Gene Expression Response in Peripheral Blood Cells of Petroleum Workers Exposed to Sub-Ppm Benzene Levels

 , , ,
, , ,
Abstract
:
1. Introduction
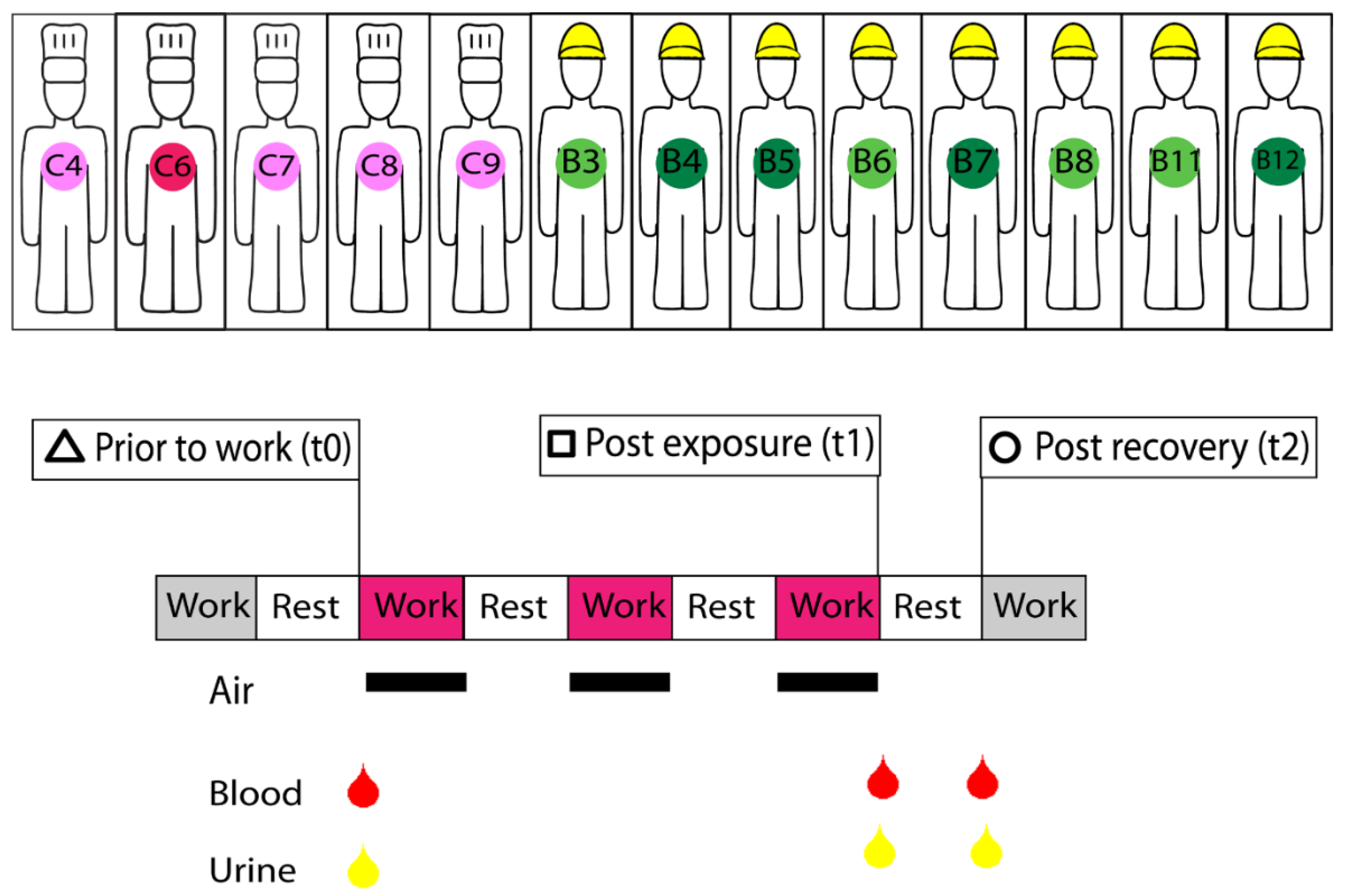
2. Materials and Methods
3. Results
3.1. Study Population and Exposure
3.2. Selecting Marker Genes Predictive of Exposure Group at Current Exposure
3.3. Considering Gene Expression Differences in Worker Groups Prior to Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lan, Q.; Zhang, L.; Li, G.; Vermeulen, R.; Weinberg, R.S.; Dosemeci, M.; Rappaport, S.M.; Shen, M.; Alter, B.P.; Wu, Y.; et al. Hematotoxicity in workers exposed to low levels of benzene. Science 2004, 306, 1774–1776. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). IARC Monographs on the Evaluation of Carcinogenic Risks of Chemicals to Humans; Chemical Agents and Related Occupations; IARC: Lyon, France, 2012; Volume 100F. [Google Scholar]
- Glass, D.C.; Gray, C.N.; Jolley, D.J.; Gibbons, C.; Sim, M.R.; Fritschi, L.; Adams, G.G.; Bisby, J.A.; Manuell, R. Leukemia risk associated with low-level benzene exposure. Epidemiology 2003, 14, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Vlaanderen, J.; Portengen, L.; Rothman, N.; Lan, Q.; Kromhout, H.; Vermeulen, R. Flexible meta-regression to assess the shape of the benzene-leukemia exposure-response curve. Environ. Health Perspect. 2010, 118, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Loomis, D.; Guyton, K.Z.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Vilahur, N.; Mattock, H.; Straif, K. Carcinogenicity of benzene. Lancet Oncol. 2017, 18, 1574–1575. [Google Scholar] [CrossRef]
- Irons, R.D.; Kerzic, P.J. Cytogenetics in benzene-associated myelodysplastic syndromes and acute myeloid leukemia: New insights into a disease continuum. Ann. N. Y. Acad. Sci. 2014, 1310, 84–88. [Google Scholar] [CrossRef] [PubMed]
- McHale, C.M.; Zhang, L.; Lan, Q.; Vermeulen, R.; Li, G.; Hubbard, A.E.; Porter, K.E.; Thomas, R.; Portier, C.J.; Shen, M.; et al. Global gene expression profiling of a population exposed to a range of benzene levels. Environ. Health Perspect. 2011, 119, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Hubbard, A.E.; McHale, C.M.; Zhang, L.; Rappaport, S.M.; Lan, Q.; Rothman, N.; Vermeulen, R.; Guyton, K.Z.; Jinot, J.; et al. Characterization of changes in gene expression and biochemical pathways at low levels of benzene exposure. PLoS ONE 2014, 9, e91828. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, C.; McHale, C.M.; Hubbard, A.E.; Zhang, L.; Thomas, R.; Vermeulen, R.; Li, G.; Shen, M.; Rappaport, S.M.; Yin, S.; et al. Identification of gene expression predictors of occupational benzene exposure. PLoS ONE 2018, 13, e0205427. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M.; Kim, S.; Lan, Q.; Vermeulen, R.; Waidyanatha, S.; Zhang, L.; Li, G.; Yin, S.; Hayes, R.B.; Rothman, N.; et al. Evidence that humans metabolize benzene via two pathways. Environ. Health Perspect. 2009, 117, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Kirkeleit, J.; Ulvestad, E.; Riise, T.; Bratveit, M.; Moen, B.E. Acute suppression of serum IgM and IgA in tank workers exposed to benzene. Scand. J. Immunol. 2006, 64, 690–698. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Schwartz, D.M.; Villarino, A.V.; Gadina, M.; McInnes, I.B.; Laurence, A. The JAK-STAT Pathway: Impact on Human Disease and Therapeutic Intervention. Annu. Rev. Med. 2015, 66, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.; Haque, T.; Gritsman, K. Inflammatory Signaling Pathways in Preleukemic and Leukemic Stem Cells. Front. Oncol. 2017, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Irish, J.M.; Hovland, R.; Krutzik, P.O.; Perez, O.D.; Bruserud, Ø.; Gjertsen, B.T.; Nolan, G.P. Single cell profiling of potentiated phospho-protein networks in cancer cells. Cell 2004, 118, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Kirkeleit, J.; Riise, T.; Bratveit, M.; Pekari, K.; Mikkola, J.; Moen, B.E. Biological monitoring of benzene exposure during maintenance work in crude oil cargo tanks. Chem.-Biol. Interact. 2006, 164, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Hopf, N.B.; Kirkeleit, J.; Bratveit, M.; Succop, P.; Talaska, G.; Moen, B.E. Evaluation of exposure biomarkers in offshore workers exposed to low benzene and toluene concentrations. Int. Arch. Occup. Environ. Health 2012, 85, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Pekari, K.; Vainiotalo, S.; Heikkila, P.; Palotie, A.; Luotamo, M.; Riihimaki, V. Biological monitoring of occupational exposure to low levels of benzene. Scand. J. Work Environ. Health 1992, 18, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Ettre, L.S.; Jones, E. Quantitative analysis with headspace gas chromatography using multiple headspace extraction. Chromatogr. Newslett. 1984, 12, 1–3. [Google Scholar]
- Hopf, N.B.; Kirkeleit, J.; Kramer, S.L.; Moen, B.; Succop, P.; Genter, M.B.; Carreón, T.; Mack, J.; Talaska, G. Urinary 1-hydroxypyrene levels in offshore workers. Int. Arch. Occup. Environ. Health 2010, 83, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Hornung, R.W.; Reed, L.D. Estimation of average concentration in the presence of non-detectable values. Appl. Occup. Environ. Hyg. 1990, 5, 46–51. [Google Scholar] [CrossRef]
- A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 4 September 2018).
- Stavrum, A.K.; Petersen, K.; Jonassen, I.; Dysvik, B. Analysis of gene expression data using J-express. Curr. Protoc. Bioinf. 2008, 21, 7.3.1–7.3.25. [Google Scholar] [CrossRef]
- Brazma, A.; Hingamp, P.; Quackenbush, J.; Sherlock, G.; Spellman, P.; Stoeckert, C.; Aach, J.; Ansorge, W.; Ball, C.A.; Causton, H.C. Minimum information about a microarray experiment (MIAME)—Toward standards for microarray data. Nat. Genet. 2001, 29, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Arak, T.; Vilo, J. g:Profiler—A web server for functional interpretation of gene lists (2011 update). Nucleic Acids Res. 2011, 39, W307–W315. [Google Scholar] [CrossRef] [PubMed]
- Hotelling, H. Analysis of a Complex of Statistical Variables into Principal Components. J. Educ. Psychol. 1933, 24, 417–441. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Hastie, T. Regularization and variable selection via the elastic net. J. R. Stat. Soc. B 2005, 67, 301–320. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Reid, L.H.; Jones, W.D.; Shippy, R.; Warrington, J.A.; Baker, S.C.; Collins, P.J.; De Longueville, F.; Kawasaki, E.S.; Lee, K.Y.; et al. The MicroArray Quality Control (MAQC) project shows inter- and intraplatform reproducibility of gene expression measurements. Nat. Biotechnol. 2006, 24, 1151–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollati, V.; Baccarelli, A.; Hou, L.; Bonzini, M.; Fustinoni, S.; Cavallo, D.; Byun, H.M.; Jiang, J.; Marinelli, B.; Pesatori, A.C.; et al. Changes in DNA methylation patterns in subjects exposed to low-dose benzene. Cancer Res. 2007, 67, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Rota, F.; Conti, A.; Campo, L.; Favero, C.; Cantone, L.; Motta, V.; Polledri, E.; Mercadante, R.; Dieci, G.; Bollati, V.; et al. Epigenetic and Transcriptional Modifications in Repetitive Elements in Petrol Station Workers Exposed to Benzene and MTBE. Int. J. Environ. Res. Public Health 2018, 15, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batliner, J.; Mancarelli, M.M.; Jenal, M.; Reddy, V.A.; Fey, M.F.; Torbett, B.E.; Tschan, M.P. CLEC5A (MDL-1) is a novel PU.1 transcriptional target during myeloid differentiation. Mol. Immunol. 2011, 48, 714–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrest, M.S.; Lan, Q.; Hubbard, A.E.; Zhang, L.; Vermeulen, R.; Zhao, X.; Li, G.; Wu, Y.Y.; Shen, M.; Yin, S.; et al. Discovery of novel biomarkers by microarray analysis of peripheral blood mononuclear cell gene expression in benzene-exposed workers. Environ. Health Perspect. 2005, 113, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Selvik, L.K.; Fjeldbo, C.S.; Flatberg, A.; Steigedal, T.S.; Misund, K.; Anderssen, E.; Doseth, B.; Langaas, M.; Tripathi, S.; Beisvag, V.; et al. The duration of gastrin treatment affects global gene expression and molecular responses involved in ER stress and anti-apoptosis. BMC Genom. 2013, 14, 429. [Google Scholar] [CrossRef] [PubMed]
- Stenehjem, J.S.; Kjærheim, K.; Bråtveit, M.; Samuelsen, S.O.; Barone-Adesi, F.; Rothman, N.; Lan, Q.; Grimsrud, T.K. Benzene exposure and risk of lymphohaematopoietic cancers in 25,000 offshore oil industry workers. Br. J. Cancer 2015, 112, 1603–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkeleit, J.; Riise, T.; Bratveit, M.; Moen, B.E. Increased risk of acute myelogenous leukemia and multiple myeloma in a historical cohort of upstream petroleum workers exposed to crude oil. Cancer Cause Control. 2008, 19, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Vlaanderen, J.; Lan, Q.; Kromhout, H.; Rothman, N.; Vermeulen, R. Occupational benzene exposure and the risk of chronic myeloid leukemia: A meta-analysis of cohort studies incorporating study quality dimensions. Am. J. Ind. Med. 2012, 55, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Tamiya, T.; Kashiwagi, I.; Takahashi, R.; Yasukawa, H.; Yoshimura, A. Suppressors of Cytokine Signaling (SOCS) Proteins and JAK/STAT Pathways Regulation of T-Cell Inflammation by SOCS1 and SOCS3. Arterioscl. Throm. Vas. 2011, 31, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Johan, M.F.; Bowen, D.T.; Frew, M.E.; Goodeve, A.C.; Reilly, J.T. Aberrant methylation of the negative regulators RASSFIA, SHP-1 and SOCS-1 in myelodysplastic syndromes and acute myeloid leukaemia. Br. J. Haematol. 2005, 129, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, D.B. Temporal variation in the association between benzene and leukemia mortality. Environ. Health Perspect. 2008, 116, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Darrall, K.G.; Figgins, J.A.; Brown, R.D.; Phillips, G.F. Determination of benzene and associated volatile compounds in mainstream cigarette smoke. Analyst 1998, 123, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Schnatter, A.R.; Kerzic, P.J.; Zhou, Y.; Chen, M.; Nicolich, M.J.; Lavelle, K.; Armstrong, T.W.; Bird, M.G.; Lin, L.; Fu, H.; et al. Peripheral blood effects in benzene-exposed workers. Chem. Biol. Interact. 2010, 184, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Bird, M.G.; Wetmore, B.A.; Letinski, D.J.; Nicolich, M.; Chen, M.; Schnatter, A.R.; Whitman, F.T. Influence of toluene co-exposure on the metabolism and genotoxicity of benzene in mice using continuous and intermittent exposure. Chem. Biol. Interact. 2010, 184, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Kirkeleit, J.; Riise, T.; Bratveit, M.; Moen, B.E. Benzene exposure on a crude oil production vessel. Ann. Occup. Hyg. 2006, 50, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Campagna, M.; Satta, G.; Campo, L.; Flore, V.; Ibba, A.; Meloni, M.; Giuseppina Tocco, M.; Avataneo, G.; Flore, C.; Fustinoni, S.; et al. Biological monitoring of low-level exposure to benzene. Med. Lav. 2012, 103, 338–346. [Google Scholar] [PubMed]
- Weisel, C.P. Benzene exposure: An overview of monitoring methods and their findings. Chem. Biol. Interact. 2010, 184, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoet, P.; De Smedt, E.; Ferrari, M.; Imbriani, M.; Maestri, L.; Negri, S.; De Wilde, P.; Lison, D.; Haufroid, V. Evaluation of urinary biomarkers of exposure to benzene: Correlation with blood benzene and influence of confounding factors. Int. Arch. Occup. Environ. Health 2009, 82, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Manini, P.; De Palma, G.; Andreoli, R.; Poli, D.; Petyx, M.; Corradi, M.; Mutti, A.; Apostoli, P. Biological monitoring of low benzene exposure in Italian traffic policemen. Toxicol. Lett. 2008, 181, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Lovreglio, P.; D’Errico, M.N.; Fustinoni, S.; Drago, I.; Barbieri, A.; Sabatini, L.; Carrieri, M.; Apostoli, P.; Soleo, L. Biomarkers of internal dose for the assessment of environmental exposure to benzene. J. Environ. Monit. 2011, 13, 2921–2928. [Google Scholar] [CrossRef] [PubMed]
- Allan, L.L.; Sherr, D.H. Disruption of human plasma cell differentiation by an environmental polycyclic aromatic hydrocarbon: A mechanistic immunotoxicological study. Environ. Health 2010, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- American Conference of Governmental Industrial Hygenists (ACGIH). TLVs and BEIs Based on the Documentation of the Threshold Limit Values for Chemical Substances and Physical Agents and Biological Substances; ACGIH: Cincinnatti, OH, USA, 2018.
- Committee for Risk Assessment (RAC). Opinion on Scientific Evaluation of Occupational Exposure Limits for Benzene; Contract No.: ECHA/RAC/0-000000-1412-86-187/F; Committee for Risk Assessment (RAC): Helsinki, Finland, 2018. [Google Scholar]
- Zhang, X.; Xue, Z.; Li, H.; Yan, L.; Yang, Y.; Wang, Y.; Duan, J.; Li, L.; Chai, F.; Cheng, M.; et al. Ambient volatile organic compounds pollution in China. J. Environ. Sci. 2017, 55, 69–75. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unbalanced | Balanced | ||
|---|---|---|---|---|
| Tank Workers | Referents | Tank Workers | Referents | |
| Subjects | 8 | 5 | 4 | 4 |
| Sex (male/female) | 8/0 | 4/1 | 4/0 | 4/0 |
| Age in years (range) | 37.6 (27–54) | 48.2 (42–59) | 43.5 (38–54) | 45.5 (42–53) |
| Current smokers (%) | 3 (37.5%) | 1 (20%) | 0 (0%) | 1 (25%) |
| Former smokers (%) | 4 (50%) | 2 (40%) | 4 (100%) | 1 (25%) |
| Benzene in air (ppm) | AM (SD) = 0.21 (0.17) GM (GSD) = 0.14 (2.9) Range 0.03–0.45 | — | AM (SD) = 0.25 (0.16) GM (GSD) = 0.19 (2.6) Range 0.08–0.50 | — |
| Toluene in air (ppm) | AM (SD) = 0.36 (0.23) GM (GSD) = 0.22 (2.6) Range 0.10–0.8 | — | AM (SD) = 0.37 (0.22) GM (GSD) = 0.30 (2.5) Range 0.14–0.71 | — |
| Ethylbenzene in air (ppm) | AM (SD) = 0.10 (0.08) GM (GSD) = 0.07 (2.56) Range 0.02 – 0.22 | — | AM (SD) = 0.12 (0.08) GM (GSD) = 0.10 (2.29) Range 0.03–0.21 | — |
| Xylene in air, all isomers (ppm) | AM (SD) = 0.54 (0.40) GM (GSD) = 0.42 (2.27) Range 0.13–1.14 | — | AM (SD) = 0.63 (0.37) GM (GSD) = 0.53(2.07) Range 0.20–1.04 | — |
| Benzene in blood (nmol/L) | AM (SD) = 19.3 (12) GM (GSD) = 15.4 (2.2) Range 4.0–38.0 | AM (SD) = 0.9 (0.2) GM (GSD) = 0.87 (1.4) Range 0.5–1.0 | AM (SD) = 22.8 (12.4) GM (GSD) = 19.7 (1.9) Range 8.0–38.0 | AM (SD) = 0.9 (0.3) GM (GSD) = 0.84 (1.4) Range 0.5–1.0 |
| Toluene in blood (nmol/L) | AM (SD) = 28.8 (21.6) GM (GSD) = 21.7 (2.3) Range 6.0–73.0 | AM (SD) = 0.9 (0.2) GM (GSD) = 0.87 (1.4) Range 0.5–1.0 | AM (SD) = 36.5 (27.0) GM (GSD) = 28.0 (2.5) Range 8.0–73.0 | AM (SD) = 1.0 (-) GM (GSD) = (-) Range 1.0–1.0 |
| Benzene in urine (nmol/L) | AM (SD) = 71.3 (110) GM (GSD) = 36.6 (2.9) Range 15.0–333.0 | AM (SD) = 1.1 (0.8) GM (GSD) = 0.87 (1.4) Range 0.5–2.0 | AM (SD) = 121 (146) GM (GSD) = 65.9 (3.7) Range 20.0–333.0 | AM (SD) = 0.88 (0.8) GM (GSD) = 0.71 (2.0) Range 0.5–2.0 |
| Toluene in urine (nmol/L) | AM (SD) = 15.5 (5,3) GM (GSD) = 14.38 (1.6) Range 5.0–23.0 | AM (SD) = 3.6 (1.7) GM (GSD) = 3.13 (2.0) Range 1.0–5.0 | AM (SD) = 18.8 (3.8) GM (GSD) = 18.45 (1.2) Range 14.0–23.0 | AM (SD) = 3.3 (1.7) GM (GSD) = 2.78 (2.0) Range 1.0–5.0 |
| t,t-muconic acid (µmol/L) | AM (SD) = 4.2 (4.1) GM (GSD) = 3.0 (1.9) Range 1.6–14.1 | AM (SD) = 2.6 (1.5) GM (GSD) = 2.27 (1.7) Range 1.1–5.1 | AM (SD) = 5.8 (5.6) GM (GSD) = 4.35 (2.2) Range 2.5–14.1 | AM (SD) = 2.7 (1.7) GM (GSD) = 2.35 (1.9) Range 1.1–5.1 |
| 1-Hydroxypyrene in urine (1-HP, µg/g creatinine) | AM (SD) = 0.43 (1.1) GM (GSD) = 0.05 (7.0) Range 0.01–3.18 | AM (SD) = 0.01 (0.01) GM (GSD) = 0.01 (1.9) Range 0.006–0.036 | AM (SD) = 0.8 (1.6) GM (GSD) = 0.09 (13.4) Range 0.007–3.18 | AM (SD) = 0.01 (0.01) GM (GSD) = 0.01 (2.1) Range 0.006–0.036 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jørgensen, K.M.; Færgestad Mosleth, E.; Hovde Liland, K.; Hopf, N.B.; Holdhus, R.; Stavrum, A.-K.; Gjertsen, B.T.; Kirkeleit, J. Gene Expression Response in Peripheral Blood Cells of Petroleum Workers Exposed to Sub-Ppm Benzene Levels. Int. J. Environ. Res. Public Health 2018, 15, 2385. https://doi.org/10.3390/ijerph15112385
Jørgensen KM, Færgestad Mosleth E, Hovde Liland K, Hopf NB, Holdhus R, Stavrum A-K, Gjertsen BT, Kirkeleit J. Gene Expression Response in Peripheral Blood Cells of Petroleum Workers Exposed to Sub-Ppm Benzene Levels. International Journal of Environmental Research and Public Health. 2018; 15(11):2385. https://doi.org/10.3390/ijerph15112385
Chicago/Turabian StyleJørgensen, Katarina M., Ellen Færgestad Mosleth, Kristian Hovde Liland, Nancy B. Hopf, Rita Holdhus, Anne-Kristin Stavrum, Bjørn Tore Gjertsen, and Jorunn Kirkeleit. 2018. "Gene Expression Response in Peripheral Blood Cells of Petroleum Workers Exposed to Sub-Ppm Benzene Levels" International Journal of Environmental Research and Public Health 15, no. 11: 2385. https://doi.org/10.3390/ijerph15112385
APA StyleJørgensen, K. M., Færgestad Mosleth, E., Hovde Liland, K., Hopf, N. B., Holdhus, R., Stavrum, A. -K., Gjertsen, B. T., & Kirkeleit, J. (2018). Gene Expression Response in Peripheral Blood Cells of Petroleum Workers Exposed to Sub-Ppm Benzene Levels. International Journal of Environmental Research and Public Health, 15(11), 2385. https://doi.org/10.3390/ijerph15112385