Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization—Biotechnological Potential and Biosafety Considerations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Ochrobactrum sp. POC9, Culture Conditions and Screening of Enzymatic Activities
2.2. Amplification and Sequencing of the 16S rRNA Gene
2.3. Draft Genome Sequencing
2.4. Bioinformatic Analyses
2.5. Analytical Methods
2.6. Simulation of the Anaerobic Digestion Process
2.7. Antibiotic Susceptibility Testing
2.8. Heavy Metal Resistance Testing
2.9. Adherence of Bacteria to Artificial Surface (Biofilm Formation) Testing
2.10. Nucleotide Sequence Accession Number
3. Results and Discussion
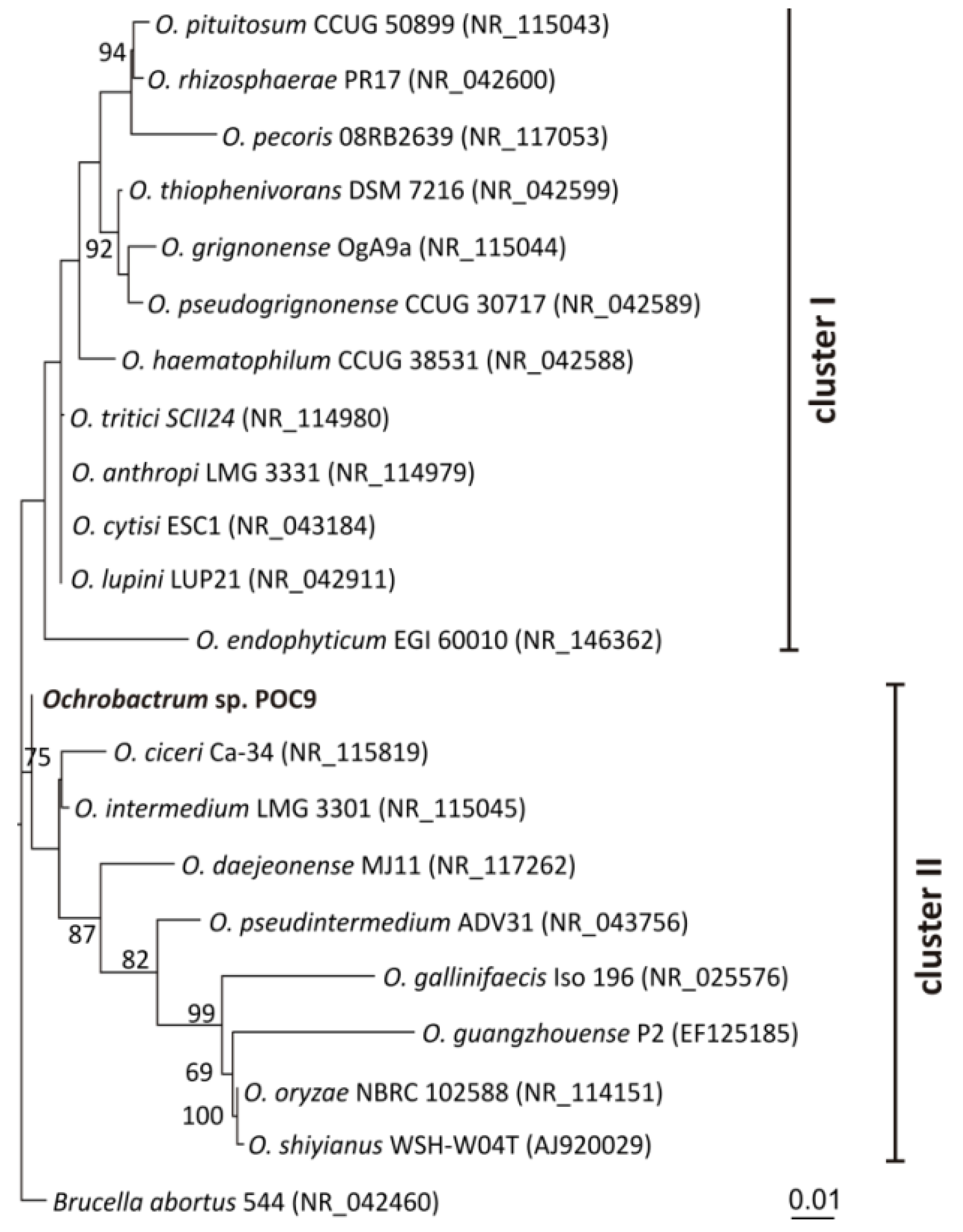
3.1. Isolation and Identification of Ochrobactrum sp. POC9
3.2. Bioaugmentation of Sewage Sludge Anaerobic Digestion
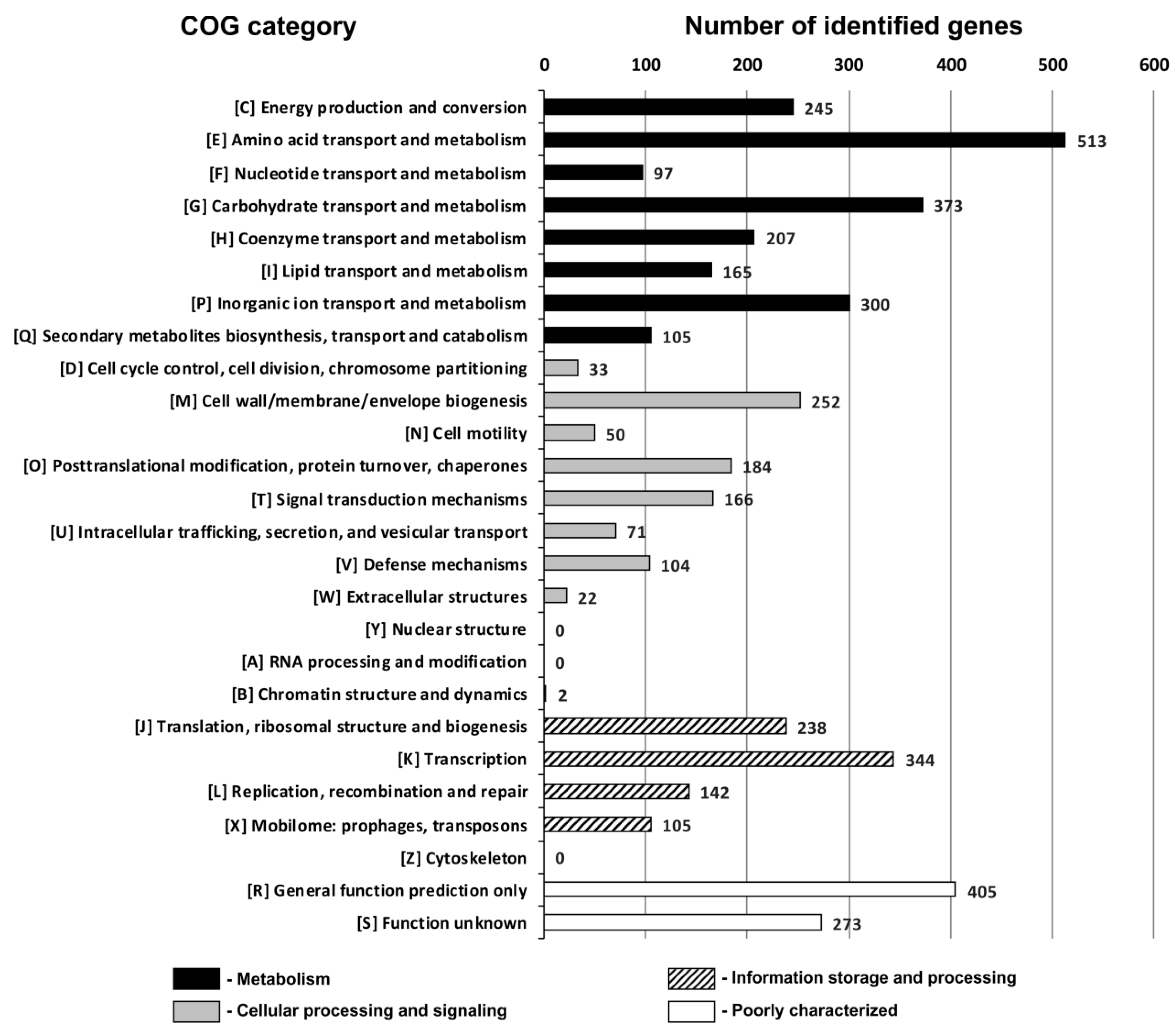
3.3. Genome-Based Insight into the Metabolic Potential of the POC9 Strain
3.4. Heavy Metal Metabolism
3.5. Antibiotic Resistance Genes and Virulence Factors
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malla, M.A.; Dubey, A.; Yadav, S.; Kumar, A.; Hashem, A.; Abd_Allah, E.F. Understanding and designing the strategies for the microbe-mediated remediation of environmental contaminants using omics approaches. Front. Microbiol. 2018, 9, 1132. [Google Scholar] [CrossRef] [PubMed]
- Danilovich, M.E.; Sanchez, L.A.; Acosta, F.; Delgado, O.D. Antarctic bioprospecting: In pursuit of microorganisms producing new antimicrobials and enzymes. Polar Biol. 2018, 1–17. [Google Scholar] [CrossRef]
- Head, I.M.; Jones, D.M.; Roling, W.F. Marine microorganisms make a meal of oil. Nat. Rev. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Harayama, S.; Kasai, Y.; Hara, A. Microbial communities in oil-contaminated seawater. Curr. Opin. Biotechnol. 2004, 15, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Kazy, S.K.; Gupta, A.; Dutta, A.; Mohapatra, B.; Roy, A.; Bera, P.; Mitra, A.; Sar, P. Biostimulation of indigenous microbial community for bioremediation of petroleum refinery sludge. Front. Microbiol. 2016, 7, 1407. [Google Scholar] [CrossRef] [PubMed]
- Aislabie, J.; Saul, D.J.; Foght, J.M. Bioremediation of hydrocarbon-contaminated polar soils. Extremophiles 2006, 10, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Vogt, C.; Richnow, H.H. Bioremediation via in situ microbial degradation of organic pollutants. Adv. Biochem. Eng. Biotechnol. 2014, 142, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Przygodzinska, D.; Matyjewicz, K.; Popowska, M. Occurrence and variety of β-lactamase genes among Aeromonas spp. isolated from urban wastewater treatment plant. Front. Microbiol. 2017, 8, 863. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, T. Pathogenic bacteria in sewage treatment plants as revealed by 454 pyrosequencing. Environ. Sci. Technol. 2011, 45, 7173–7179. [Google Scholar] [CrossRef] [PubMed]
- Lood, R.; Erturk, G.; Mattiasson, B. Revisiting antibiotic resistance spreading in wastewater treatment plants—Bacteriophages as a much neglected potential transmission vehicle. Front. Microbiol. 2017, 8, 2298. [Google Scholar] [CrossRef] [PubMed]
- Aujoulat, F.; Romano-Bertrand, S.; Masnou, A.; Marchandin, H.; Jumas-Bilak, E. Niches, population structure and genome reduction in Ochrobactrum intermedium: Clues to technology-driven emergence of pathogens. PLoS ONE 2014, 9, e83376. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, P.; Huber, B.; Busse, H.J.; Scholz, H.C.; Tomaso, H.; Hotzel, H.; Melzer, F. Ochrobactrum pecoris sp. nov. isolated from farm animals. Int. J. Syst. Evol. Microbiol. 2011, 61, 2278–2283. [Google Scholar] [CrossRef] [PubMed]
- Jackel, C.; Hertwig, S.; Scholz, H.C.; Nockler, K.; Reetz, J.; Hammerl, J.A. Prevalence, host range, and comparative genomic analysis of temperate Ochrobactrum phages. Front. Microbiol. 2017, 8, 1207. [Google Scholar] [CrossRef] [PubMed]
- Bathe, S.; Achouak, W.; Hartmann, A.; Heulin, T.; Schloter, M.; Lebuhn, M. Genetic and phenotypic microdiversity of Ochrobactrum spp. FEMS Microbiol. Ecol. 2006, 56, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Chain, P.S.; Lang, D.M.; Comerci, D.J.; Malfatti, S.A.; Vergez, L.M.; Shin, M.; Ugalde, R.A.; Garcia, E.; Tolmasky, M.E. Genome of Ochrobactrum anthropi ATCC 49188 T, a versatile opportunistic pathogen and symbiont of several eukaryotic hosts. J. Bacteriol. 2011, 193, 4274–4275. [Google Scholar] [CrossRef] [PubMed]
- Menezes, F.G.; Abreu, M.G.; Kawagoe, J.Y.; Warth, A.N.; Deutsch, A.D.; Dornaus, M.F.; Martino, M.D.; Correa, L. Ochrobactrum anthropi bacteremia in a preterm infant with cystic fibrosis. Braz. J. Microbiol. 2014, 45, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Moller, L.V.; Arends, J.P.; Harmsen, H.J.; Talens, A.; Terpstra, P.; Slooff, M.J. Ochrobactrum intermedium infection after liver transplantation. J. Clin. Microbiol. 1999, 37, 241–244. [Google Scholar] [PubMed]
- Galanakis, E.; Bitsori, M.; Samonis, G.; Christidou, A.; Georgiladakis, A.; Sbyrakis, S.; Tselentis, Y. Ochrobactrum anthropi bacteraemia in immunocompetent children. Scand. J. Infect. Dis. 2002, 34, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Aujoulat, F.; Roger, F.; Bourdier, A.; Lotthe, A.; Lamy, B.; Marchandin, H.; Jumas-Bilak, E. From environment to man: Genome evolution and adaptation of human opportunistic bacterial pathogens. Genes 2012, 3, 191–232. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.S.; Sarwari, A.R.; Khan, M.A.; Sophie, Z.; Khan, E.; Sami, S. Infective endocarditis and septic embolization with Ochrobactrum anthropi: Case report and review of literature. J. Infect. 2000, 40, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Zarinviarsagh, M.; Ebrahimipour, G.; Sadeghi, H. Lipase and biosurfactant from Ochrobactrum intermedium strain MZV101 isolated by washing powder for detergent application. Lipids Health Dis. 2017, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Zu, L.; Xiong, J.; Li, G.; Fang, Y.; An, T. Concurrent degradation of tetrabromobisphenol A by Ochrobactrum sp. T under aerobic condition and estrogenic transition during these processes. Ecotoxicol. Environ. Saf. 2014, 104, 220–225. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, W.S.; Ibrahim, M.K.; Abu-Shady, M.; El-Beih, F.; Ohmura, N.; Saiki, H.; Ando, A. Isolation and identification of a novel strain of the genus Ochrobactrum with phenol-degrading activity. J. Biosci. Bioeng. 2003, 96, 310–312. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Zhang, G.-S.; Zhang, Z.-H.; Xu, J.-H.; Li, S.-P. Isolation and characterization of a dichlorvos-degrading strain DDV-1 of Ochrobactrum sp. Pedosphere 2006, 16, 64–71. [Google Scholar] [CrossRef]
- Ermakova, I.T.; Shushkova, T.V.; Sviridov, A.V.; Zelenkova, N.F.; Vinokurova, N.G.; Baskunov, B.P.; Leontievsky, A.A. Organophosphonates utilization by soil strains of Ochrobactrum anthropi and Achromobacter sp. Arch. Microbiol. 2017, 199, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Arulazhagan, P.; Vasudevan, N. Biodegradation of polycyclic aromatic hydrocarbons by a halotolerant bacterial strain Ochrobactrum sp. VA1. Mar. Pollut. Bull. 2011, 62, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, B.; Karray, F.; Mhiri, N.; Aloui, F.; Sayadi, S. Application of a continuously stirred tank bioreactor (CSTR) for bioremediation of hydrocarbon-rich industrial wastewater effluents. J. Hazard. Mater. 2011, 189, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Katsivela, E.; Moore, E.R.; Maroukli, D.; Strompl, C.; Pieper, D.; Kalogerakis, N. Bacterial community dynamics during in-situ bioremediation of petroleum waste sludge in landfarming sites. Biodegradation 2005, 16, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Katsivela, E.; Moore, E.R.B.; Kalogerakis, N. Biodegradation of aliphatic and aromatic hydrocarbons: Specificity among bacteria isolated from refinery waste sludge. Water Air Soil Pollut. 2003, 3, 103–115. [Google Scholar] [CrossRef]
- Calvo, C.; Silva-Castro, G.A.; Uad, I.; Garcia Fandino, C.; Laguna, J.; Gonzalez-Lopez, J. Efficiency of the EPS emulsifier produced by Ochrobactrum anthropi in different hydrocarbon bioremediation assays. J. Ind. Microbiol. Biotechnol. 2008, 35, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Hendricks, C.W.; Doyle, J.D.; Hugley, B. A new solid medium for enumerating cellulose-utilizing bacteria in soil. Appl. Environ. Microbiol. 1995, 61, 2016–2019. [Google Scholar] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, E.P.; Burge, S.W.; Bateman, A.; Daub, J.; Eberhardt, R.Y.; Eddy, S.R.; Floden, E.W.; Gardner, P.P.; Jones, T.A.; Tate, J.; et al. Rfam 12.0: Updates to the RNA families database. Nucleic Acids Res. 2014, 43, D130–D137. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef] [PubMed]
- Eaton, A.D.; Clesceri, L.S.; Greenberg, A.E.; Franson, M.A.H. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA): Washington, DC, USA, 1998. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Available online: http://www.eucast.org (accessed on 13 May 2018).
- Dziewit, L.; Pyzik, A.; Matlakowska, R.; Baj, J.; Szuplewska, M.; Bartosik, D. Characterization of Halomonas sp. ZM3 isolated from the Zelazny Most post-flotation waste reservoir, with a special focus on its mobile DNA. BMC Microbiol. 2013, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Nieto, J.J.; Ventosa, A.; Ruiz-Berraquero, F. Susceptibility of halobacteria to heavy metals. Appl. Environ. Microbiol. 1987, 53, 1199–1202. [Google Scholar] [PubMed]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shanab, R.A.; van Berkum, P.; Angle, J.S. Heavy metal resistance and genotypic analysis of metal resistance genes in gram-positive and gram-negative bacteria present in Ni-rich serpentine soil and in the rhizosphere of Alyssum murale. Chemosphere 2007, 68, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Dib, J.; Motok, J.; Zenoff, V.F.; Ordonez, O.; Farias, M.E. Occurrence of resistance to antibiotics, UV-B, and arsenic in bacteria isolated from extreme environments in high-altitude (above 4400 m) Andean wetlands. Curr. Microbiol. 2008, 56, 510–517. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Kuttuva Rajarao, G.; Land, C.J.; Dalhammar, G. Biofilm formation and interactions of bacterial strains found in wastewater treatment systems. FEMS Microbiol. Lett. 2008, 283, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deflaun, M.F.; Steffan, R.J. Bioaugmentation. In Encyclopedia of Environmental Microbiology; Bitton, G., Ed.; Wiley-Interscience: New York, NY, USA, 2002; Volume 1, pp. 434–442. [Google Scholar]
- Bagi, Z.; Acs, N.; Balint, B.; Horvath, L.; Dobo, K.; Perei, K.R.; Rakhely, G.; Kovacs, K.L. Biotechnological intensification of biogas production. Appl. Microbiol. Biotechnol. 2007, 76, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.B.; Mladenovska, Z.; Ahring, B.K. Bioaugmentation of a two-stage thermophilic (68 °C/55 °C) anaerobic digestion concept for improvement of the methane yield from cattle manure. Biotechnol. Bioeng. 2007, 97, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Tauber, M.; Somitsch, W.; Meincke, R.; Muller, H.; Berg, G.; Guebitz, G.M. Enhancement of biogas production by addition of hemicellulolytic bacteria immobilised on activated zeolite. Water Res. 2010, 44, 1970–1980. [Google Scholar] [CrossRef] [PubMed]
- Tsapekos, P.; Kougias, P.G.; Vasileiou, S.A.; Treu, L.; Campanaro, S.; Lyberatos, G.; Angelidaki, I. Bioaugmentation with hydrolytic microbes to improve the anaerobic biodegradability of lignocellulosic agricultural residues. Bioresour. Technol. 2017, 234, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Cater, M.; Fanedl, L.; Malovrh, S.; Marinsek Logar, R. Biogas production from brewery spent grain enhanced by bioaugmentation with hydrolytic anaerobic bacteria. Bioresour. Technol. 2015, 186, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Fotidis, I.A.; Wang, H.; Fiedel, N.R.; Luo, G.; Karakashev, D.B.; Angelidaki, I. Bioaugmentation as a solution to increase methane production from an ammonia-rich substrate. Environ. Sci. Technol. 2014, 48, 7669–7676. [Google Scholar] [CrossRef] [PubMed]
- Poszytek, K.; Ciezkowska, M.; Sklodowska, A.; Drewniak, L. Microbial Consortium with High Cellulolytic Activity (MCHCA) for enhanced biogas production. Front. Microbiol. 2016, 7, 324. [Google Scholar] [CrossRef] [PubMed]
- Ariunbaatar, J.; Panico, A.; Esposito, G.; Pirozzi, F.; Lens, P.N.L. Pretreatment methods to enhance anaerobic digestion of organic solid waste. Appl. Energy 2014, 123, 143–156. [Google Scholar] [CrossRef]
- Parawira, W.; Tekere, M. Biotechnological strategies to overcome inhibitors in lignocellulose hydrolysates for ethanol production: Review. Crit. Rev. Biotechnol. 2011, 31, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Stuckey, D.C. Bioaugmentation and its application in wastewater treatment: A review. Chemosphere 2015, 140, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Cevallos, M.A.; Cervantes-Rivera, R.; Gutierrez-Rios, R.M. The repABC plasmid family. Plasmid 2008, 60, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Textor, S.; Wendisch, V.F.; De Graaf, A.A.; Muller, U.; Linder, M.I.; Linder, D.; Buckel, W. Propionate oxidation in Escherichia coli: Evidence for operation of a methylcitrate cycle in bacteria. Arch. Microbiol. 1997, 168, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Lv, P.; Luo, J.; Zhuang, X.; Zhang, D.; Huang, Z.; Bai, Z. Diversity of culturable aerobic denitrifying bacteria in the sediment, water and biofilms in Liangshui River of Beijing, China. Sci. Rep. 2017, 7, 10032. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.; Liu, C.; Liu, L.; Zhang, K.; Zou, S.; Hong, J.; Zhang, M. Hydrogen sulfide formation as well as ethanol production in different media by cysND- and/or cysIJ-inactivated mutant strains of Zymomonas mobilis ZM4. Bioprocess. Biosyst. Eng. 2013, 36, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Ciriminna, R.; Meneguzzo, F.; Delisi, R.; Pagliaro, M. Citric acid: Emerging applications of key biotechnology industrial product. Chem. Cent. J. 2017, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Jormakka, M.; Byrne, B.; Iwata, S. Formate dehydrogenase—A versatile enzyme in changing environments. Curr. Opin. Struct. Biol. 2003, 13, 418–423. [Google Scholar] [CrossRef]
- Oleszczuk, P. Phytotoxicity of municipal sewage sludge composts related to physico-chemical properties, PAHs and heavy metals. Ecotoxicol. Environ. Saf. 2008, 69, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Werle, S.; Wilk, R.K. A review of methods for the thermal utilization of sewage sludge: The Polish perspective. Renew. Energy 2010, 35, 1914–1919. [Google Scholar] [CrossRef]
- Cieslik, B.M.; Namiesnik, J.; Konieczka, P. Review of sewage sludge management: Standards, regulations and analytical methods. J. Clean. Prod. 2015, 90, 1–15. [Google Scholar] [CrossRef]
- Dixit, R.; Malaviya, D.; Pandiyan, K.; Singh, B.U.; Sahu, A.; Shukla, R.; Singh, P.B.; Rai, P.J.; Sharma, K.P.; Lade, H.; et al. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef]
- Binet, M.R.; Poole, R.K. Cd(II), Pb(II) and Zn(II) ions regulate expression of the metal-transporting P-type ATPase ZntA in Escherichia coli. FEBS Lett. 2000, 473, 67–70. [Google Scholar] [CrossRef]
- Hou, Z.; Mitra, B. The metal specificity and selectivity of ZntA from Escherichia coli using the acylphosphate intermediate. J. Biol. Chem. 2003, 278, 28455–28461. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.J.; Narindrasorasak, S.; Bhushan, B.; Sarkar, B.; Mitra, B. Functional analysis of chimeric proteins of the Wilson Cu(I)-ATPase (ATP7B) and ZntA, a Pb(II)/Zn(II)/Cd(II)-ATPase from Escherichia coli. J. Biol. Chem. 2001, 276, 40858–40863. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.H.; Moreno-Sanchez, R.; Cervantes, C. Chromate efflux by means of the ChrA chromate resistance protein from Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 7398–7400. [Google Scholar] [PubMed]
- Anton, A.; Grosse, C.; Reissmann, J.; Pribyl, T.; Nies, D.H. CzcD is a heavy metal ion transporter involved in regulation of heavy metal resistance in Ralstonia sp. strain CH34. J. Bacteriol. 1999, 181, 6876–6881. [Google Scholar] [PubMed]
- Munkelt, D.; Grass, G.; Nies, D.H. The chromosomally encoded cation diffusion facilitator proteins DmeF and FieF from Wautersia metallidurans CH34 are transporters of broad metal specificity. J. Bacteriol. 2004, 186, 8036–8043. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.; Colleran, E. Arsenical resistance in the IncHI2 plasmids. Plasmid 2002, 47, 234–240. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Nadjar, D.; Labia, R.; Cerceau, C.; Bizet, C.; Philippon, A.; Arlet, G. Molecular characterization of chromosomal class C beta-lactamase and its regulatory gene in Ochrobactrum anthropi. Antimicrob. Agents Chemother. 2001, 45, 2324–2330. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, K.; Kehrenberg, C.; Schwarz, S. Efflux-mediated resistance to florfenicol and/or chloramphenicol in Bordetella bronchiseptica: Identification of a novel chloramphenicol exporter. J. Antimicrob. Chemother. 2007, 59, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Pradel, E.; Pages, J.M. The AcrAB-TolC efflux pump contributes to multidrug resistance in the nosocomial pathogen Enterobacter aerogenes. Antimicrob. Agents Chemother. 2002, 46, 2640–2643. [Google Scholar] [CrossRef] [PubMed]
- Natale, P.; Bruser, T.; Driessen, A.J. Sec- and Tat-mediated protein secretion across the bacterial cytoplasmic membrane-distinct translocases and mechanisms. Biochim. Biophys. Acta 2008, 1778, 1735–1756. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, S.P.; Baumler, A.J. Evolution of the chaperone/usher assembly pathway: Fimbrial classification goes Greek. Microbiol. Mol. Biol. Rev. 2007, 71, 551–575. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Quinn, P.J.; Yan, A. Kdo2-lipid A: Structural diversity and impact on immunopharmacology. Biol. Rev. Camb. Philos. Soc. 2015, 90, 408–427. [Google Scholar] [CrossRef] [PubMed]
- Polissi, A.; Sperandeo, P. The lipopolysaccharide export pathway in Escherichia coli: Structure, organization and regulated assembly of the Lpt machinery. Mar. Drugs 2014, 12, 1023–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, P.A.; Stroeher, U.H.; Karageorgos, L.E.; Morona, R. Putative O-antigen transport genes within the rfb region of Vibrio cholerae O1 are homologous to those for capsule transport. Gene 1995, 158, 1–7. [Google Scholar] [CrossRef]
- Marczak, M.; Mazur, A.; Koper, P.; Zebracki, K.; Skorupska, A. Synthesis of rhizobial exopolysaccharides and their importance for symbiosis with legume plants. Genes 2017, 8, 360. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’Toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Loupasaki, E.; Diamadopoulos, E. Attached growth systems for wastewater treatment in small and rural communities: A review. J. Chem. Technol. Biotechnol. 2013, 88, 190–204. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain/ Microbial Consortium | Scale | Substrate for Anaerobic Digestion | Effect | Reference |
|---|---|---|---|---|
| Caldicellulosiruptor saccharolyticus, Enterobacter cloacae | Laboratory | Waste water sludge, pig manure slurry and dried plant biomass from Jerusalem artichoke | Increased biogas production of up to 160–170% | [57] |
| Caldicellulosiruptor lactoaceticus 6A, Dictyoglomus sp. B4a | Laboratory (batch experiments and CSTR bioreactor) | Cattle manure | Increased methane yield of up to 93% | [58] |
| Hemicellulolytic consortium immobilized on activated zeolite | Laboratory (batch experiments and CSTR bioreactor) | Xylan from birch wood | Increased methane yield of up to 5% | [59] |
| Clostridium thermocellum, Melioribacter roseus | Laboratory scale (batch experiments and CSTR bioreactor) | Wheat straw | Increased methane yield of up to 34% | [60] |
| Ruminococcus flavefaciens 007C, Pseudobutyrivibrio xylanivorans Mz5T, Fibrobacter succinogenes S85 and Clostridium cellulovorans | Laboratory | Brewery spent grain | Increased biogas production of up to 5–18% | [61] |
| Methanoculleus bourgensis MS2 | Laboratory | Ammonia-rich substrates (mixed pig and chicken manure, slaughterhouse residues, and food industry waste) | Increased methane yield of up to 34% | [62] |
| Microbial consortium with high cellulolytic activity (MCHCA) | Laboratory (two-stage anaerobic digestion) | Maize silage (lignocellulose biomass) | Increased biogas production of up to 38%, increased methane yield of up to 64% | [63] |
| Parameter (unit) | Control | Culture with the POC9 Strain | ||||
|---|---|---|---|---|---|---|
| 3 days | 7 days | 30 days | 3 days | 7 days | 30 days | |
| Cumulative biogas production (L/kgvs) | 229.58 ± 13.92 | 294.58 ± 44.98 | ||||
| CH4 content (%) | 43.41 | 61.34 | 49.18 | 46.00 | 66.48 | 58.87 |
| Genomic feature | Calculation |
|---|---|
| Number of contigs | 298 |
| Estimated genome size (bp) | 4,976,112 |
| GC content (%) | 55.68% |
| Coding density (%) | 89.07% |
| Number of genes | 5217 |
| Number of tRNA genes | 66 |
| Number of 16S-23S-5S rRNA clusters | 3 |
| Protein Name | Localization within the POC9 Draft Genome Sequence (GenBank acc. no.) | Predicted Protein Function | Homologous Protein Based on Best BLASTP Hit (GenBank acc. no.) |
|---|---|---|---|
| arsB | contig00008 (QGST01000008.1) coordinates: 148,637-147,576 | Export of As(III) ions | arsenic transporter of O. anthropi FRAF13 (KXO76567) |
| arsC | contig00008 (QGST01000008.1) coordinates: 147,579-147,169 | Reduction of As(V) to As(III) | arsenate reductase of O. intermedium LMG 3301 (EEQ95705) |
| arsH | contig00008 (QGST01000008.1) coordinates: 147,172-146,459 | Unknown function, probably regulatory protein | arsenical resistance protein ArsH of Ochrobactrum sp. 30A/1000/2015 (PJT26941) |
| chrA | contig00005 (QGST01000005.1) coordinates: 67,497-66,199 | Export of Cr(VI) | chromate transporter of Ochrobactrum sp. EGD-AQ16 (ERI13917) |
| czcD | contig00031 (QGST01000031.1) coordinates: 6786-7745 | Export of Cd(II), Co(II), and Zn(II) | cation transporter of Ochrobactrum sp. MYb71 (PQZ25943) |
| fieF | contig00005 (QGST01000005.1) coordinates: 103,790-102,813 | Export of Cd(II), Co(II), Fe(II), Ni(II), and Zn(II) | cadmium transporter of O. anthropi FRAF13 (KXO76051) |
| zntA | contig00025 (QGST01000025.1) coordinates: 28,104-26,260 | Export of Cd(II), Co(II), Cu(II), Ni(II), Pb(II), and Zn(II) | haloacid dehalogenase of O. anthropi FRAF13 (KXO73917) |
| zntA | contig00032 (QGST01000032.1) coordinates: 21,766-24,129 | Export of Cd(II), Co(II), Cu(II), Ni(II), Pb(II), and Zn(II) | lead, cadmium, zinc and mercury transporting ATPase; copper-translocating P-type ATPase of O. haematophilum FI11154 (SPL62610) |
| zntA | contig00034 (QGST01000034.1) coordinates: 5822-7666 | Export of Cd(II), Co(II), Cu(II), Ni(II), Pb(II), and Zn(II) | cadmium-translocating P-type ATPase of O. rhizosphaerae PR17 (OYR19288) |
| zntA | contig00065 (QGST01000065.1) coordinates: 984-2825 | Export of Cd(II), Co(II), Cu(II), Ni(II), Pb(II), and Zn(II) | heavy metal translocating P-type ATPase of O. anthropi ATCC 49188 (ABS17306) |
| zntA | contig00001 (QGST01000001.1) coordinates: 97,090-99,573 | Export of Cd(II), Co(II), Cu(II), Ni(II), Pb(II), and Zn(II) | ATPase of O. anthropi FRAF13 (KXO79927) |
| zntA | contig00004 (QGST01000004.1) coordinates: 22,193-19,692 | Export of Cd(II), Co(II), Cu(II), Ni(II), Pb(II), and Zn(II) | copper-translocating P-type ATPase of O. lupini LUP21 (OYR29555) |
| Gene/Gene Cluster Name | Localization within the POC9 Draft Genome Sequence (GenBank acc. no.) | Protein | Best BLAST Hits: [% Identity] Organism (GeneBank acc. no.) | Predicted Antimicrobial Resistance Profile | Tested Antibiotics | Profile |
|---|---|---|---|---|---|---|
| gcvA- blaOCH | contig00009 (QGST01000009.1) coordinates: 164,062–165,431 | class C beta-lactamase | [98%] Ochrobactrum tritici C8846-N36 (SME85995) | penams, penems, cephalosporins, cephamycins, monobactams | AMP CFM CTX CRO | R R R R |
| transcriptional regulator GcvA | [98%] Ochrobactrum anthropi (WP_061347328) | |||||
| acrAB- TolC | contig00001 (QGST01000001.1) coordinates: 191,438-196,605 | transcriptional regulator TetR | [93%] Ochrobactrum oryzae (WP_104756164) | tetracyclines, cephalosporins penams, phenicols, ryfamycins, fluoroquinolones | AMP CIP CFM CTX CRO TE MXF RIF | R S R R R S S R |
| efflux RND transporter periplasmic adaptor subunit | [98%] Ochrobactrum anthropi (WP_061344971) | |||||
| efflux RND transporter permease subunit | [99%] Ochrobactrum anthropi (WP_061344972) | |||||
| acrAB- TolC | contig00010 (QGST01000010.1) coordinates: 49,666–54,163 | efflux RND transporter periplasmic adaptor unit | [98%] Ochrobactrum sp. (WP_024900215) | |||
| efflux RND transporter permease subunit | [99%] Ochrobactrum oryzae (WP_104755654) | |||||
| qacH | contig00007 (QGST01000007.1) coordinates: 192,104–191,772 | efflux SMR transporter | [100%] Ochrobactrum sp. (WP_010661279) | fluoroquinolones | CIP MXF | S S |
| cmlA/floR | contig00016 (QGST01000016.1) coordinates: 56,907–55,714 | CmlA/floR chloramphenicol efflux MFS transporter | [94%] Ochrobactrum anthropi (WP_061345584) | chloramphenicol | C | R |
| acc (6’) | contig00016 (QGST01000016.1) coordinates: 92,931–92,485 | aminoglycoside 6’-acetyl-transferase | [93%] Ochrobactrum anthropi FRAF13(KXO77791) | aminoglycosides | CN | S |
| tetG- tetR | contig00014 (QGST01000014.1) coordinates: 7148–9063 | Tet(A/B/C) family MFS transporter | [87%] Ochrobactrum oryzae (WP_104755825) | tetracyclines | TE | S |
| transcriptional regulator tetR | [89%] Ochrobactrum oryzae (WP_104755986) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poszytek, K.; Karczewska-Golec, J.; Ciok, A.; Decewicz, P.; Dziurzynski, M.; Gorecki, A.; Jakusz, G.; Krucon, T.; Lomza, P.; Romaniuk, K.; et al. Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization—Biotechnological Potential and Biosafety Considerations. Int. J. Environ. Res. Public Health 2018, 15, 1501. https://doi.org/10.3390/ijerph15071501
Poszytek K, Karczewska-Golec J, Ciok A, Decewicz P, Dziurzynski M, Gorecki A, Jakusz G, Krucon T, Lomza P, Romaniuk K, et al. Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization—Biotechnological Potential and Biosafety Considerations. International Journal of Environmental Research and Public Health. 2018; 15(7):1501. https://doi.org/10.3390/ijerph15071501
Chicago/Turabian StylePoszytek, Krzysztof, Joanna Karczewska-Golec, Anna Ciok, Przemyslaw Decewicz, Mikolaj Dziurzynski, Adrian Gorecki, Grazyna Jakusz, Tomasz Krucon, Pola Lomza, Krzysztof Romaniuk, and et al. 2018. "Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization—Biotechnological Potential and Biosafety Considerations" International Journal of Environmental Research and Public Health 15, no. 7: 1501. https://doi.org/10.3390/ijerph15071501
APA StylePoszytek, K., Karczewska-Golec, J., Ciok, A., Decewicz, P., Dziurzynski, M., Gorecki, A., Jakusz, G., Krucon, T., Lomza, P., Romaniuk, K., Styczynski, M., Yang, Z., Drewniak, L., & Dziewit, L. (2018). Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization—Biotechnological Potential and Biosafety Considerations. International Journal of Environmental Research and Public Health, 15(7), 1501. https://doi.org/10.3390/ijerph15071501