
Overexpression of RACK1 Promotes Metastasis by Enhancing Epithelial-Mesenchymal Transition and Predicts Poor Prognosis in Human Glioma

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Follow-Up
2.2. Cell Culture and siRNA Interference
2.3. Cell Proliferation Assays
2.4. Colony Formation Assay
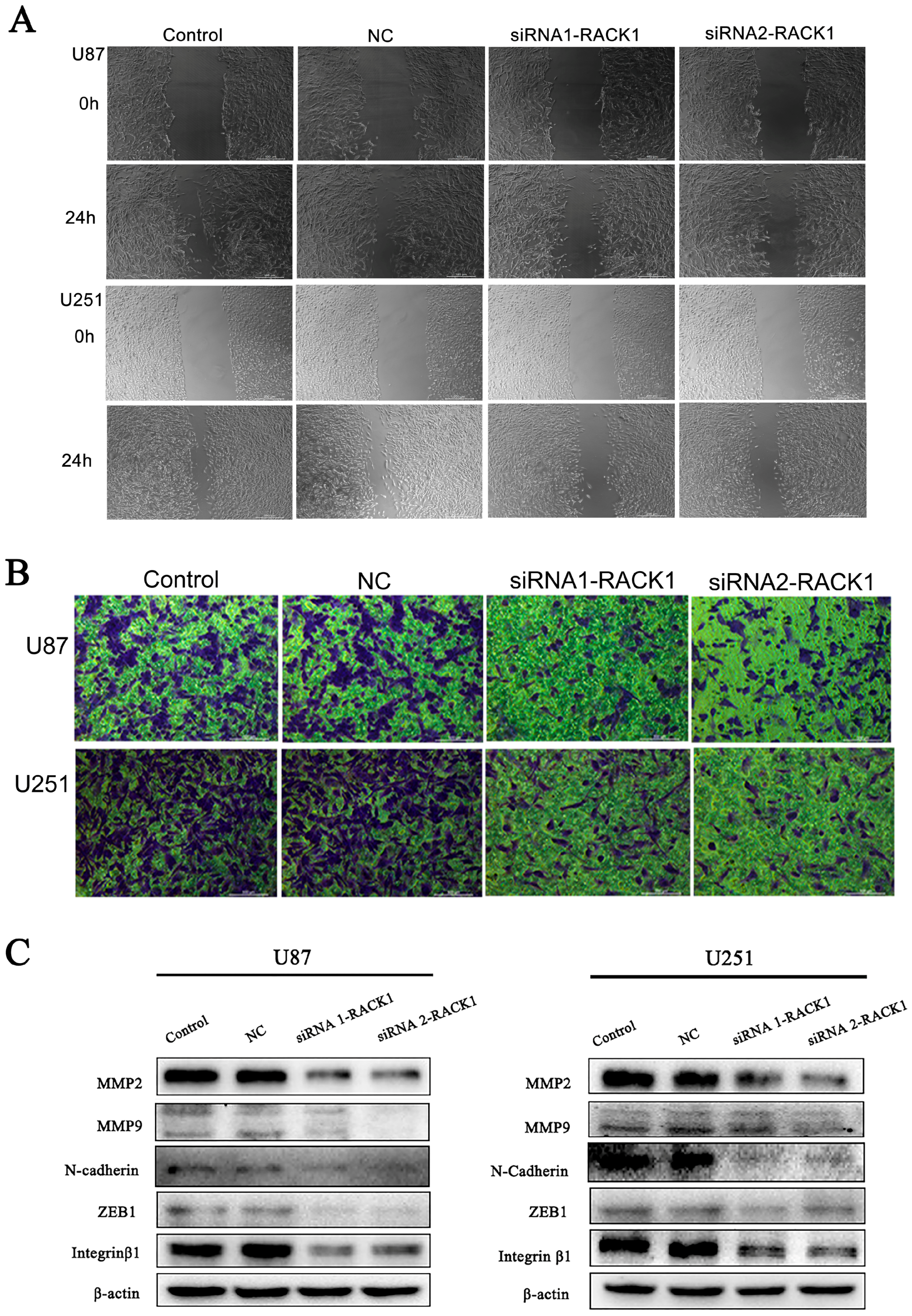
2.5. Wound Healing Assay
2.6. Transwell Invasion Assay
2.7. Real-Time PCR Analysis
2.8. Western Blot Analysis
2.9. Immunohistochemistry (IHC)
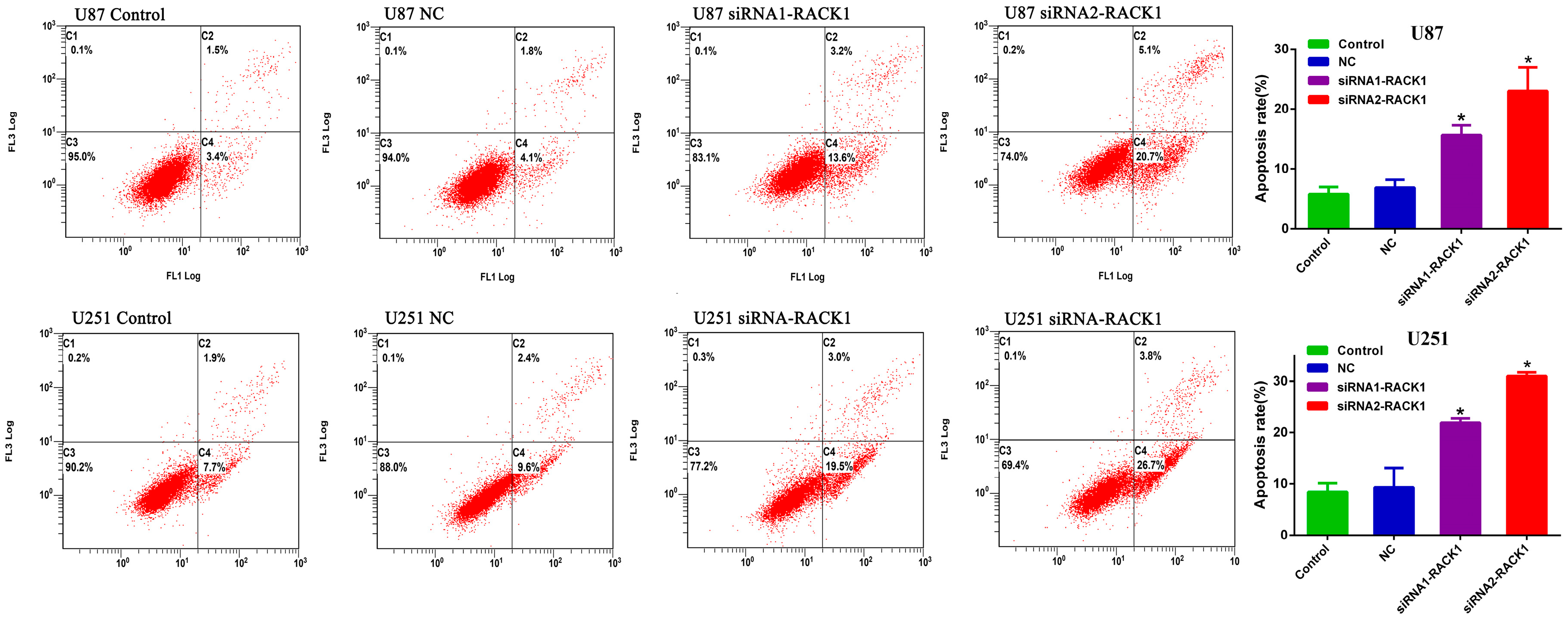
2.10. Flow Cytometry Analysis
2.11. Statistical Analysis
3. Results
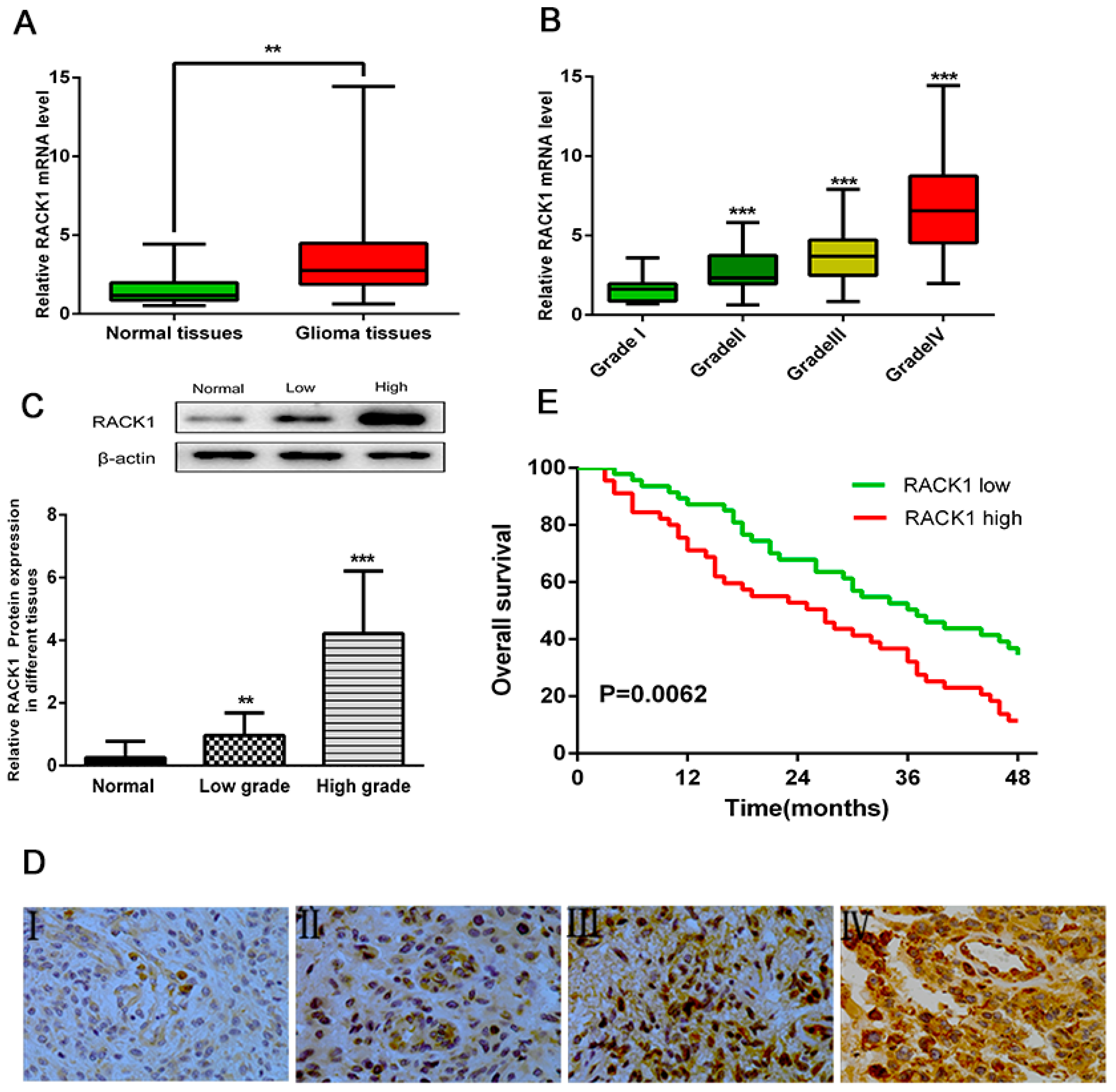
3.1. Overexpression of RACK1 Correlates with Histological Malignancy and Poor Prognosis in Human Gliomas
3.2. Upregulation of RACK1 Is Associated with Poor Prognosis of Glioma Patients
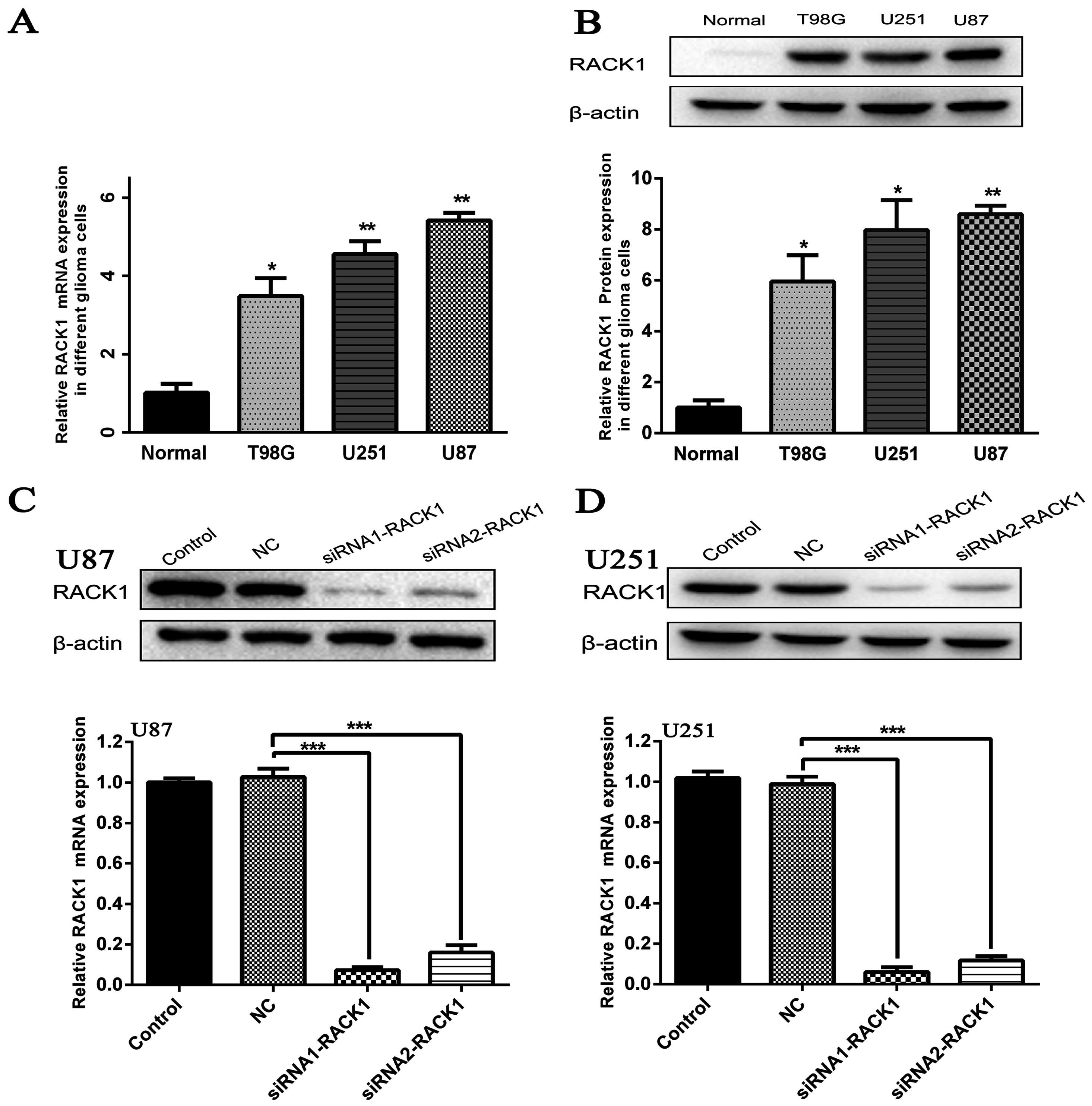
3.3. Silencing of RACK1 Expression in Glioma Cells by siRNA
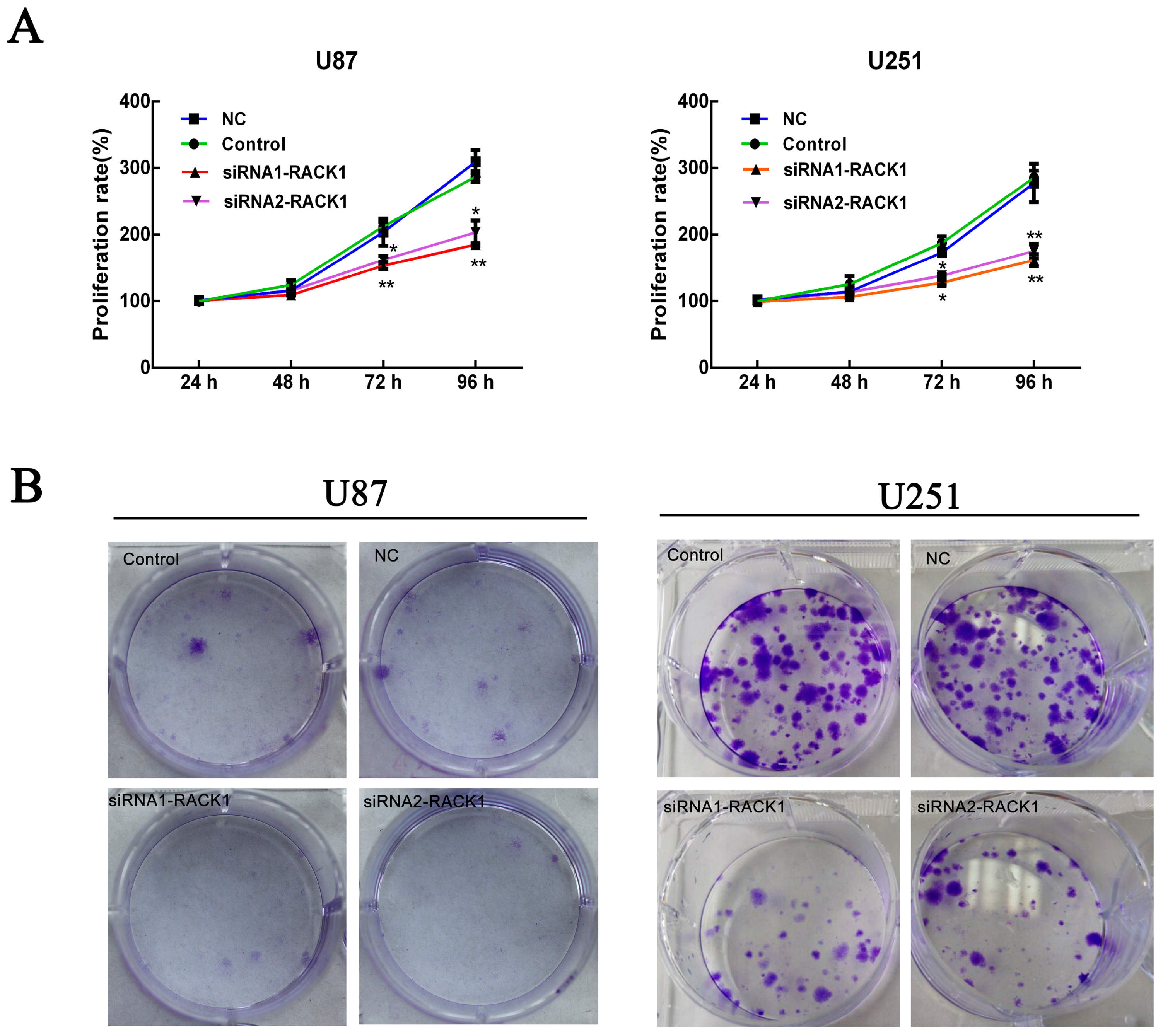
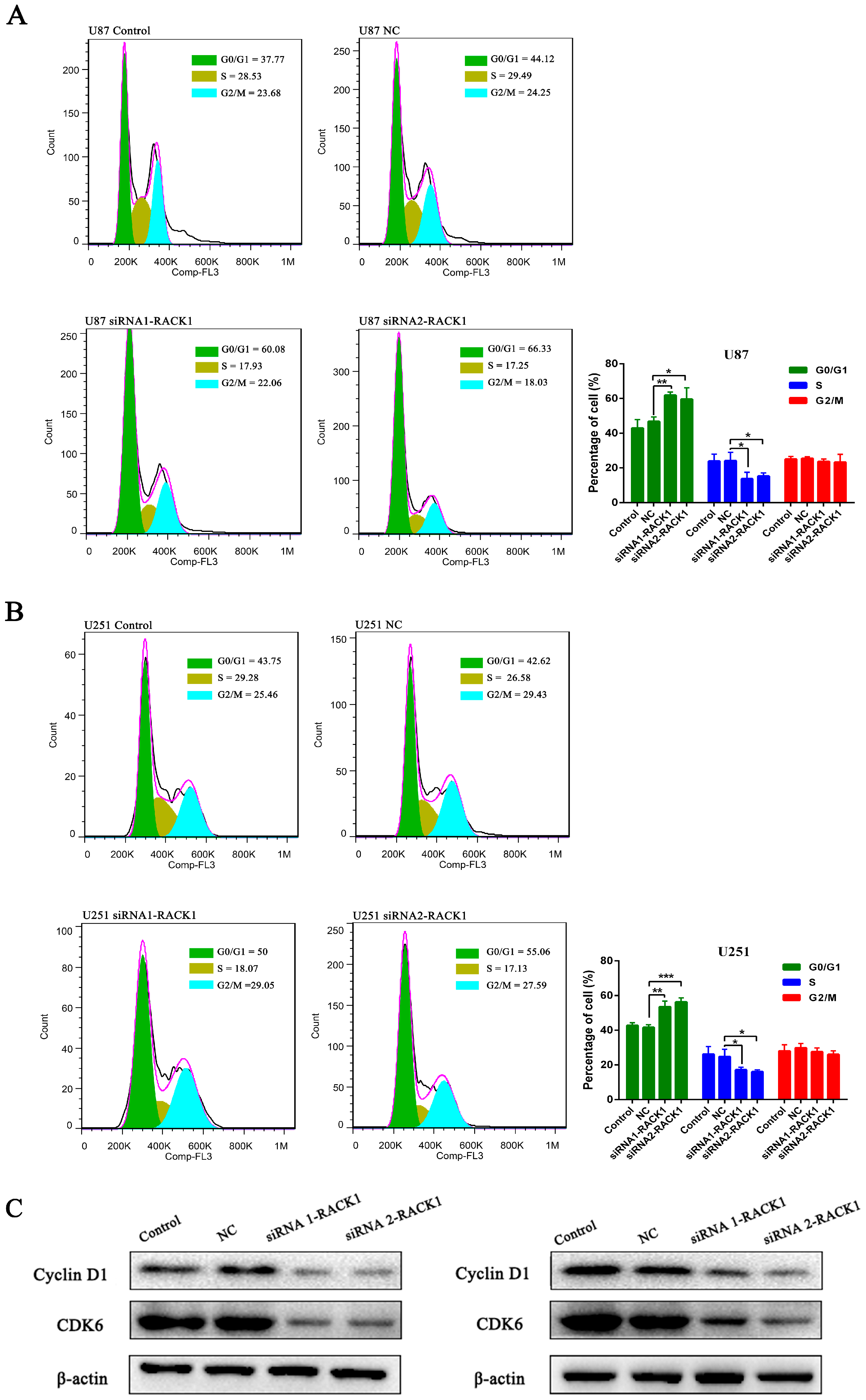
3.4. Effects of RACK1 on U87 and U251 Cell Proliferation
3.5. Downregulation of RACK1 Inhibited the Migration and Invasion of Glioma Cells
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Armstrong, T.S.; Wen, P.Y.; Gilbert, M.R.; Schiff, D. Management of treatment-associated toxicites of anti-angiogenic therapy in patients with brain tumors. Neuro-Oncology 2012, 14, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.J.; Qi, Z.X.; Hua, W.; Zhu, J.J.; Zhang, X.; Yao, Y.; Mao, Y. Increased expression of Capn4 is associated with the malignancy of human glioma. CNS Neurosci. Ther. 2014, 20, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Kong, B.; Kuang, Y.Q.; Cheng, L.; Gu, J.W.; Zhang, J.H.; Shu, H.F.; Yu, S.X.; He, W.Q.; Xing, X.M.; et al. Overexpression of MACC1 protein and its clinical implications in patients with glioma. Tumour Biol. 2014, 35, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Zhu, V.F.; Yang, J.; Lebrun, D.G.; Li, M. Understanding the role of cytokines in glioblastoma multiforme pathogenesis. Cancer Lett. 2012, 316, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Sathornsumetee, S.; Reardon, D.A.; Desjardins, A.; Quinn, J.A.; Vredenburgh, J.J.; Rich, J.N. Molecularly targeted therapy for malignant glioma. Cancer 2007, 110, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, G.; Hegi, M.; Stupp, R.; Weller, M. Clinical implications of molecular neuropathology and biomarkers for malignant glioma. Curr. Neurol. Neurosci. Rep. 2012, 12, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Mochly-Rosen, D.; Khaner, H.; Lopez, J. Identification of intracellular receptor proteins for activated protein kinase C. Proc. Natl. Acad. Sci. USA 1991, 88, 3997–4000. [Google Scholar] [CrossRef] [PubMed]
- McCahill, A.; Warwicker, J.; Bolger, G.B.; Houslay, M.D.; Yarwood, S.J. The RACK1 scaffold protein: A dynamic cog in cell response mechanisms. Mol. Pharmacol. 2002, 62, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.R.; Ron, D.; Kiely, P.A. RACK1, a multifaceted scaffolding protein: Structure and function. Cell Commun. Signal. 2011, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.C.; Rovnak, J.; Quackenbush, S.L. Walleye dermal sarcoma virus Orf B functions through receptor for activated C kinase (RACK1) and protein kinase C. Virology 2008, 375, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Hermanto, U.; Zong, C.S.; Li, W.; Wang, L.H. RACK1, an insulin-like growth factor I (IGF-I) receptor-interacting protein, modulates IGF-I-dependent integrin signaling and promotes cell spreading and contact with extracellular matrix. Mol. Cell. Biol. 2002, 22, 2345–2365. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Xie, D. RACK1, a versatile hub in cancer. Oncogene 2015, 34, 1890–1898. [Google Scholar] [CrossRef] [PubMed]
- Wehner, P.; Shnitsar, I.; Urlaub, H.; Borchers, A. RACK1 is a novel interaction partner of PTK7 that is required for neural tube closure. Development 2011, 138, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.X.; Xu, J.D.; Xu, J.W.; Liu, X.L.; Cheng, Y.Y.; Wang, W.J.; Li, Q.Q.; Chen, Q.; Xu, Z.D.; Liu, X.P. RACK1 promotes breast carcinoma proliferation and invasion/metastasis in vitro and in vivo. Breast Cancer Res. Treat. 2010, 123, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.D.; Wen, Z.; Ji, W.; Ma, Y. RACK1 expression contributes to JNK activity, but JNK activity does not enhance RACK1 expression in hepatocellular carcinoma SMMC-7721 cells. Oncol. Lett. 2015, 9, 2767–2770. [Google Scholar] [CrossRef] [PubMed]
- Campagne, C.; Jule, S.; Alleaume, C.; Bernex, F.; Ezagal, J.; Chateau-Joubert, S.; Estrada, M.; Aubin-Houzelstein, G.; Panthier, J.J.; Egidy, G. Canine melanoma diagnosis: RACK1 as a potential biological marker. Vet. Pathol. 2013, 50, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Min, L.; Wang, X.; Zhao, J.; Chen, H.; Qin, J.; Chen, W.; Shen, Z.; Tang, Z.; Gan, Q.; et al. Loss of RACK1 promotes metastasis of gastric cancer by inducing a miR-302c/IL8 signaling loop. Cancer Res. 2015, 75, 3832–3841. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guo, Y.; Feng, X.; Wang, Z.; Wang, Y.; Deng, P.; Zhang, D.; Wang, R.; Xie, L.; Xu, X.; et al. Receptor for activated C kinase 1 (RACK1): A regulator for migration and invasion in oral squamous cell carcinoma cells. J. Cancer Res. Clin. Oncol. 2012, 138, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Cui, M.; Teng, H.; Wang, F.; Yu, W.; Xu, T. Silencing the receptor of activated C-kinase 1 (RACK1) suppresses tumorigenicity in epithelial ovarian cancer in vitro and in vivo. Int. J. Oncol. 2014, 44, 1252–1258. [Google Scholar] [PubMed]
- Atala, A. Re: Ion channel TRPM8 promotes hypoxic growth of prostate cancer cells via an O2-independent and RACK1-mediated mechanism of HIF-1α stabilization. J. Urol. 2015, 194, 260. [Google Scholar] [CrossRef] [PubMed]
- Berns, H.; Humar, R.; Hengerer, B.; Kiefer, F.N.; Battegay, E.J. RACK1 is up-regulated in angiogenesis and human carcinomas. FASEB J. 2000, 14, 2549–2558. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Sun, L.; Hao, Y.; Wang, L.; Xu, J.; Zhang, W.; Xie, J.; Guo, L.; Zhou, L.; Yun, X.; et al. Ribosomal RACK1 promotes chemoresistance and growth in human hepatocellular carcinoma. J. Clin. Investig. 2012, 122, 2554–2566. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, F.; Cao, F.; Jia, Y.; Wang, J.; Ma, W.; Tan, B.; Wang, K.; Song, Q.; Cheng, Y. RACK1 predicts poor prognosis and regulates progression of esophageal squamous cell carcinoma through its epithelial-mesenchymal transition. Cancer Biol. Ther. 2015, 16, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.Y.; Lee, S.Y.; Lee, W.K.; Jeon, H.S.; Lee, E.B.; Lee, H.C.; Choi, J.E.; Kang, H.G.; Lee, E.J.; Bae, E.Y.; et al. RACK1 is a candidate gene associated with the prognosis of patients with early stage non-small cell lung cancer. Oncotarget 2015, 6, 4451–4466. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.X.; Xu, J.D.; Liu, X.L.; Xu, J.W.; Wang, W.J.; Li, Q.Q.; Chen, Q.; Xu, Z.D.; Liu, X.P. RACK1: A superior independent predictor for poor clinical outcome in breast cancer. Int. J. Cancer 2010, 127, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, W.; Wang, J.; Feng, J.; Wang, Q.; Jin, J.; Lv, M.; Li, X.; Li, Y.; Ma, Y.; et al. Receptor for activated C kinase 1 promotes hepatocellular carcinoma growth by enhancing mitogen-activated protein kinase kinase 7 activity. Hepatology 2013, 57, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Z.; Yao, F.; Li, J.J.; Mao, Z.F.; Hu, P.T.; Long, L.Y.; Li, G.; Ji, X.D.; Shi, S.; Guan, D.X.; et al. RACK1 suppresses gastric tumorigenesis by stabilizing the β-catenin destruction complex. Gastroenterology 2012, 142, 812–823.e15. [Google Scholar] [CrossRef] [PubMed]
- Van Meir, E.G.; Hadjipanayis, C.G.; Norden, A.D.; Shu, H.K.; Wen, P.Y.; Olson, J.J. Exciting new advances in neuro-oncology: The avenue to a cure for malignant glioma. Cancer J. Clin. 2010, 60, 166–193. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Jiang, B.; Ma, J.; Ma, Z.; Wan, X.; Liu, H.; Chen, Z.; Cheng, Q.; Chen, R. Forced downregulation of RACK1 inhibits glioma development by suppressing Src/Akt signaling activity. Onco. Rep. 2013, 30, 2195–2202. [Google Scholar]
- Li, X.; Xiao, Y.; Fan, S.; Xiao, M.; Wang, X.; Chen, X.; Li, C.; Zong, G.; Zhou, G.; Wan, C. RACK1 overexpression associates with pancreatic ductal adenocarcinoma growth and poor prognosis. Exp. Mol. Pathol. 2016, 101, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Yan, C.; Liu, M.; Feng, Y.; Chen, Y. RACK1 promotes prostate cancer cell proliferation, invasion and metastasis. Mol. Med. Rep. 2013, 8, 999–1004. [Google Scholar] [PubMed]
- Zhang, X.; Liu, N.; Ma, D.; Liu, L.; Jiang, L.; Zhou, Y.; Zeng, X.; Li, J.; Chen, Q. Receptor for activated C kinase 1 (RACK1) promotes the progression of OSCC via the Akt/mTOR pathway. Int. J. Oncol. 2016, 49, 539–548. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, H.C.; Kiely, P.A.; O’Connor, R. Effects of RACK1 on cell migration and IGF-I signalling in cardiomyoctes are not dependent on an association with the IGF-IR. Cell Signal. 2007, 19, 2588–2595. [Google Scholar] [CrossRef] [PubMed]
- Kiely, P.A.; Leahy, M.; O’Gorman, D.; O’Connor, R. RACK1-mediated integration of adhesion and insulin-like growth factor I (IGF-I) signaling and cell migration are defective in cells expressing an IGF-I receptor mutated at tyrosines 1250 and 1251. J. Biol. Chem. 2005, 280, 7624–7633. [Google Scholar] [CrossRef] [PubMed]
- Vincent-Salomon, A.; Thiery, J.P. Host microenvironment in breast cancer development: Epithelial-mesenchymal transition in breast cancer development. Breast Cancer Res. 2003, 5, 101–106. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Q.-L.; Huang, Y.-T.; Wang, G.-H.; Liu, Y.-L.; Huang, J.; Qu, Q.; Sun, B.; Hu, L.; Cheng, L.; Chen, S.-H.; et al. Overexpression of RACK1 Promotes Metastasis by Enhancing Epithelial-Mesenchymal Transition and Predicts Poor Prognosis in Human Glioma. Int. J. Environ. Res. Public Health 2016, 13, 1021. https://doi.org/10.3390/ijerph13101021
Lv Q-L, Huang Y-T, Wang G-H, Liu Y-L, Huang J, Qu Q, Sun B, Hu L, Cheng L, Chen S-H, et al. Overexpression of RACK1 Promotes Metastasis by Enhancing Epithelial-Mesenchymal Transition and Predicts Poor Prognosis in Human Glioma. International Journal of Environmental Research and Public Health. 2016; 13(10):1021. https://doi.org/10.3390/ijerph13101021
Chicago/Turabian StyleLv, Qiao-Li, Yuan-Tao Huang, Gui-Hua Wang, Yan-Ling Liu, Jin Huang, Qiang Qu, Bao Sun, Lei Hu, Lin Cheng, Shu-Hui Chen, and et al. 2016. "Overexpression of RACK1 Promotes Metastasis by Enhancing Epithelial-Mesenchymal Transition and Predicts Poor Prognosis in Human Glioma" International Journal of Environmental Research and Public Health 13, no. 10: 1021. https://doi.org/10.3390/ijerph13101021
APA StyleLv, Q. -L., Huang, Y. -T., Wang, G. -H., Liu, Y. -L., Huang, J., Qu, Q., Sun, B., Hu, L., Cheng, L., Chen, S. -H., & Zhou, H. -H. (2016). Overexpression of RACK1 Promotes Metastasis by Enhancing Epithelial-Mesenchymal Transition and Predicts Poor Prognosis in Human Glioma. International Journal of Environmental Research and Public Health, 13(10), 1021. https://doi.org/10.3390/ijerph13101021