Di-Ethylhexylphthalate (DEHP) Modulates Cell Invasion, Migration and Anchorage Independent Growth through Targeting S100P in LN-229 Glioblastoma Cells


 and
and 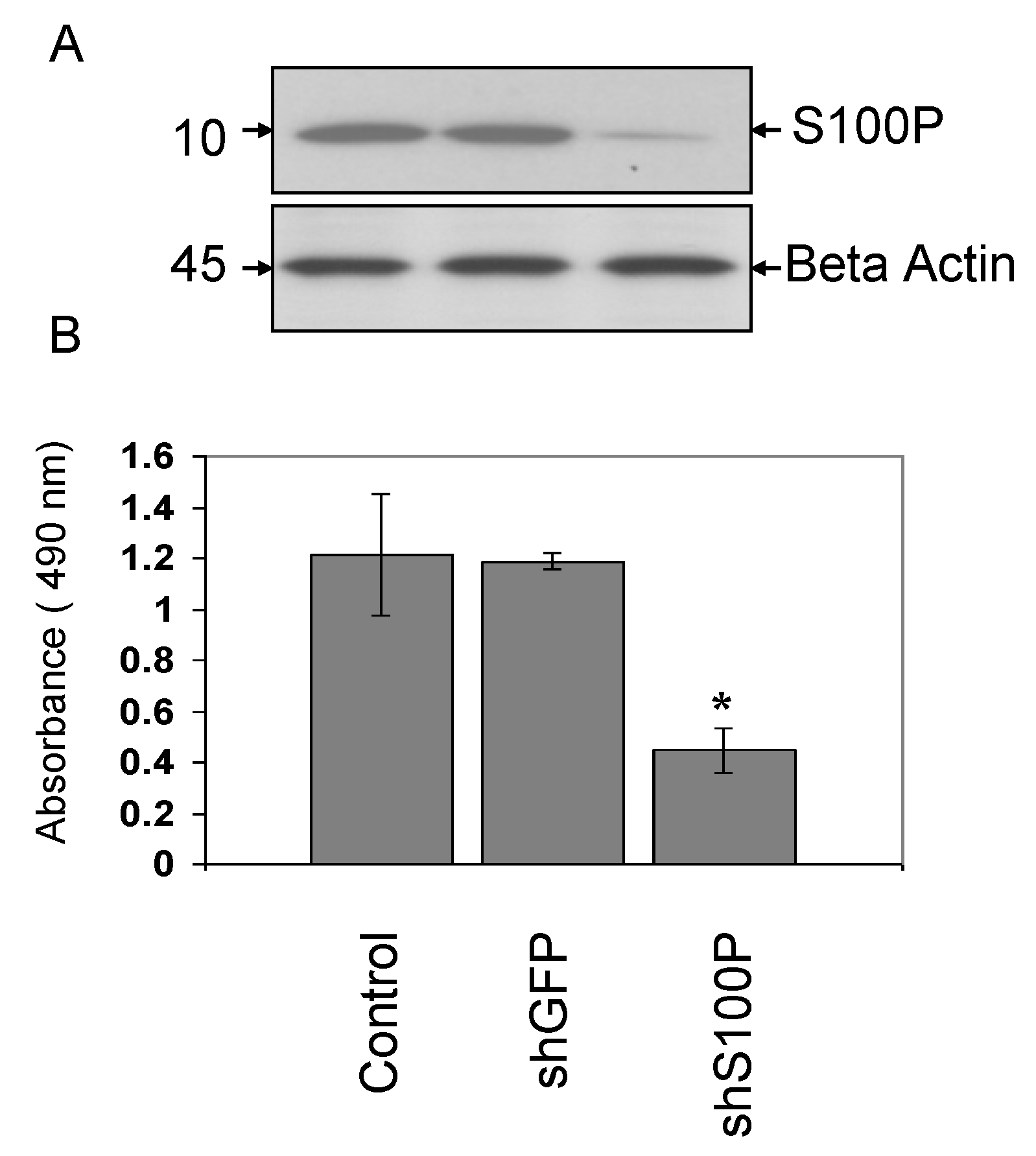{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents
2.2. Cell Culture
2.4. shS100P Infections
2.5. Western Blot Analysis
2.6. Cell Proliferation Assay
2.7. Sub G1 Apoptosis Assay
2.8. Soft Agar Assay
2.9. Wound Scratch Assay
2.10. 3D Spheroid BME Cell Invasion Assay
2.11. Statistical Analyses
3. Results
3.1. Verification of S100P Knockdown in Glioblastoma

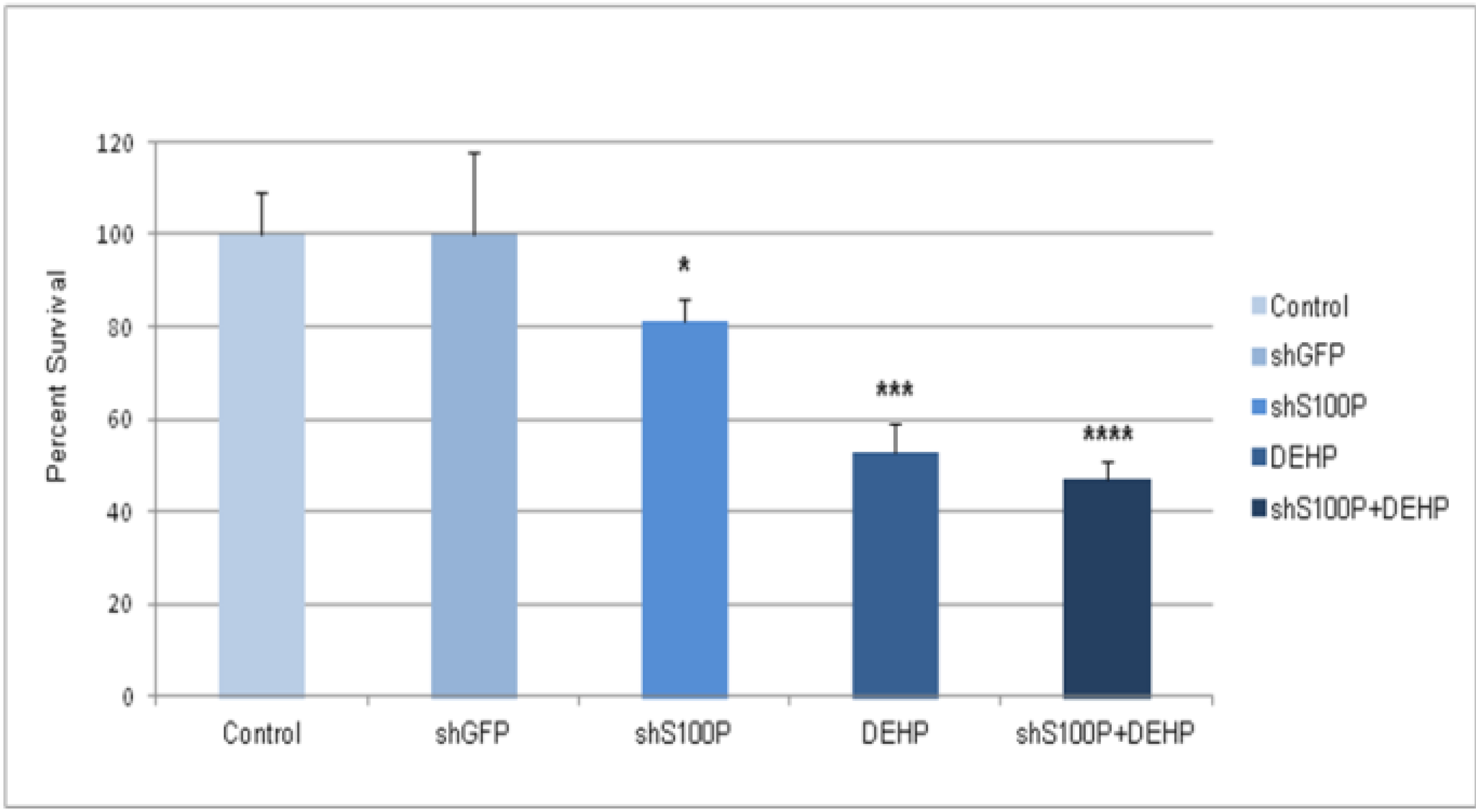
3.2. shS100P and DEHP Exposure Suppressed Glioblastoma Cell Proliferation

3.3. S100P Knockdown Increased Apoptosis in Glioblastoma Cells
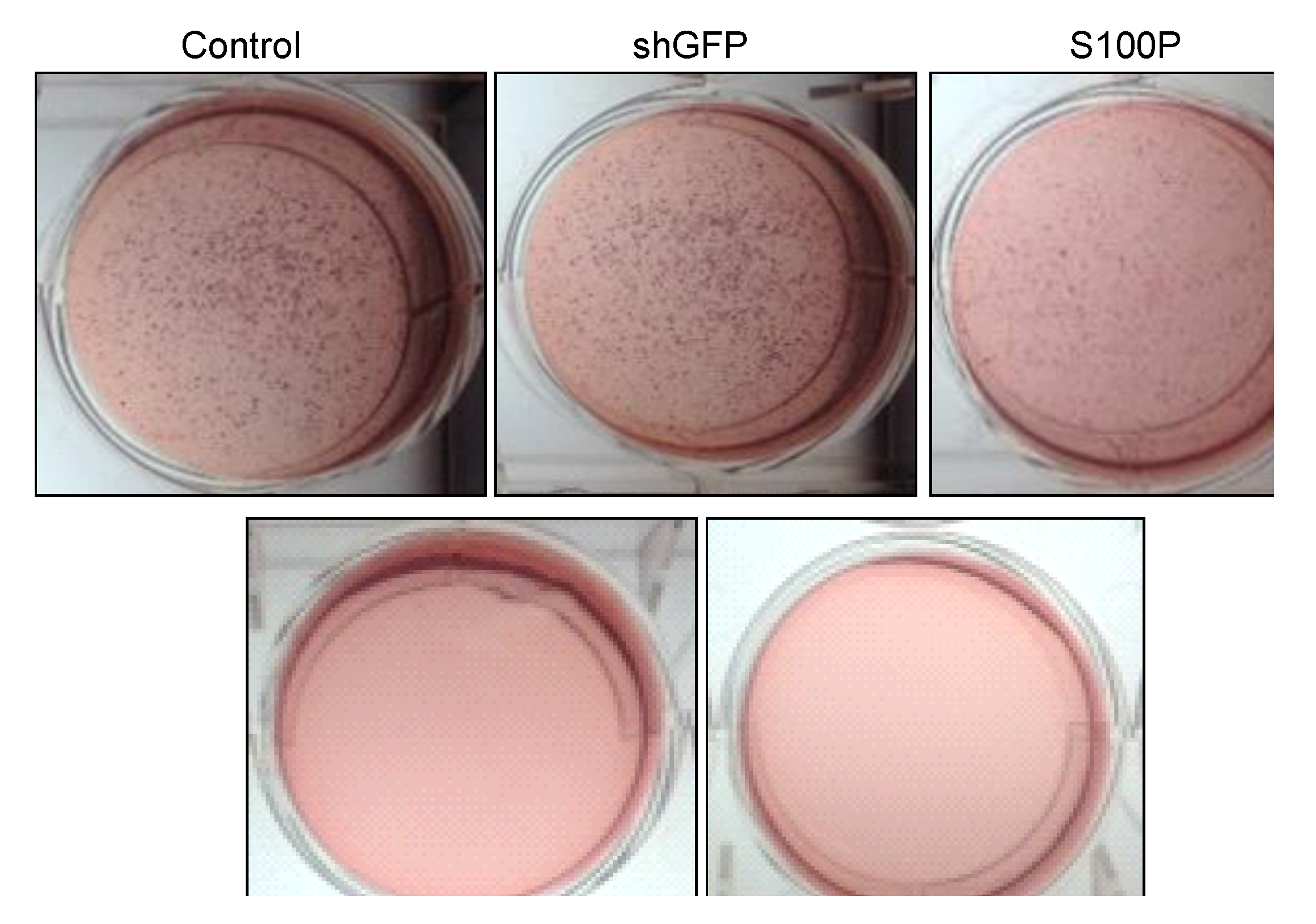
3.4. S100P Knockdown Reduced Anchorage Independent Growth in Glioblastoma Cells
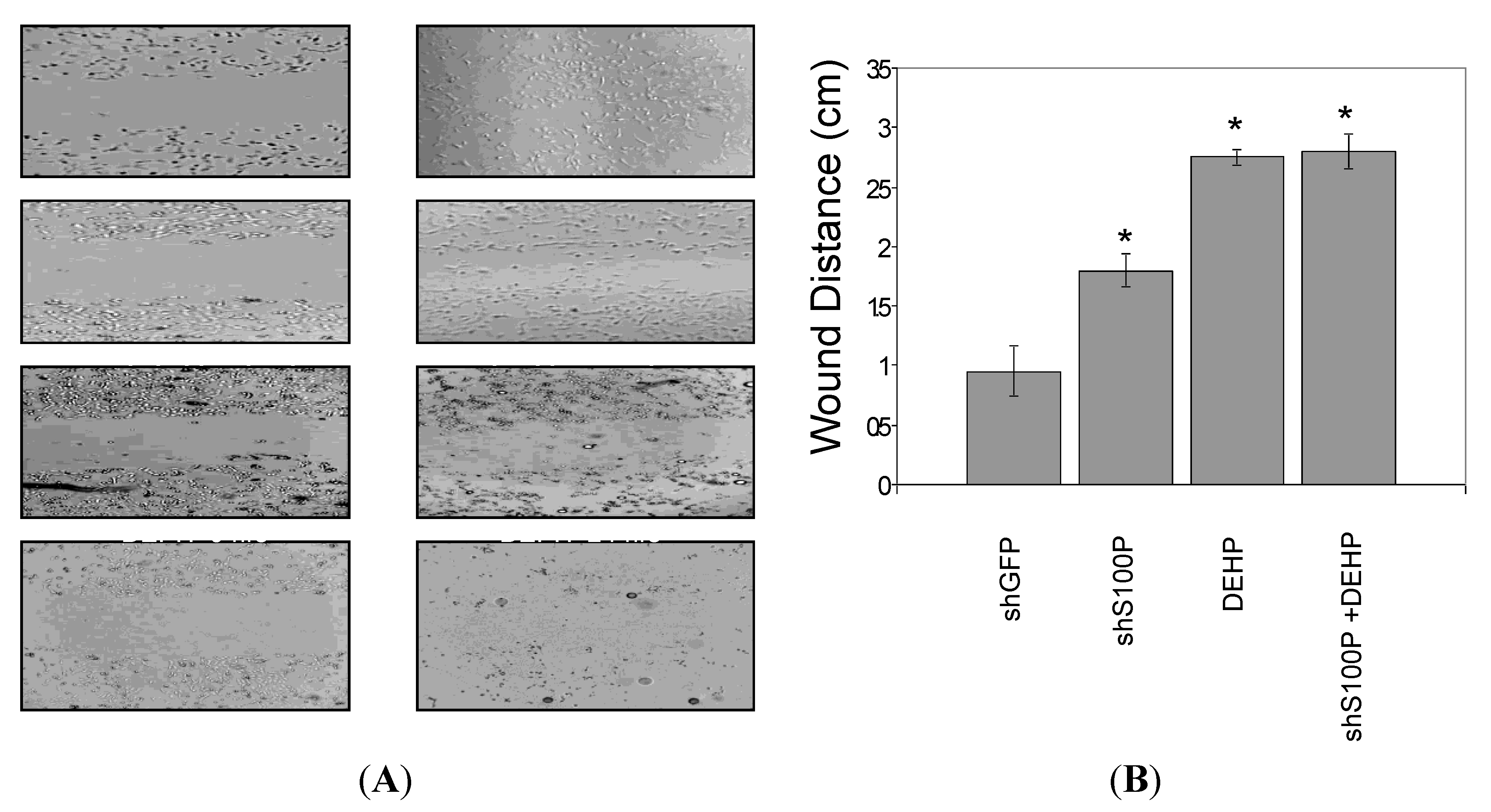
3.5. S100P Knockdown and DEHP Exposure Inhibited Cell Migration



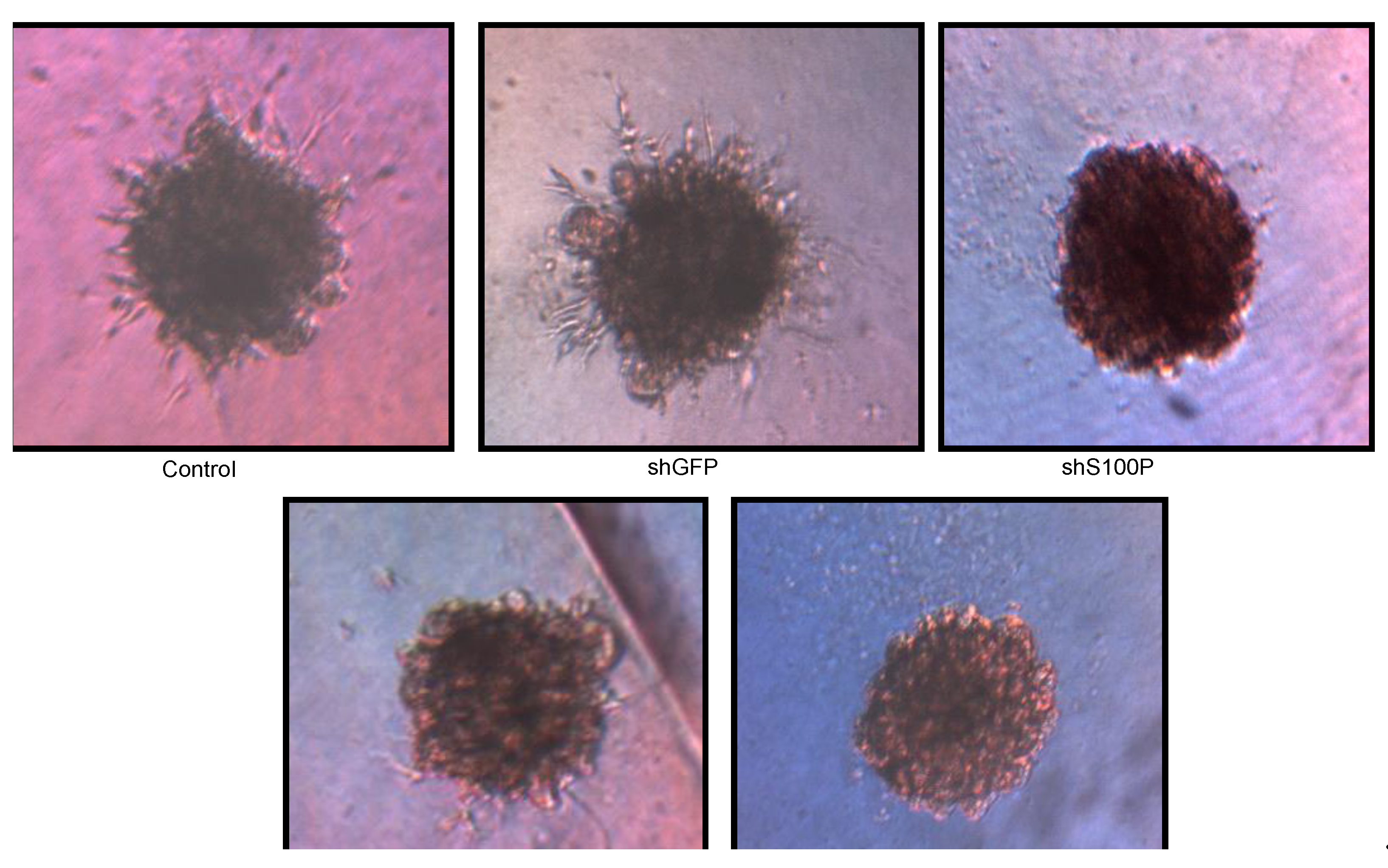
3.6. S100P Knockdown and DEHP Exposure Reduced Spheroid Formation and Expansion
4. Discussion

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Polivka, J., Jr.; Polivka, J.; Rohan, V.; Topolcan, O.; Ferda, J. New molecularly targeted therapies for glioblastoma multiforme. Anticancer Res. 2012, 32, 2935–2946. [Google Scholar]
- Amberger-Murphy, V. Hypoxia helps glioma to fight therapy. Curr. Cancer Drug Targets 2009, 9, 381–390. [Google Scholar] [CrossRef]
- Burton, E.C.; Lamborn, K.R.; Forsyth, P.; Scott, J.; O’Campo, J.; Uyehara-Lock, J.; Prados, M.; Berger, M.; Passe, S.; Uhm, J.; et al. Aberrant p53, mdm2, and proliferation differ in glioblastomas from long-term compared with typical survivors. Clin. Cancer Res. 2002, 8, 180–187. [Google Scholar]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef]
- Clapp, R.W.; Jacobs, M.M.; Loechler, E.L. Environmental and occupational causes of cancer: New evidence 2005–2007. Rev. Environ. Health 2008, 23, 1–37. [Google Scholar] [CrossRef]
- Lyche, J.L.; Gutleb, A.C.; Bergman, A.; Eriksen, G.S.; Murk, A.J.; Ropstad, E.; Saunders, M.; Skaare, J.U. Reproductive and developmental toxicity of phthalates. J. Toxicol. Environ. Health Pt. B 2009, 12, 225–249. [Google Scholar] [CrossRef]
- Casals-Casas, C.; Feige, J.N.; Desvergne, B. Interference of pollutants with PPARs: Endocrine disruption meets metabolism. Int J. Obesity 2008, 32, S53–S61. [Google Scholar] [CrossRef]
- Li, L.; Zhang, T.; Qin, X.S.; Ge, W.; Ma, H.G.; Sun, L.L.; Hou, Z.M.; Chen, H.; Chen, P.; Qin, G.Q.; et al. Exposure to diethylhexyl phthalate (DEHP) results in a heritable modification of imprint genes DNA methylation in mouse oocytes. Mol. Biol Rep. 2014, 41, 1227–1235. [Google Scholar] [CrossRef]
- Becker, T.; Gerke, V.; Kube, E.; Weber, K. S100P, a novel Ca(2+)-binding protein from human placenta. cDNA cloning, recombinant protein expression and Ca2+ binding properties. Eur. J. Biochem. 1992, 207, 541–547. [Google Scholar] [CrossRef]
- Schafer, B.W.; Wicki, R.; Engelkamp, D.; Mattei, M.G.; Heizmann, C.W. Isolation of a YAC clone covering a cluster of nine S100 genes on human chromosome 1q21: Rationale for a new nomenclature of the S100 calcium-binding protein family. Genomics 1995, 25, 638–643. [Google Scholar] [CrossRef]
- Norremolle, A.; Budtz-Jorgensen, E.; Fenger, K.; Nielsen, J.E.; Sorensen, S.A.; Hasholt, L. 4p16.3 haplotype modifying age at onset of Huntington disease. Clin. Genet. 2009, 75, 244–250. [Google Scholar] [CrossRef]
- Ingersoll, R.G.; Hetmanski, J.; Park, J.W.; Fallin, M.D.; McIntosh, I.; Wu-Chou, Y.H.; Chen, P.K.; Yeow, V.; Chong, S.S.; Cheah, F.; et al. Association between genes on chromosome 4p16 and non-syndromic oral clefts in four populations. Eur. J. Hum. Genet. 2010, 18, 726–732. [Google Scholar] [CrossRef]
- Zollino, M.; Murdolo, M.; Neri, G. The terminal 760 kb region on 4p16 is unlikely to be the critical interval for growth delay in Wolf-Hirschhorn syndrome. J. Med. Genet. 2008, 45. [Google Scholar] [CrossRef]
- Zenteno, J.C.; Ruiz, G.; Perez-Cano, H.J.; Camargo, M. Familial Wolfram syndrome due to compound heterozygosity for two novel WFS1 mutations. Mol. Vis. 2008, 14, 1353–1357. [Google Scholar]
- Raelson, J.V.; Little, R.D.; Ruether, A.; Fournier, H.; Paquin, B.; van Eerdewegh, P.; Bradley, W.E.; Croteau, P.; Nguyen-Huu, Q.; Segal, J.; et al. Genome-wide association study for Crohn’s disease in the Quebec Founder Population identifies multiple validated disease loci. Proc. Natl. Acad. Sci. USA 2007, 104, 14747–14752. [Google Scholar] [CrossRef]
- Singh, R.K.; Indra, D.; Mitra, S.; Mondal, R.K.; Basu, P.S.; Roy, A.; Roychowdhury, S.; Panda, C.K. Deletions in chromosome 4 differentially associated with the development of cervical cancer: Evidence of slit2 as a candidate tumor suppressor gene. Hum. Genet. 2007, 122, 71–81. [Google Scholar] [CrossRef]
- Yuan, R.H.; Chang, K.T.; Chen, Y.L.; Hsu, H.C.; Lee, P.H.; Lai, P.L.; Jeng, Y.M. S100P expression is a novel prognostic factor in hepatocellular carcinoma and predicts survival in patients with high tumor stage or early recurrent tumors. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Da Silva, I.D.G.; Hu, Y.F.; Russo, I.H.; Ao, X.; Salicioni, A.M.; Yang, X.; Russo, J. S100P calcium-binding protein overexpression is associated with immortalization of human breast epithelial cells in vitro and early stages of breast cancer development in vivo. Int. J. Oncol. 2000, 16, 231–240. [Google Scholar]
- Kita, H.; Hikichi, Y.; Hikami, K.; Tsuneyama, K.; Cui, Z.G.; Osawa, H.; Ohnishi, H.; Mutoh, H.; Hoshino, H.; Bowlus, C.L.; et al. Differential gene expression between flat adenoma and normal mucosa in the colon in a microarray analysis. J. Gastroenterol. 2006, 41, 1053–1063. [Google Scholar] [CrossRef]
- Fuentes, M.K.; Nigavekar, S.S.; Arumugam, T.; Logsdon, C.D.; Schmidt, A.M.; Park, J.C.; Huang, E.H. RAGE activation by S100P in colon cancer stimulates growth, migration, and cell signaling pathways. Dis. Colon Rectum 2007, 50, 1230–1240. [Google Scholar] [CrossRef]
- Jiang, H.; Hu, H.; Tong, X.; Jiang, Q.; Zhu, H.; Zhang, S. Calcium-binding protein S100P and cancer: Mechanisms and clinical relevance. J. Cancer Res. Clin. Oncol. 2012, 138, 1–9. [Google Scholar]
- Johnston, J.C.; Gasmi, M.; Lim, L.E.; Elder, J.H.; Yee, J.K.; Jolly, D.J.; Campbell, K.P.; Davidson, B.L.; Sauter, S.L. Minimum requirements for efficient transduction of dividing and nondividing cells by feline immunodeficiency virus vectors. J. Virol. 1999, 73, 4991–5000. [Google Scholar]
- Arumugam, T.; Simeone, D.M.; van Golen, K.; Logsdon, C.D. S100P promotes pancreatic cancer growth, survival, and invasion. Clin. Cancer Res. 2005, 11, 5356–5364. [Google Scholar] [CrossRef]
- Basu, G.D.; Azorsa, D.O.; Kiefer, J.A.; Rojas, A.M.; Tuzmen, S.; Barrett, M.T.; Trent, J.M.; Kallioniemi, O.; Mousses, S. Functional evidence implicating S100P in prostate cancer progression. Int. J. Cancer 2008, 123, 330–339. [Google Scholar] [CrossRef]
- Van Meir, E.G.; Kikuchi, T.; Tada, M.; Li, H.; Diserens, A.C.; Wojcik, B.E; Cavenee, W.K. Analysis of the p53 gene and its expression in human glioblastoma cells. Cancer Res. 1994, 54, 649–652. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sims, J.N.; Graham, B.; Pacurari, M.; Leggett, S.S.; Tchounwou, P.B.; Ndebele, K. Di-Ethylhexylphthalate (DEHP) Modulates Cell Invasion, Migration and Anchorage Independent Growth through Targeting S100P in LN-229 Glioblastoma Cells. Int. J. Environ. Res. Public Health 2014, 11, 5006-5019. https://doi.org/10.3390/ijerph110505006
Sims JN, Graham B, Pacurari M, Leggett SS, Tchounwou PB, Ndebele K. Di-Ethylhexylphthalate (DEHP) Modulates Cell Invasion, Migration and Anchorage Independent Growth through Targeting S100P in LN-229 Glioblastoma Cells. International Journal of Environmental Research and Public Health. 2014; 11(5):5006-5019. https://doi.org/10.3390/ijerph110505006
Chicago/Turabian StyleSims, Jennifer Nicole, Barbara Graham, Maricica Pacurari, Sophia S. Leggett, Paul B. Tchounwou, and Kenneth Ndebele. 2014. "Di-Ethylhexylphthalate (DEHP) Modulates Cell Invasion, Migration and Anchorage Independent Growth through Targeting S100P in LN-229 Glioblastoma Cells" International Journal of Environmental Research and Public Health 11, no. 5: 5006-5019. https://doi.org/10.3390/ijerph110505006
APA StyleSims, J. N., Graham, B., Pacurari, M., Leggett, S. S., Tchounwou, P. B., & Ndebele, K. (2014). Di-Ethylhexylphthalate (DEHP) Modulates Cell Invasion, Migration and Anchorage Independent Growth through Targeting S100P in LN-229 Glioblastoma Cells. International Journal of Environmental Research and Public Health, 11(5), 5006-5019. https://doi.org/10.3390/ijerph110505006