Systematic Review: Land Cover, Meteorological, and Socioeconomic Determinants of Aedes Mosquito Habitat for Risk Mapping


Abstract
:1. Introduction
2. Methods
2.1. Search Query
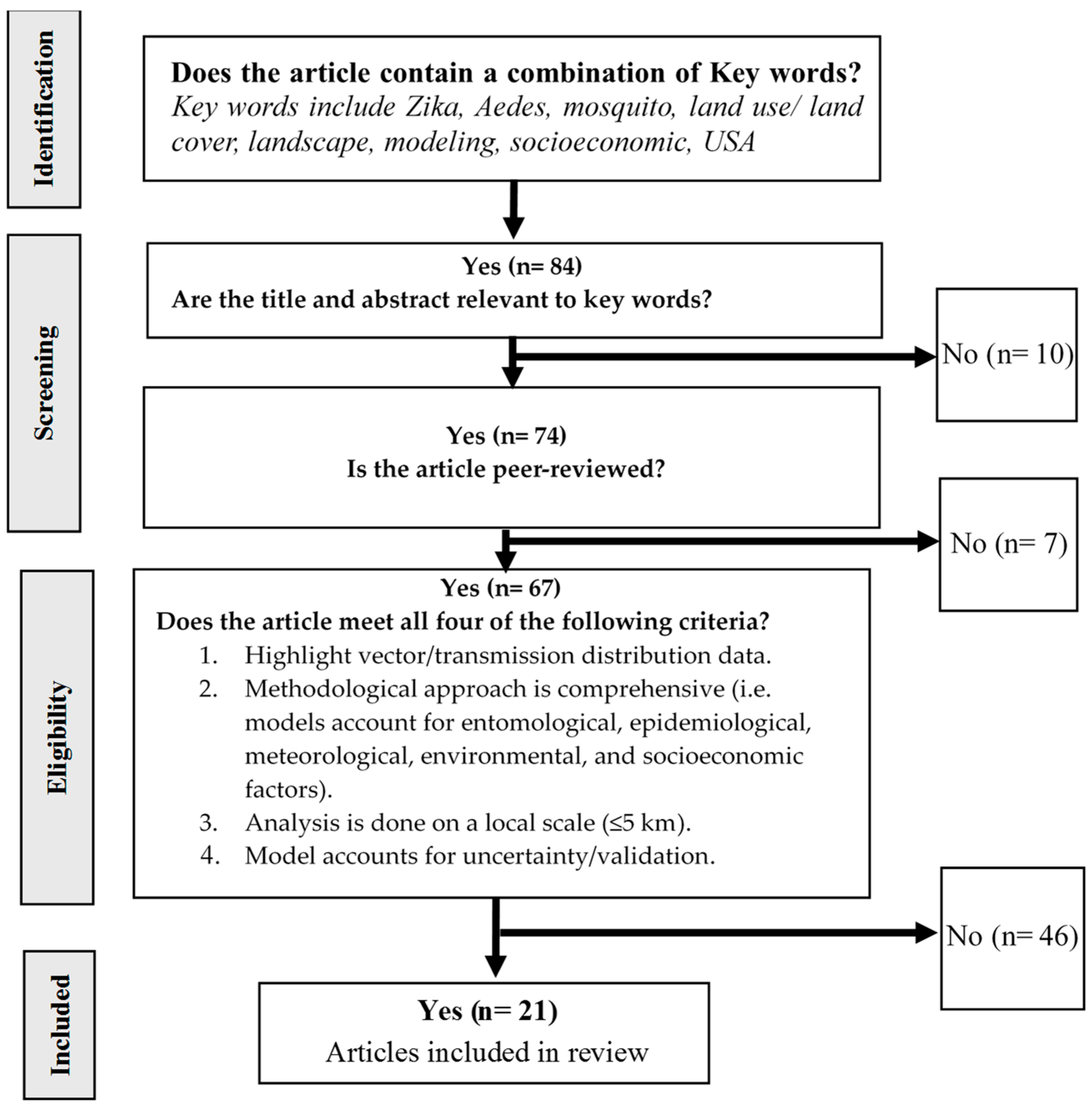
2.2. Hierarchical Decision Tree for Article Selection
3. Results and Discussion
3.1. Relevant and Key Articles
3.2. Dependent Variables
Entomological/Incidence Data
3.3. Explanatory Variables
3.3.1. Meteorological Data
3.3.2. Socioeconomic Data
3.3.3. Environmental Data
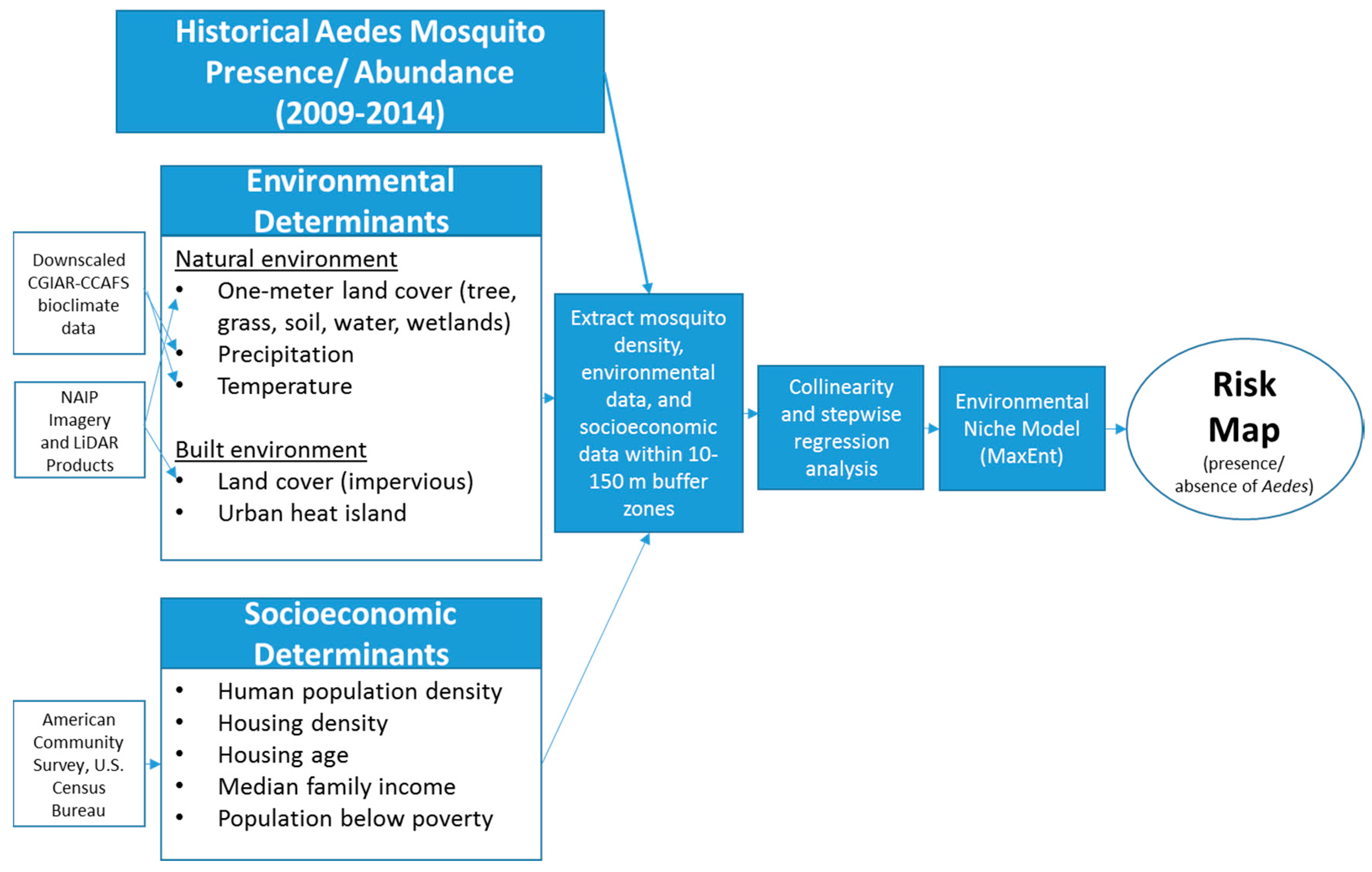
4. Modeling Approaches
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- WHO. The global burden of disease: 2004 update. In World Health Organization Report; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- WHO. World health statistics 2013. In World Health Organization Report; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Lance-Parker, S.; Rebmann, C.; Kramer, S.; Kelly, R. Arboviruses in Georgia. Emergence of West Nile virus. J. Med. Assoc. Ga. 2002, 91, 32–34. [Google Scholar] [PubMed]
- Clements, A.; Paterson, G. The analysis of mortality and survival rates in wild populations of mosquitoes. J. Appl. Ecol. 1981, 18, 373–399. [Google Scholar] [CrossRef]
- Clements, A.N. The Biology of Mosquitoes: Sensory, Reception, and Behaviour; CABI: New York, NY, USA, 1999. [Google Scholar]
- Clements, A.N. The Biology of Mosquitoes, Sensory Reception and Behaviour; CAB International: Wallingford, UK, 1999; Volume 2. [Google Scholar]
- Marini, F.; Caputo, B.; Pombi, M.; Tarsitani, G.; Della Torre, A. Study of Aedes albopictus dispersal in Rome, Italy, using sticky traps in mark–release–recapture experiments. Med. Vet. Entomol. 2010, 24, 361–368. [Google Scholar] [CrossRef] [PubMed]
- David, M.R.; Lourenço-de-Oliveira, R.; Freitas, R.M.D. Container productivity, daily survival rates and dispersal of Aedes aegypti mosquitoes in a high income dengue epidemic neighbourhood of Rio de Janeiro: Presumed influence of differential urban structure on mosquito biology. Mem. Inst. Oswaldo Cruz 2009, 104, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate Change and Range Expansion of the Asian Tiger Mosquito (Aedes albopictus) in Northeastern USA: Implications for Public Health Practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef] [PubMed]
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of The Tiger: Global Risk of Invasion by The Mosquito Aedes albopictus. Vector-Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Delatte, H.; Desvars, A.; Bouétard, A.; Bord, S.; Gimonneau, G.; Vourc’h, G.; Fontenille, D. Blood-feeding behavior of Aedes albopictus, a vector of Chikungunya on La Réunion. Vector Borne Zoonotic Dis. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A.; Shriram, A.N.; Sunish, I.P.; Vidhya, P.T. Host-feeding pattern of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in heterogeneous landscapes of South Andaman, Andaman and Nicobar Islands, India. Parasitol. Res. 2015, 114, 3539–3546. [Google Scholar] [CrossRef] [PubMed]
- GDCH. Georgia Department of Community Health. Mosquito-Borne Viral Diseases Web Page. Available online: https://dph.georgia.gov/EnvironmentalHealth (accessed on 16 October 2017).
- Godsey, M.S.; Blackmore, M.S.; Panella, N.A.; Burkhalter, K.; Gottfried, K.; Halsey, L.A.; Rutledge, R.; Langevin, S.A.; Gates, R.; Lamonte, K.M. West Nile epizootiology in the southeastern United States, 2001. Vector-Borne Zoonotic Dis. 2005, 5, 82–89. [Google Scholar] [CrossRef] [PubMed]
- DeGroote, J.; Mercer, D.R.; Fisher, J.; Sugumaran, R. Spatiotemporal Investigation of Adult Mosquito (Diptera: Culicidae) Populations in an Eastern Iowa County, USA. J. Med. Entomol. 2007, 44, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Golding, N.; Pigott, D.M.; Kraemer, M.U.G.; Messina, J.P.; Reiner, R.C., Jr.; Scott, T.W.; Smith, D.L.; Gething, P.W.; Hay, S.I. Global temperature constraints on Aedes aegypti and Ae. albopictus persistence and competence for dengue virus transmission. Parasites Vectors 2014, 7, 338. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, D.; Thomas, S.M.; Suk, J.E.; Sudre, B.; Hess, A.; Tjaden, N.B.; Beierkuhnlein, C.; Semenza, J.C. Climate change effects on Chikungunya transmission in Europe: Geospatial analysis of vector’s climatic suitability and virus’ temperature requirements. Int. J. Health Geogr. 2013, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Nihei, N.; Kurihara, T. Analysis of Northern Distribution of Aedes albopictus (Diptera: Culicidae) in Japan by Geographical Information System. J. Med. Entomol. 2002, 39, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.; Gething, P.; Bhatt, S.; Messina, J.; Brownstein, J.; Hoen, A.; Moyes, C.; Farlow, A.; Scott, T.; Hay, S. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brady, O.; Messina, J.; Scott, T.; Hay, S. Mapping the global epidemiology of dengue. In Dengue and Dengue Hemorrhagic Fever, 2nd ed.; Gubler, D.J., Ooi, E.E., Vasudevan, S., Farrar, J., Eds.; CABI: Wallingford, UK, 2014; pp. 30–49. [Google Scholar]
- Fischer, D.; Thomas, S.M.; Neteler, M.; Tjaden, N.B.; Beierkuhnlein, C. Climatic suitability of Aedes albopictus in Europe referring to climate change projections: Comparison of mechanistic and correlative niche modelling approaches. Euro Surveill. 2014, 19, 20696. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Alem, I.; De Majo, M.; Campos, R.; Schweigmann, N. Cold season mortality and hatching behavior of Aedes aegypti L. (Diptera: Culicidae) eggs in Buenos Aires City, Argentina. J. Vector Ecol. 2011, 36, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Caminade, C.; Ndione, J.A.; Diallo, M.; MacLeod, D.A.; Faye, O.; Ba, Y.; Dia, I.; Morse, A.P. Rift Valley Fever Outbreaks in Mauritania and Related Environmental Conditions. Int. J. Environ. Res. Public Health 2014, 11, 903–918. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.P.; Luther, C.; Moo-Llanes, D.; Ramsey, J.M.; Danis-Lozano, R.; Peterson, A.T. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed]
- Buckner, E.A.; Blackmore, M.S.; Golladay, S.W.; Covich, A.P. Weather and landscape factors associated with adult mosquito abundance in southwestern Georgia, U.S.A. J. Vector Ecol. 2011, 36, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.H.; Uejio, C.K.; Walker, K.; Ramberg, F.; Moreno, R.; Rosales, C.; Gameros, M.; Mearns, L.O.; Zielinski-Gutierrez, E.; Janes, C.R. Microclimate and Human Factors in the Divergent Ecology of Aedes aegypti along the Arizona, U.S./Sonora, MX Border. EcoHealth 2010, 7, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Landau, K.I.; van Leeuwen, W.J.D. Fine scale spatial urban land cover factors associated with adult mosquito abundance and risk in Tucson, Arizona. J. Vector Ecol. 2012, 37, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Lockaby, G.; Noori, N.; Morse, W.; Zipperer, W.; Kalin, L.; Governo, R.; Sawant, R.; Ricker, M. Climatic, ecological, and socioeconomic factors associated with West Nile virus incidence in Atlanta, Georgia, U.S.A. J. Vector Ecol. 2016, 41, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Reiter, M.E.; LaPointe, D.A. Landscape Factors Influencing the Spatial Distribution and Abundance of Mosquito Vector Culex quinquefasciatus (Diptera: Culicidae) in a Mixed Residential–Agricultural Community in Hawai‘i. J. Med. Entomol. 2007, 44, 861–868. [Google Scholar] [CrossRef]
- Rey, J.R.; Nishimura, N.; Wagner, B.; Braks, M.A.; O’Connell, S.M.; Lounibos, L.P. Habitat Segregation of Mosquito Arbovirus Vectors in South Florida. J. Med. Entomol. 2006, 43, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Apperson, C.S.; Ghosh, S.K.; Cheshire, H.M.; Zeichner, B.C. Spatial Analysis of Aedes albopictus (Diptera: Culicidae) Oviposition in Suburban Neighborhoods of a Piedmont Community in North Carolina. J. Med. Entomol. 2006, 43, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.A.; Christofferson, R.C.; Silva, N.J.B.; Vasquez, C.; Mores, C.N.; Wearing, H.J. Modeling Mosquito-Borne Disease Spread in U.S. Urbanized Areas: The Case of Dengue in Miami. PLoS ONE 2016, 11, e0161365. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, I.; Turbow, D.; Gomez, F.; Ninivaggi, D.V.; Campbell, S.R. Predictive Mapping of Human Risk for West Nile Virus (WNV) Based on Environmental and Socioeconomic Factors. PLoS ONE 2011, 6, e23280. [Google Scholar] [CrossRef] [PubMed]
- Sallam, M.F.; Xue, R.-D.; Pereira, R.M.; Koehler, P.G. Ecological niche modeling of mosquito vectors of West Nile virus in St. John’s County, Florida, USA. Parasites Vectors 2016, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, A.J.; Morin, C.W.; Steinhoff, D.F.; Wilhelmi, O.; Hayden, M.; Quattrochi, D.A.; Reiskind, M.; Lloyd, A.L.; Smith, K.; Schmidt, C.A.; et al. On the Seasonal Occurrence and Abundance of the Zika Virus Vector Mosquito Aedes Aegypti in the Contiguous United States. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Ashby, J.; Moreno-Madriñán, M.; Yiannoutsos, C.; Stanforth, A. Niche Modeling of Dengue Fever Using Remotely Sensed Environmental Factors and Boosted Regression Trees. Remote Sens. 2017, 9, 328. [Google Scholar] [CrossRef]
- Gleiser, R.M.; Zalazar, L.P. Distribution of mosquitoes in relation to urban landscape characteristics. Bull. Entomol. Res. 2010, 100, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Koyadun, S.; Butraporn, P.; Kittayapong, P. Ecologic and Sociodemographic Risk Determinants for Dengue Transmission in Urban Areas in Thailand. Int. Perspect. Infect. Dis. 2012, 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Rubio, A.; Cardo, M.V.; Carbajo, A.E.; Vezzani, D. Imperviousness as a predictor for infestation levels of container-breeding mosquitoes in a focus of dengue and Saint Louis encephalitis in Argentina. Acta Trop. 2013, 128, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Troyo, A.; Fuller, D.O.; Calderón-Arguedas, O.; Beier, J.C. A geographical sampling method for surveys of mosquito larvae in an urban area using high-resolution satellite imagery. J. Vector Ecol. J. Soc. Vector Ecol. 2008, 33, 1–7. [Google Scholar] [CrossRef]
- Wijayanti, S.P.M.; Porphyre, T.; Chase-Topping, M.; Rainey, S.M.; McFarlane, M.; Schnettler, E.; Biek, R.; Kohl, A. The Importance of Socio-Economic Versus Environmental Risk Factors for Reported Dengue Cases in Java, Indonesia. PLoS Negl. Trop. Dis. 2016, 10, e0004964. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Munga, S.; Minakawa, N.; Githeko, A.K.; Yan, G. Spatial relationship between adult malaria vector abundance and environmental factors in western Kenya highlands. Am. J. Trop. Med. Hyg. 2007, 77, 29–35. [Google Scholar] [PubMed]
- Massad, E.; Tan, S.-H.; Khan, K.; Wilder-Smith, A. Estimated Zika virus importations to Europe by travellers from Brazil. Glob. Health Action 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Kraemer, M.U.G.; Brady, O.J.; Pigott, D.M.; Shearer, F.M.; Weiss, D.J.; Golding, N.; Ruktanonchai, C.W.; Gething, P.W.; Cohn, E.; et al. Mapping global environmental suitability for Zika virus. eLife 2016, 5, e15272. [Google Scholar] [CrossRef] [PubMed]
- Manrique, P.D.; Xu, C.; Hui, P.M.; Johnson, N.F. Atypical viral dynamics from transport through popular places. Phys. Rev. 2016, 94. [Google Scholar] [CrossRef] [PubMed]
- Sallam, M.F.; Lippi, C.; Xue, R.-D. Spatial analysis of arbovirus transmisson in St. Johns County, Florida. Tech. Bull. Fla. Mosq. Control Assoc. 2016, 10, 18–29. [Google Scholar]
- Sallam, M.F.; Michaels, S.R.; Riegel, C.; Tian, Y.; Pereira, R.M.; Koehler, P.G. Habitat suitability model and risk assessment of Zika Virus mosquito vectors in city of New Orleans, LA, USA. in preparation.
- Sallam, M.F.; Michaels, S.R.; Riegel, C.; Pereira, R.M.; Zipperer, W.; Lockaby, B.G.; Koehler, P.G. Spatio-Temporal Distribution of Vector-Host Contact (VHC) Ratios and Ecological Niche Modelingof the West Nile Virus Mosquito Vector, Culex quinquefasciatus, in the City of New Orleans, LA, USA. Int. J. Environ. Res. Public Health 2017, 14, 892. [Google Scholar] [CrossRef] [PubMed]
- Honório, N.A.; Silva, W.D.C.; Leite, P.J.; Gonçalves, J.M.; Lounibos, L.P.; Lourenço-de-Oliveira, R. Dispersal of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in an urban endemic dengue area in the State of Rio de Janeiro, Brazil. Mem. Inst. Oswaldo Cruz 2003, 98, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P.; Amador, M.A.; Anderson, R.A.; Clark, G.G. Dispersal of Aedes aegypti in an urban area after blood feeding as demonstrated by rubidium-marked eggs. Am. J. Trop. Med. Hyg. 1995, 52, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.G.; Cline, B.L.; Ruiz-Tibén, E.; Lee, D.; Romney-Joseph, H.; Rivera-Correa, E. Aedes aegypti in Puerto Rico: environmental determinants of larval abundance and relation to dengue virus transmission. Am. J. Trop. Med. Hyg. 1978, 27, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Aiken, S.R.; Frost, D.B.; Leigh, C.H. Dengue hemorrhagic fever and rainfall in Peninsular Malaysia: Some suggested relationships. Soc. Sci. Med. Part D Med. Geogr. 1980, 14, 307–316. [Google Scholar] [CrossRef]
- Chadee, D.D. Aedes aegypti surveillance in Tobago, West Indies (1983–1988). J. Am. Mosquito Control Assoc. 1990, 6, 148–150. [Google Scholar]
- Scott, T.W.; Morrison, A.C.; Lorenz, L.H.; Clark, G.G.; Strickman, D.; Kittayapong, P.; Zhou, H.; Edman, J.D. Longitudinal Studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: Population Dynamics. J. Med. Entomol. 2000, 37, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Focks, D.A.; Daniels, E.; Haile, D.G.; Keesling, J.E. A simulation model of the epidemiology of urban dengue fever: literature analysis, model development, preliminary validation, and samples of simulation results. Am. J. Trop. Med. Hyg. 1995, 53, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Jetten, T.H.; Focks, D.A. Changes in the distribution of dengue transmission under climate warming scenarios. Am. J. Trop. Med. Hyg. 1997, 57, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Koopman, J.S.; Prevots, D.R.; Vaca Marin, M.A.; Gomez Dantes, H.; Zarate Aquino, M.L.; Longini, I.M.J.; Sepulveda Amor, J. Determinants and predictors of dengue infection in Mexico. Am. J. Epidemiol. 1991, 133, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Basto, E.; Prevots, D.R.; Zarate, M.L.; Silva, J.L.; Sepulveda-Amor, J. First Reported Outbreak of Classical Dengue Fever at 1700 Meters above Sea Level in Guerrero State, Mexico, June 1988. Am. J. Trop. Med. Hyg. 1992, 46, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Hylton, A.R. Low humidity water-retention ability in Eretmapodites chrysogaster and Aedes albopictus. J. Insect Physiol. 1967, 13, 153–157. [Google Scholar] [CrossRef]
- Ruiz, M.O.; Tedesco, C.; McTighe, T.J.; Austin, C.; Kitron, U.D. Environmental and social determinants of human risk during a West Nile virus outbreak in the greater Chicago area, 2002. Int. J. Health Geogr. 2004, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrigan, R.J.; Thomassen, H.A.; Buermann, W.; Cummings, R.F.; Kahn, M.E.; Smith, T.B. Economic Conditions Predict Prevalence of West Nile Virus. PLoS ONE 2010, 5, e15437. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Takahashi, R.M.; Carroll, B.D.; Quiring, R. Delinquent Mortgages, Neglected Swimming Pools, and West Nile Virus, California. Emerg. Infect. Dis. 2008, 14, 1747–1749. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, P.; Wilcox, B.A. Parasites, ecosystems and sustainability: An ecological and complex systems perspective. Int. J. Parasitol. 2005, 35, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.; Lu, Y.; Lin, T.; Chu, P.; Chang, S.; Huang, J.; Chen, K.; King, C. Predisposing factors of dengue cases by random effect model in the largest dengue haemorrhagic fever epidemic in Taiwan in 1998. Dengue Bull. 2000, 24, 46–52. [Google Scholar]
- Thammapalo, S.; Supaporn, M.; Virasakdi, C. Effectiveness of Space Spraying on the Transmission of Dengue/Dengue Hemorrhagic Fever (Df/Dhf) in an Urban Area of Southern Thailand. J. Trop. Med. 2012, 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Barker, C.M.; Paulson, S.L.; Cantrell, S.; Davis, B.S. Habitat Preferences and Phenology of Ochlerotatus triseriatus and Aedes albopictus (Diptera: Culicidae) in Southwestern Virginia. J. Med. Entomol. 2003, 40, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Swanson, J.; Lancaster, M.; Anderson, J.; Crandell, M.; Haramis, L.; Grimstad, P.; Kitron, U. Overwintering and establishment of Aedes albopictus (Diptera: Culicidae) in an urban La Crosse Virus enzootic site in Illinois. J. Med. Entomol. 2000, 37, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Jemal, Y.; Al-Thukair, A.A. Combining GIS application and climatic factors for mosquito control in Eastern Province, Saudi Arabia. Saudi J. Biol. Sci. 2016. [Google Scholar] [CrossRef]
- Dixon, P. A Primer of Ecological Statistics by Nicholas J. Gotelli and Aaron M. Ellison. Q. Rev. Biol. 2014, 89, 168–169. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Sallam, M.F.; Al Ahmed, A.M.; Abdel-Dayem, M.S.; Abdullah, M.A.R. Ecological Niche Modeling and Land Cover Risk Areas for Rift Valley Fever Vector, Culex tritaeniorhynchus Giles in Jazan, Saudi Arabia. PLoS ONE 2013, 8, e65786. [Google Scholar] [CrossRef] [PubMed]
- Pickard, B.R.; Daniel, J.; Mehaffey, M.; Jackson, L.E.; Neale, A. EnviroAtlas: A new geospatial tool to foster ecosystem services science and resource management. Ecosyst. Serv. 2015, 14, 45–55. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Reference | Model Type | Threshold/Validation Indicators |
|---|---|---|
| Buckner et al. [27] | Mechanistic, Priori RA | β, P, AICc-values |
| Hayden et al. [28] | MLM | AUC, ROC, QIC-values |
| Landau and van Leeuwen [29] | SMRA | R2, CV, residual plot, |
| Lockaby et al. [30] | SMRA | β, R2, p-values |
| Reiter and LaPointe [31] | MLM | AICc values |
| Rey et al. [32] | SMRA, PCA | β, Z, p-values |
| Richards et al. [33] | RA, Kriging | R2, CV, residual plot, predicted vs. observed, goodness-of fit, RMSSE |
| Robert et al. [34] | GLS | Pr, p-values |
| Rochlin et al. [35] | MLM | β, p-values |
| Sallam et al. [36] | SMRA, MaxEnt | AICc, AUC, ROC, CV, β, p-values |
| Monaghan et al. [37] | SB, DMSiM | Mean of ensemble models, mean of two life stages model |
| Ashby et al. [38] | BRT | RMSE, Pr, p-values |
| Gleiser and Zalazar [39] | RA | R2, p-values |
| Koyadun et al. [40] | ANOVA, MLM, LR, WT | R2, p-values |
| Rubio et al. [41] | GLMM, ML | R2, p-values |
| Troyo et al. [42] | ANOVA | P, Kappa values |
| Wijayanti et al. [43] | BPSA, INLA | IRR, DIC, predicted vs. observed |
| Zhou et al. [44] | SA, GI, SMRA | p-values |
| Massad et al. [45] | MDM | NA |
| Messina et al. [46] | BRT | AUC value, 10% omission rate value, CV |
| Manrique et al. [47] | SIR, SIS | NA |
| Reference | Entomol./Inc. * | Meteorology * | Socioeconomic ** | Environment ** | Topography ** |
|---|---|---|---|---|---|
| Buckner et al. [27] | A | P, T, RH, DI | NA | 10 (1 m, aerial) | NA |
| Hayden et al. [28] | E | T, RH | 6 | 2 (1 m, Ikonos-aerial) | NA |
| Landau and van Leeuwen [29] | log A | NA | NA | Sq. root 11 (1 m, NAIP, aerial, LiDAR) | NA |
| Lockaby et al. [30] | A | P, T, PET, SM | 2 | 7 (1 m, aerial) | NA |
| Reiter and LaPointe [31] | A, IR | P | NA | 4 (30 m, LSTM) | Elevation |
| Rey et al. [32] | log E, log L | NA | NA | Arcsine sq. root 17 (1 m, aerial) | NA |
| Richards et al. [33] | E, A | P, T | NA | 2 (1 m, Ikonos) | NA |
| Robert et al. [34] | VHR | NA | 2 | NA | NA |
| Rochlin et al. [35] | DIn3 | NA | 4 | 7 (30 m, USGS, MODIS) | NA |
| Sallam et al. [36] | A, Ser. | P, T | 1 | 2 (250 m, MODIS, USGS) | 5 |
| Monaghan et al. [37] | E, L, P, A | P, T, RH | 2 | NA | NA |
| Ashby et al. [38] | DIn1 | LST, nLST | 1 | 9 (250 m, MODIS) | Elevation |
| Gleiser and Zalazar [39] | A | NA | NA | 4 (30 m, LSTM) | NA |
| Koyadun et al. [40] | DIn1 | NA | 22 | 4 (household level) | NA |
| Rubio et al. [41] | L | NA | NA | 1 (30 m, LSTM) | NA |
| Troyo et al. [42] | L | NA | NA | 5 (0.5–15 m, QB-ASTER) | NA |
| Wijayanti et al. [43] | DIn1 | P, LST, nLST | 53 | 1 (1 km, MODIS) | NA |
| Zhou et al. [44] | L | NA | 2 | 4 (1 m, Ikonos) | Elevation |
| Massad et al. [45] | DIn2 | NA | 4 | NA | NA |
| Messina et al. [46] | DIn2 | P, T, RH | 1 | 1 (5 km, MODIS) | NA |
| Manrique et al. [47] | DIn2 | NA | 1 | NA | NA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sallam, M.F.; Fizer, C.; Pilant, A.N.; Whung, P.-Y. Systematic Review: Land Cover, Meteorological, and Socioeconomic Determinants of Aedes Mosquito Habitat for Risk Mapping. Int. J. Environ. Res. Public Health 2017, 14, 1230. https://doi.org/10.3390/ijerph14101230
Sallam MF, Fizer C, Pilant AN, Whung P-Y. Systematic Review: Land Cover, Meteorological, and Socioeconomic Determinants of Aedes Mosquito Habitat for Risk Mapping. International Journal of Environmental Research and Public Health. 2017; 14(10):1230. https://doi.org/10.3390/ijerph14101230
Chicago/Turabian StyleSallam, Mohamed F., Chelsea Fizer, Andrew N. Pilant, and Pai-Yei Whung. 2017. "Systematic Review: Land Cover, Meteorological, and Socioeconomic Determinants of Aedes Mosquito Habitat for Risk Mapping" International Journal of Environmental Research and Public Health 14, no. 10: 1230. https://doi.org/10.3390/ijerph14101230
APA StyleSallam, M. F., Fizer, C., Pilant, A. N., & Whung, P. -Y. (2017). Systematic Review: Land Cover, Meteorological, and Socioeconomic Determinants of Aedes Mosquito Habitat for Risk Mapping. International Journal of Environmental Research and Public Health, 14(10), 1230. https://doi.org/10.3390/ijerph14101230