Occurrence of Vibrio Pathotypes in the Final Effluents of Five Wastewater Treatment Plants in Amathole and Chris Hani District Municipalities in South Africa


Abstract
:1. Introduction
2. Experimental Section
2.1. Description of Study Site

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WWTP | Amathole D.M WWTPs | Chris Hani D.M WWTPs | |||
|---|---|---|---|---|---|
| Plant M | Plant R | Plant E | Plant W | Plant Q | |
| Technology | Biofilters, anaerobic digestion and sludge drying beds | Activated sludge and sludge lagoons | Activated sludge and marine outfall | Biofilters, sludge composting | Biofilters, anaerobic digestion |
| Design Capacity (ML/d) | 24 | 2.5 | 40 | 4.99 | NI * |
| Operational % in relation to Design Capacity | 43.8% | 44% | 85.5% | 50.1% | NI * |
2.2. Sample Collection
2.3. Enumeration and Isolation of Presumptive Vibrio Species
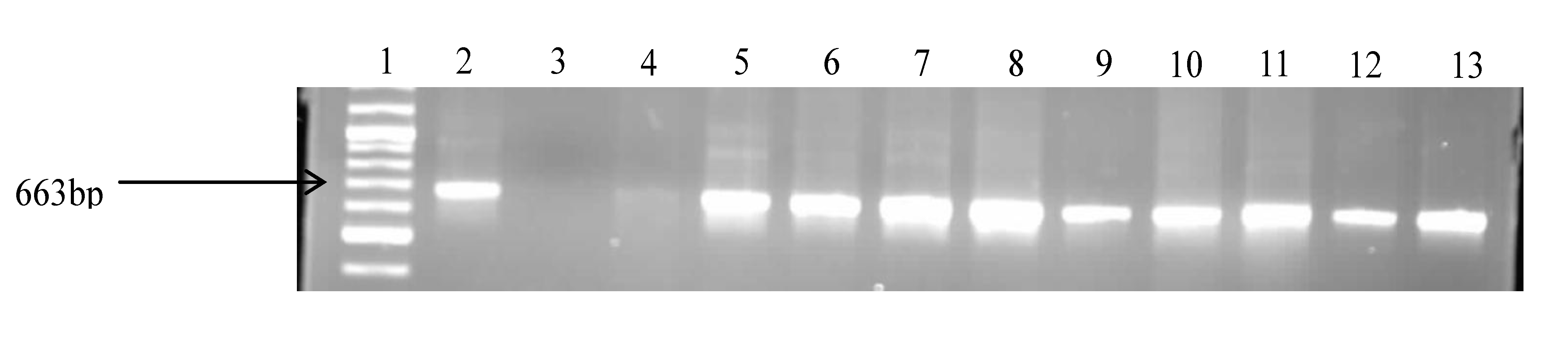
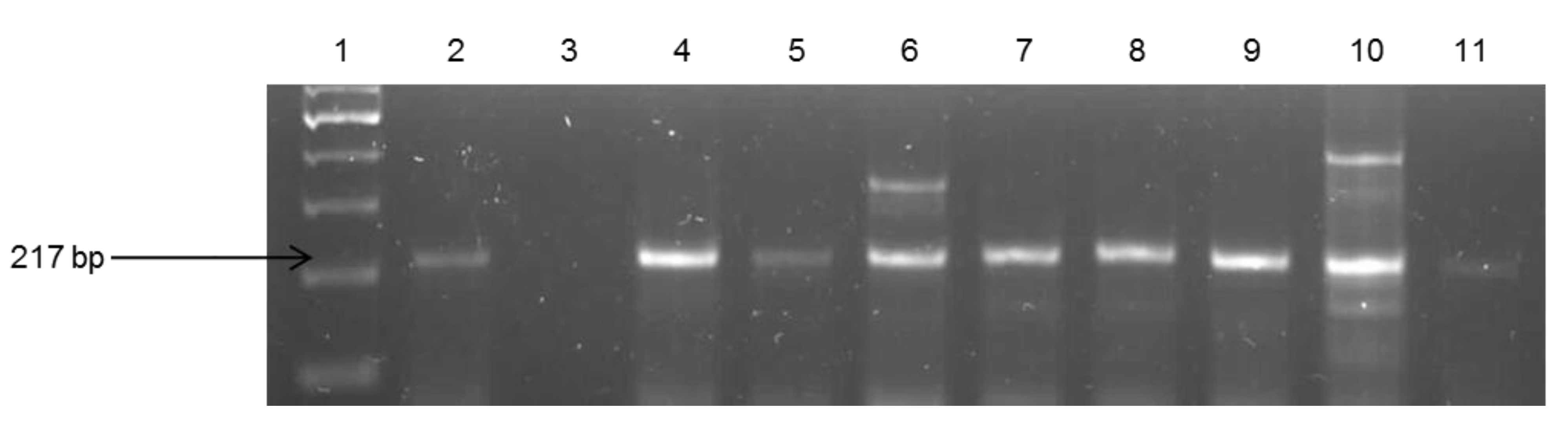
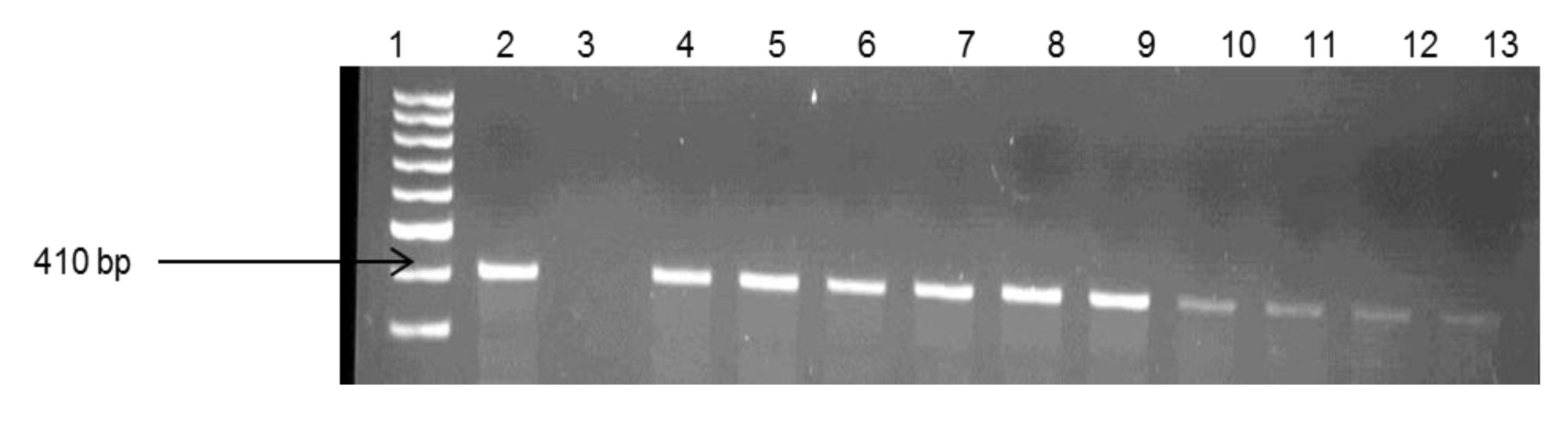
2.4. Molecular Confirmation of Pathogenic Vibrio Species
| Target Species | Primers | Sequences (5’3’) | Target Gene | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| All Vibrio spp. | V. 16S-700F V. 16s-1325R | CGG TGA AAT GCG TAG AGA T TTA CTA GCG ATT CCG AGT TC | 16SrRNA | 663 | [27] |
| V. parahaemolyticus | Vp.toxR R Vp.toxR F | GTC TTC TGA CGC AAT CGT TG ATA CGA GTG GTT GCT GTC ATG | toxR | 368 | [29] |
| V. vulnificus | Vv. hsp-326F Vv. hsp-697R | GTC TTA AAG CGG TTG CTG C CGC TTC AAG TGC TGG TAG AAG | hsp60 | 410 | [30] |
| V. fluvialis | Vf- toxR F Vf- toxR R | GAC CAG GGC TTT GAG GTG GAC AGG ATA CGG CAC TTG AGT AAG ACT C | toxR | 217 | [31] |
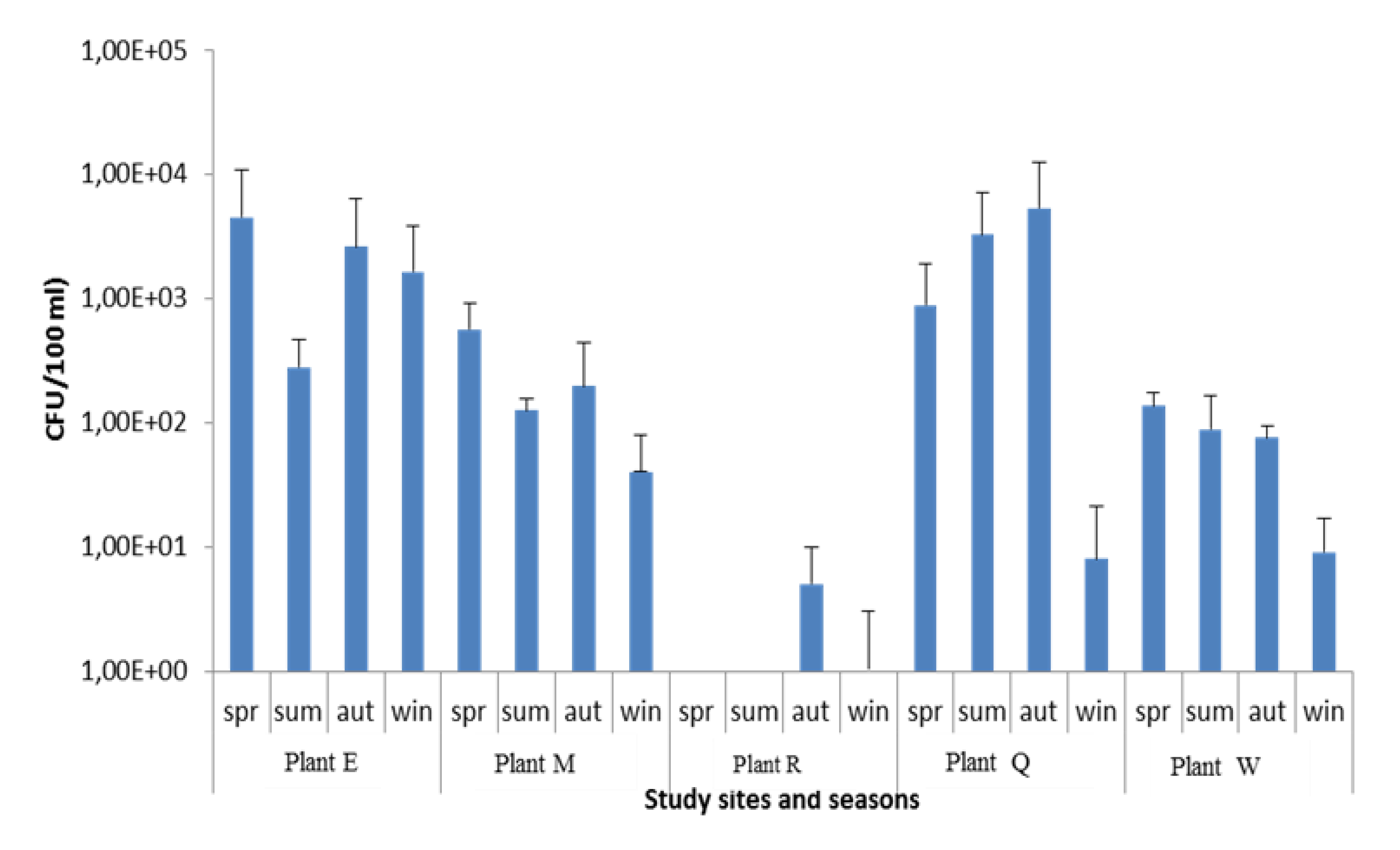
3. Results and Discussion
| Vibrio spp. (CFU/100 mL) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WWTP | SEPT ’12 | OCT ’12 | NOV ’12 | DEC ‘12 | JAN’13 | FEB’13 | MAR’13 | APR’13 | MAY’13 | JUNE ’13 | JULY’13 | AUG’13 |
| PLANT M | 9.2 × 102 | 4.8 × 101 | 6.2 × 102 | 1.6 × 102 | 1.0 × 102 | 2.6 × 101 | 5.2 × 102 | 6.0 × 101 | 1.3 × 101 | 1.4 × 101 | 9.5 × 101 | 1.3 × 101 |
| PLANT R | <1 | <1 | 1.3 × 100 | 2 × 100 | 1 × 100 | <1 | <1 | 8.7 × 100 | 9 × 10° | <1 | 5 × 100 | <1 |
| PLANT E | 3.6 × 101 | 8.5 × 102 | 1.28 × 104 | 6.1 × 102 | 5.2 × 102 | 3.5 × 101 | 6.2 × 101 | 7.7 × 103 | 6.0 × 101 | 3.5 × 102 | 4.5 × 103 | 1.29 × 103 |
| PLANT W | N/S | 1.14 × 102 | 1.61 × 102 | 1.8 × 102 | 4.0 × 101 | 3.9 × 101 | 7.2 × 101 | 1.6 × 101 | 2.1 × 101 | < 1 | 1.6 × 101 | 7 × 100 |
| PLANT Q | 6.8 × 101 | 3.4 × 102 | 2.26 × 103 | 1.48 × 103 | 9.6 × 101 | 8.3 × 103 | 1.24 × 103 | 5.8 × 101 | 1.48 × 104 | 2.5 × 101 | 1 × 100 | <1 |





4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Department of Water Affairs and Forestry, (DWAF). South African water quality guidelines. In Domestic Uses, Department of Water Affairs and Forestry; Holmes, S., Ed.; DWAF: Pretoria, South Africa, 1996; pp. 132–135. Available online: http://www.dwaf.gov.za/iwqs/wq_guide/Pol_saWQguideFRESH_vol6_AquaCulture.pdf (accessed on 17 November 2013). [Google Scholar]
- Bahlaoui, M.A.; Baleux, B.; Troussellier, M. Dynamics of pollution indicators and pathogenic bacteria in high rate oxidation wastewater treatment ponds. Water Res. 1997, 31, 574–582. [Google Scholar] [CrossRef]
- Momba, M.N.B.; Osode, A.N.; Sibewu, M. The impact of inadequate wastewater treatment on the receiving water bodies—Case study: Buffalo City and Nkonkobe Municipalities of the Eastern Cape Province. Water SA. 2006, 32. Available online: http://www.ewisa.co.za/literature/files/275%20Momba.pdf (accessed on 3 June 2014).
- Shuval, H.; Badri, F. Handbook of Water and Wastewater Microbiology, 1st ed.; Academic Press: San Diego, CA, USA, 2003; pp. 241–262. [Google Scholar]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Griffith, D.C.; Kelly-Hope, L.A.; Miller, M.A. Review of reported cholera outbreaks worldwide, 1995–2005. Am. J. Trop. Med. Hyg. 2006, 75, 973–977. [Google Scholar]
- Daniels, N.A.; MacKinnon, L.; Bishop, R.; Altekruse, S.; Ray, B.; Hammond, R.M.; Slutsker, L. Vibrio parahaemolyticus infections in the United States, 1973–1998. J. Infect. Dis. 2002, 181, 1661–1666. [Google Scholar]
- Thompson, J.R.; Randa, M.A.; Marcelino, L.A.; Tomita-Mitchell, A.; Lim, E.; Polz, M.F. Diversity and dynamics of a North Atlantic coastal Vibrio community. Appl. Environ. Microbiol. 2004, 70, 4103–4110. [Google Scholar] [CrossRef]
- Cavallo, R.N.; Loredana, S. Culturable vibrios biodiversity in the Northern Ionian Sea (Italian coasts). Sci. Mar. 2004, 68, 23–29. [Google Scholar]
- Mezrioui, N.; Oufdou, K. Abundance and antibiotic resistance of non-O1 Vibrio cholerae strains in domestic wastewater before and after treatment in stabilization ponds in an arid region (Marrakesh, Morocco). FEMS Microbiol. Ecol. 1996, 21, 277–284. [Google Scholar]
- Mishra, M.; Mohammed, F.; Akulwar, S.L.; Katkar, V.J.; Tankhiwale, N.S.; Powar, R.M. Re-emergence of El Tor Vibrio in outbreak of cholera in and around Nagpur, Indian. J. Med. Res. 2004, 120, 478–480. [Google Scholar]
- Tantillo, G.M.; Fontanarosa, M.; Di Pinto, A.; Musti, M. Updated perspectives on emerging vibrios associated with human infections. Lett. Appl. Microbiol. 2004, 39, 117–126. [Google Scholar] [CrossRef]
- Bross, M.H.; Soch, K.; Morales, R.; Mitchell, R.B. Vibrio vulnificus infection: Diagnosis and treatment. Am. Fam. Physician 2007, 76, 539–544. [Google Scholar]
- Anuradha De, M.M. Vibrio vulnificus diarrhea in a child with respiratory infection. J. Glob. Infect. Dis. 2011, 3, 300–302. [Google Scholar]
- Morris, J.G. Vibrio vulnificus—A new monster of the deep? Ann. Intern. Med. 1988, 109, 261–263. [Google Scholar] [CrossRef]
- McCarter, L. The multiple identities of Vibrio parahaemolyticus. J. Mol. Microbiol. Biotechnol. 1991, 1, 51–57. [Google Scholar]
- Nair, G.B.; Ramamurthy, T.; Bhattacharya, S.K.; Dutta, B.; Takeda, Y.; Sack, D.A. Global dissemination of Vibrio parahaemolyticus serotype O3: K6 and its serovariants. Clin. Microbiol. Rev. 2007, 20, 39–48. [Google Scholar] [CrossRef]
- Tunung, R.; Ghazali, F.M.; Noranizan, M.A.; Haresh, K.K.; Lesley, M.B.; Nakaguchi, Y.; Son, R. Rapid detection and enumeration of pathogenic Vibrio parahaemolyticus in raw vegetables from retail outlets. Int. Food Res. J. 2011, 18, 67–78. [Google Scholar]
- Huq, M.I.; Alam, A.K.; Brenner, D.J.; Morris, G.K. Isolation of Vibrio-like group, EF-6, from patients with diarrhea. J. Clin. Microbiol. 1980, 5, 621–624. [Google Scholar]
- Hlady, W.G.; Klontz, K.C. The epidemiology of Vibrio infections in Florida, 1981–1993. J. Infect. Dis. 1996, 173, 1176–1183. [Google Scholar] [CrossRef]
- Lesmana, M.; Subekti, D.S.; Tjaniadi, P.; Simanjuntak, C.H.; Punjabi, N.H.; Campbell, J.; Rand Oyofo, B.A. Spectrum of Vibrio species associated with acute diarrhea in North Jakarta, Indonesia. Diagn. Microbiol. Infect. Dis. 2002, 43, 91–97. [Google Scholar] [CrossRef]
- Tall, B.D.; Fall, S.; Pereira, M.R.; Ramos-Valle, M.; Curtis, S.K.; Kothary, M.H.; Chu, D.M.; Monday, S.R.; Kornegay, L.; Donkar, T.; et al. Characterization of Vibrio fluvialis-like strains implicated in limp lobster disease. Appl. Environ. Microbiol. 2003, 5, 7435–7446. [Google Scholar]
- Chakraborty, R.; Chakraborty, S.; De, K.; Sinha, S.; Mukhopadhyay, A.K.; Khanam, J.; Ramamurthy, T.; Takeda, Y.; Bhattacharya, S.K.; Nair, G.B. Cytotoxic and cell vacuolating activity of Vibrio fluvialis isolated from paediatric patients with diarrhoea. Int. J. Med. Microbiol. 2005, 8, 707–716. [Google Scholar]
- Igbinosa, E.O.; Obi, C.L.; Okoh, A.I. Occurrence of potentially pathogenic vibrios in the final effluents of a wastewater treatment facility in a rural community of the Eastern Cape Province of South Africa. Res. Microbiol. 2009, 160, 531–537. [Google Scholar] [CrossRef]
- Igbinosa, E.O.; Okoh, A.I. Vibrio fluvialis: An unusual enteric pathogen of increasing public health concern. Int. J. Environ. Res. Public Health 2010, 7, 3628–3643. [Google Scholar] [CrossRef]
- Mafuya, P.N.; Shukla, N. Factors that could motivate people to adopt safe hygienic practices in the Eastern Cape Province, South Africa. Afr. Health Sci. 2005, 5, 21–28. [Google Scholar]
- Kwok, A.Y.; Wilson, J.T.; Coulthart, M.; Ng, L.K.; Mutharia, L.; Chow, A.W. Phylogenetic study and identification of human pathogenic Vibrio species based on partial hsp60 gene sequences. Can. J. Micriobiol. 2002, 48, 903–910. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Carbone, M.; Fera, M.T.; Gugliandolo, C. Detection and differentiation of Vibrio vulnificus in seawater and plankton of coastal zone of the Mediterranean Sea. Res. Microbiol. 2006, 157, 194–200. [Google Scholar] [CrossRef]
- Kim, Y.B.; Okuda, J.; Matsumoto, C.; Takahashi, N.; Hashimoto, S.; Nishibichi, M. Identification of Vibrio parahaemolyticus strains at the species level by PCR targeted to the toxR gene. J. Clin. Microbiol. 1997, 37, 1173–1177. [Google Scholar]
- Wong, R.S.; Chow, A.W. Identification of enteric pathogens by heat shock protein60kDa (HSP60) gene sequesnces. FEMS Microbiol. Lett. 2002, 206, 107–113. [Google Scholar] [CrossRef]
- Chakraborty, R.; Sinha, S.; Mukhopadhyay, A.K.; Asakura, M.; Yamasaki, S.; Bhattacharya, S.K.; Nair, G.; Ramamurthy, T. Species-specific identification of Vibrio fluvialis by PCR targeted to the conserved transcriptional activation and variable membrane tether regions of the toxR gene. J. Med. Microbiol. 2006, 55, 805–808. [Google Scholar] [CrossRef]
- Uddin, M.A.; Ullah, M.W.; Noor, R. Prevalence of Vibrio cholerae in human, poultry, animal excreta and compost samples. Stamford J. Microbiol. 2013, 2, 38–41. [Google Scholar]
- Lin, M.; Schwarz, J.R. Seasonal shifts in population structure of Vibrio vulnificus in an estuarine environment as revealed by partial 16S ribosomal DNA sequencing. FEMS Microbiol. Ecol. 2003, 45, 23–27. [Google Scholar]
- Maugeri, T.L.; Carbone, M.; Fera, M.T.; Irrera, G.P.; Gugliandolo, C. Distribution of potentially pathogenic bacteria as free living and plankton associated in a marine coastal zone. J. Appl. Microbiol. 2004, 97, 354–361. [Google Scholar]
- Gugliandolo, C.; Carbone, M.; Fera, M.T.; Irrera, G.P.; Maugeri, T.L. Occurrence of potentially pathogenic vibrios in the marine environment of the Straits of Messina (Italy). Mar. Pollut. Bull. 2005, 50, 692–697. [Google Scholar] [CrossRef]
- Sousa, O.V.D.; Vieira, R.H.S.D.F.; Menezes, F.G.R.D.; Reis, C.M.F.D.; Hofer, E. Detection of Vibrio parahaemolyticus and Vibrio cholerae in oyster, Crassostrea rhizophorae, collected from a natural nursery in the Cocó river estuary, Fortaleza, Ceará, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2004, 46, 59–62. [Google Scholar] [CrossRef]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Vasta, G.R. Emerging marine diseases—Climate links and anthropogenic factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef]
- Kelly, M.T.; Dan Stroh, E.M. Occurrence of Vibrionaceae in natural and cultivated oyster populations in the Pacific Northwest. Diagn. Microbiol. Infect. Dis. 1988, 9, 1–5. [Google Scholar]
- Liang, P.; Cui, X.; Du, X.; Kan, B.; Liang, W. The virulence phenotypes and molecular epidemiological characteristics of Vibrio fluvialis in China. Gut 2013, 5. [Google Scholar] [CrossRef]
- Daniels, N.A.; Shafaie, A. A review of pathogenic Vibrio infections for clinician. Infect. Med. 2000, 17, 665–685. [Google Scholar]
- Elhadi, N.; Radu, S.; Chen, C.H.; Nishibuchi, M. Prevalence of potentially pathogenic Vibrio species in the seafood marketed in Malaysia. J. Food Prot. 2004, 67, 1469–1475. [Google Scholar]
- Ji, H.; Chen, Y.; Guo, Y.; Liu, X.; Wen, J.; Liu, H. Occurrence and characteristics of Vibrio vulnificus in retail marine shrimp in China. Food Control 2011, 22, 1935–1940. [Google Scholar] [CrossRef]
- Yano, Y.; Yokoyama, M.; Satomi, M.; Oikawa, H.; Chen, S. Occurrence of Vibrio vulnificus in fish and shellfish available from markets in China. J. Food Prot. 2004, 67, 1617–1623. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nongogo, V.; Okoh, A.I. Occurrence of Vibrio Pathotypes in the Final Effluents of Five Wastewater Treatment Plants in Amathole and Chris Hani District Municipalities in South Africa. Int. J. Environ. Res. Public Health 2014, 11, 7755-7766. https://doi.org/10.3390/ijerph110807755
Nongogo V, Okoh AI. Occurrence of Vibrio Pathotypes in the Final Effluents of Five Wastewater Treatment Plants in Amathole and Chris Hani District Municipalities in South Africa. International Journal of Environmental Research and Public Health. 2014; 11(8):7755-7766. https://doi.org/10.3390/ijerph110807755
Chicago/Turabian StyleNongogo, Vuyokazi, and Anthony I. Okoh. 2014. "Occurrence of Vibrio Pathotypes in the Final Effluents of Five Wastewater Treatment Plants in Amathole and Chris Hani District Municipalities in South Africa" International Journal of Environmental Research and Public Health 11, no. 8: 7755-7766. https://doi.org/10.3390/ijerph110807755
APA StyleNongogo, V., & Okoh, A. I. (2014). Occurrence of Vibrio Pathotypes in the Final Effluents of Five Wastewater Treatment Plants in Amathole and Chris Hani District Municipalities in South Africa. International Journal of Environmental Research and Public Health, 11(8), 7755-7766. https://doi.org/10.3390/ijerph110807755