. Tetrahedron Lett 1970, 11, 4551–4552. [Google Scholar]
- Gutiérrez, M.; Capson, T.L.; Guzmán, H.M.; González, J.; Ortega-Barría, E.; Quiñoá, E.; Riguera, R. Antiprotozoal activity against Plasmodium falciparum and Trypanosoma cruzi of aeroplysinin-1 isolated from the new sponge Aplysina chiriquensis. Pharm. Biol 2005, 43, 762–765. [Google Scholar]
- Córdoba, R.; Tormo, N.S.; Medarde, A.F.; Plumet, J. Antiangiogenic versus cytotoxic activity in analogues of aeroplysinin-1. Bioorg. Med. Chem 2007, 15, 5300–5315. [Google Scholar]
- Simões, C.M.O.; Petrovick, P.R. Farmacognosia: Da Planta ao Medicamento, 6 ed; Editora da UFSC: Santa Catarina, Brasil, 2010. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
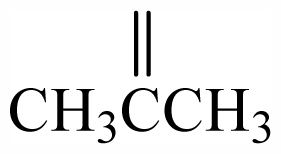{kind=link}
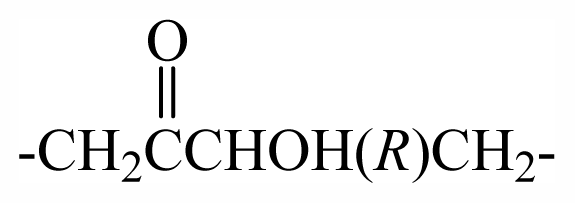{kind=link}
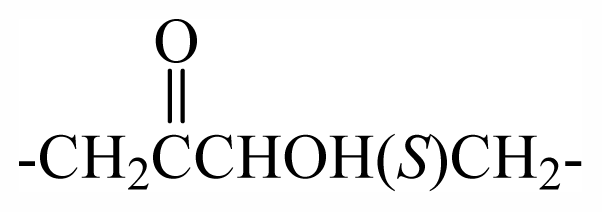{kind=link}
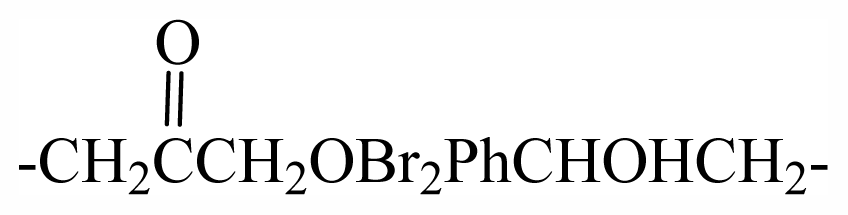{kind=link}
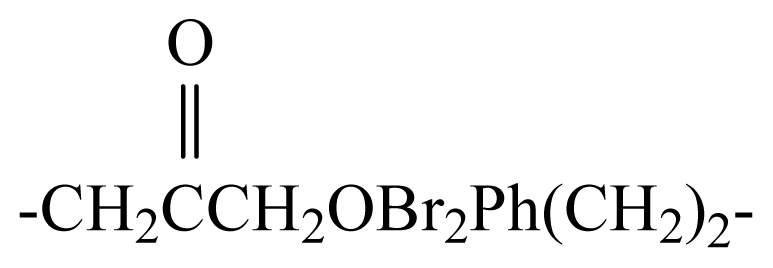{kind=link}
| Activity/Species Name | Type of Extract | Bioassays Models, Organism, Dose or Route of Administration | Result | Ref. |
|---|---|---|---|---|
| Antibacterial activity | ||||
| Aplysina archeri | MeOH Ext. | Agar plate-Bacillus subtilis; Escherichia coli-1.0 mg/Disc | Active | [24] |
| Aplysina fistularis | MeOH Ext. | Agar plate-Bacillus subtilis; Escherichia coli | Active | [25] |
| MeOH-Toluene | Agar plate-Bacillus subtilis; Escherichia coli | Active | [26] | |
| Chromatographic Fraction | Agar plate-Staphylococcus aureus; Sarcinalutea; Klebsiella pneumonia; Proteus vulgaris; Bacteroides fragilis; Clostridium perfringens; Mycobacterium aviun | Active | [26] | |
| Aplysina lacunosa | MeOH Ext. | Agar plate-Bacillus subtilis; Escherichia coli | Active | [25] |
| MeOH Ext. | Agar plate-Bacillus subtilis; Escherichia coli-1.0 mg/Disc | Active | [24] | |
| Aplysina laevis | Acetone Ext. | Agar plate-Bacillus subtilis; Escherichia coli | Active | [27] |
| Aplysina mollis | Ether Ext. | Agar plate-Staphylococcus aureus-0.2μL/Disc | Active | [28] |
| Ether Ext. | Agar plate-Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] | |
| CHCl3 Ext. | Agar plate-Staphylococcus aureus-0.2 μL/Disc | Active | [28] | |
| CHCl3 Ext. | Agar plate-Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] | |
| Acetone Ext. | Agar plate-Staphylococcus aureus-0.2 μL/Disc | Inactive | [28] | |
| ETOH (95%) Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] | |
| Benzene Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] | |
| Aplysina species | Ether Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] |
| Acetone Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] | |
| ETOH (95%) Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] | |
| Antibacterial activity | Benzene Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] |
| Aplysina species | CHCl3 Ext. | Agar plate-Staphylococcus aureus; Escherichia coli; Pseudomonas aeruginosa-0.2 μL/Disc | Inactive | [28] |
| Antiyeast activity | ||||
| Aplysina archeri | MeOH Ext. | Agar plate-Saccharomyces cerevisiae-1.0 mg/Disc | Inactive | [24] |
| Aplysina lacunosa | MeOH Ext. | Agar plate-Saccharomyces cerevisiae-1.0 mg/Disc | Weak | [24] |
| Activity | ||||
| Antifungal activity | ||||
| Aplysina archeri | MeOH Ext. | Agar plate-Penicillium atrovenetum-1.0 mg/Disc | Inactive | [24] |
| Aplysina lacunosa | MeOH Ext. | Agar plate-Penicillium atrovenetum-1.0 mg/Disc | Inactive | [24] |
| Antiviral activity | ||||
| Aplysina archeri | MeOH-Toluene | Cell culture-Virus-Feline Leukemia | Active | [29] |
| Cytotoxic activity | ||||
| Aplysina cauliformis | CHCl3-MeOH Ext. (1:1) | Cell culture-Cells-Cho K-1 | Active | [30] |
| Aplysina fistularis | MeOH Ext. | Cell culture-Leuk L-1210-ED50 50 mcg/mL | Active | [25] |
| MeOH-Toluene | Cell culture-CA-9KB | Active | [26] | |
| Chromatographic Fraction | Cell culture-Leuk L-1210-IC50 0.14 mcg/mL | Active | [26] | |
| Aplysina fulva | Isopropanol Ext. | Cell culture-CA-9KB < ED50 20 mcg/mL | Active | [31] |
| Isopropanol Ext. | Cell culture-Leuk L-1210 < ED50 20 mcg/mL | Active | [31] | |
| Isopropanol Ext. | Cell culture-Leuk P-388 < ED50 20 mcg/mL | Active | [31] | |
| Aplysina lacunosa | MeOH Ext. | Cell culture-Leuk L-1210-ED50 8.2 mcg/mL | Inactive | [25] |
| Hyperglycemic activity | ||||
| Aplysina mollis | ETOH (95%) Ext. | Mouse-Intragastric-Dose 200 mg/kg | Active | [32] |
| Species | Halogenated Substances | Substance Code | Nucleus | Ref. |
|---|---|---|---|---|
| A. aerophoba (Schmidt, 1862) | Aeroplysinine 2 | 25 | E1 | [63] |
| Aplysinadiene | 26 | E1 | [64] | |
| (7S*,11R*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 29 | F1 | [65] | |
| (R,R)-5[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)] methoxyphenyl]-2-oxazolidone | 31 | F1 | [64] | |
| Aerothionin | 41 | G1 | [34] | |
| Homoaerothionin | 47 | G2 | [66] | |
| Isofistularin-3 | 48 | G2 | [67] | |
| Aerophobin-1 | 76 | G7 | [68] | |
| 2-(3,5-Dibromo-2-hydroxy-4-methoxyphenyl) acetamide | 84 | H | [69] | |
| 3,5-Dibromo-4-methoxyphenol | 85 | H | [64] | |
| Methyl 2-(3,5-dibromo-2-hydroxy-4-methoxyphenyl) acetate | 86 | H | [69] | |
| Dibromoverongiaquinol or dienone or 3-5-dibromo-1-hydroxy-4-oxocyclohexa-2-5-diene-1-acetamide | 94 | I1 | [64] | |
| Aeroplysinin 1 | 100 | J | [66,70] | |
| A. archeri (Higgin, 1875) | (7S*,11R*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 29 | F1 | [71] |
| 11,19-Dideoxyfistularin 3 | 33 | G1 | [72] | |
| Archerine | 43 | G1 | [73] | |
| Fistularin-3 | 46 | G2 | [29] | |
| 11-Ketofistularin 3 | 51 | G2 | [29] | |
| Aplysina compound 1 or 1-Oxa-2-azaspiro[4,5]deca-2,6-diene-3-carboxamide, N,N′-(2-oxo-1,4-butanediyl)bis[7,9-dibromo-10-hydroxy-8-oxo, [5R-[5α(5′R*,9′R*,10′S*),9α,10β]]-(9CI) | 57 | G4 | [74] | |
| Aeroplysinin 1 | 100 | J | [74] | |
| (+) Aeroplysinin 1 | 100 | J | [71] | |
| (−) Aeroplysinin 1 | 100 | J | [71] | |
| A. caissara (Pinheiro & Hajdu, 2001) | 2-(3,5-Dibromo-4,4-dimethoxy-1-hydroxy-2,5-cyclohexadien-1-yl) acetamide | 21 | D1 | [75] |
| Caissarine C | 42 | G1 | [76] | |
| Caissarine B | 53 | G2 | [75] | |
| Agelocaissarine A1 | 58 | G4 | [76] | |
| Agelocaissarine A2 | 59 | G4 | [76] | |
| Agelocaissarine B1 | 60 | G4 | [76] | |
| Agelocaissarine B2 | 61 | G4 | [76] | |
| Caissarine A | 79 | G8 | [75] | |
| A. cauliformis (Carter, 1882) | 2-(3,5-Dibromo-1-hydroxy-4,4-dimethoxycyclohexa-2,5-dienyl)acetamide | 21 | D1 | [71] |
| (7S*,11R*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 29 | F1 | [71] | |
| 11-Oxoaerothionin | 52 | G2 | [77] | |
| Aplysinametabolite or Methyl 4-((5S,10R)-7,9-dibromo-10-hydroxy-8-methoxy-1-oxa-2-azaspiro-[4.5]deca-2,6,8-trienecarboxamido)-2-oxobutylcarbamate | 64 | G7 | [78] | |
| Methyl 4-((5S,10R)-7,9-dibromo-10-hydroxy-8-methoxy-1-oxa-2-azaspiro-[4.5]deca-2,6,8-trienecarboxamido)-3-oxobutylcarbamate or Aplysina compound 13 | 65 | G7 | [78] | |
| Methyl-4-((5S,10R)-7,9-dibromo-10-hydroxy-8-methoxy-1-oxa-2-azaspiro[4.5]deca-2,6,8-trienecarboxamido)butylcarbamate or Aplysina metabolite 14 | 66 | G7 | [78] | |
| Aplysinamisine-1 | 67 | G7 | [30] | |
| Aplysinamisine-2 | 68 | G7 | [30] | |
| Aplysinamisine-3 | 69 | G7 | [30] | |
| Aeroplysinin 1 | 100 | J | [78] | |
| A. cavernicola (Vacelet, 1959) | Cavernicolin-1 | 5 | B1 | [79,80] |
| 5-Bromo-7α-chlorocavernicolin | 7 | B2 | [81] | |
| 5-Bromo-7β-chlorocavernicolin | 8 | B2 | [81] | |
| 7β-Bromo-5-chlorocavernicolin | 9 | B2 | [81] | |
| 7α-Bromo-5-chlorocavernicolin | 10 | B2 | [81] | |
| Monobromocavernicolin or 5-Bromocavernicolin | 11 | B2 | [81] | |
| 5-Chlorocavernicolin | 12 | B2 | [81,82] | |
| 7-Bromocavernicolenone | 13 | B3 | [82] | |
| 7-Chlorocavernicolenone | 14 | B3 | [63] | |
| 2-(3,5-Dibromo-1-hydroxy-4,4-dimethoxycyclohexa-2,5-dienyl) acetamide | 21 | D1 | [63] | |
| Aeroplysinine 2 | 25 | E1 | [63] | |
| 11,19-Dideoxyfistularin 3 | 33 | G1 | [63] | |
| 12(R)-Hydroxy-11-oxoaerothionin | 36 | G1 | [63] | |
| Aerothionin | 41 | G1 | [63] | |
| Oxohomoaerothionin | 44 | G1 | [63] | |
| 11-Deoxyfistularin-3 | 45 | G2 | [63] | |
| Homoaerothionin | 47 | G2 | [63] | |
| Isofistularin3 | 48 | G2 | [63] | |
| 11-Oxoaerothionin | 52 | G2 | [63] | |
| (+) 3-Bromo-5-chloroverongiaquinol or (+)-3-Bromo-5-chloro-1-hydroxy-4-oxo-2,5-cyclohexadiene-1-acetamide | 91 | I1 | [81] | |
| (+) 3-Bromoverongiaquinol or (+)-3-Bromo-1-hydroxy-4-oxo-2,5-cyclohexadiene-1-acetamide | 92 | I1 | [81] | |
| (DL) 5-Bromoverongiaquinol | 93 | I1 | [81] | |
| Dichloroverongiaquinol | 95 | I1 | [80] | |
| Aeroplysinin 1 | 100 | J | [63] | |
| A. conulosa (Pulitzer-Finali, 1986) | Aeroplysinine 2 | 25 | E1 | [83] |
| A. fistularis (Pallas, 1766) | Aplysamine1 | 1 | A | [84] |
| Aplysamine2 | 2 | A | [84] | |
| Aplysfistularine | 4 | A | [46] | |
| 5-Amino-2,6-dichloro-4-hydroxycyclohex-2-enone acetic acid lactam | 15 | B4 | [26] | |
| 5-Amino-2-bromo-6-chloro-4-hydroxy-cyclohex-2-enone acetic acid lactam (5-bromo-7-chlorocavernicolin) | 16 | B4 | [26] | |
| 5-Amino-2-6-dibromo-4-hydroxy-cyclohex-2-enone acetic acid lactam or Cavernicolin | 17 | B4 | [26] | |
| 4,6-Dibromohomogentisamide | 18 | C | [85] | |
| 3,5-Dibromohydroquinone-2-acetamide | 19 | C | [86] | |
| 2-(3,5-Dibromo-1-hydroxy-4,4-dimethoxycyclohexa-2,5-dienyl) acetamide | 21 | D1 | [87] | |
| Aeroplysinine 2 | 25 | E1 | [88] | |
| (7S*,11R*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 29 | F1 | [71] | |
| Aerothionin | 41 | G1 | [88,89] | |
| Fistularin-3 | 46 | G2 | [88] | |
| Homoaerothionin | 47 | G2 | [89] | |
| 11-Oxoaerothionin | 52 | G2 | [88] | |
| Purealidin-L | 78 | G7 | [90] | |
| 2-(3,5-Dibromo-2-hydroxy-4-methoxyphenyl) acetamide | 84 | H | [26,69] | |
| 2,6-Dibromo-1,4-benzoquinone | 87 | I1 | [91] | |
| 2,6-Dichloro-4-hydroxycyclohexa-2-5-dienone-4-acetamide | 89 | I1 | [26] | |
| 2-Bromo-6-chloro-4-hydroxycyclohexa-2,5-dienone-4-acetamide | 90 | I1 | [26] | |
| Dibromoverongiaquinol or dienone or 3-5-dibromo-1-hydroxy-4-oxocyclohexa-2-5-diene-1-acetamide | 94 | I1 | [61–65,92–96] | |
| Aeroplysinin 1 | 100 | J | [96] | |
| A. fulva (Pallas, 1766) | Cavernicolin-1 | 5 | B1 | [83,97] |
| Cavernicolin-2 | 6 | B1 | [83,97] | |
| 3,5-Dibromohydroquinone-2-acetamide | 19 | C | [34] | |
| 2’-(3,5-Dibromo-4-hydroxyphenyl) acetamide | 20 | C | [97] | |
| 2-(3,5-Dibromo-1-hydroxy-4,4-dimethoxycyclohexa-2,5-dienyl) acetamide | 21 | D1 | [97] | |
| Aeroplysinine 2 | 25 | E1 | [34] | |
| (7S*,11R*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 29 | F1 | [97] | |
| 11-Epi-fistularin-3 | 34 | G1 | [98] | |
| 11-Hydroxyfistularin-3 | 35 | G1 | [99] | |
| 12(R)-Hydroxy-11-oxoaerothionine | 36 | G1 | [34] | |
| 12(S)-Hydroxy-11-oxoaerothionine | 37 | G1 | [34] | |
| Aerothionin | 41 | G1 | [34,83,97] | |
| Fistularin-3 | 46 | G2 | [31,34,83,97] | |
| Homoaerothionin | 47 | G2 | [34] | |
| 11-Hydroxyaerothionin | 50 | G2 | [34,97] | |
| 11-Oxoaerothionin | 52 | G2 | [34,97] | |
| Aplysinamisine-1 | 67 | G7 | [97] | |
| AraplysillinN9-sulfamate | 70 | G7 | [99] | |
| Fistularin-1 | 72 | G7 | [34] | |
| Fistularin-2 | 73 | G7 | [31] | |
| N-[5S,10R)-7,9-Dibromo-10-hydroxy-8-methoxy-1-oxa-2-azaspiro[4.5]deca-2,6,8-triene-3-carboxy]-4-aminobutanoic acid | 74 | G7 | [99] | |
| Aerophobin-1 | 76 | G7 | [97] | |
| Aerophobin-2 | 77 | G7 | [97] | |
| Aeroplysinin 1 | 100 | J | [34,66] | |
| Aplysinafulvin | 101 | J | [97] | |
| A. gerardogreeni (Gomes & Bakus, 1992) | Aerothionin | 41 | G1 | [100] |
| Homoaerothionin | 47 | G2 | [101] | |
| Aplysinone A | 54 | G3 | [101] | |
| Aplysinone D | 55 | G3 | [101] | |
| Aplysinone B | 56 | G4 | [101] | |
| Calafianin | 62 | G5 | [100] | |
| Aplysinone C | 63 | G6 | [101] | |
| 2-(3,5-dibromo-2-hydroxy-4-methoxyphenyl) acetic acid | 83 | H | [100] | |
| A. insularis (Duchassaing & Michelotti, 1864) | 5-((2,6-Dibromo-4-(2-oxooxazolidin-5-yl)-phenoxy)-methyl)-5-methoxyoxazolidin-2-one | 32 | F3 | [74] |
| 11,19-Dideoxyfistularin 3 | 33 | G1 | [102] | |
| Aerothionin | 41 | G1 | [74,103] | |
| Fistularin-3 | 46 | G2 | [74,102,103] | |
| Homoaerothionin | 47 | G2 | [103] | |
| 11-Dihydroaerothionin | 49 | G2 | [102] | |
| 11-Oxoaerothionin | 52 | G2 | [102,103] | |
| Aplysina metabolite 14 | 66 | G7 | [74] | |
| 14-Oxoaerophobin-2 | 75 | G7 | [102] | |
| Aerophobin-1 | 76 | G7 | [102] | |
| Aerophobin-2 | 77 | G7 | [102] | |
| (5S,10R)-Methyl 7,9-dibromo-10-hydroxy-8-methoxy-1-oxa-2-azaspiro[4.5]deca-2,6,8-triene-3-carboxylate | 80 | G9 | [102] | |
| 2-(3-Dibromo-4-hydroxyphenyl)-N,N,N-trimethylethanaminium | 82 | H | [102] | |
| A. lacunose (Lamarck, 1814) | (7S*,11R*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 29 | F1 | [65] |
| (7R*,11S*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 30 | F2 | [71] | |
| 11,19-Dideoxyfistularin-3 | 33 | G1 | [104] | |
| Aerothionin | 41 | G1 | [104] | |
| Fistularin-3 | 46 | G2 | [104] | |
| 11-Hydroxyaerothionin | 50 | G2 | [104] | |
| 11-Oxoaerothionin | 52 | G2 | [104] | |
| A. laevis (Carter, 1885) | (1′R,5′R,6′S)-2-(3′,5′-Dibromo-1′,6′-dihydroxy-4′-oxo-cyclohex-2′-enyl) acetonitrile | 98 | I2 | [27] |
| (1′R,5’S,6′S)-2-(3′,5′-Dibromo-1′,6′-dihydroxy-4′-oxo-cyclohex-2′-enyl) acetonitrile | 99 | I2 | [27] | |
| (+) Aeroplysinin 1 | 100 | J | [27] | |
| A. species | Aplysamine1 | 1 | A | [84] |
| Aplysamine2 | 2 | A | [84] | |
| Aplyzanzine A | 3 | A | [105] | |
| 2-(3,5-Dibromo-4-ethoxy-1-hydroxy-4-methoxy-2,5-cyclohexadien-1-yl)-ethanamide | 22 | D2 | [71,84,106] | |
| Aeroplysinine 2 | 25 | E1 | [106] | |
| (7R*,11S*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidinone | 30 | F2 | [71] | |
| (R,R)-5[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]methoxyphenyl]-2-oxazolidone | 31 | F1 | [106] | |
| 11,19-Dideoxyfistularin-3 | 33 | G1 | [72] | |
| 11-Oxofistularin-3 | 38 | G1 | [72] | |
| 19-Deoxy-11-oxofistularin | 39 | G1 | [72] | |
| 19-Deoxyfistularin-3 | 40 | G1 | [72] | |
| Aerothionin | 41 | G1 | [107] | |
| Hemifistularin-3 | 71 | G7 | [72] | |
| (10R)-Ethyl-7,9-dibromo-10-hydroxy-8-methyl-1-oxa-2-azaspiro[4.5]deca-2,6,8-triene-3-carboxylate | 81 | G9 | [106] | |
| 2-(3-Bromo-4-hydroxyphenyl)-N,N,N-trimethylethanaminium | 82 | H | [108] | |
| A. thiona (Laubenfels, 1950) | Aplysinketal A | 23 | D1 | [109] |
| Aplysinketal B | 24 | D1 | [109] | |
| Aplysinolide | 27 | E2 | [109] | |
| Aplysinimine | 28 | E2 | [109] | |
| (7R*,11S*)-5-[3,5-Dibromo-4-[(2-oxo-5-oxazolidinyl)]-methoxyphenyl]-2-oxazolidinone | 30 | F2 | [109] | |
| Aerothionin | 41 | G1 | [109] | |
| Homoaerothionin | 47 | G2 | [109] | |
| 2-(3,5-Dibromo-2-hydroxy-4-methoxyphenyl)-acetamide | 84 | H | [109] | |
| 2,6-Dibromo-4-acetamide-4-hydroxycyclohexadienone | 88 | I1 | [109] | |
| Aplysina hydroxydienone or Dibromo compound 10 | 96 | I1 | [109] | |
| Aplysina hydroxydienoic methyl esther | 97 | I1 | [109] |


| Halogen Compound | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | R9 | Nucleus |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Me | Me | - | - | - | - | - | - | - | A |
| 2 | H | (E) BrMeOPhCH2CNOHCO- | - | - | - | - | - | - | - | A |
| 3 | H | Br2MeOPhCH2CHN(CH3)2CO- | - | - | - | - | - | - | - | A |
| 4 | Me | Me | Me | - | - | - | - | - | - | A |
| 5 | Br | H | Br | - | - | - | - | - | - | B1 |
| 6 | H | Br | Br | - | - | - | - | - | - | B1 |
| 7 | Cl | H | Br | - | - | - | - | - | - | B2 |
| 8 | H | Cl | Br | - | - | - | - | - | - | B2 |
| 9 | Br | H | Cl | - | - | - | - | - | - | B2 |
| 10 | H | Br | Cl | - | - | - | - | - | - | B2 |
| 11 | H | H | Br | - | - | - | - | - | - | B2 |
| 12 | H | H | Cl | - | - | - | - | - | - | B2 |
| 13 | Br | - | - | - | - | - | - | - | - | B3 |
| 14 | Cl | - | - | - | - | - | - | - | - | B3 |
| 15 | H | Cl | Cl | - | - | - | - | - | - | B4 |
| 16 | H | Cl | Br | - | - | - | - | - | - | B4 |
| 17 | H | Br | Br | - | - | - | - | - | - | B4 |
| 18 | OH | H | Br | OH | Br | - | - | - | - | C |
| 19 | Br | OH | Br | H | OH | - | - | - | - | C |
| 20 | H | Br | OH | Br | H | - | - | - | - | C |
| 21 | Me | Me | - | - | - | - | - | - | - | D1 |
| 22 | Et | Me | - | - | - | - | - | - | - | D2 |
| 23 | Me | Butyl | - | - | - | - | - | - | - | D1 |
| 24 | Me | Pentyl | - | - | - | - | - | - | - | D1 |
| 25 | MeO | OH | H | - | - | - | - | - | - | E1 |
| 26 | H | Br | (E) CH2=CH=CH=CH- | OH | Br | - | - | - | - | E1 |
| 27 | Br | OMe |  | Br | H | - | - | - | - | E2 |
| 28 | Br | OMe | H | Br | H | - | - | - | - | E2 |
| 29 | -CH2(R)oxz | - | - | - | - | - | - | - | - | F1 |
| 30 | -CH2(S)oxz | - | - | - | - | - | - | - | - | F2 |
| 31 | -CH2(R)oxz | - | - | - | - | - | - | - | - | F1 |
| 32 | -CH2MeOoxz | - | - | - | - | - | - | - | - | F3 |
| 33 | -(CH2)3OBr2Ph(CH2)2- | - | - | - | - | - | - | - | - | G1 |
| 34 | -CH2(R)CHOHCH2OBr2PhCHOHCH2- | - | - | - | - | - | - | - | - | G1 |
| 35 | -(CH2)3OBr2Ph(R)CHOHCH2- | - | - | - | - | - | - | - | - | G1 |
| 36 |  | - | - | - | - | - | - | - | - | G1 |
| 37 |  | - | - | - | - | - | - | - | - | G1 |
| 38 |  | - | - | - | - | - | - | - | - | G1 |
| 39 |  | - | - | - | - | - | - | - | - | G1 |
| 40 | -CH2CHOHCH2OBr2Ph(CH2)2- | - | - | - | - | - | - | - | - | G1 |
| 41 | -(CH2)4- | - | - | - | - | - | - | - | - | G1 |
| 42 | -CH2CHOH(CH2)3- | - | - | - | - | - | - | - | - | G1 |
| 43 | -(CH2)2CH(imz)2(CH2)3- | - | - | - | - | - | - | - | - | G1 |
| 44 |  | - | - | - | - | - | - | - | - | G1 |
| 45 | -(CH2)3OBr2PhCHOHCH2- | - | - | - | - | - | - | - | - | G2 |
| 46 | -CH2(S)CHOHCH2OBr2PhCHOHCH2- | - | - | - | - | - | - | - | - | G2 |
| 47 | -(CH2)5- | - | - | - | - | - | - | - | - | G2 |
| 48 | -CH2CHOHCH2OBr2PhCHOHCH2- | - | - | - | - | - | - | - | - | G2 |
| 49 | -CH2(CHOH)2CH2- | - | - | - | - | - | - | - | - | G2 |
| 50 | -CH2CHOH(CH)2- | - | - | - | - | - | - | - | - | G2 |
| 51 |  | - | - | - | - | - | - | - | - | G2 |
| 52 | -CH2CH2O(CH2)2- | - | - | - | - | - | - | - | - | G2 |
| 53 | -(CH2)5COHCH2- | - | - | - | - | - | - | - | - | G2 |
| 54 | -(CH2)5- | - | - | - | - | - | - | - | - | G3 |
| 55 | -(CH2)4- | - | - | - | - | - | - | - | - | G3 |
| 56 | -(CH2)5- | H | Br | OH | H | Br | H | H | OH | G4 |
| 57 | -CH2CH2O(CH2)2- | Br | H | H | OH | H | Br | OH | H | G4 |
| 58 | -CH2CHOH(CH)2- | H | Br | OH | H | Br | H | H | OH | G4 |
| 59 | -CH2CHOH(CH)2- | Br | H | OH | H | H | Br | H | OH | G4 |
| 60 | -CH2CHOH(CH2)3- | H | Br | OH | H | Br | H | H | OH | G4 |
| 61 | -CH2CHOH(CH2)3- | Br | H | OH | H | H | Br | H | OH | G4 |
| 62 | -(CH2)4- | - | - | - | - | - | - | - | - | G5 |
| 63 | -(CH2)5- | - | - | - | - | - | - | - | - | G6 |
| 64 |  | - | - | - | - | - | - | - | - | G7 |
| 65 |  | - | - | - | - | - | - | - | - | G7 |
| 66 |  | - | - | - | - | - | - | - | - | G7 |
| 67 | -CH2(Z)(CH)2imzNH2 | - | - | - | - | - | - | - | - | G7 |
| 68 | -(CH2)5gnd | - | - | - | - | - | - | - | - | G7 |
| 69 | -(CH2)3OBr2PhCHOHCH2NHAc | - | - | - | - | - | - | - | - | G7 |
| 70 | -(CH2)3OBr2Ph(CH2)2NHSO3Na | - | - | - | - | - | - | - | - | G7 |
| 71 | -CH2CHOHBr2PhOH | - | - | - | - | - | - | - | - | G7 |
| 72 | -(CH2)3OBr2Phoxz | - | - | - | - | - | - | - | - | G7 |
| 73 | -CH2Br2PhOoxz | - | - | - | - | - | - | - | - | G7 |
| 74 |  | - | - | - | - | - | - | - | - | G7 |
| 75 | -(CH2)3imzONH2 | - | - | - | - | - | - | - | - | G7 |
| 76 | -(CH2)2imz | - | - | - | - | - | - | - | - | G7 |
| 77 | -(CH2)3imzNH2 | - | - | - | - | - | - | - | - | G7 |
| 78 | -(CH2)4gnd | - | - | - | - | - | - | - | - | G7 |
| 79 | -CH2CHOH(CH2)2gnd | - | - | - | - | - | - | - | - | G8 |
| 80 | Me | - | - | - | - | - | - | - | - | G9 |
| 81 | Et | - | - | - | - | - | - | - | - | G9 |
| 82 | -(CH2)2N(Me)3 | H | - | - | - | - | - | - | - | H |
| 83 |  | OH | - | - | - | - | - | - | - | H |
| 84 |  | OH | - | - | - | - | - | - | - | H |
| 85 | OH | H | - | - | - | - | - | - | - | H |
| 86 |  | OH | - | - | - | - | - | - | - | H |
| 87 | Br | H | H | O | Br | - | - | - | - | I1 |
| 88 | Br | H | OH |  | Br | - | - | - | - | I1 |
| 89 | Cl | H | OH |  | Cl | - | - | - | - | I1 |
| 90 | Br | H | OH |  | Cl | - | - | - | - | I1 |
| 91 | Br | OH | OH |  | Cl | - | - | - | - | I1 |
| 92 | Br | OH | OH |  | H | - | - | - | - | I1 |
| 93 | H | OH | OH |  | Br | - | - | - | - | I1 |
| 94 | Br | H | OH |  | Br | - | - | - | - | I1 |
| 95 | Cl | H | OH |  | Cl | - | - | - | - | I1 |
| 96 | Br | H | OH |  | Br | - | - | - | - | I1 |
| 97 | Br | H | O | CH2CH2OCH3 | Br | - | - | - | - | I1 |
| 98 | OH | OH | - | - | - | - | - | - | - | I2 |
| 99 | OH | OH | - | - | - | - | - | - | - | I2 |
| 100 | OH | OH | CH2CN | - | - | - | - | - | - | J |
| 101 | MeO | OH | CH2CONH2 | - | - | - | - | - | - | J |
| Position | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| 1 | 140.3 | 140.3 | 137.94 | 138.58 |
| 2 | 134.4 | 134.4 | 132.78 | 132.80 |
| 3 | 118.7 | 118.7 | 117.74 | 118.02 |
| 4 | 152.1 | 152.1 | 150.87 | 151.30 |
| 5 | 118.7 | 118.7 | 117.74 | 118.02 |
| 6 | 134.4 | 134.4 | 132.78 | 132.80 |
| 7 | 35.2 | 35.2 | 34.20 | 32.26 |
| 8 | 41.3 | 41.3 | 39.81 | 60.20 |
| 9 | - | 165.8 | 170.82 | - |
| 10 | - | 152.9 | 69.84 | - |
| 11 | - | 28.7 | 31.57 | - |
| 12 | - | 113.1 | 137.66 | - |
| 13 | - | 134.7 | 133.21 | - |
| 14 | - | 130.3 | 117.58 | - |
| 15 | - | 155.8 | 152.34 | - |
| 16 | - | 112.1 | 117.58 | - |
| 17 | - | 131.7 | 133.21 | - |
| 1’ | 71.7 | 71.7 | 69.71 | 71.00 |
| 2’ | 26.4 | 26.4 | 25.38 | 26.79 |
| 3’ | 56.9 | 56.9 | 55.41 | 55.77 |
| MeO | - | 56.7 | 60.39 | - |
| +N(Me)2 | 43.7/43.6 | 43.7 | 41.51/42.92 | 44.06 |
| +N(Me)3 | - | - | - | 44.74 |
| Position | 5 | 6 | 9 | 11 | 12 | 13 | 14 |
|---|---|---|---|---|---|---|---|
| 1 | 184.0 | 184.0 | 183.4 | 188.91 | 188.89 | 80.90 | 82.7 |
| 2 | 56.9 | 53.1 | 58.0 | 39.78 | 40.08 | - | - |
| 3 | 68.7 | 64.7 | 67.9 | 58.60 | 58.56 | 168.30 | 171.4 |
| 4 | 76.6 | 75.7 | 74.4 | 74.15 | 73.41 | 43.1 | 44.0 |
| 5 | 149.9 | 149.0 | 146.3 | 150.07 | 145.84 | 69.5 | 69.5 |
| 6 | 120.8 | 120.8 | 127.4 | 122.45 | 130.18 | 156.90 | 153.2 |
| 7 | 43.0 | 45.1 | 42.4 | 43.58 | 43.78 | 118.80 | 129.5 |
| 8 | 173.9 | 173.9 | 173.6 | 172.65 | 172.72 | 187.8 | 188.9 |
| 9 | - | - | - | - | - | 46.6 | 47.6 |
| Position | 20 |
|---|---|
| 1 | 130.0 |
| 2 | 133.0 |
| 3 | 111.0 |
| 4 | 150.0 |
| 5 | 111.0 |
| 6 | 133.0 |
| 2’ | 40.0 |
| 3’ | 173.4 |
| Position | 22 | 23 |
|---|---|---|
| 1 | 71.3 | 123.55 |
| 2 | 140.3 | 123.55 |
| 3 | 124.1 | 142.11 |
| 4 | 96.7 | 71.92 |
| 5 | 124.1 | 142.11 |
| 6 | 140.3 | 123.55 |
| 7 | 44.2 | 45.03 |
| 8 | 173.2 | 172.99 |
| 9 | 51.3 | 51.04 |
| 10 | 60.2 | 63.98 |
| 11 | 15.4 | 32.34 |
| 12 | - | 20.04 |
| 13 | - | 14.10 |
| 14 | - | - |
| Position | 25 | 26 | 27 | 28 |
|---|---|---|---|---|
| 1 | 89.1 | 148.85 | 154.29 | 161.3 |
| 2 | 135.5 | 109.12 | 100.74 | 106.7 |
| 3 | 119.3 | 103.75 | 161.97 | 155.2 |
| 4 | 151.2 | 147.16 | 111.80 | 110.8 |
| 5 | 107.6 | 103.75 | 125.75 | 135.2 |
| 6 | 77.0 | 113.68 | 122.35 | 117.0 |
| 7 | 42.2 | 146.35 | 117.53 | 40.6 |
| 8 | 173.7 | 165.91 | 165.23 | 164.0 |
| 9 | - | 144.60 | - | - |
| 10 | - | 128.12 | 150.75 | - |
| 11 | - | 128.12 | 25.21 | - |
| 12 | - | 19.79 | 23.69 | - |
| MeO | 61.5 | - | 61.0 | 60.8 |
| Position | 29 | 30 | 31 | 32 |
|---|---|---|---|---|
| 1 | 153.5 | 151.6 | 142.3 | 140.45 |
| 2 | 119.3 | 117.6 | 118.2 | 118.63 |
| 3 | 131.9 | 130.7 | 130.8 | 131.45 |
| 4 | 140.6 | 138.9 | 140.0 | 152.60 |
| 5 | 131.9 | 130.7 | 130.8 | 131.45 |
| 6 | 119.3 | 117.6 | 142.3 | 118.63 |
| 7 | 76.3 | 74.3 | 54.3 | 75.78 |
| 8 | 47.9 | 46.9 | 47.0 | 48.42 |
| 9 | 159.8 | 158.5 or 158.2 | 157.8 | 159.05 |
| 10 | 75.0 | 73.0 | 53.0 | 72.05 |
| 11 | 75.5 | 73.5 | 53.55 | |
| 12 | 41.7 | 41.1 | 41.47 | 46.97 |
| 13 | 160.2 | 161.35 | 158.4 | 161.35 |
| MeO | - | - | - | 50.25 |
| Position | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 75.28 | 75.0 | 75.42 | 75.47 | 75.47 | 75.29 | 75.11 | 75.09 | 75.5 |
| 2 | 122.07 | 122.0 | 122.74 | 114.14 | 114.14 | 122.11 | 122.11 | 122.10 | 114.2 |
| 3 | 148.77 | 148.6 | 149.28 | 149.28 | 149.28 | 148.82 | 148.73 | 148.75 | 149.3 |
| 4 | 113.81 | 113.7 | 114.16 | 122.78 | 122.78 | 113.85 | 113.85 | 113.78 | 122.7 |
| 5 | 132.37 | 132.0 | 132.24 | 132.23 | 132.23 | 132.24 | 132.27 | 132.28 | 133.2 |
| 6 | 91.67 | 91.8 | 92.48 | 92.64 | 92.64 | 90.04 | 91.95 | 91.56 | 92.6 |
| 7 | 40.13 | 39.8 | 40.13 | 40.09 | 40.09 | 39.99 | 39.87 | 40.03 | 40.1 |
| 8 | 155.24 | 154.9 | 155.27 | 155.12 | 155.12 | 154.80 | 154.79 | 155.14 | 155.5 |
| 9 | 160.07 | 160.4 | 161.81 | 161.93 | 161.93 | 160.29 | 160.24 | 160.00 | 161.6 |
| 1′ | 75.28 | 75.1 | 75.42 | 75.47 | 75.47 | 75.29 | 75.18 | 75.20 | 75.5 |
| 2′ | 122.07 | 122.1 | 122.74 | 114.14 | 114.14 | 122.11 | 122.11 | 122.10 | 114.2 |
| 3′ | 148.77 | 148.6 | 149.28 | 149.28 | 149.28 | 148.82 | 148.74 | 148.75 | 149.3 |
| 4′ | 113.81 | 113.7 | 114.16 | 122.78 | 122.78 | 113.85 | 113.85 | 113.78 | 122.7 |
| 5′ | 132.30 | 132.1 | 132.16 | 133.21 | 133.21 | 132.23 | 132.28 | 132.28 | 133.2 |
| 6′ | 91.67 | 91.8 | 92.48 | 92.54 | 92.54 | 91.86 | 91.59 | 91.76 | 92.6 |
| 7′ | 40.09 | 39.9 | 40.04 | 40.03 | 40.03 | 39.89 | 40.09 | 40.08 | 40.1 |
| 8′ | 155.14 | 155.0 | 154.12 | 154.92 | 154.92 | 155.16 | 155.16 | 155.16 | 155.5 |
| 9′ | 160.01 | 160.5 | 161.55 | 161.80 | 161.80 | 160.48 | 160.03 | 160.41 | 161.6 |
| 10 | 37.47 | 43.4 | 37.94 | 49.87 | 49.87 | 47.52 | 47.46 | 43.49 | 38.4 |
| 11 | 30.55 | 69.7 | 30.59 | 207.84 | 207.84 | 200.82 | 200.87 | 69.68 | 26.1 |
| 12 | 72.16 | 75.7 | 71.61 | 75.73 | 75.73 | 76.52 | 76.48 | 75.79 | 26.1 |
| 13 | 152.30 | 152.5 | 153.56 | 43.43 | 43.43 | 151.93 | 151.23 | 151.97 | 38.4 |
| 14 | 118.48 | 118.3 | 118.99 | - | - | 118.11 | 118.07 | 118.31 | - |
| 15 | 134.06 | 131.3 | 131.69 | - | - | 131.55 | 134.21 | 134.14 | - |
| 16 | 139.63 | 142.9 | 143.06 | - | - | 143.93 | 140.53 | 139.88 | - |
| 17 | 134.06 | 131.3 | 131.69 | - | - | 131.55 | 134.21 | 134.14 | - |
| 18 | 118.48 | 118.3 | 118.99 | - | - | 118.11 | 118.07 | 118.31 | - |
| 19 | 34.75 | 71.3 | 72.18 | - | - | 71.46 | 34.74 | 34.71 | - |
| 20 | 40.96 | 47.5 | 47.63 | - | - | 47.60 | 40.85 | 40.91 | - |
| MeO | 60.21 | 60.2 | 60.35 | 60.39 | 60.39 | 60.23 | 60.19 | 60.19 | 60.4 |
| Position | 42 | 43 | 44 | 45 | 46 | 49 | 50 | 51 | 52 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 72.4 | 75.41 | 75.50 | 74.60 | 74.73 | 73.55 | 73.8 | 74.67 | 73.55 |
| 2 | 119.6 | 114.24 | 120.80 | 121.66 | 121.80 | 120.81 | 121.4 | 121.85 | 120.88 |
| 3 | 145.9 | 149.29 | 149.31 | 147.92 | 148.06 | 147.15 | 147.7 | 148.06 | 147.13 |
| 4 | 111.9 | 122.75 | 114.15 | 115.16 | 115.20 | 113.09 | 113.1 | 115.20 | 113.08 |
| 5 | 130.1 | 132.30 | 131.32 | 132.15 | 132.36 | 131.25 | 130.6 | 132.30 | 131.20 |
| 6 | 89.0 | 92.39 | 92.63 | 91.72 | 91.87 | 90.32 | 91.9 | 91.87 | 90.50 |
| 7 | 38.5 | 40.26 | 40.21 | 40.27 | 40.30 | 42.53 | 38.7 | 40.30 | 39.70 |
| 8 | 153.3 | 155.34 | 155.29 | 155.10 | 155.17 | 154.47 | 153.9 | 155.18 | 154.37 |
| 9 | 157.4 | 161.44 | 161.81 | 160.05 | 160.52 | 158.98 | 160.0 | 160.46 | 159.11 |
| 1′ | 72.4 | 75.51 | 75.50 | 74.67 | 74.73 | 73.55 | 73.8 | 74.67 | 73.55 |
| 2′ | 119.6 | 114.24 | 120.80 | 121.66 | 121.80 | 120.81 | 121.3 | 121.85 | 20.84 |
| 3′ | 145.9 | 149.29 | 149.31 | 147.92 | 148.06 | 147.15 | 147.7 | 148.06 | 147.11 |
| 4′ | 111.9 | 122.75 | 114.15 | 115.16 | 115.20 | 113.09 | 113.1 | 115.20 | 113.08 |
| 5′ | 130.1 | 132.30 | 132.25 | 132.31 | 132.36 | 131.25 | 130.6 | 132.30 | 131.20 |
| 6′ | 88.9 | 92.45 | 92.36 | 91.78 | 91.93 | 90.32 | 91.8 | 91.13 | 90.24 |
| 7′ | 38.5 | 40.26 | 40.04 | 40.27 | 40.30 | 42.53 | 38.7 | 40.16 | 39.45 |
| 8′ | 153.4 | 155.41 | 154.93 | 155.23 | 155.25 | 154.47 | 153.9 | 154.77 | 154.05 |
| 9′ | 157.4 | 161.57 | 161.81 | 160.44 | 160.52 | 158.98 | 160.0 | 160.56 | 158.86 |
| 10 | 44.2 | 39.07 | 49.17 | 37.13 | 43.95 | 39.04 | 36.2 | 47.49 | 48.52 |
| 11 | 67.2 (65.7)c | 33.92 | 206.18 | 30.37 | 69.47 | 71.04 | 68.0 | 201.34 | 204.43 |
| 12 | 30.6 | 33.51 | 37.57 | 71.51 | 76.13 | 71.04 | 45.0 | 76.32 | 38.59 |
| 13 | 24.0 | 135.87 | 24.11 | 152.27 | 152.29 | 39.02 | 33.6 | 151.22 | 33.82 |
| 14 | 35.0 (32.5)c | 150.68 | 39.61 | 118.35 | 118.42 | - | - | 118.06 | - |
| 15 | - | 111.32 | - | 130.90 | 131.09 | - | - | 131.10 | - |
| 16 | - | 126.67 | - | 143.35 | 143.52 | - | - | 144.23 | - |
| 17 | - | 149.52 | - | 130.90 | 131.09 | - | - | 131.10 | - |
| 18 | - | 126.09 | - | 118.35 | 118.42 | - | - | 118.06 | - |
| 19 | - | 23.09 | - | 70.70 | 69.47 | - | - | 70.70 | - |
| 20 | - | 30.10 | - | 47.99 | 48.15 | - | - | 48. 01 | - |
| 21 | - | 40.03 | - | - | - | - | - | - | - |
| MeO | 58.4 | 60.44 | 60.38 | 59.75 | 59.86 | 59,63 | 60.0 | 59.86 | 59.63 |
| Position | 53 | 54 | 55 | 56 | 57 | 58 | 59 | 60 | 61 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 74.1 | 75.3 | 75.2 | 75.0 | 74.73 | 74.5 | 74.0 | 74.5 | 68.5 |
| 2 | 113.5 | 113.9 | 113.9 | 57.4 | 57.11 | 57.2 | 55.0 | 57.2 | 54.6 |
| 3 | 147.6 | 148.7 | 148.7 | 183.7 | 183.51 | 184.1 | 184.1 | 184.2 | 183.0 |
| 4 | 121.2 | 122.0 | 122.0 | 122.5 | 122.45 | 122.7 | 123.0 | 122.7 | 123.2 |
| 5 | 131.7 | 132.4 | 132.4 | 149.5 | 149.09 | 149.3 | 146.2 | 149.4 | 146.3 |
| 6 | 90.7 | 91.5 | 91.5 | 91.4 | 91.67 | 91.5 | 90.5 | 91.4 | 90.5 |
| 7 | 40.0 | 40.3 | 40.2 | 38.6 | 38.24 | 38.2 | 41.4 | 38.2 | 41.4 |
| 8 | 155.0 | 155.4 | 155.3 | 154.9 | 154.53 | 154.8 | 155.5 | 154.8 | 155.5 |
| 9 | 159.3 | 159.9 | 159.9 | 159.7 | 159.76 | 160.1 | 160.1 | 160.1 | 159.7 |
| 1′ | 74.1 | 75.1 | 75.0 | 75.0 | 74.78 | 74.5 | 74.0 | 74.5 | 68.5 |
| 2′ | 113.5 | 57.4 | 57.5 | 57.4 | 57.07 | 57.2 | 55.0 | 57.2 | 54.6 |
| 3′ | 147.6 | 183.7 | 183.7 | 183.7 | 183.51 | 184.1 | 184.1 | 184.2 | 183.0 |
| 4′ | 121.2 | 122.5 | 122.5 | 122.5 | 122.36 | 122.7 | 123.0 | 122.7 | 123.2 |
| 5′ | 131.7 | 149.5 | 149.5 | 149.5 | 149.17 | 149.3 | 146.2 | 149.4 | 146.3 |
| 6′ | 90.7 | 91.5 | 91.5 | 91.4 | 91.42 | 91.5 | 90.5 | 91.4 | 90.5 |
| 7′ | 40.0 | 38.6 | 38.6 | 38.6 | 38.10 | 38.2 | 41.4 | 38.2 | 41.4 |
| 8′ | 155.0 | 154.9 | 154.9 | 154.9 | 154.27 | 154.8 | 155.5 | 154.8 | 155.5 |
| 9′ | 159.3 | 159.7 | 159.7 | 159.7 | 159.64 | 160.1 | 160.1 | 160.1 | 159.7 |
| 10 | 45.7 | 39.7 | 39.5 | 39.7 | 49.17 | 45.9 | 45.9 | 45.9 | 45.9 |
| 11 | 67.4 and 68.9 | 29.7 | 27.5 | 29.7 | 204.70 | 68.8 | 68.8 | 70.3 | 70.3 |
| 12 | 32.2 | 24.7 | 27.5 | 24.7 | 39.63 | 34.6 | 34.6 | 32.4 | 32.4 |
| 13 | 25.5 | 29.7 | 39.5 | 29.7 | 34.78 | 37.0 | 37.0 | 26.0 | 26.0 |
| 14 | 36.6 | 39.7 | - | 39.7 | - | - | - | 37.0 | 37.0 |
| 15 | 34.4 | - | - | - | - | - | - | - | - |
| 16 | 39.4 | - | - | - | - | - | - | - | - |
| MeO | 60.1 | 60.2 | 60.2 | - | - | - | - | - | - |
| Position | 62 | 63 | 64 | 65 | 66 | 67 | 68 | 69 | 70 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 56.9 | 75.3 | 73.55 | 73.56 | 73.57 | 75.45 | 73.44 | 75.40 | 73.6 |
| 2 | 53.0 | 113.8 | 120.80 | 120.79 | 120.86 | 122.68 | 120.64 | 122.72 | 120.8 |
| 3 | 186.0 | 149.0 | 147.13 | 147.12 | 147.16 | 149.23 | 147.07 | 149.23 | 147.3 |
| 4 | 122.8 | 122.0 | 113.04 | 113.04 | 113.11 | 114.12 | 113.00 | 114.10 | 113.1 |
| 5 | 143.7 | 132.4 | 131.19 | 131.21 | 131.23 | 132.21 | 131.22 | 132.14 | 131.2 |
| 6 | 84.0 | 91.5 | 90.16 | 90.24 | 90.52 | 92.39 | 90.12 | 92.55 | 90.2 |
| 7 | 43.6 | 40.2 | 39.50 | 39.50 | 39.33 | 40.19 | 39.50 | 40.06 | 39.3 |
| 8 | 154.9 | 155.3 | 154.34 | 154.34 | 154.08 | 155.32 | 154.44 | 155.11 | 154.3 |
| 9 | 158.3 | 159.3 | 158.37 | 158.87 | 159.13 | 161.33 | 158.82 | 161.61 | 158.9 |
| 1′ | 56.9 | 58.1 | - | - | - | - | - | - | - |
| 2′ | 53.0 | 54.2 | - | - | - | - | - | - | - |
| 3′ | 186.0 | 186.0 | - | - | - | - | - | - | - |
| 4′ | 122.8 | 124.1 | - | - | - | - | - | - | - |
| 5′ | 143.7 | 144.1 | - | - | - | - | - | - | - |
| 6′ | 84.0 | 85.3 | - | - | - | - | - | - | - |
| 7′ | 43.6 | 44.5 | - | - | - | - | - | - | - |
| 8′ | 154.9 | 155.6 | - | - | - | - | - | - | - |
| 9′ | 158.3 | 159.9 | - | - | - | - | - | - | - |
| 10 | 38.6 | 39.8 | 33.83 | 49.89 | 48.58 | 39.49 | 38.53 | 37.86 | 36.2 |
| 11 | 26.4 | 29.7 | 38.29 | 205.66 | 205.75 | 123.47 | 28.23 | 30.76 | 29.4 |
| 12 | 26.4 | 24.7 | 205.66 | 38.29 | 39.75 | 122.33 | 23.28 | 72.30 | 71.2 |
| 13 | 38.6 | 29.7 | 49.89 | 33.83 | 35.21 | 130.81 | 27.99 | 153.61 | 150.8 |
| 14 | - | 39.7 | 156.98 | 156.98 | 157.03 | 151.45 | 40.65 | 118.94 | 117.0 |
| 15 | - | - | - | - | - | 118.11 | 157.20 | 131.66 | 133.0 |
| 16 | - | - | - | - | - | - | - | 142.82 | 140.6 |
| 17 | - | - | - | - | - | - | - | 131.66 | 133.0 |
| 18 | - | - | - | - | - | - | - | 118.94 | 117.0 |
| 19 | - | - | - | - | - | - | - | 71.64 | 33.7 |
| 20 | - | - | - | - | - | - | - | 47.62 | 44.7 |
| 21 | - | - | - | - | - | - | - | 173.29 | - |
| 22 | - | - | - | - | - | - | - | 47.62 | - |
| 23 | - | - | - | - | - | - | - | 22.68 | - |
| MeO-C3 | - | 60.2 | 59.60 | 59.60 | 59.79 | 60.42 | 59.56 | 60.42 | 59.6 |
| MeO-C14 | - | - | 51.54 | 51.54 | 51.28 | - | - | - | - |
| Position | 71 | 74 | 75 | 76 | 79 | 80 | 81 |
|---|---|---|---|---|---|---|---|
| 1 | 75.21 | 73.4 | 75.5 | 73.57 | 76.4 | 76.3 | 75.1 |
| 2 | 122.15 | 120.8 | 114.1 | 120.87 | 115.1 | 114.9 | 113.8 |
| 3 | 148.78 | 147.0 | 149.3 | 147.18 | 150.2 | 150.1 | 149.0 |
| 4 | 113.90 | 113.3 | 122.8 | 113.08 | 123.7 | 123.5 | 122.3 |
| 5 | 132.27 | 131.2 | 132.3 | 131.21 | 133.2 | 133.0 | 132.3 |
| 6 | 91.84 | 90.1 | 92.3 | 90.33 | 93.4 | 93.2 | 92.6 |
| 7 | 40.08 | 39.7 | 40.2 | 39.26 | 41.0 | 40.1 | 40.0 |
| 8 | 155.14 | 154.8 | 155.3 | 154.37 | 156.0 | 159.5 | 153.3 |
| 9 | 160.42 | 158.9 | 161.6 | 159.10 | 162.8 | 162.5 | 161.0 |
| 10 | 47.71 | 39.4 | 39.9 | 37.66 | 44.7 | 40.5 | 63.0 |
| 11 | 71.43 | 24.5 | 29.8 | 24.12 | 78.4 | - | 14.4 |
| 12 | 138.5 | 34.5 | 25.6 | 130.79 | 25.7 | - | - |
| 13 | 111.38 | 174.7 | 61.9 | 133.84 | 40.2 | - | - |
| 14 | 130.90 | - | 190.6 | 116.21 | 157.5 | - | - |
| 15 | 150.72 | - | 171.4 | - | - | - | - |
| 16 | 111.38 | - | - | - | - | - | - |
| 17 | 130.90 | - | - | - | - | - | - |
| MeO-C3 | 60.22 | 59.6 | 60.4 | 59.63 | 61.3 | 59.2 | 60.8 |
| Position | 82 | 83 |
|---|---|---|
| 1 | 154.87 | 152.19 |
| 2 | 111.18 | 117.30 |
| 3 | 134.63 | 133.35 |
| 4 | 129.20 | 135.92 |
| 5 | 130.33 | 133.35 |
| 6 | 117.68 | 117.30 |
| 7 | 29.07 | 26.88 |
| 8 | 68.51 | 65.07 |
| OMe | 59.88 | 60.31 |
| (Me)3N+ | 53.77 | 52.23 |
| Position | 90 | 97 | 98 |
|---|---|---|---|
| 1 | 172.6 | 183.0 | 183.0 |
| 2 | 119.9 | 122.7 | 123.7 |
| 3 | 153.2 | 151.7 | 146.6 |
| 4 | 70.8 | 75.5 | 74.2 |
| 5 | 148.8 | 78.4 | 78.9 |
| 6 | 127.6 | 56.1 | 57.1 |
| 1′ | - | 116.9 | 116.9 |
| 2′ | - | 28.4 | 28.4 |
| CH2 | 45.1 | - | - |
| CONH2 | 169.4 | - | - |
| Position | 101 |
|---|---|
| 1 | 86.3 |
| 2 | 109.1 |
| 3 | 149.7 |
| 4 | 113.6 |
| 5 | 140.4 |
| 6 | 76.5 |
| 7 | 42.1 |
| 8 | 172.8 |
| 9 | 60.2 |
| 10 | 60.3 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lira, N.S.; Montes, R.C.; Tavares, J.F.; Silva, M.S.d.; Cunha, E.V.L.d.; Athayde-Filho, P.F.d.; Rodrigues, L.C.; Dias, C.d.S.; Barbosa-Filho, J.M. Brominated Compounds from Marine Sponges of the Genus Aplysina and a Compilation of Their 13C NMR Spectral Data. Mar. Drugs 2011, 9, 2316-2368. https://doi.org/10.3390/md9112316
Lira NS, Montes RC, Tavares JF, Silva MSd, Cunha EVLd, Athayde-Filho PFd, Rodrigues LC, Dias CdS, Barbosa-Filho JM. Brominated Compounds from Marine Sponges of the Genus Aplysina and a Compilation of Their 13C NMR Spectral Data. Marine Drugs. 2011; 9(11):2316-2368. https://doi.org/10.3390/md9112316
Chicago/Turabian StyleLira, Narlize Silva, Ricardo Carneiro Montes, Josean Fechine Tavares, Marcelo Sobral da Silva, Emidio V. L. da Cunha, Petronio Filgueiras de Athayde-Filho, Luis Cezar Rodrigues, Celidarque da Silva Dias, and Jose Maria Barbosa-Filho. 2011. "Brominated Compounds from Marine Sponges of the Genus Aplysina and a Compilation of Their 13C NMR Spectral Data" Marine Drugs 9, no. 11: 2316-2368. https://doi.org/10.3390/md9112316
APA StyleLira, N. S., Montes, R. C., Tavares, J. F., Silva, M. S. d., Cunha, E. V. L. d., Athayde-Filho, P. F. d., Rodrigues, L. C., Dias, C. d. S., & Barbosa-Filho, J. M. (2011). Brominated Compounds from Marine Sponges of the Genus Aplysina and a Compilation of Their 13C NMR Spectral Data. Marine Drugs, 9(11), 2316-2368. https://doi.org/10.3390/md9112316