The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe

 , ,
, ,
Abstract
:1. Introduction
1.1. Viruses Associated with Transmission by Culex pipiens
1.1.1. Emergence of West Nile Virus in Europe
1.1.2. Emergence of Usutu Virus in Europe
1.1.3. Sindbis Virus in Europe
1.1.4. Other Viruses Transmitted by Culex Mosquitoes
1.2. Culex pipiens Taxonomy
1.3. Delineation of Species, Ecoforms and Hybrids
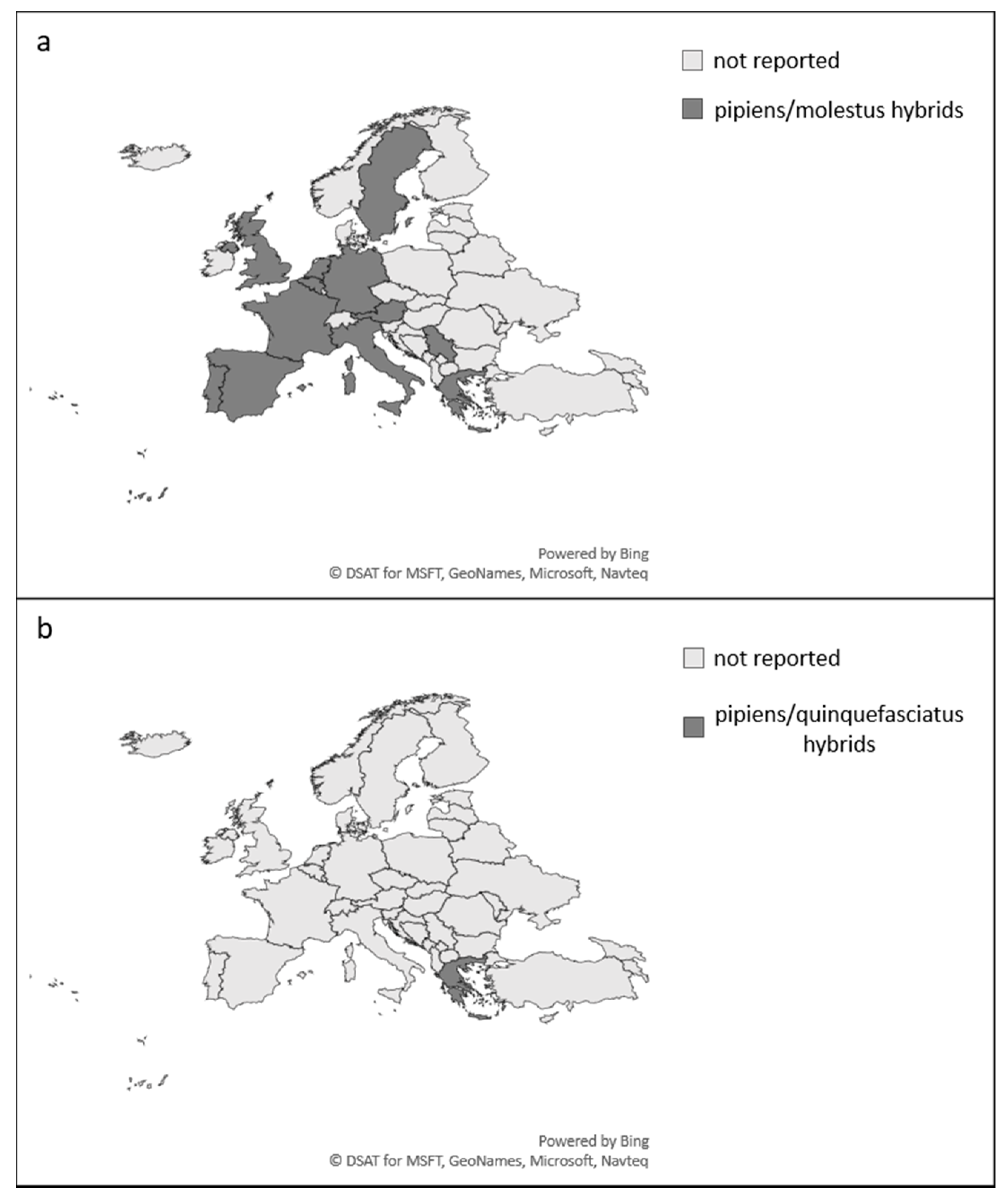
1.4. Distribution and Hybridisation
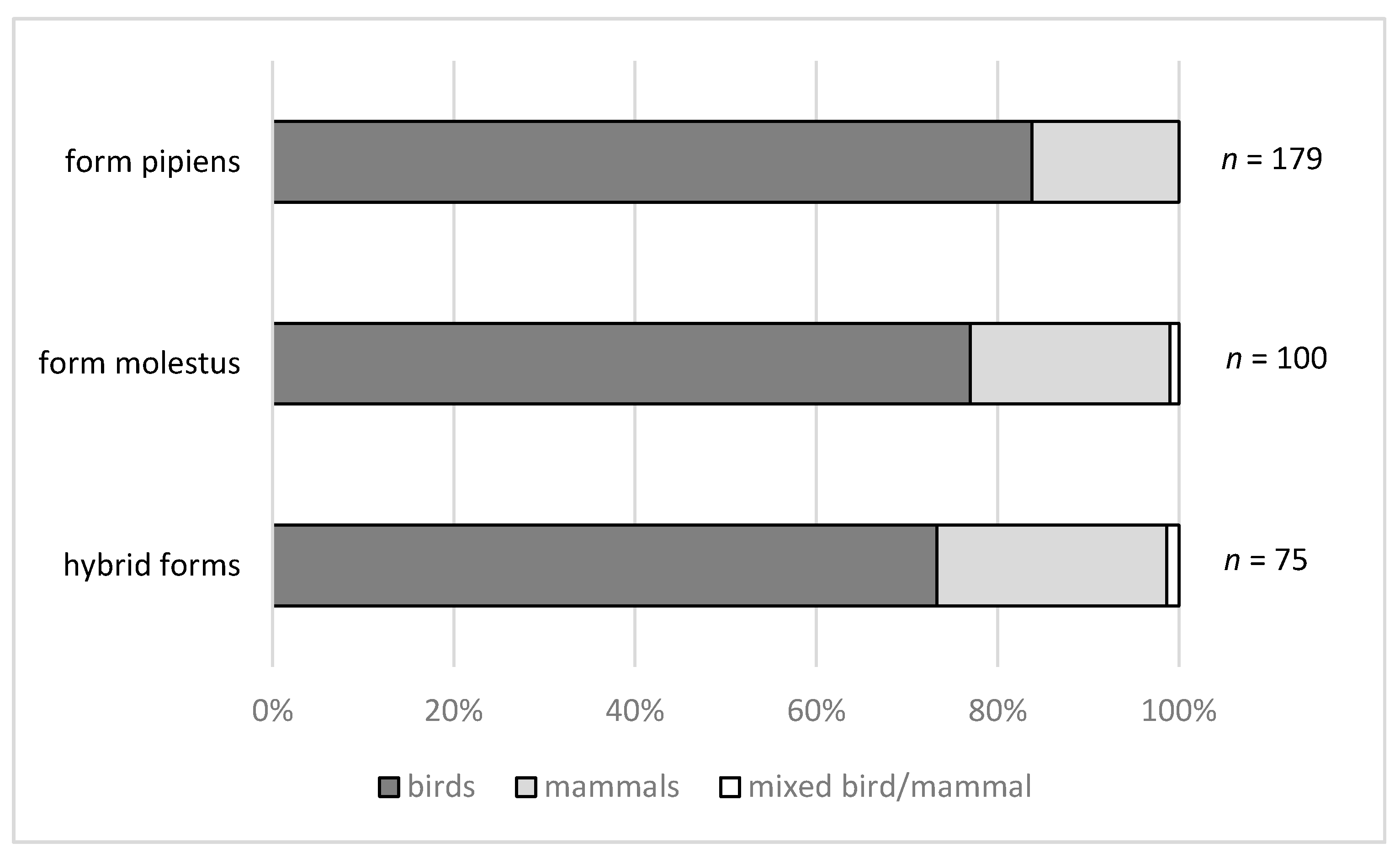
1.5. Culex pipiens Blood-Feeding Behaviour
2. Future Research Directions
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
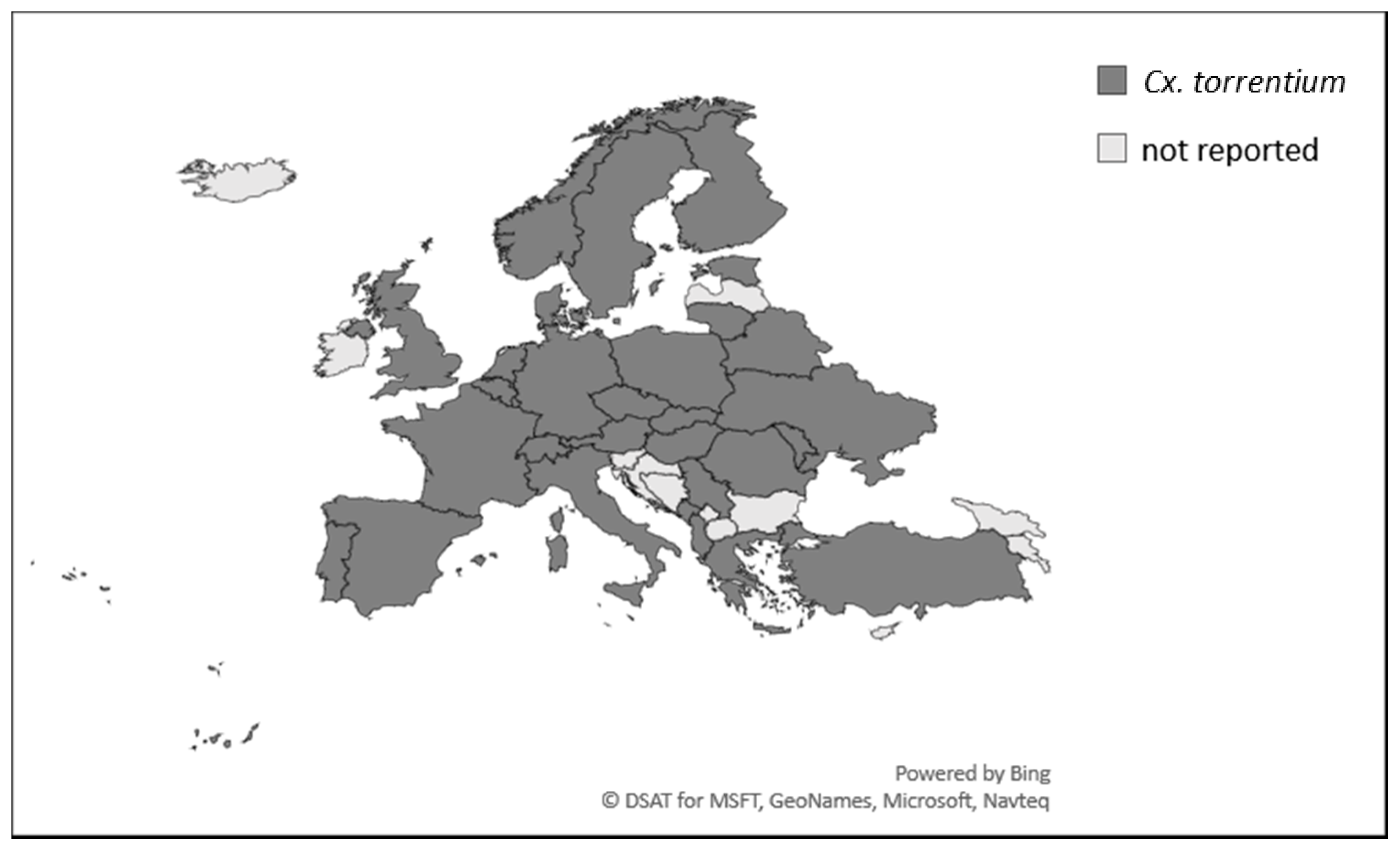
- Culex torrentium (Figure 3)
References
- Gratz, N.G. The Vector- and Rodent-Borne Diseases of Europe and North America: Their Distribution and Public Health Burden; Cambridge University Press: Cambridge, UK, 2006; ISBN 9780511541896. [Google Scholar]
- Harbach, R.E. Mosquito Taxonomic Inventory. Available online: http://mosquito-taxonomic-inventory.info (accessed on 16 September 2015).
- Harbach, R.E.; Howard, T.M. Index of currently recognized mosquito species (Diptera: Culicidae). Eur. Mosq. Bull. 2007, 23, 1–66. [Google Scholar]
- Medlock, J.M.; Hansford, K.M.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; van Bortel, W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 2015, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Adhami, J.; Reiter, P. Introduction and establishment of Aedes (Stegomyia) albopictus Skuse (Diptera: Culicidae) in Albania. J. Am. Mosq. Control Assoc. 1998, 14, 340–343. [Google Scholar] [PubMed]
- Sabatini, A.; Raineri, V.; Trovato, G.; Coluzzi, M. Aedes albopictus in Italy and possible diffusion of the species into the Mediterranean area. Parassitologia 1990, 32, 301–304. [Google Scholar] [PubMed]
- European Centre for Disease Prevention and Control. Aedes Albopictus—Factsheet for Experts; European Centre for Disease Prevention and Control: Solna, Sweden, 2016. [Google Scholar]
- Margarita, Y.; Grácio, A.J.; Lencastre, I.; Silva, A.C.; Novo, M.T.; Sousa, C.A. First record of Aedes (Stegomyia) aegypti (Linnaeus, 1762) (Diptera, Culicidae) in Madeira Island-Portugal. Acta Parasitológica Port. 2006, 13, 59–61. [Google Scholar]
- Akiner, M.M.; Demirci, B.; Babuadze, G.; Robert, V.; Schaffner, F. Spread of the invasive mosquitoes Aedes aegypti and Aedes albopictus in the Black Sea region increases risk of Chikungunya, Dengue, and Zika outbreaks in Europe. PLoS Negl. Trop. Dis. 2016, 10, e0004664. [Google Scholar] [CrossRef]
- Versteirt, V.; de Clercq, E.M.; Fonseca, D.M.; Pecor, J.; Schaffner, F.; Coosemans, M.; van Bortel, W. Bionomics of the established exotic mosquito species Aedes koreicus in Belgium, Europe. J. Med. Entomol. 2012, 49, 1226–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capelli, G.; Drago, A.; Martini, S.; Montarsi, F.; Soppelsa, M.; Delai, N.; Ravagnan, S.; Mazzon, L.; Schaffner, F.; Mathis, A.; et al. First report in Italy of the exotic mosquito species Aedes (Finlaya) koreicus, a potential vector of arboviruses and filariae. Parasit. Vectors 2011, 4, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrame, A.; Angheben, A.; Bisoffi, Z.; Monteiro, G.; Marocco, S.; Calleri, G.; Lipani, F.; Gobbi, F.; Canta, F.; Castelli, F.; et al. Imported chikungunya infection, Italy. Emerg. Infect. Dis. 2007, 13, 1264–1266. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Maioli, G.; Galletti, G.; Gelati, A.; Barbieri, I.; Tamba, M.; et al. Evidence of simultaneous circulation of West Nile and Usutu viruses in mosquitoes sampled in Emilia-Romagna region (Italy) in 2009. PLoS ONE 2010, 5, e14324. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Xanthopoulou, K.; Gewehr, S.; Mourelatos, S. Detection of West Nile virus lineage 2 in mosquitoes during a human outbreak in Greece. Clin. Microbiol. Infect. 2011, 17, 1176–1180. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Capelli, G.; Monaco, F.; Polci, A.; Russo, F.; Di Gennaro, A.; Marini, V.; Teodori, L.; Montarsi, F.; Pinoni, C.; et al. Evidence of West Nile virus lineage 2 circulation in Northern Italy. Vet. Microbiol. 2012, 158, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.P.G.; Freitas, F.B.; Novo, M.T.; Sousa, C.A.; Rodrigues, J.C.; Alves, R.; Esteves, A. Mosquito surveys and West Nile virus screening in two different areas of southern Portugal, 2004–2007. Vector-Borne Zoonotic Dis. 2010, 10, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Engler, O.; Savini, G.; Papa, A.; Figuerola, J.; Groschup, M.H.; Kampen, H.; Medlock, J.M.; Vaux, A.G.C.; Wilson, A.J.; Werner, D.; et al. European surveillance for West Nile virus in mosquito populations. Int. J. Environ. Res. Public Health 2013, 10, 4869–4895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortuna, C.; Remoli, M.E.; Di Luca, M.; Severini, F.; Toma, L.; Benedetti, E.; Bucci, P.; Montarsi, F.; Minelli, G.; Boccolini, D.; et al. Experimental studies on comparison of the vector competence of four Italian Culex pipiens populations for West Nile virus. Parasit. Vectors 2015, 8, 463. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Miesen, P.; Vogels, C.B.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Heal. 2015, 1, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Engel, D.; Jöst, H.; Wink, M.; Börstler, J.; Bosch, S.; Garigliany, M.M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; et al. Reconstruction of the evolutionary history and dispersal of Usutu virus, a neglected emerging arbovirus in Europe and Africa. mBio 2016, 7, e01938. [Google Scholar] [CrossRef] [PubMed]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Kilpatrick, A.M. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, E.B. Culex pipiens pipiens mosquitoes: Taxonomy, distribution, ecology, physiology, genetics, applied importance and control. Pensoft Ser. Parasitol. 2000, 2, 250. [Google Scholar]
- Lundström, J.O. Mosquito-borne viruses in western Europe: A review. J. Vector Ecol. 1999, 24, 1–39. [Google Scholar] [PubMed]
- Hubálek, Z. Mosquito-borne viruses in Europe. Parasitol. Res. 2008, 103, S29–S43. [Google Scholar] [CrossRef] [PubMed]
- Snow, K.R.; Medlock, J.M. The potential impact of climate change on the distribution and prevalence of mosquitoes in Britain. Eur. Mosq. Bull. 2006, 21, 1–10. [Google Scholar]
- Becker, N. Influence of climate change on mosquito development and mosquito-borne diseases in Europe. Parasitol. Res. 2008, 103, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Peinado, S.A.; Osorio, J.E.; Bartholomay, L.C. Culex pipiens and Aedes triseriatus mosquito susceptibility to Zika Virus. Emerg. Infect. Dis. 2016, 22, 1857–1859. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.L.; Romo, H.; Duggal, N.K.; Tzeng, W.P.; Burkhalter, K.L.; Brault, A.C.; Savage, H.M. Transmission incompetence of Culex quinquefasciatus and Culex pipiens pipiens from North America for Zika virus. Am. J. Trop. Med. Hyg. 2017, 96, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, A.; Jansen, S.; Lühken, R.; Leggewie, M.; Badusche, M.; Pluskota, B.; Becker, N.; Vapalahti, O.; Schmidt-Chanasit, J.; Tannich, E. Experimental transmission of zika virus by mosquitoes from central Europe. Eurosurveillance 2017, 22, 30437. [Google Scholar] [CrossRef] [PubMed]
- Amraoui, F.; Atyame-Nten, C.; Vega-Rúa, A.; Lourenço-De-Oliveira, R.; Vazeille, M.; Failloux, A.B. Culex mosquitoes are experimentally unable to transmit zika virus. Eurosurveillance 2016, 21, 30333. [Google Scholar] [CrossRef] [PubMed]
- Boccolini, D.; Toma, L.; Di Luca, M.; Severini, F.; Romi, R.; Remoli, M.E.; Sabbatucci, M.; Venturi, G.; Rezza, G.; Fortuna, C. Experimental investigation of the susceptibility of Italian Culex pipiens mosquitoes to zika virus infection. Eurosurveillance 2016, 21, 30328. [Google Scholar] [CrossRef] [PubMed]
- Hart, C.E.; Roundy, C.M.; Azar, S.R.; Huang, J.H.; Yun, R.; Reynolds, E.; Leal, G.; Nava, M.R.; Vela, J.; Stark, P.M.; et al. Zika virus vector competency of mosquitoes, Gulf Coast, United States. Emerg. Infect. Dis. 2017, 23, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Rückert, C.; Chotiwan, N.; Nguyen, C.; Garcia Luna, S.M.; Fauver, J.R.; Foy, B.D.; Perera, R.; Black, W.C.; Kading, R.C.; et al. Vector competence of American mosquitoes for three strains of Zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0005101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, T.; Lai, Z.; Zhang, Z.; Jia, Z.; Zhou, G.; Williams, T.; Xu, J.; Gu, J.; Zhou, X.; et al. Competence of Aedes aegypti, Ae. albopictus, and Culex quinquefasciatus mosquitoes as Zika virus vectors, China. Emerg. Infect. Dis. 2017, 23, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-J.S.; Ayers, V.B.; Lyons, A.C.; Unlu, I.; Alto, B.W.; Cohnstaedt, L.W.; Higgs, S.; Vanlandingham, D.L. Culex species mosquitoes and Zika virus. Vector-Borne Zoonotic Dis. 2016, 16, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.X.; Li, C.X.; Deng, Y.Q.; Xing, D.; Liu, Q.M.; Wu, Q.; Sun, A.J.; Dong, Y.D.; Cao, W.C.; Qin, C.F.; et al. Culex pipiens quinquefasciatus: A potential vector to transmit Zika virus. Emerg. Microbes Infect. 2016, 5, e102. [Google Scholar] [CrossRef] [PubMed]
- Guedes, D.R.D.; Paiva, M.H.S.; Donato, M.M.A.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.W.S.; Saraiva, K.L.A.; Crespo, M.M.; Rezende, T.M.T.; Wallau, G.L.; et al. Zika virus replication in the mosquito Culex quinquefasciatus in Brazil. Emerg. Microbes Infect. 2017, 6, e69. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Snow, K.R.; Leach, S.A. Possible ecology and epidemiology of medically important mosquito-borne arboviruses in Great Britain. Epidemiol. Infect. 2007, 135, 466–482. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Tomasello, D.; Schlagenhauf, P. Chikungunya and dengue autochthonous cases in Europe, 2007–2012. Travel Med. Infect. Dis. 2013, 11, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.; Tagliazucchi, S.; Savini, G.; Monaco, F.; Simone, M.L.; Lelli, R.; et al. First human case of Usutu virus neuroinvasive infection, Italy, August–September 2009. Euro. Surveill. 2009, 14, 19446. [Google Scholar] [PubMed]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne Flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile virus. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef]
- Bunning, M.L.; Bowen, R.A.; Bruce Cropp, C.; Sullivan, K.G.; Davis, B.S.; Komar, N.; Godsey, M.S.; Baker, D.; Hettler, D.L.; Holmes, D.A.; et al. Experimental infection of horses with West Nile virus. Emerg. Infect. Dis. 2002, 8, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar] [CrossRef]
- Work, T.H.; Hurlbut, H.S.; Taylor, R.M. Indigenous wild birds of the Nile-delta as potential West Nile virus circulating reservoirs. Am. J. Trop. Med. Hyg. 1955, 4, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Cabre, O.; Grandadam, M.; Marié, J.-L.; Gravier, P.; Prangé, A.; Santinelli, Y.; Rous, V.; Bourry, O.; Durand, J.-P.; Tolou, H.; et al. West Nile Virus in horses, sub-Saharan Africa. Emerg. Infect. Dis. 2006, 12, 1958–1960. [Google Scholar] [CrossRef] [PubMed]
- Melnick, J.L.; Paul, J.R.; Riordan, J.T.; Barnett, V.H.; Goldblum, N.; Zabin, E. Isolation from human sera in Egypt of a virus apparently identical to West Nile Virus. Proc. Soc. Exp. Biol. Med. 1951, 77, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Peyrefitte, C.N.; El Harrak, M.; Murri, S.; Leblond, A.; Zeller, H.G. West Nile Virus in Morocco, 2003. Emerg. Infect. Dis. 2005, 11, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Lafri, I.; Prat, C.M.; Bitam, I.; Gravier, P.; Besbaci, M.; Zeroual, F.; Ben-Mahdi, M.H.; Davoust, B.; Leparc-Goffart, I. Seroprevalence of West Nile virus antibodies in equids in the North-East of Algeria and detection of virus circulation in 2014. Comp. Immunol. Microbiol. Infect. Dis. 2017, 50, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Rappole, J.H.; Hubálek, Z. Migratory birds and West Nile virus. J. Appl. Microbiol. 2003, 94, 47S–58S. [Google Scholar] [CrossRef] [PubMed]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.J.; Sabatier, P. Bird migration routes and risk for pathogen dispersion into western Mediterranean wetlands. Emerg. Infect. Dis. 2007, 13, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Fall, G.; Di Paola, N.; Faye, M.; Dia, M.; de Melo Freire, C.C.; Loucoubar, C.; de Andrade Zanotto, P.M.; Faye, O.; Sall, A.A. Biological and phylogenetic characteristics of West African lineages of West Nile virus. PLoS Negl. Trop. Dis. 2017, 11, e0006078. [Google Scholar] [CrossRef] [PubMed]
- Cernescu, C.; Nedelcu, N.I.; Tardei, G.; Ruta, S.; Tsai, T.F. Continued transmission of West Nile virus to humans in southeastern Romania, 1997–1998. J. Infect. Dis. 2000, 181, 710–712. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S. Origin of the West Nile virus responsible for an outbreak of encephalitis in the Northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.B.; Komar, N.; Nasci, R.S.; Montgomery, S.P.; O’Leary, D.R.; Campbell, G.L. Epidemiology and transmission dynamics of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Hamer, G.; Kitron, U.D.; Goldberg, T.L.; Brawn, J.D.; Loss, S.R.; Ruiz, M.O.; Hayes, D.B.; Walker, E.D. Host selection by Culex pipiens mosquitoes and West Nile virus amplification. Am. J. Trop. Med. Hyg. 2009, 80, 268–278. [Google Scholar] [PubMed]
- Andreadis, T.G. The contribution of Culex pipiens complex mosquitoes to transmission and persistence of West Nile virus in North America. J. Am. Mosq. Control Assoc. 2012, 28, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Tamba, M.; Tassinari, M.; Gelati, A.; Cordioli, P.; Angelini, P.; et al. Usutu virus persistence and West Nile virus inactivity in the Emilia-Romagna region (Italy) in 2011. PLoS ONE 2013, 8, e63978. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, E.; Fernández-Pinero, J.; Llorente, F.; Vázquez, A.; Moreno, A.; Agüero, M.; Cordioli, P.; Tenorio, A.; Jimeénez-Clavero, M.Á. Phylogenetic relationships of Western Mediterranean West Nile virus strains (1996–2010) using full-length genome sequences: Single or multiple introductions? J. Gen. Virol. 2011, 92, 2512–2522. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J.M. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [PubMed]
- Mani, P.; Rossi, G.; Perrucci, S.; Bertini, S. Mortality of Turdus merula in Tuscany. Sel. Vet. 1998, 8, 749–753. [Google Scholar]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, Western Europe, 2016. Eurosurveillance 2017, 22, 30452. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Erdélyi, K.; Brunthaler, R.; Dán, Á.; Weissenböck, H.; Nowotny, N. Usutu virus, Austria and Hungary, 2010–2016. Emerg. Microbes Infect. 2017, 6, e85. [Google Scholar] [CrossRef] [PubMed]
- Cavrini, F.; Gaibani, P.; Longo, G.; Pierro, A.M.; Rossini, G.; Bonilauri, P.; Gerundi, G.E.; Di Benedetto, F.; Pasetto, A.; Girardis, M.; et al. Usutu virus infection in a patient who underwent orthotropic liver transplantation, Italy, August–September 2009. Eurosurveillance 2009, 14, 19448. [Google Scholar] [PubMed]
- Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savic, V.; Stevanovic, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection in Croatia, August–September 2013: Clinical and laboratory features. J. Neurovirol. 2014, 21, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Maier, P.; Müller, S.; Kress, J.; Chudy, M.; Bialonski, A.; Schlaphof, A.; Jansen, S.; Jöst, H.; Tannich, E.; et al. Blood donor screening for west nile virus (WNV) revealed acute usutu virus (USUV) infection, Germany, September 2016. Eurosurveillance 2017, 22, 30501. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.M.; Hurlbut, H.S.; Work, T.H.; Kingston, J.R.; Frothingham, T.E. Sindbis virus: A newly recognized arthropod-transmitted virus. Am. J. Trop. Med. Hyg. 1955, 4, 844–862. [Google Scholar] [CrossRef] [PubMed]
- Espmark, A.; Niklasson, B. Ockelbo disease in Sweden: epidemiological, clinical, and virological data from the 1982 outbreak. Am. J. Trop. Med. Hyg. 1984, 33, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, S.; Human, S.; Williams, J.; van Wilpe, E.; Pretorius, M.; Swanepoel, R.; Venter, M. Sindbis and Middelburg old world alphaviruses associated with neurologic disease in horses, South Africa. Emerg. Infect. Dis. 2015, 21, 2225–2229. [Google Scholar] [CrossRef] [PubMed]
- Adouchief, S.; Smura, T.; Sane, J.; Vapalahti, O.; Kurkela, S. Sindbis virus as a human pathogen—Epidemiology, clinical picture and pathogenesis. Rev. Med. Virol. 2016, 26, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Lundström, J.O.; Pfeffer, M. Phylogeographic structure and evolutionary history of Sindbis virus. Vector-Borne Zoonotic Dis. 2010, 10, 889–907. [Google Scholar] [CrossRef]
- Lundström, J.O.; Niklasson, B.; Francy, D.B. Swedish Culex torrentium and Cx. pipiens (Diptera: Culicidae) as experimental vectors of Okelbo virus. J. Med. Entomol. 1990, 27, 561–563. [Google Scholar]
- Lundström, J.O.; Turell, M.J.; Niklasson, B. Effect of environmental temperature on the vector competence of Culex pipiens and Cx. torrentium for Okelbo virus. Am. J. Trop. Med. Hyg. 1990, 43, 534–542. [Google Scholar] [CrossRef]
- Hesson, J.C.; Verner-Carlsson, J.; Larsson, A.; Ahmed, R.; Lundkvist, Å.; Lundström, J.O. Culex torrentium mosquito role as major enzootic vector defined by rate of Sindbis virus infection, Sweden, 2009. Emerg. Infect. Dis. 2015, 21, 875–878. [Google Scholar] [CrossRef] [PubMed]
- Eiden, M.; Ziegler, U.; Keller, M.; Müller, K.; Granzow, H.; Jöst, H.; Schmidt-Chanasit, J.; Groschup, M.H. Isolation of sindbis virus from a hooded crow in Germany. Vector-Borne Zoonotic Dis. 2014, 14, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Hesson, J.C.; Ostman, O.; Schäfer, M.; Lundström, J.O. Geographic distribution and relative abundance of the sibling vector species Culex torrentium and Culex pipiens in Sweden. Vector-Borne Zoonotic Dis. 2011, 11, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Málková, D.; Danielová, V.; Minár, B.; Rosický, B.; Casals, J. Isolation of Yaba 1 arbovirus in Czechoslovakia. Acta Virol. 1972, 16, 93. [Google Scholar] [PubMed]
- Danielová, V. To the problem of the vector of Lednice virus. Folia Parasitol. (Praha) 1984, 31, 379–382. [Google Scholar]
- Bakonyi, T.; Hubálek, Z.; Rudolf, I.; Nowotny, N. Novel flavivirus or new lineage of West Nile virus, Central Europe. Emerg. Infect. Dis. 2005, 11, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Jöst, H.; Bialonski, A.; Schmetz, C.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation and phylogenetic analysis of Batai virus, Germany. Am. J. Trop. Med. Hyg. 2011, 84, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Weitzel, T.; Collado, A.; Jöst, A.; Pietsch, K.; Storch, V.; Becker, N. Genetic differentiation of populations within the Culex pipiens complex and phylogeny of related species. J. Am. Mosq. Control Assoc. 2009, 25, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Jöst, A.; Weitzel, T. The Culex pipiens complex in Europe. J. Am. Mosq. Control Assoc. 2012, 28, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E. Culex pipiens: Species versus species complex—taxonomic history and perspective. J. Am. Mosq. Control Assoc. 2012, 28, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C. A review of the status and significance of the species within the Culex pipiens group in Australia. J. Am. Mosq. Control Assoc. 2012, 28, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, D.M.; Lapointe, D.A.; Fleischer, R.C. Bottlenecks and multiple introductions: Population genetics of the vector of avian malaria in Hawaii. Mol. Ecol. 2000, 9, 1803–1814. [Google Scholar] [CrossRef] [PubMed]
- Hesson, J.C.; Rettich, F.; Merdić, E.; Vignjević, G.; Ostman, O.; Schäfer, M.; Schaffner, F.; Foussadier, R.; Besnard, G.; Medlock, J.M.; et al. The arbovirus vector Culex torrentium is more prevalent than Culex pipiens in northern and central Europe. Med. Vet. Entomol. 2014, 28, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, D.M.; Keyghobadi, N.; Malcolm, C.A.; Mehmet, C.; Schaffner, F.; Mogi, M.; Fleischer, R.C.; Wilkerson, R.C. Emerging vectors in the Culex pipiens complex. Science 2004, 303, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Shaikevich, E.V. PCR-RFLP of the COI gene reliably differentiates Cx. pipiens, Cx. pipiens f. molestus and Cx. torrentium of the Pipiens Complex. Eur. Mosq. Bull. 2007, 23, 25–30. [Google Scholar]
- Byrne, K.; Nichols, R.A. Culex pipiens in London Underground tunnels: Differentiation between surface and subterranean populations. Heredity (Edinb.) 1999, 82, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kothera, L.; Godsey, M.; Mutebi, J.-P.; Savage, H.M. A comparison of aboveground and belowground populations of Culex pipiens (Diptera: Culicidae) mosquitoes in Chicago, Illinois, and New York City, New York, using microsatellites. J. Med. Entomol. 2010, 47, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Dobrotworsky, N.V. The problem of the Culex pipiens complex in the South Pacific (including Australia). Bull. World Health Organ. 1967, 37, 251–255. [Google Scholar] [PubMed]
- Kent, R.J.; Harrington, L.C.; Norris, D.E. Genetic differences between Culex pipiens f. molestus and Culex pipiens pipiens (Diptera: Culicidae) in New York. J. Med. Entomol. 2007, 44, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Keyghobadi, N.; Matrone, M.A.; Ebel, G.D.; Kramer, L.D.; Fonseca, D.M. Microsatellite loci from the northern house mosquito (Culex pipiens), a principal vector of West Nile virus in North America. Mol. Ecol. Notes 2004, 4, 20–22. [Google Scholar] [CrossRef]
- Bahnck, C.M.; Fonseca, D.M. Rapid assay to identify the two genetic forms of Culex (Culex) pipiens L. (Diptera: Culicidae) and hybrid populations. Am. J. Trop. Med. Hyg. 2006, 75, 251–255. [Google Scholar] [PubMed]
- Smith, J.L.; Fonseca, D.M. Rapid assays for identification of members of the Culex (Culex) pipiens complex, their hybrids, and other sibling species (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 2004, 70, 339–345. [Google Scholar] [PubMed]
- Börstler, J.; Lühken, R.; Rudolf, M.; Steinke, S.; Melaun, C.; Becker, S.; Garms, R.; Krüger, A. The use of morphometric wing characters to discriminate female Culex pipiens and Culex torrentium. J. Vector Ecol. 2014, 39, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; van de Peppel, L.J.J.; van Vliet, A.J.H.; Westenberg, M.; Ibañez-Justicia, A.; Stroo, A.; Buijs, J.A.; Visser, T.M.; Koenraadt, C.J.M. Winter activity and aboveground hybridization between the two biotypes of the West Nile virus vector Culex pipiens. Vector-Borne Zoonotic Dis. 2015, 15, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Danabalan, R.; Ponsonby, D.J.; Linton, Y.-M. A critical assessment of available molecular identification tools for determining the status of Culex pipiens s.L. in the United Kingdom. J. Am. Mosq. Control Assoc. 2012, 28, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Shaikevich, E.V.; Vinogradova, E.B.; Bouattour, A.; Gouveia de Almeida, A.P. Genetic diversity of Culex pipiens mosquitoes in distinct populations from Europe: Contribution of Cx. quinquefasciatus in Mediterranean populations. Parasit. Vectors 2016, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Manley, R.; Harrup, L.E.; Veronesi, E.; Stubbins, F.; Stoner, J.; Gubbins, S.; Wilson, A.; Batten, C.; Koenraadt, C.J.M.; Henstock, M.; et al. Testing of UK populations of Culex pipiens L. for Schmallenberg virus vector competence and their colonization. PLoS ONE 2015, 10, e0134453. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Lunt, D.H.; Zhang, D.-X.; Szymura, J.M.; Hewitt, O.M. The insect cytochrome oxidase I gene: Evolutionary patterns and conserved primers for phylogenetic studies. Insect Mol. Biol. 1996, 5, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, M.V.; Valsangiacomo, C.; Piffaretti, J.C.; Ward, P.I. Phylogenetic relationships among Muscoidea (Diptera: Calyptratae) based on mitochondrial DNA sequences. Insect Mol. Biol. 2000, 9, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Shaikevich, E.V.; Zakharov, I.A. Polymorphism of mitochondrial COI and nuclear ribosomal ITS2 in the Culex pipiens complex and in Culex torrentium (Diptera: Culicidae). Comp. Cytogenet. 2010, 4, 161–174. [Google Scholar] [CrossRef]
- Bourguet, D.; Fonseca, D.; Vourch, G.; Dubois, M.P.; Chandre, F.; Severini, C.; Raymond, M. The acetylcholinesterase gene Ace: A diagnostic marker for the Pipiens and Quinquefasciatus forms of the Culex pipiens complex. J. Am. Mosq. Control Assoc. 1998, 14, 390–396. [Google Scholar] [PubMed]
- Atyame, C.M.; Delsuc, F.; Pasteur, N.; Weill, M.; Duron, O. Diversification of Wolbachia endosymbiont in the Culex pipiens mosquito. Mol. Biol. Evol. 2011, 28, 2761–2772. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, M.; Czajka, C.; Börstler, J.; Melaun, C.; Jöst, H.; von Thien, H.; Badusche, M.; Becker, N.; Schmidt-Chanasit, J.; Krüger, A.; et al. First nationwide surveillance of Culex pipiens complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS ONE 2013, 8, e71832. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Parola, P.; Lindström, A.; Lilja, T.; L’Ambert, G.; Bondesson, U.; Berenger, J.M.; Raoult, D.; Almeras, L. Identification of European mosquito species by MALDI-TOF MS. Parasitol. Res. 2014, 113, 2375–2378. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, C.; Schaffner, F.; Ziegler, D.; Pflüger, V.; Mathis, A. Identification of field-caught Culicoides biting midges using matrix-assisted laser desorption/ionization time of flight mass spectrometry. Parasitology 2012, 139, 248–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yssouf, A.; Flaudrops, C.; Drali, R.; Kernif, T.; Socolovschi, C.; Berenger, J.M.; Raoult, D.; Parola, P. Matrix-assisted laser desorption ionization-time of flight mass spectrometry for rapid identification of tick vectors. J. Clin. Microbiol. 2013, 51, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Zittra, C.; Flechl, E.; Kothmayer, M.; Vitecek, S.; Rossiter, H.; Zechmeister, T.; Fuehrer, H.-P. Ecological characterization and molecular differentiation of Culex pipiens complex taxa and Culex torrentium in eastern Austria. Parasit. Vectors 2016, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.B.E.M.; De Vries, A.; Buijs, J.; Braks, M.A.H.; Den Hartog, W.; Scholte, E.J. First evidence for presence of Culex pipiens biotype molestus in the Netherlands, and of hybrid biotype pipiens and molestus in Northern Europe. J. Vector Ecol. 2010, 35, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Barriga, D.; Gomes, B.; Almeida, A.P.G.; Serrano-Aguilera, F.J.; Pérez-Martín, J.E.; Calero-Bernal, R.; Reina, D.; Frontera, E.; Pinto, J. The mosquito fauna of the western region of Spain with emphasis on ecological factors and the characterization of Culex pipiens forms. J. Vector Ecol. 2017, 42, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Osório, H.C.; Zé-Zé, L.; Amaro, F.; Nunes, A.; Alves, M.J. Sympatric occurrence of Culex pipiens (Diptera, Culicidae) biotypes pipiens, molestus and their hybrids in Portugal, Western Europe: Feeding patterns and habitat determinants. Med. Vet. Entomol. 2014, 28, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Di Luca, M.; Toma, L.; Boccolini, D.; Severini, F.; La Rosa, G.; Minelli, G.; Bongiorno, G.; Montarsi, F.; Arnoldi, D.; Capelli, G.; et al. Ecological distribution and CQ11 genetic structure of Culex pipiens complex (Diptera: Culicidae) in Italy. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.; Sousa, C.A.; Novo, M.T.; Freitas, F.B.; Alves, R.; Côrte-Real, A.R.; Salgueiro, P.; Donnelly, M.J.; Almeida, A.P.G.; Pinto, J. Asymmetric introgression between sympatric molestus and pipiens forms of Culex pipiens (Diptera: Culicidae) in the Comporta region, Portugal. BMC Evol. Biol. 2009, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; Möhlmann, T.W.R.; Melsen, D.; Favia, G.; Wennergren, U.; Koenraadt, C.J.M. Latitudinal diversity of Culex pipiens biotypes and hybrids in farm, peri-urban, and wetland habitats in Europe. PLoS ONE 2016, 11, e0166959. [Google Scholar] [CrossRef] [PubMed]
- Shaikevich, E.V.; Vinogradova, E.B. The discovery of a hybrid population of mosquitoes of the Culex pipiens L. complex (Diptera, Culicidae) on the Kos Island (Greece) by means of molecular markers. Entomol. Rev. 2014, 94, 35–39. [Google Scholar] [CrossRef]
- Amraoui, F.; Tijane, M.; Sarih, M.; Failloux, A.-B. Molecular evidence of Culex pipiens form molestus and hybrids pipiens/molestus in Morocco, North Africa. Parasit. Vectors 2012, 5, 83. [Google Scholar] [CrossRef] [PubMed]
- Cornel, A.J.; McAbee, R.D.; Rasgon, J.; Stanich, M.A.; Scott, T.W.; Coetzee, M. Differences in extent of genetic introgression between sympatric Culex pipiens and Culex quinquefasciatus (Diptera: Culicidae) in California and South Africa. J. Med. Entomol. 2003, 40, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; Hartemink, N.; Koenraadt, C.J.M. Modelling West Nile virus transmission risk in Europe: Effect of temperature and mosquito biotypes on the basic reproduction number. Sci. Rep. 2017, 7, 5022. [Google Scholar] [CrossRef] [PubMed]
- Han, L.L.; Popovici, F.; Alexander, J.P., Jr.; Laurentia, V.; Tengelsen, L.A.; Cernescu, C.; Gary, H.E., Jr.; Ion-Nedelcu, N.; Campbell, G.L.; Tsai, T.F.; et al. Risk factors for West Nile virus infection and meningoencephalitis, Romania, 1996. J. Infect. Dis. 1999, 179, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.L.; Walker, E.D.; Miller, J.R.; Severson, D.W.; Dworkin, I. Divergent host preferences of above- and below-ground Culex pipiens mosquitoes and their hybrid offspring. Med. Vet. Entomol. 2015, 29, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; Fros, J.J.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J.M. Vector competence of northern European Culex pipiens biotypes and hybrids for West Nile virus is differentially affected by temperature. Parasit. Vectors 2016, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Weitzel, T.; Jawień, P.; Rydzanicz, K.; Lonc, E.; Becker, N. Culex pipiens s.l. and Culex torrentium (Culicidae) in Wrocław area (Poland): Occurrence and breeding site preferences of mosquito vectors. Parasitol. Res. 2015, 114, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Lühken, R.; Steinke, S.; Leggewie, M.; Tannich, E.; Krüger, A.; Becker, S.; Kiel, E. Physico-chemical characteristics of Culex pipiens sensu lato and Culex torrentium (Diptera: Culicidae) breeding sites in Germany. J. Med. Entomol. 2015, 52, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Service, M.W. The taxonomy and biology of two sympatric sibling species of Culex, C. pipiens and C. torrentium (Diptera, Culicidae). J. Zool. 1968, 156, 313–323. [Google Scholar] [CrossRef]
- Simpson, J.E.; Hurtado, P.J.; Medlock, J.; Molaei, G.; Andreadis, T.G.; Galvani, A.P.; Diuk-Wasser, M.A. Vector host-feeding preferences drive transmission of multi-host pathogens: West Nile virus as a model system. Proc. R. Soc. B Biol. Sci. 2012, 279, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Takken, W.; Verhulst, N.O. Host preferences of blood-feeding mosquitoes. Annu. Rev. Entomol. 2013, 58, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P. West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLoS Biol. 2006, 4, e82. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.D.; Scott, T.W. Host defensive behaviour and the feeding success of mosquitoes. Int. J. Trop. Insect Sci. 1987, 8, 617–622. [Google Scholar] [CrossRef]
- De Boer, J.G.; Robinson, A.; Powers, S.J.; Burgers, S.L.G.E.; Caulfield, J.C.; Birkett, M.A.; Smallegange, R.C.; van Genderen, P.J.J.; Bousema, T.; Sauerwein, R.W.; et al. Odours of Plasmodium falciparum-infected participants influence mosquito-host interactions. Sci. Rep. 2017, 7, 9283. [Google Scholar] [CrossRef] [PubMed]
- Cornet, S.; Nicot, A.; Rivero, A.; Gandon, S. Both infected and uninfected mosquitoes are attracted toward malaria infected birds. Malar. J. 2013, 12, 179. [Google Scholar] [CrossRef] [PubMed]
- Cornet, S.; Nicot, A.; Rivero, A.; Gandon, S. Malaria infection increases bird attractiveness to uninfected mosquitoes. Ecol. Lett. 2013, 16, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; Fros, J.J.; Pijlman, G.P.; van Loon, J.J.A.; Gort, G.; Koenraadt, C.J.M. Virus interferes with host-seeking behaviour of mosquito. J. Exp. Biol. 2017, 220, 3598–3603. [Google Scholar] [CrossRef] [PubMed]
- Busula, A.O.; Bousema, T.; Mweresa, C.K.; Masiga, D.; Logan, J.G.; Sauerwein, R.W.; Verhulst, N.O.; Takken, W.; de Boer, J.G. Gametocytemia and attractiveness of Plasmodium falciparum-infected Kenyan children to Anopheles gambiae mosquitoes. J. Infect. Dis. 2017, 216, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E.; Harrison, B.A.; Gad, A.M. Culex (Culex) molestus Forskal (Diptera: Culicidae): Neotype designation, description, variation and taxonomic status. Proc. Entomol. Soc. Washingt. 1984, 86, 521–542. [Google Scholar]
- Harbach, R.E.; Dahl, C.; White, G.B. Culex (Culex) pipiens Linnacus (Diptera: Culicidae): Concepts, type designations, and description. Proc. Entomol. Soc. Washingt. 1985, 87, 1–24. [Google Scholar]
- Platonov, A.E.; Fedorova, M.V.; Karan, L.S.; Shopenskaya, T.A.; Platonova, O.V.; Zhuravlev, V.I. Epidemiology of West Nile infection in Volgograd, Russia, in relation to climate change and mosquito (Diptera: Culicidae) bionomics. Parasitol. Res. 2008, 103. [Google Scholar] [CrossRef] [PubMed]
- Martínez-de la Puente, J.; Moreno-Indias, I.; Hernández-Castellano, L.E.; Argüello, A.; Ruiz, S.; Soriguer, R.; Figuerola, J. Host-feeding pattern of Culex theileri (Diptera: Culicidae), potential vector of Dirofilaria immitis in the Canary Islands, Spain. J. Med. Entomol. 2012, 49, 1419–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osório, H.C.; Zé-Zé, L.; Alves, M.J. Host-feeding patterns of Culex pipiens and other potential mosquito vectors (Diptera: Culicidae) of West Nile virus (Flaviviridae) collected in Portugal. J. Med. Entomol. 2012, 49, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, S.; Yildirim, A.; Duzlu, O.; Ciloglu, A.; Onder, Z.; Inci, A. Blood meal identification of the mosquito (Diptera: Culicidae) specimens belong to Culex pipiens complex that were collected from Kayseri Province. Turkish J. Parasitol. 2017, 40, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Martínez-de la Puente, J.; Muñoz, J.; Capelli, G.; Montarsi, F.; Soriguer, R.; Arnoldi, D.; Rizzoli, A.; Figuerola, J. Avian malaria parasites in the last supper: identifying encounters between parasites and the invasive Asian mosquito tiger and native mosquito species in Italy. Malar. J. 2015, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Martínez-de la Puente, J.; Ferraguti, M.; Ruiz, S.; Roiz, D.; Soriguer, R.C.; Figuerola, J. Culex pipiens forms and urbanization: Effects on blood feeding sources and transmission of avian Plasmodium. Malar. J. 2016, 15, 589. [Google Scholar] [CrossRef] [PubMed]
- Börstler, J.; Jöst, H.; Garms, R.; Krüger, A.; Tannich, E.; Becker, N.; Schmidt-Chanasit, J.; Lühken, R. Host-feeding patterns of mosquito species in Germany. Parasit. Vectors 2016, 9, 318. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.; Sousa, C.A.; Vicente, J.L.; Pinho, L.; Calderón, I.; Arez, E.; Almeida, A.P.G.; Donnelly, M.J.; Pinto, J. Feeding patterns of molestus and pipiens forms of Culex pipiens (Diptera: Culicidae) in a region of high hybridization. Parasit. Vectors 2013, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Alcaide, M.; Rico, C.; Ruiz, S.; Soriguer, R.; Muñoz, J.; Figuerola, J. Disentangling vector-borne transmission networks: A universal DNA barcoding method to identify vertebrate hosts from arthropod bloodmeals. PLoS ONE 2009, 4, e7092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-de la Puente, J.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effect of blood meal digestion and DNA extraction protocol on the success of blood meal source determination in the malaria vector Anopheles atroparvus. Malar. J. 2013, 12, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, J.; Eritja, R.; Alcaide, M.; Montalvo, T.; Soriguer, R.C.; Figuerola, J. Host-feeding patterns of native Culex pipiens and invasive Aedes albopictus mosquitoes (Diptera: Culicidae) in urban zones from Barcelona, Spain. J. Med. Entomol. 2011, 48, 956–960. [Google Scholar] [CrossRef] [Green Version]
- Cancrini, G.; Magi, M.; Gabrielli, S.; Arispici, M.; Tolari, F.; Dell’Omodarme, M.; Prati, M.C. Natural vectors of Dirofilariasis in rural and urban areas of the Tuscan region, central Italy. J. Med. Entomol. 2006, 43, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Curtotti, A. Characterization of East London Culex pipiens s.l. in Relation to the Risk of Transmission to Humans of the West Nile Virus in Great Britain. PhD Thesis, Queen Mary University of London, London, UK, 2009. [Google Scholar]
- Radrova, J.; Seblova, V.; Votypka, J. Feeding behavior and spatial distribution of Culex mosquitoes (Diptera: Culicidae) in wetland areas of the Czech Republic. J. Med. Entomol. 2013, 50, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Schönenberger, A.C.; Wagner, S.; Tuten, H.C.; Schaffner, F.; Torgerson, P.; Furrer, S.; Mathis, A.; Silaghi, C. Host preferences in host-seeking and blood-fed mosquitoes in Switzerland. Med. Vet. Entomol. 2016, 30, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Service, M.W. Feeding behaviour and host preferences of British mosquitoes. Bull. Entomol. Res. 1971, 60, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Service, M.W. A reappraisal of the role of mosquitoes in the transmission of myxomatosis in Britain. J. Hyg. (Lond). 1971, 69, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Balenghien, T.; Fouque, F.; Sabatier, P.; Bicout, D.J. Horse-, bird-, and human-seeking behavior and seasonal abundance of mosquitoes in a West Nile virus focus of southern France. J. Med. Entomol. 2006, 43, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Service, M.W. Observations on the ecology of some British mosquitoes. Bull. Entomol. Res. 1969, 59, 161–193. [Google Scholar] [CrossRef]
- Rizzoli, A.; Bolzoni, L.; Chadwick, E.A.; Capelli, G.; Montarsi, F.; Grisenti, M.; Martínez-de la Puente, J.; Muñoz, J.; Figuerola, J.; Soriguer, R.; et al. Understanding West Nile virus ecology in Europe: Culex pipiens host feeding preference in a hotspot of virus emergence. Parasit. Vectors 2015, 8, 213. [Google Scholar] [CrossRef] [PubMed]
- Fyodorova, M.V.; Savage, H.M.; Lopatina, J.V.; Bulgakova, T.A.; Ivanitsky, A.V.; Platonova, O.V.; Platonov, A.E. Evaluation of potential West Nile virus vectors in Volgograd Region, Russia, 2003 (Diptera: Culicidae): Species composition, bloodmeal host utilization, and virus infection rates of mosquitoes. J. Med. Entomol. 2006, 43, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G.T.; Niklasson, B. Feeding patterns of mosquitoes (Diptera: Culicidae) in relation to the transmission of Ockelbo disease in Sweden. Bull. Entomol. Res. 1986, 76, 375. [Google Scholar] [CrossRef]
- Muñoz, J.; Ruiz, S.; Soriguer, R.; Alcaide, M.; Viana, D.S.; Roiz, D.; Vázquez, A.; Figuerola, J. Feeding patterns of potential West Nile virus vectors in south-west Spain. PLoS ONE 2012, 7, e39549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyeka, J.O.A.; Boreham, P.F.L. Population studies, physiological state and mortality factors of overwintering adult populations of females of Culex pipiens L. (Diptera:Culicidae). Bull. Entomol. Res. 1987, 77, 99–112. [Google Scholar] [CrossRef]
- Ferraguti, M.; Martínez-de la Puente, J.; Muñoz, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Avian Plasmodium in Culex and Ochlerotatus mosquitoes from Southern Spain: Effects of season and host-feeding source on parasite dynamics. PLoS ONE 2013, 8, e66237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiz, D.; Vazquez, A.; Rosà, R.; Muñoz, J.; Arnoldi, D.; Rosso, F.; Figuerola, J.; Tenorio, A.; Rizzoli, A. Blood meal analysis, flavivirus screening, and influence of meteorological variables on the dynamics of potential mosquito vectors of West Nile virus in northern Italy. J. Vector Ecol. 2012, 37, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Brugman, V.A.; Hernández-Triana, L.M.; England, M.E.; Medlock, J.M.; Mertens, P.P.C.; Logan, J.G.; Wilson, A.J.; Fooks, A.R.; Johnson, N.; Carpenter, S. Blood-feeding patterns of native mosquitoes and insights into their potential role as pathogen vectors in the Thames estuary region of the United Kingdom. Parasites Vectors 2017, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Triana, L.M.; Brugman, V.A.; Prosser, S.W.J.; Weland, C.; Nikolova, N.; Thorne, L.; Fernández de Marco, M.; Fooks, A.R.; Johnson, N. Molecular approaches for blood meal analysis and species identification of mosquitoes (Insecta: Diptera: Culicidae) in rural locations in southern England, United Kingdom. Zootaxa 2017, 4250, 67. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P.; Fonseca, D.M. Genetic influences on mosquito feeding behavior and the emergence of zoonotic pathogens. Am. J. Trop. Med. Hyg. 2007, 77, 667–671. [Google Scholar] [PubMed]
- Huang, S.; Hamer, G.L.; Molaei, G.; Walker, E.D.; Goldberg, T.L.; Kitron, U.D.; Andreadis, T.G. Genetic variation associated with mammalian feeding in Culex pipiens from a West Nile virus epidemic region in Chicago, Illinois. Vector-Borne Zoonotic Dis. 2009, 9, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Silver, J.B. Mosquito Ecology: Field Sampling Methods, 3rd ed.; Springer Netherlands: Berlin, Germany, 2007; ISBN 140206666X. [Google Scholar]
- Brugman, V.A.; England, M.E.; Stoner, J.; Tugwell, L.; Harrup, L.E.; Wilson, A.J.; Medlock, J.M.; Logan, J.G.; Fooks, A.R.; Mertens, P.P.C.; et al. How often do mosquitoes bite humans in southern England? A standardised summer trial at four sites reveals spatial, temporal and site-related variation in biting rates. Parasit. Vectors 2017, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Sieg, M.; Schmidt, V.; Ziegler, U.; Keller, M.; Höper, D.; Heenemann, K.; Rückner, A.; Nieper, H.; Muluneh, A.; Groschup, M.H.; et al. Outbreak and Cocirculation of Three Different Usutu Virus Strains in Eastern Germany. Vector-Borne Zoonotic Dis. 2017, 17, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G.C. Impacts of the creation, expansion and management of English wetlands on mosquito presence and abundance—Developing strategies for future disease mitigation. Parasit. Vectors 2015, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Townroe, S.; Callaghan, A. British container breeding mosquitoes: The impact of urbanisation and climate change on community composition and phenology. PLoS ONE 2014, 9, e95325. [Google Scholar] [CrossRef] [PubMed]
- Dekoninck, W.; Hendrickx, F.; Vasn Bortel, W.; Versteirt, V.; Coosemans, M.; Damiens, D.; Hance, T.; De Clercq, E.M.; Hendrickx, G.; Schaffner, F.; et al. Human-induced expanded distribution of Anopheles plumbeus, experimental vector of West Nile virus and a potential vector of human malaria in Belgium. J. Med. Entomol. 2011, 48, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G.C. Colonization of a newly constructed urban wetland by mosquitoes in England: Implications for nuisance and vector species. J. Vector Ecol. 2014, 39, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Kassim, N.F.A.; Webb, C.E.; Russell, R.C. Is the expression of autogeny by Culex molestus Forskal (Diptera: Culicidae) influenced by larval nutrition or by adult mating, sugar feeding, or blood feeding? J. Vector Ecol. 2012, 37, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, I.; Betášová, L.; Blažejová, H.; Venclíková, K.; Straková, P.; Šebesta, O.; Mendel, J.; Bakonyi, T.; Schaffner, F.; Nowotny, N.; et al. West Nile virus in overwintering mosquitoes, central Europe. Parasit. Vectors 2017, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Drummond, C.L.; Drobnack, J.; Ruby, M.A.; Kramer, L.D.; Ebel, G.D. Emergence of Culex pipiens from overwintering hibernacula. J. Am. Mosq. Control Assoc. 2011, 27, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Nelms, B.M.; Macedo, P.A.; Kothera, L.; Savage, H.M.; Reisen, W.K. Overwintering biology of Culex (Diptera: Culicidae) mosquitoes in the Sacramento Valley of California. J. Med. Entomol. 2013, 50, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Climatic effects on mosquito abundance in Mediterranean wetlands. Parasit. Vectors 2014, 7, 333. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, M.; Martínez-de la Puente, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effects of landscape anthropization on mosquito community composition and abundance. Sci. Rep. 2016, 6, 29002. [Google Scholar] [CrossRef] [PubMed]
- LaDeau, S.L.; Allan, B.F.; Leisnham, P.T.; Levy, M.Z. The ecological foundations of transmission potential and vector-borne disease in urban landscapes. Funct. Ecol. 2015, 29, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Ibañez-Justicia, A.; Cianci, D. Modelling the spatial distribution of the nuisance mosquito species Anopheles plumbeus (Diptera: Culicidae) in the Netherlands. Parasit. Vectors 2015, 8, 258. [Google Scholar] [CrossRef] [PubMed]
- Heym, E.C.; Kampen, H.; Fahle, M.; Hohenbrink, T.L.; Schäfer, M.; Scheuch, D.E.; Walther, D. Anopheles plumbeus (Diptera: Culicidae) in Germany: updated geographic distribution and public health impact of a nuisance and vector mosquito. Trop. Med. Int. Heal. 2017, 22, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Misslin, R.; Telle, O.; Daudé, E.; Vaguet, A.; Paul, R.E. Urban climate versus global climate change-what makes the difference for dengue? Ann. N. Y. Acad. Sci. 2016, 1382, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Medlock, J.M.; Murray, V. Impact of drought on vector-borne diseases—how does one manage the risk? Public Health 2014, 128, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Beji, M.; Rhim, A.; Roiz, D.; Bouattour, A. Ecophysiological characterization and molecular differentiation of Culex pipiens forms (Diptera: Culicidae) in Tunisia. Parasit. Vectors 2017, 10, 327. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.; Russell, I. Flying in tune: Sexual recognition in mosquitoes. Curr. Biol. 2006, 16, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Pennetier, C.; Warren, B.; Dabiré, K.R.; Russell, I.J.; Gibson, G. “Singing on the Wing” as a mechanism for species recognition in the malarial mosquito Anopheles gambiae. Curr. Biol. 2010, 20, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Raymond, M.; Weill, M. Many compatible Wolbachia strains coexist within natural populations of Culex pipiens mosquito. Heredity (Edinb.) 2011, 106, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.J.; Harrup, L.E.; Tugwell, L.A.; Brugman, V.A.; England, M.; Carpenter, S. Quantification of within- and between-farm dispersal of Culicoides biting midges using an immunomarking technique. J. Appl. Ecol. 2017, 54, 1429–1439. [Google Scholar] [CrossRef] [PubMed]
- Verhulst, N.O.; Loonen, J.A.C.M.; Takken, W. Advances in methods for colour marking of mosquitoes. Parasit. Vectors 2013, 6, 200. [Google Scholar] [CrossRef] [PubMed]
- Syed, Z.; Leal, W.S. Acute olfactory response of Culex mosquitoes to a human- and bird-derived attractant. Proc. Natl. Acad. Sci. USA 2009, 106, 18803–18808. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.A.; Bernier, U.R.; Kline, D.L. Laboratory evaluation of avian odors for mosquito (Diptera: Culicidae) attraction. J. Med. Entomol. 2006, 43, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Mauer, D.J.; Rowley, W.A. Attraction of Culex pipiens pipiens (Diptera: Culicidae) to flower volatiles. J. Med. Entomol. 1999, 36, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Jhumur, U.S.; Dötterl, S.; Jürgens, A. Naïve and conditioned responses of Culex pipiens pipiens biotype molestus (Diptera: Culicidae) to flower odors. J. Med. Entomol. 2006, 43, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Hesson, J.C.; Ignell, R.; Hill, S.R.; Östman, Ö.; Lundström, J.O. Trapping biases of Culex torrentium and Culex pipiens revealed by comparison of captures in CDC traps, ovitraps, and gravid traps. J. Vector Ecol. 2015, 40, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Weitzel, T.; Braun, K.; Collado, A.; Jost, A.; Becker, N. Distribution and frequency of Culex pipiens and Culex torrentium (Culicidae) in Europe and diagnostic allozyme markers. Eur. Mosq. Bull. 2011, 29, 22–37. [Google Scholar]
- Calzolari, M.; Pautasso, A.; Montarsi, F.; Albieri, A.; Bellini, R.; Bonilauri, P.; Defilippo, F.; Lelli, D.; Moreno, A.; Chiari, M.; et al. West Nile virus surveillance in 2013 via mosquito screening in Northern Italy and the influence of weather on virus circulation. PLoS ONE 2015, 10, e014915. [Google Scholar] [CrossRef] [PubMed]
- Vaux, A.G.C.; Gibson, G.; Hernández-Triana, L.M.; Cheke, R.A.; McCracken, F.; Jeffries, C.L.; Horton, D.L.; Springate, S.; Johnson, N.; Fooks, A.R.; et al. Enhanced West Nile virus surveillance in the North Kent marshes, UK. Parasit. Vectors 2015, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Boukraa, S.; de La Grandiere, M.A.; Bawin, T.; Raharimalala, F.N.; Zimmer, J.Y.; Haubruge, E.; Thiry, E.; Francis, F. Diversity and ecology survey of mosquitoes potential vectors in Belgian equestrian farms: A threat prevention of mosquito-borne equine arboviruses. Prev. Vet. Med. 2016, 124, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.; Kioulos, E.; Papa, A.; Almeida, A.P.G.; Vontas, J.; Pinto, J. Distribution and hybridization of Culex pipiens forms in Greece during the West Nile virus outbreak of 2010. Infect. Genet. Evol. 2013, 16, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Petrić, D.; Petrović, T.; Hrnjaković Cvjetković, I.; Zgomba, M.; Milošević, V.; Lazić, G.; Ignjatović Ćupina, A.; Lupulović, D.; Lazić, S.; Dondur, D.; et al. West Nile virus “circulation” in Vojvodina, Serbia: Mosquito, bird, horse and human surveillance. Mol. Cell. Probes 2017, 31, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Brustolin, M.; Talavera, S.; Santamaría, C.; Rivas, R.; Pujol, N.; Aranda, C.; Marquès, E.; Valle, M.; Verdún, M.; Pagès, N.; et al. Culex pipiens and Stegomyia albopicta (=Aedes albopictus) populations as vectors for lineage 1 and 2 West Nile virus in Europe. Med. Vet. Entomol. 2016, 30, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Rogozi, E.; Velo, E.; Bino, S.; Kadriaj, P.; Severini, F.; Schaffner, F. An update of the mosquito fauna of Albania, based on a country-wide field survey 2011–2012. In Proceedings of the E-Sove: From biology to integrated in a changing world, Montpellier, France, 8–11 October 2012. [Google Scholar]
- Snow, K.; Ramsdale, C. Distribution chart for European mosquitoes. Eur. Mosq. Bull. 1999, 3, 14–31. [Google Scholar]
- Boukraa, S.; Dekoninck, W.; Versteirt, V.; Schaffner, F.; Coosemans, M.; Haubruge, E.; Francis, F. Updated checklist of the mosquitoes (Diptera: Culicidae) of Belgium. J. Vector Ecol. 2015, 40, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Huldén, L.; Huldén, L. Checklist of the family Culicidae (Diptera) in Finland. Zookeys 2014. [Google Scholar] [CrossRef] [PubMed]
- Culverwell, C.L. A report on the mosquitoes of mainland Åland, southwestern Finland and revised list of Finnish mosquitoes. Med. Vet. Entomol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, F. A revised cheddist of the French Culicidae. Eur. Mosq. Bull. 1998, 2, 1–9. [Google Scholar]
- Tóth, S.; Kenyeres, Z. Revised checklist and distribution maps of mosquitoes (Diptera, Culicidae) of Hungary. Eur. Mosq. Bull. 2012, 30, 30–65. [Google Scholar]
- Pakalniškis, S.; Bernotienė, R.; Lutovinovas, E.; Petrašiūnas, A.; Podėnas, S.; Rimšaitė, J.; Saether, O.A.; Spungis, V. Checklist of Lithuanian Diptera. New Rare Lith. Insect Species 2006, 18, 16–154. [Google Scholar] [CrossRef]
- Beck, M.; Galm, M.; Weitzel, T.; Fohlmeister, V.; Kaiser, A.; Amold, A. Preliminary studies on the mosquito fauna of Luxembourg. Eur. Mosq. Bull. 2003, 14, 21–24. [Google Scholar]
- Sulesco, T.M.; Toderas, I.K.; Toderas, L.G. Annotated checklist of the mosquitoes of the Republic of Moldova. J. Am. Mosq. Control Assoc. 2013, 29, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, H.; Ramos, H.; Pires, C.; Capela, R. An annotated checklist of the mosquitoes of continental Portugal (Diptera, Culicidae). Acta III Congr. Iber. Entomol. Spec. 1988, 233–254. [Google Scholar]
- Nicolescu, G.; Vladimirescu, A.; Ciolpan, O. The distribution of mosquitoes in Romania (Diptera: Culicidae). Part I: Anopheles, Aedes and Culex. Eur. Mosq. Bull. 2002, 13, 17–26. [Google Scholar]
- Božičić-Lothrop, B.; Vujić, A. Fauna of mosquitoes (Diptera: Culicidae) of Stara Planina, Serbia. Acta Entomol. Serbica 1996, 1, 31–38. [Google Scholar]
- Jalili, N.; Országh, I.; Halgoš, J.; Labuda, M. Mosquitoes (Diptera, Culicidae) of Slovakia. Eur. Mosq. Bull. 2000, 6, 20–26. [Google Scholar]
- Aranda, C.; Eritja, R.; Schaffner, F.; Escosa, R. Culex (Culex) torrentium Martini (Diptera: Culicidae) a new species from Spain. Eur. Mosq. Bull. 2000, 8, 7–9. [Google Scholar]
- Eritja, R.; Aranda, C.; Padrós, J.; Goula, M.; Lucientes, J.; Escosa, R.; Marques, E.; Cáceres, F. An annotaded checklist and bibliography of the mosquitoes of Spain (Diptera: Culicidae). Eur. Mosq. Bull. 2000, 8, 10–18. [Google Scholar] [CrossRef]
- Gunay, F.; Alten, B.; Simsek, F.; Aldemir, A.; Linton, Y.M. Barcoding Turkish Culex mosquitoes to facilitate arbovirus vector incrimination studies reveals hidden diversity and new potential vectors. Acta Trop. 2015, 143, 112–120. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Primary Vertebrate Hosts | Principal Vectors | Medical/Veterinary Importance |
|---|---|---|---|
| Batai virus (Bunyaviridae) | Pigs, horses, ruminants, and isolations from wild birds. | Anopheles maculipennis s.l., Anopheles claviger (Meigen, 1804), Coquillettidia richiardii (Ficalbi, 1889) | Mild illness in sheep/goats. Influenza-like illness in humans. |
| Chikungunya virus (Togaviridae) | Humans as primary reservoirs during epidemics. Non-human reservoirs include monkeys, rodents and birds. | Aedes aegypti, Aedes albopictus | Fever, joint pain (also chronic), occasional neurological involvement with some deaths reported. |
| Dengue virus (Flaviviridae) | Humans. | Aedes aegypti, Aedes albopictus | Serotype 1 recorded from Europe. Cases range from asymptomatic to severe haemorrhagic fever. |
| Inkoo virus (Bunyaviridae) | Mountain hares. | Aedes communis (De Geer, 1776) | Influenza-like illness in humans. |
| Lednice virus (Bunyaviridae) | Birds, primarily of the order Anseriformes. | Culex modestus (Ficalbi, 1889) | Unknown, avian fatalities not recorded. |
| Sindbis/Sindbis-like viruses (Togaviridae) | Birds (Passeriformes), occasionally rodents and amphibians. | Culex pipiens, Culex torrentium (Martini, 1925), Culiseta morsitans (Theobald, 1901), Coquillettidia richiardii, Aedes communis, Aedes excrucians (Walker, 1856), Aedes cinereus (Meigen, 1818) and Anopheles hyrcanus s.l. | Sporadic illness in birds, including mortality in chickens. Fever, malaise and potentially chronic arthritis in humans, no mortality. |
| Snowshoe hare virus (Bunyaviridae) | Snowshoe hare, voles, lemmings. | Aedes cinereus, Aedes vexans (Meigen, 1830), Aedes communis, Aedes punctor (Kirby, 1837), Aedes cataphylla (Dyar, 1916), Culiseta inornata (Williston, 1893) and Culiseta impatiens (Walker, 1848) | Non-fatal encephalitis in horses. Fever and occasional CNS involvement in humans. |
| Tahyna virus (Bunyaviridae) | Brown hares, hedgehogs, rodents. | Aedes vexans | Influenza-like illness in humans with occasional CNS involvement. |
| Usutu virus (Flaviviridae) | Birds, particularly the Passeriformes. | Culex spp. including Culex pipiens | Avian mortality recorded in several species. Limited neuroinvasive cases reported from Italy. |
| West Nile virus (Flaviviridae) | Wild birds. Mammals including horses and humans incidental hosts. | Culex pipiens, Culex modestus, Coquillettidia richiardii | Limited avian mortality in Europe, equine febrile illness with ~25% mortality. Severe neurological disease in <1% human infections. |
| Ecoform | Trait | ||||
|---|---|---|---|---|---|
| Mating | Egg-Laying Requirements | Blood-Feeding Preference | Habitat Associations | Overwintering | |
| form pipiens | Eurogamous (mating requires open spaces) | Anautogenous (blood meal required for first egg batch) | Primarily birds | Rural and urban, aboveground | Heterodynamic (undergoes diapause) |
| form molestus | Stenogamous (can mate in confined spaces) | Autogenous (no blood meal required for first egg batch) | Birds and mammals | Principally urban, aboveground and underground | Homodynamic (active throughout the year) |
| Method | Target | Primer Sequences | Identification Output | References |
|---|---|---|---|---|
| Gel electrophoresis | Electrophoreticpolymorphisms in various genetic targets, often enzymes | n/a | Provides estimates of genetic differentiation between populations in target genes | [40,92] |
| Multiplex end-point PCR | ace-2 | FOR ACEtorr 5′-TGCCTGTGCTACCAGTGATGTT-3′ FOR ACEpip (5′-GGAAACAACGACGTATGTACT-3′) REV B1246s (5′-TGGAGCCTCCTCTTCACGG-3′) | Cx. pipiens complex: Cx. pipiens, Cx. quinquefasciatus, Cx. p. pallens, Cx. australicus, Cx. torrentium, Cx. pervigilans, Cx pipiens/Cx. quinquefasciatus hybrids | [98] |
| CQ11 | FOR CQ11F (5′-GATCCTAGCAAGCGAGAAC-3′) REV pipCQ11R (5′-CATGTTGAGCTTCGGTGAA-3′ REV molCQ11R (5′-CCCTCCAGTAAGGTATCAAC-3′ | Cx. pipiens form pipiens and form molestus | [97] | |
| PCR-DNA sequencing | COI | FOR LCO1490 (5′-GGTCAACAAATCATAAAGATATTGG-3′) REV HCO2198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′) | Enables universal identification to species level with comparison to sequence database | [104] |
| COI | FOR TY-J-1460 (5′-TACAATCTATCGCCTAAACTTCAGCC-3′) REV UEA10 (5′ -TCCAATGCACTAATCTGCCATATTA-3′) | [105,106,107] | ||
| PCR-RFLP | COI | FOR COIF (5′-TTGAGCTGGA-ATAGTTGGAACTT-3′) REV COIR (5′-CCTCCAATTGGATCAAAGAATGA-3′) | Cx. pipiens form pipiens and form molestus, Cx. torrentium | [91] |
| ace-2 | FOR F1457 (5′-GAGGAGATGTGGAATC CCAA-3′) REV B1246 (5′-TGGAGCCTCCTCTTCACGG C-3′) | Cx. pipiens, Cx. quinquefasciatus and their hybrids | [108] | |
| Wolbachia pipientis markers, ank2, pk1 | ank2 FOR (5′-CTTCTTCTGTGAGTGTACGT-3′) ank2 REV (5′-TCCATATCGATCTACTGCGT-3′) pk1 FOR (5′-CCACTACATTGCGCTATAGA-3′) pk1 REV (5′-ACAGTAGAACTACACTCCTCCA-3′) | Five groups of W. pipientis: wPip-I to wPip-V | [102,109] | |
| Real-time PCR | CQ11 | FOR Culex pipiens (5′-GCGGCCAAATATTGAGACTT-3′) REV Culex pipiens (5′-CGTCCTCAAACATCCA-GACA-3′) Probes Cx. pipiens all (59-Cy55-GGAACATGTTGAGCTTCGGK-BBQ-1-39 Cx. pipiens pipiens form pipiens (5′-JOE-GCTTCGGTGAAGGT TTGTGT-BHQ1-3′) Cx. pipiens pipiens form molestus (5′-Rox-TGAACCCTCC AGTAAGGTATCAACTAC-BHQ2-3′) | Collectively enables separation Cx. pipiens and its ecoforms and hybrids, plus Cx. torrentium | [110] |
| ace-2 | FOR Cx. torrentium (5′ -GACACAGGACGACAGAAA-3′) REV Cx. torrentium (5′-GCCTACGCAACTACTAAA-3′) Probe Cx. torrentium (5′-FAM-CGAT-GATGCCTGTGCTACCA-3BHQ1-3′) | |||
| CQ11 | FOR Cx_pip_F (5′-GCGGCCAAATATTGAGACTTTC-3′) REV Cx_pip_R (5′- ACTCGTCCTCAAACATCCAGACATA-3′) Probes Cpp_mol_P (5′-FAM-TGAACCCTCCAGTAAGGTA-MGB-3′) Cpp_pip_P1 (5′-VIC-CACA CAAAYCTTCACCGAA-MGB-3′) Cpp_pip_P2 (5′-VIC-ACACAAACCTTCATCGAA-MGB-3′) | Collectively enables separation Cx. pipiens and its ecoforms and hybrids, plus Cx. torrentium | [100] (modified from Rudolf et al. [110]) | |
| ace-2 | FOR Cx_tor_F (5′-CTTATTAGTATGACACAGGACGACAG AAA-3′) Cx_tor_R (5′-GCATAAACGCCTACGCAACTACTAA-3′) Probe Cx_tor_P (5′-FAM-ATGATGCCTGTG CTACCA-MGB-3′) |
| Order | Family | Genus Species | Common Name | Locations | References |
|---|---|---|---|---|---|
| Mammals | |||||
| Mammal, unidentified | - | - | - | Russia | [142] |
| Artiodactyla | Bovidae | Capra hircus | Goat | Spain (Canary Islands) | [143] |
| Ovis aries | Sheep | Portugal, Turkey | [117,144,145] | ||
| Bos taurus | Cow | Portugal, Turkey, Italy, Spain, Germany | [144,145,146,147,148,149] | ||
| Cervidae | Capreolus capreolus | Roe deer | Germany | [148] | |
| Suidae | Sus scrofa | Wild boar | Italy, Germany, Spain | [146,147,148] | |
| Carnivora | Canidae | Canis lupus familiaris | Dog | Spain, Turkey, Italy, Germany, UK | [145,146,147,148,150,151,152,153,154] |
| Felidae | Felis catus | Domestic cat | Spain, Czech Republic, Switzerland, Italy | [146,150,152,155,156] | |
| Felis silvestris | Wildcat | Spain | [147] | ||
| Herpestidae | Herpestes ichneumon | Egyptian mongoose | Spain | [150] | |
| Chiroptera | Vespertilionidae | Nyctalus noctula | Common Noctule | Czech Republic | [155] |
| Eulipotyphla | Erinaceidae | Erinaceus europaeus | European hedgehog | Italy | [146] |
| Lagomorpha | Leporidae | Oryctolagus cuniculus | Rabbit | UK, Germany, Spain | [147,148,157,158] |
| Lepus granatensis | Granada hare | Spain | [147] | ||
| Perissodactyla | Equidae | Equus caballus | Horse | France, Italy, Spain | [146,147,159] |
| Primates | Hominidae | Homo sapiens | Human | UK, Spain, Portugal, Czech Republic, Switzerland, Turkey, Italy, Russia, Germany | [142,144,145,146,147,148,149,150,152,154,155,156,160,161,162] |
| Rodentia | Caviidae | Cavia porcellus | Guinea pig | Sweden | [163] |
| Muridae | Rattus rattus | Rat | Spain | [147] | |
| Reptiles | |||||
| Reptile unidentified | - | - | - | Spain, Italy | [161,164] |
| Anura | Ranidae | Rana sp. | Frog | Czech Republic | [155] |
| Hylidae | Hyla arborea | European tree frog | Czech Republic | [155] | |
| Squamata | Lacertidae | Podarcis muralis | Common wall lizard | Italy | [146] |
| Lacerta sp. | Frog | Italy | [146] | ||
| Birds | |||||
| Bird, unidentified | - | - | - | UK, Spain, Switzerland, France, Russia, Portugal, Sweden | [117,142,151,156,159,160,162,163,164,165,166] |
| Accipitriformes | Accipitridae | Hieraaetus pennatus | Booted eagle | Turkey | [145] |
| Buteo buteo | Buzzard | Turkey | [145] | ||
| Neophron percnopterus | Egyptian vulture | Switzerland | [156] | ||
| Accipiter nisus | Eurasian sparrowhawk | Switzerland, Italy | [146,156] | ||
| Circus aeruginosus | Western marsh harrier | Czech Republic | [155] | ||
| Anseriformes | Anatidae | Cygnus atratus | Black swan | Spain | [151] |
| Anas sp. | Duck | Czech Republic | [155] | ||
| Anas crecca | Eurasian teal | Spain | [147] | ||
| Tachyeres pteneres | Flightless steamerduck | Switzerland | [156] | ||
| Anas strepera | Gadwall | Czech Republic | [155] | ||
| Anser sp. | Goose | Czech Republic | [155] | ||
| Anser albifrons | Greater white-fronted goose | Czech Republic | [155] | ||
| Anser anser | Greylag goose | Czech Republic | [155] | ||
| Anas platyrhynchos | Mallard | Portugal, Czech Republic, Switzerland, Italy, Germany | [144,146,148,155,156] | ||
| Cairina moschata | Muscovy duck | Spain, Portugal, Italy | [144,146,150] | ||
| Branta sandvicensis | Nene | Spain | [151] | ||
| Charadriiformes | Laridae | Larus ridibundus | Black-headed gull | Spain | [147] |
| Larus fuscus | Lesser black-backed gull | Portugal | [144] | ||
| Burhinidae | Burhinus oedicnemus | Eurasian stone-curlew | Spain | [147] | |
| Columbiformes | Columbidae | Streptopelia decaocto | Eurasian collared dove | Spain, Switzerland, Turkey, Italy | [145,146,147,150,152,156,161,164,167] |
| Columba livia | Rock dove | UK, Spain, Italy | [146,147,154,161] | ||
| Columba oenas | Stock dove | UK | [168] | ||
| Columba palumbus | Wood pigeon | Spain, Italy, UK | [146,152,168] | ||
| Falconiformes | Falconidae | Falco tinnunculus | Common kestrel | Portugal | [144] |
| Galliformes | Phasianidae | Gallus gallus | Chicken | Spain, Portugal, Switzerland, Italy, Russia, UK | [144,146,147,149,151,152,156,161,162,168] |
| Pavo cristatus | Common peacock | Switzerland | [156] | ||
| Phasianus colchicus | Common pheasant | Czech Republic, Italy | [146,155] | ||
| Coturnix coturnix | Common quail | Czech Republic | [155] | ||
| Alectoris rufa | Red-legged partridge | Spain | [150] | ||
| Alectoris rufa | Red-legged partridge | Spain | [147] | ||
| Meleagris gallopavo | Turkey | Portugal, Italy | [144,146] | ||
| Numididae | Numida meleagris | Helmeted guineafowl | Italy | [146] | |
| Gruiformes | Gruidae | Grus sp. | - | Spain | [151] |
| Grus grus | Common crane | Spain | [150] | ||
| Anthropoides virgo | Demoiselle crane | Switzerland | [156] | ||
| Rallidae | Rallus aquaticus | Water rail | Czech Republic | [155] | |
| Gallinula chloropus | Common moorhen | Italy | [146] | ||
| Passeriformes | Acrocephalidae | Acrocephalus scirpaceus | Eurasian reed warbler | Czech Republic | [155] |
| Hippolais polyglotta | Melodious warbler | Portugal, Spain | [144,147] | ||
| Alaudidae | Galerida cristata | Crested lark | Spain, Turkey, Portugal | [144,145,147,150] | |
| Alauda arvensis | Eurasian skylark | UK | [168] | ||
| Corvidae | Corvus corone | Carrion crow | Switzerland | [156] | |
| Garrulus glandarius | Eurasian jay | Turkey | [145] | ||
| Pica pica | Eurasian magpie | Czech Republic, Switzerland, Turkey, Italy | [145,146,155,156,161] | ||
| Cyanopica cooki | Iberian magpie | Portugal | [144] | ||
| Cyanocorax chrysops | Plush-crested jay | Switzerland | [156] | ||
| Emberizidae | Miliaria calandra | Corn bunting | Portugal | [144] | |
| Emberiza citrinella | Yellowhammer | Czech Republic, Germany | [148,155] | ||
| Fringillidae | Serinus canaria | Atlantic canary | Portugal | [144] | |
| Fringilla coelebs | Common chaffinch | Czech Republic | [155] | ||
| Carduelis chloris | European greenfinch | Spain, Italy | [146,151] | ||
| Serinus serinus | European serin | Italy | [167] | ||
| Carduelis chloris | Greenfinch | Spain | [147] | ||
| Hirundinidae | Hirundo rustica | Barn swallow | Czech Republic, UK | [155,168,169] | |
| Delichon urbica | House martin | Portugal, Czech Republic, Italy, Germany | [144,148,155,167] | ||
| Locustellidae | Bradypterus tacsanowskius | Chinese bush warbler | Portugal | [144] | |
| Motacillidae | Anthus pratensis | Meadow pipit | Spain, UK | [147,168] | |
| Motacilla alba | Pied wagtail | Czech Republic, Switzerland | [155,156] | ||
| Motacilla flava | Yellow wagtail | UK | [168] | ||
| Muscicapidae | Erithacus rubecula | European robin | Italy, Germany | [148,167] | |
| Oriolidae | Oriolus oriolus | Eurasian golden oriole | Italy | [146] | |
| Paridae | Cyanistes caeruleus | Blue tit | Portugal, Czech Republic, Switzerland, Germany | [144,148,149,155,156] | |
| Parus major | Great tit | Switzerland, Italy, UK | [146,156,169] | ||
| Passeridae | Passer montanus | Eurasian tree sparrow | Italy | [146] | |
| Passer domesticus | House sparrow | Spain, Portugal, Switzerland, Italy, UK, Germany | [144,146,147,148,149,150,151,152,156,161,164,167,168] | ||
| Sturnidae | Sturnus sp. | - | Spain | [147] | |
| Sturnus vulgaris | European starling | Spain, Czech Republic, Italy, UK | [146,150,155,161,168] | ||
| Sylviidae | Sylvia sp. | - | Spain | [147] | |
| Sylvia communis | Common whitethroat | Portugal, Czech Republic, Germany | [144,148,155] | ||
| Sylvia atricapilla | Eurasian blackcap | Czech Republic, Italy | [146,155] | ||
| Sylvia borin | Garden warbler | Portugal | [144] | ||
| Sylvia melanocephala | Sardinian warbler | Portugal, Spain | [144,147,150] | ||
| Turdidae | Turdus merula | Blackbird | Spain, Portugal, Czech Republic, Switzerland, Italy, UK, Germany | [144,146,147,148,149,150,152,155,156,161,164,167,168,169] | |
| Turdus philomelos | Song thrush | Czech Republic, Germany | [148,155] | ||
| Pelecaniformes | Ardeidae | Nycticorax nycticorax | Black-crowned night heron | Portugal, Italy, Spain | [144,146,147] |
| Bubulcus ibis | Cattle egret | Spain | [147] | ||
| Ardea cinerea | Grey heron | Czech Republic, UK | [155,168] | ||
| Ixobrychus minutus | Little bittern | Spain | [147] | ||
| Ardeola ralloides | Squacco heron | Spain | [147] | ||
| Piciformes | Picidae | Jynx torquilla | Eurasian wryneck | Italy | [146] |
| Psittaciformes | Cacatuidae | Nymphicus hollandicus | Cockatiel | Portugal | [144] |
| Psittacidae | Myiopsitta monachus | Monk parakeet | Spain | [152] | |
| Cyanoliseus patagonus | Patagonian conure | Switzerland | [156] | ||
| Sphenisciformes | Spheniscidae | Spheniscus humboldti | Humboldt's penguin | Switzerland | [156] |
| Strigiformes | Tytonidae | Tyto alba | Barn owl | UK | [168] |
| Tyto alba guttata | Dark-breasted barn owl | UK | [168] | ||
| Strigidae | Athene noctua | Little owl | Turkey, Italy | [145,146] | |
| Asio otus | Long-eared owl | UK, Portugal, Spain | [147,149,168] | ||
| Suliformes | Sulidae | Morus bassanus | Northern gannet | Portugal | [144] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brugman, V.A.; Hernández-Triana, L.M.; Medlock, J.M.; Fooks, A.R.; Carpenter, S.; Johnson, N. The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe. Int. J. Environ. Res. Public Health 2018, 15, 389. https://doi.org/10.3390/ijerph15020389
Brugman VA, Hernández-Triana LM, Medlock JM, Fooks AR, Carpenter S, Johnson N. The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe. International Journal of Environmental Research and Public Health. 2018; 15(2):389. https://doi.org/10.3390/ijerph15020389
Chicago/Turabian StyleBrugman, Victor A., Luis M. Hernández-Triana, Jolyon M. Medlock, Anthony R. Fooks, Simon Carpenter, and Nicholas Johnson. 2018. "The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe" International Journal of Environmental Research and Public Health 15, no. 2: 389. https://doi.org/10.3390/ijerph15020389
APA StyleBrugman, V. A., Hernández-Triana, L. M., Medlock, J. M., Fooks, A. R., Carpenter, S., & Johnson, N. (2018). The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe. International Journal of Environmental Research and Public Health, 15(2), 389. https://doi.org/10.3390/ijerph15020389